Enzimas
Enzimas
Diferenças em relação aos catalisadores químicos:
1.Maiores velocidades de reacção
Com enzima
P
reacção
Sem enzima
tempo
A
P
2. Condições de reacção mais suaves
3. Maior especificidade
3. Capacidade de regulação
História
Anos:
1700 – estudos sobre
a digestão da carne
por secreções do
estômago.
1800- Conversão do amido em açucar pela saliva e
extractos de plantas (amilase)
1850 – Pasteur – a fermentação do açucar em álcool
pelas leveduras era catalisada por “fermentos”,
inseparáveis das células vivas – vitalismo.
1893 – Ostwald - enzima = catalisador
1894 - Fischer: -teoria chave-fechadura
1897 – Eduard Buchner – as
moléculas do extracto de levedura
responsáveis pela conversão de
açucar a etanol, continuavam a
funcionar quando removidas das
células.
É uma molécula !?
1877 Kühne: in zyme
enzima
1926 - Sumner: - urease cristal
É uma
proteína !
1905 Harden e Young: coenzima isolado(NAD+)
1909 Sorensen: escala de pH
1913 Michaelis e Menten: teoria cinética
1925 Briggs e Haldane: teoria cinética modificada
1931 Lineweaver e Burk determinação das
constantes cinéticas.
1960 Hirs, Moore e Stein ribonuclease (estrutura
primária)
1962 Phillips lisozima (estrutura terciária)
1970 Chang e Cohen clonagem do gene do
β-lactamase de Staphylococcus aureus em E. Coli
1982 produção de mutante de tirosil-RNA sintetase
por mutagénese dirigida
Mas todos os enzimas são realmente
proteínas?
Cech 1968 “ribozimas”- RNA com actividade
catalítica
Wade 1997 “abzimas”- anticorpo monoclonal
com actividade catalítica.
Aplicações industriais de enzimas
DISTRIBUIÇÃO DO MERCADO
14%
29%
32%
15%
p.lácteos
deterg.
texteis
amido
outros
10%
1983-1995 (2x)
-2005 (2x)
Godfrey,T., West, S.(eds)Industrial Enzimology, Macmillan, London(1990)
Indústria
Função dos enzimas
alimentos
quebra do xarope de amido em xarope de glucose (osidases);
isomerização de glucose em frutose, mais doce logo menor
quantidade (isomerase);
hidrólise da sacarose em glucose e frutose (hidrolases);
fabrico de adoçantes artificiais;
alimentos
bébé
início da digestão da comida (proteases e lipases)
sumos de
frutos
impedir turvação (pectinase)
detergentes
biológicos
anti-nódoas (proteases para as proteinas e lipases para as
gorduras);
amaciadores (celulases quebram os “borbotos” )
carne
tornar mais tenra
medicamentos
cancro, produção de drogas sintéticas (penicilina)
Enzimas em medicina
enfarte miocárdio
(diagnóstico)
terapêutica
Desenvolvimentos da última década
Mais estruturas
Mais sequências
•Sobre-expressão
•Mutagénese dirigida
+
Modificar e estudar propriedades
Novos métodos de cálculo automático
+
desenho molecular
Desenho de novos biocatalisadores
enzimas
artificiais
Enzimas artificiais
maior estabilidade térmica
Ex: subtilisina com 8 aa substituidos
vida média a 100o passa de 30s para 170min
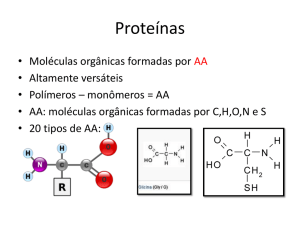
A maioria dos enzimas são proteínas!
MM 12 000 – 1 000 000
Classificação internacional de enzimas
Nº
Classe
Tipo de reacção
1
Oxidoreductases
Transferência de electrões
2
Transferases
Reacções de transferência de grupos
3
Hidrolases
Reacções de hidrólise (transferência
de grupos para a água)
4
Liases
Adição de grupos a ligações duplas ou
formação de ligações duplas por
remoção de grupos
5
Isomerases
Transferência de grupos dentro da
molécula para dar origem a formas
isoméricas
6
Ligases
Formação de ligações C-C, C-S, C-O e
C-N, por reacções de condensação
acopladas à hidrólise de ATP
Cada enzima tem um nome sistemático, um nº de
classificação e um nome trivial
Ex: O enzima que catalisa a reacção
ATP + D-glucose → ADP + D-glucose6-P
Nome sistemático : ATP glucose fosfotransferase
Nº de classificação: E.C. 2.7.1.1.
E.C. – Enzyme Commission
2- classe das transferases
7- subclasse fosfotransferase
1- fosfotransferase com um grupo hidroxilo como aceitador
1- D-glucose como aceitadora do grupo fosforilo
Nome trivial : hexoquinase
Há enzimas que precisam de outras moléculas
ou iões para a sua actividade
Cofactores – iões ou pequenas moléculas orgânicas
(coenzimas) necessários à actividade enzimática.
Os coenzimas podem estar transientemente ou
permanentemente – grupo prostético -ligados aos
enzimas .
Enzima + cofactor → holoenzima
Enzima sem cofactor → apoenzima
Cofactores - iões inorgânicos
Ião
Enzima
Cu 2+
Fe 2+ Fe 2+
K+
Mg 2+
Citocromo oxidase
Mn 2+
Mo
Ni 2+
Se
Zn 2+
Arginase,ribonucleotido redutase
Peroxidase, catalase, citocromo oxidase
Piruvato cinase
Hexocinase, 6-P-glucose fosfatase, piruvato
cinase
Dinitrogenase
Urease
Glutationo peroxidase
Anidrase carbónico, alcool desidrogenase,
carboxipeptidases A e B
Coenzimas: moléculas orgânicas complexas ou
organometálicas que transferem átomos específicos ou grupos
funcionais
coenzima
Grupo transferido
Precursor na dieta do
mamífero
Biocitina
CO2
biotina
Coenzima A
Grupos acilo
Ácido pantoténico
Coenzima B12
H, grupos alquilo
Vit B12
FAD
Electrões
Riboflavina (Vit B2)
Lipoato
Electrões, grupos
acilo
Não necess. na dieta
NAD+
Ião hidreto (:H-)
Ácido nicotínico
Fosfato de piridoxal
Grupos amina
Piridoxina (Vit B6)
Tetrahidrofolato
Grupos de 1C
Folato
Pirofosfato de tiamina
Aldeídos
Tiamina (Vit B1)
Os enzimas são altamente específicos:
Estereoespecificidade – reconhecem substratos quirais
Especificidade geométrica – são selectivos em relação
aos grupos químicos dos substratos.
Os enzimas variam no grau de especificidade geométrica
Regulação da actividade enzimática
Controlo da quantidade de enzima
Controlo da actividade enzimática
Funções termodinâmicas úteis para compreender o
funcionamento dos enzimas
• Variação de energia livre de Gibbs entre os produtos e
os reagentes
•Energia necessária para iniciar a conversão de
reagentes em produtos
A variação da energia livre padrão de uma reacção
está relacionada com a constante de equilíbrio
Relação entre K’eq e ∆G’o
S ↔P
K eq´= [P]/[S]
∆Go = -RT lnK´eq
Como é que os enzimas funcionam?
Os enzimas fornecem um
ambiente em que a reacção
ocorre mais rapidamente.
A reacção tem lugar no local
activo do enzima.
A molécula que se liga ao
local activo é chamada
substrato. Forma-se um
complexo enzima-substrato
Os enzimas afectam as velocidades de reacção e
não o equilibrio
E + S ↔ES ↔ EP ↔E + P
Os catalisadores aumentam a velocidade das reacções
baixando a energia de activação
A reacção chega mais depressa ao equilíbrio
A velocidade de reacção é determinada pela
concentração do(s) reagente(s) e pela constante de
velocidade, k.
v = k[S]
A constante de velocidade de uma reacção está
relacionada com a energia de activação:
k = kT/h . e- ∆G*/RT
k – constante de Boltzman
h – constante de Planck
Uma energia de activação mais baixa significa uma
maior velocidade de reacção – para um reacção ser
acelerada por um factor de 10, a energia de activação
tem que baixar cerca de 5,7 kJ/mole
2 H2O2
2 H2O + O2
Catalisador
Eactiv (Jmol-1)
-----------------
------------------------
Pt coloidal
Catalase
75 600
49 140
8 400
Como é que os enzimas conseguem aumentar tanto as
velocidades das reacções?
Interacções covalentes entre enzimas e substratos
baixam a energia de activação fornecendo uma via
reaccional alternativa com energia mais baixa.
Formação de um complexo específico enzima substrato
em que são formadas muitas interacções fracas. A
energia libertada aquando da formação destas ligações
contribui para a especificidade e para a catálise.
As ligações fracas são optimizadas no estado de
transição. Os locais activos são complementares do
estado de transição e não do substrato.
Formação de um complexo enzima-substrato.
As interacções fracas entre o enzima e o substrato são
optimizadas no estado de transição
Para baixar a energia de activação o sistema deve adquirir
uma quantidade de energia equivalente à que diminui.
Muita desta energia (∆GB )vem da formação de ligações
fracas entre o enzima e o substrato no estado de transição.
glucocinase
PDB ID 2YHX
“A catálise pode ser descrita formalmente
em termos de estabilização do estado de
transição através de ligação forte (tight
binding) do reagente ao catalisador”
Jencks 1975
Ligação do reagente
(substrato) ao centro activo
do enzima(quimotripsina)
Aumentos de velocidade produzidos por
enzimas:
Ciclofilina
Anidrase carbónico
P-triose isomerase
Carboxipeptidase A
Fosfoglucomutase
Succinil - coA transferase
Urease
Monofosfato de oritidina descarboxilase
O tamanho dos enzimas reflecte a necessidade de a
superestrutura ter os grupos reactivos posicionados de
modo correcto e de manter o local activo formado.
Os enzimas podem ser inibidos por moléculas
específicas
Inibição irreversível - os inibidores ligam-se
covalentemente (ou não)ou destroiem um grupo
funcional essencial para a actividade do enzima.
A penicilina modifica por meio de ligações covalentes o
enzima transpeptidase, impedindo a síntese das
paredes bacterianas.
A aspirina inibe o enzima que catalisa o 1º passo da
síntese das prostaglandinas, compostos envolvidos nos
processos de produção de dor.
Inactivadores-suicidas
Compostos pouco reactivos que se ligam ao
enzima no local activo, sofrem os primeiros
passos da reacção normal mas não se
transformam em produto. Convertem-se num
composto muito reactivo que se combina
irreversivelmente com o enzima.
Design de fármacos
Especificidade – menos efeitos secundários
Inibição reversível – Rápida
dissociação do complexo
enzima-inibidor.
Inibição competitiva – um
inibidor competitivo compete
com o substrato para o local
activo do enzima. O inibidor é
semelhante ao substrato.
Substrato
Enzima
Inibidor
competitivo
Enzima
Ex. malonato (inibidor) e succinato (substrato da
succinato desidrogenase)
-OOCCH
2
-
COO-
malonato
OOCCH2 CH2 COO- succinato
Metotrexato – anticancerígeno
Inibidor competitivo do dihidrofolato reductase
Tratamento para a ingestão de metanol (solvente
existente nos anticongelantes).
Metanol ↔ formaldeído (provoca cegueira)
desidrogenase alcoólica do fígado
Etanol é inibidor competitivo do metanol e também
é substrato para o enzima dando origem a
acetaldeído.
Outros tipos de inibição reversível:
Inibição incompetitiva – o inibidor liga-se a um
local diferente do local activo. Liga-se ao
complexo ES
Substrato
Inibidor
incompetitivo
Enzima
Inibição mista – o inibidor também se
liga a um local diferente do local activo.
Liga-se a E ou a ES
Enzimas regulatórios
Aumentam ou diminuem a sua actividade
catalítica em resposta a sinais.
Os enzimas alostéreos – funcionam através de
ligações reversíveis de compostos regulatórios.
Outros enzimas são regulados por ligações
covalentes reversíveis.
Regulação alostérea
Forma activa
Forma inactiva
Substrato ligado
ao local activo
Inibidor ligado ao
local alostéreo
Alguns enzimas são regulados por modificações
covalentes reversíveis
Modificação covalente (resíduos alvo)
Fosforilação ( Tyr, Ser, Thr, His)
Inactivo
Activo
A actividade enzimática depende do pH
Pepsina – estômago
G-6-P - hepatócitos
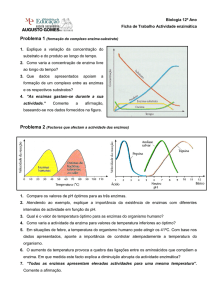
Perfil da actividade enzimática vs pH de dois
enzimas
Como estudar o mecanismo de acção dos
enzimas?
•Cinética enzimática
•Estudos estruturais
•Química de proteínas
•Mutagénese dirigida
Efeito da concentração de substrato na velocidade
inicial de uma reacção enzimática
O modelo de Michaelis - Menten
A relação entre a concentração de substrato e a
velocidade de reacção pode ser expressa
quantitativamente
Equação de
Michaelis-Menten
Vmax –velocidade máxima; KM – constante de Michaelis
Dependência da velocidade inicial da concentração de
substrato
Quando [S] <<< KM , então v0 = Vmax [S] /KM
A velocidade é directamente proporcional a [S]
Quando [S] >>> KM , então v0 = Vmax
A velocidade é independente de [S]
Quando v0 = Vmax /2, então KM = [S]
KM é a conc. de substrato para a qual v0 = Vmax /2
Transformações da equação de MichaelisMenten – a equação de Lineweaver-Burk (duplo
recíproco)
Gráfico de Lineweaver-Burk
Os parâmetros cinéticos são usados para
comparar actividades enzimáticas
A equação de Michaelis descreve o comportamento
cinético dos enzimas em que v0 tem uma dependência
hiperbólica de [S]. Estes enzimas seguem uma cinética de
Michaelis-Menten.
Excepções enzimas regulatórios.
Parâmetros cinéticos:
KM
Vmax
Significado dos parâmetros cinéticos KM e Vmax
KM – Concentração de substrato para a qual a
velocidade é igual a metade da Vmax .
[S] necessária para ocorrer uma catálise significativa.
Consequência fisiológica do KM
Fígado
etanol
↔
acetaldeído
desidrogenase alcoólica
acetaldeído
↔
acetato
desidrogenase do acetaldeído
Desidrogenase do acetaldeído:
Enzima mitocondrial – baixo KM
Enzima citosólico – elevado KM
Em pessoas susceptíveis ao etanol o enzima mitocondrial é
menos activo.
O KM é muitas usado como indicador da afinidade do
enzima para o substrato.
O KM está relacionado com as constantes de
velocidade:
Se k2 <<< k-1
Elevado KM baixa afinidade do enzima para o
substrato
Baixo KM elevada afinidade
Vmax
Vmax = kcat [Et ]
Representa o nº de moléculas de substrato
convertidas em produto por uma molécula de
enzima, numa unidade de tempo, quando o
enzima está completamente saturado com o
substrato.
kcat – Nº de turnover
Exemplo:
Foi descoberto um enzima que catalisa a reacção
Triste ↔ Contente
Uma equipa de investigadores altamente motivados
resolve estudar o enzima que chamaram contentase
e verificaram que o kcat para este enzima era 600 s-1
.
Quando [Et ] = 20 nM e [Triste] = 40 µM, a velocidade
da reacção V0 é 9,6 µM s-1 . Calcular o KM para o
substrato “Triste”.
R: 10 µM
Inibição competitiva
A competição pode ser desviada para favorecer
o substrato simplesmente adicionando mais
substrato.
Quando [S] >>> [I] a probabilidade do inibidor se
ligar é minimizada e a reacção tem a Vmax normal.
No entanto [S] para o qual V0 =1/2 Vmax (KM )
aumenta.
Vmax mantem o valor.
KM - aumenta
Representação de Lineweaver-Burk para
inibidores competitivos
Os investigadores a trabalhar na enzima
contentase descobriram que o composto
STRESS era um potente inibidor competitivo
do enzima contentase.
A adição de STRESS aumenta o valor de KM
para o dobro.
O que deverá ser feito para a reacção manter
a sua velocidade máxima normal?
Os outros tipos de inibição reversível com
diferentes variações dos parâmetros cinéticos
(inibição anticompetitiva e mista) são observados
em geral em reacções enzimáticas com mais que
dois substratos.
Enzimas regulatórios
Aumentam ou diminuem a sua actividade
catalítica em resposta a sinais.
Os enzimas alostéreos – funcionam através de
ligações reversíveis de compostos regulatórios.
Outros enzimas são regulados por ligações
covalentes reversíveis.
Os enzimas alostéreos não obedecem à
cinética de Michaelis-Menten
.
Regulação da actividade enzimática
Em muitas vias os passos de regulação são catalisados
por enzimas alostéreos.
Ex.
Inibição por “feed-back”
Os enzimas alostéreos apresentam uma cinética
sigmoidal que reflecte interacções cooperativas
entre as subunidades da proteína