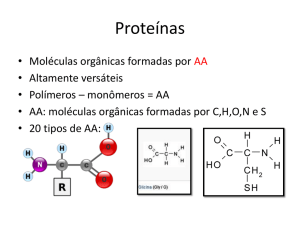
Me
etabollismo
o e En
ndocrinolo
ogia IST – FML
L 2
2º Seme
stre 200
07/2008 Re
elatório d
de: C
Cinétic
ca e reg
gulaçã
ão enziimáticca (a) Definiçã
ão e deduçção da equ
uação de Michaelis­M
M
Menten e respectiva
a aplicação
o, cálculo da enzimática
a pela equa
ação de Lin
neweaver­­Burk; acctividade e
(b
b) Inibição
o reversíve
el e irrev
versível, ap
plicação das d equaçõ
ões e reve
ersibilidad
de à enzim
ma he
exocinase e
e respectiv
va reacção
o catalisad
da; (c)) Enzima
as reguladoras, tip
pos de modulado
ores e cinética, m
modulação
o covalen
nte (fo
osforilação
o, adenilaçção, uridilação, ADP­ribosilaçã
ão e metila
ação), e zim
mogénios.. Trabalho
o realizado p
por: Miguel Am
mador nº584
484 Joana Nunes nº584
497 João Maarques nº585
513 Metabolismo e Endocrinologia – Cinética e regulação enzimática – 2º Semestre 2007/2008 Cinética Enzimática A cinética enzimática é o estudo da velocidade de uma reacção química que ocorre na presença de uma enzima e permite elucidar os pormenores do mecanismo catalítico das enzimas, o seu papel no metabolismo, como é controlada a sua actividade e como pode ser inibida. Para que consigamos estudar melhor os mecanismos de uma reacção catalisada por uma enzima é determinar as constantes dessas reacções. A velocidade da reacção é influenciada pela concentração de substrato, através de uma dependência hiperbólica, ou seja, existe um limite de velocidade que nunca é atingido (velocidade máxima) que corresponde ao caso hipotético em que todas as enzimas estariam ocupadas. Segundo Michaelis e Menten, inicialmente, a enzima liga‐se ao substrato reversivelmente formando o complexo enzima substrato (ES). Esta reacção é rápida. Seguidamente o complexo ES é dissociado na enzima livre e no produto da reacção, sendo que esta reacção é mais lenta que a primeira. (1) (2)
Isto faz com que a reacção (2), a mais lenta, limite a velocidade da reacção global, velocidade esta que será proporcional à concentração de complexo ES (o reagente dessa reacção). Em cada momento da reacção a enzima existe nas duas formas, e assim podemos concluir que a velocidade máxima (Vmáx) da reacção ocorre quando a enzima está saturada. A curva que representa a relação entre [S] e Vo tem forma idêntica para a maior parte das enzimas e é descrita algebricamente pela equação de Michaelis‐Menten. Equação de Michaelis­Menten Para obter a expressão da velocidade em função da concentração de substrato, põe‐se por hipótese que o passo limitante da velocidade das reacções é a dissociação do complexo ES, e ainda que o produto não volta a dar origem a substrato. (E)
Vo pode ser considerado como a velocidade com que ocorre a quebra da ligação ES, que é dessa forma determinada pela [ES], e é dado por (a). Define‐se [ET] como concentração total da enzima, e consideramos que a concentração de enzima livre é [ET]‐[ES]. Passo 1: As velocidades de formação e degradação do complexo ES podem ser dadas por: Velocidade de formação de ES
Velocidade de dissociação de ES
Passo 2: A reacção está num estado constante (“steady state”), logo as velocidades são iguais: Passo 3: Resolvendo a equação em ordem a [ES], vem
Definindo a expressão /
como a constante de Michaelis (Km) obtemos a expressão: Passo 4: Podemos agora obter Vo, substituindo [ES] na equação (a): Como á
. , pois a velocidade é máxima quando [ES]=[ET]: á
Metabolismo e Endocrinologia – Cinética e regulação enzimática – 2º Semestre 2007/2008 Assim obtemos a equação de Michaelis‐Menten, que nos dá a relação quantitativa entre a velocidade inicial V0, a velocidade máxima Vmáx e a quantidade de substrato [S], todas relacionadas pela constante de Michaelis Km. O Km tem unidades de concentração, mede a afinidade da enzima para com o substrato (nas enzimas que seguem a cinética de Michaelis‐Menten, ou seja, que exibem uma dependência hiperbólica de V0 em função de [S]), e corresponde à concentração de substrato para a qual V0 é metade de Vmáx. A equação de Michaelis‐Menten é muito útil para determinar os valores de Km e Vmáx das reacções. Estes parâmetros são importantes no estudo e comparação da actividade das enzimas. A equação de Lineweaver­Burk Invertendo a equação de Michaelis‐Menten, obtemos a equação de Lineweaver Burk, que nos permite um cálculo facilitado dos parâmetros Vmáx e Km , pois a intersecção da recta no eixo 1/V0 corresponde ao valor 1/Vmáx, e a intersecção com o eixo 1/[S] corresponde a ‐1/Km: 1
1
á
á
Inibição enzimática A actividade de um tipo de enzimas nem sempre é constante. Isto porque podem ser inibidas por factores do meio, que pode ser de dois tipos, reversível e irreversível. Na inibição irreversível, o inibidor liga‐se permanentemente à enzima, inutilizando‐a. O inibidor pode ligar‐se covalentemente à enzima, pode destruir um grupo funcional essencial ao funcionamento da mesma, ou pode ainda formar uma ligação não covalente particularmente estável. A inibição reversível divide‐se em 3 tipos: competitiva, anti‐competitiva e mista: Competitiva Neste tipo de inibição, o substrato e o inibidor competem pelo centro activo da enzima. Assim, é possível contrariar a inibição adicionando mais substrato ao meio, aumentando a probabilidade da enzima se ligar ao substrato e não ao inibidor. Neste tipo de inibição, é comum que o inibidor seja estruturalmente semelhante ao . Nesta equação, substrato. Em termos da equação de Michaelis‐Menten, esta é da seguinte forma:
1
, onde obtém‐se: é a concentração de inibidor e . Invertendo a equação de Michaelis‐Menten, . Assim conclui‐se que Vmax não se altera e o Km aumenta. Anti­Competitiva Neste tipo de inibição, o inibidor liga‐se a um local específico da enzima que não o centro activo. Aqui, o inibidor só se liga ao complexo enzima‐substrato (ES), formando o complexo enzima‐substrato‐inibidor (ESI). Em termos da equação de Michaelis‐Menten, esta vem afectada da seguinte forma:
onde é a concentração de inibidor e , sendo que, 1
, . Invertendo a equação de Michaelis‐Menten, obtém‐se: . Através desta equação, pode concluir‐se que Vmax e Km diminuem. Mista Neste tipo de inibição, tal como no anterior, a enzima liga‐se a um local específico da enzima que não ao centro activo. Não há competição pelo centro activo. Aqui, o inibidor pode ligar‐se ao complexo ES, formando o complexo ESI, ou pode ligar‐se apenas à enzima, formando o complexo EI, e o substrato pode ainda ligar‐se a este complexo EI formando o complexo ESI. Em termos da equação de Michaelis‐Menten, é da seguinte forma: . Nesta equação, e são definidos como nas inibições acima, bem como o e o . Metabolismo e Endocrinologia – Cinética e regulação enzimática – 2º Semestre 2007/2008 Invertendo a equação de Michaelis‐Menten, obtém‐se: Km pode aumentar, diminuir ou manter‐se, pois aparente =
. Assim conclui‐se que Vmax diminui e . O Exemplo da Hexocinase A hexocinase é uma enzima de extrema importância, principalmente porque é a enzima responsável pela fosforilação da glicose em glicose‐6‐fosfato. Esta reacção ocorre com degradação de ATP, e na presença de um ião Mg2+. A hexocinase é uma enzima de tipo indutivo, ou seja, a glicose induz na hexocinase a forma correcta para que se dê a reacção. A hexocinase não é específica da glicose, ou seja, catalisa também outras reacções. O Km para a glicose é de 0.1mM, e a concentração desta na célula está à volta dos 4mM, o que quer dizer que a hexocinase está normalmente activa dentro da célula. A fosforilação da glicose impede a sua saída da célula por difusão, pois a glicose atravessa facilmente a membrana plasmática, enquanto que a glicose‐6‐fosfato não o faz. A hexocinase é regulada alostericamente pelo seu próprio produto, ou seja, a glicose‐6‐fosfato é o inibidor da hexocinase. A fosforilação da glicose está englobada na via metabólica da glicólise. No fígado existe também uma hexocinase, mas com pouca afinidade para com a glicose, o que faz com que esta apenas esteja activa quando a concentração de açúcar no sangue está muito elevada. Aqui, a glicose é convertida em glicogénio. A reacção catalisada pela hexocinase é reversível, ou seja a glicose‐6‐fosfato pode ser reconvertida em glicose no fígado, e esta reacção faz parte da gliconeogénese. Enzimas Reguladoras No metabolismo celular, grupos de enzimas trabalham em conjunto, em caminhos sequenciais encarregues de um processo metabólico. Neste tipo de sistemas, os produtos de reacção de uma enzima tornam‐se o substrato da seguinte. O funcionamento de cada enzima é independente, pelo que normalmente seguem os padrões cinéticos que já foram descritos. A cinética enzimática depende, em grande parte, da quantidade de substrato. Assim, certas enzimas possuem um grande efeito na taxa global da reacção sequencial, chamadas enzimas reguladoras, já que aumentam ou diminuem a sua actividade em resposta a certos sinais. Esta regulação é essencial, já que permite, ao regular a actividade de certas enzimas, regular a actividade inteira da sequência metabólica, possibilitando a célula ajustar‐se às suas necessidades energéticas e biomoleculares. Geralmente, e de forma a aumentar a eficiência, a primeira enzima da sequência é a enzima reguladora. Tipos de Moduladores Para além dos tipos de factores que podem inibir a reacção existem dois modelos de regulação enzimática mais conhecidos – modulação alostérica e covalente. Nas enzimas alostéricas, a modulação funciona através da ligação não covalente, reversível, de substâncias chamadas modeladores alostéricos ou efectores alostéricos. A zona regulada e a zona activa podem estar em duas subunidades distintas ou numa só. As enzimas reguladoras sofrem, por acção dos moduladores, alterações conformacionais que alternam entre formas mais ou menos activas da enzima. Por vezes o modulador de enzimas alostéricas é o mesmo que o substrato, e neste caso a enzima é chamada homotrópica. No caso de serem diferentes diz‐se serem heterotrópicas. As propriedades de enzimas alostéricas são significativamente distintas das enzimas simples, não reguladas. Assim como cada centro activo é específico para um substrato, também os chamados centros alostéricos são específicos para cada modelador. No caso de enzimas homotrópicas, o centro activo e regulador tende a ser o mesmo. Em algumas vias metabólicas, a enzima reguladora é especificamente inibida pelo produto final da via, uma vez que a concentração deste exceda os requerimentos celulares. Este tipo de regulação é denominada inibição de “retroalimentação”. Metabolismo e Endocrinologia – Cinética e regulação enzimática – 2º Semestre 2007/2008 Na modulação covalente, as enzimas são activadas quando pedaços peptídicos são removidos por clivagem proteolítica, que ao contrário das anteriores é geralmente irreversível, já que exige formação ou destruição de ligações covalentes. Uma mesma enzima pode ser regulada de diversas formas, já que a regulação alostérica permite um ajustamento mais sensível, importante, por exemplo, em metabolismos contínuos, enquanto a regulação por modificações covalentes permite mudanças mais significativas. Cinética das Enzimas Reguladas As propriedades cinéticas das enzimas alostéricas divergem do comportamento indicado por Mechaelis‐Menten. Continua a existir uma saturação para altas concentrações de substrato, [S], mas a velocidade de reacção, em função da [S], apresenta uma saturação sigmóide ao invés da normal curva hiperbólica. Apesar de ser possível identificar o valor de [S] para o qual a velocidade de reacção é metade da velocidade máxima, não podemos identificar esse valor como Km , já que não possui o comportamento hiperbólico definido por Michaelis‐Menten, optando‐se pela simbologia [S]0,5. Para o caso das enzimas alostéricas homotrópicas a sua cinética sigmóide tem a característica de que pequenas variações na concentração do modulador, no inicio da curva, podem implicar grandes um aumento significante na velocidade de reacção, já que o substrato é ao mesmo tempo o modelador. Para as enzimas alostéricas heterotípicas, é díficil generalizar a forma da curva [S]/V. Um modelador pode alterar a actividade de uma enzima para um determinada concentração, mas não alterar a sua velocidade máxima de actividade. Outros podem alterar o valor de velocidade máxima, não tendo grande efeito na afinidade. As principais dificuldades surgem do facto de a enzima heterotípica poder ser regulada positivamente ou negativamente pelos diferentes modeladores, pelo que, dependendo dos com que interage, apresenta diferentes tipos de resposta nas suas curvas [S]/V. Modulação Covalente Outro tipo de enzimas reguladoras é modulado por modificações covalentes da molécula que a constitui. Certos grupos são colocados ou retirados da enzima com intervenção de outras enzimas. As principais modificações covalentes são: fosforilação, adenilação, uridilação, ADP‐ribosilação e metilação. A fosforilação é o tipo mais comum de modificações reguladoras. A ligação de grupos fosforilo a certos resíduos de aminoácidos de uma proteína é catalisada por proteínas denominadas cinases, a sua remoção é levada a cabo por uma proteína denominada fosfatase, permitindo a reversibilidade deste processo. Os grupos fosforilo influenciam a polaridade dos aminoácidos aos quais se encontram ligados, contribuindo com forças de atracção e repulsão entre os resíduos proteicos, para além de permitir o estabelecimento de pontes de hidrogénio com vários grupos de proteínas, sendo assim importante para a estrutura e conformação da enzima. As restantes modificações covalentes são menos frequentes, mas apresentam os mesmos mecanismos de adição de um grupo específico a um resíduo, com intervenção de enzimas específicas. Na adenilação é adicionado um grupo adenilo à enzima. Na uridilação é acrescentado um grupo uridilo. Na ADP‐Ribosilação, em que é acresentado uma ADP‐Ribose. A metilação, consiste na adição de um grupo metilo, nomeadamente em resíduos de Glutamato. Zimogénios A regulação enzimática pode passar ainda pela existência de um precursor, sem capacidade catalítica, que, no caso das proteases, são chamados zimogénios, como é exemplo o par pepsinogénio/pepsina. Uma clivagem proteolítica altera a conformação do precursor e expõe o centro activo da enzima, tornando‐a activa. Este processo é no entanto irreversível. Não só as proteases são activadas por proteólise, mas nesse caso não se chamam zimogénios, mas sim proproteínas ou proenzimas, consoante for apropriado.