Carlos Capela
ENZIMAS
ENZIMAS ................................................................... 1
A.
INTRODUÇÃO ..........................................................3
1.
CONCEITO ..................................................................... 3
2.
PROPRIEDADES ............................................................. 3
3.
NOMENCLATURA E CLASSIFICAÇÃO ........................... 4
4.
COFACTORES ................................................................ 5
4.a.
4.b.
4.c.
B.
Coenzimas .................................................................................5
Iões Metálicos ...........................................................................5
Grupos Prostéticos ....................................................................6
CINÉTICA ENZIMÁTICA..........................................7
1.
VELOCIDADE INICIAL DA REACÇÃO............................. 8
2.
INFLUÊNCIA DA [E] NA CINÉTICA ............................... 8
3.
INFLUÊNCIA DA [S] NA CINÉTICA ................................ 9
3.a.
3.b.
3.c.
Equação de Michaelis-Menten (1913).....................................9
A representação de Lineweaver-Burk....................................10
Inibição Enzimática................................................................11
3.C.I.
3.C.II.
3.C.III.
INIBIÇÃO ENZIMÁTICA REVERSÍVEL COMPETITIVA ..................... 11
INIBIÇÃO ENZIMÁTICA REVERSÍVEL NÃO-COMPETITIVA ............ 11
INIBIÇÃO ENZIMÁTICA REVERSÍVEL ANTICOMPETITIVA ............. 11
4. FACTORES EXTERNOS QUE INFLUENCIAM A
VELOCIDADE DE UMA REACÇÃO ENZIMÁTICA .................. 12
4.a.
4.b.
Temperatura............................................................................12
pH ............................................................................................12
Enzimas página 1 de 25
Carlos Capela
C.
MECANISMOS DE ACÇÃO ENZIMÁTICA...............14
1.
O CENTRO ACTIVO ...................................................... 14
2.
MODELOS .................................................................... 15
2.a.
2.b.
3.
MECANISMOS DE ACÇÃO............................................ 17
3.a.
3.b.
3.c.
3.d.
D.
Modelo Chave/Fechadura (Emil Fischer, 1890) ..................15
Modelo do Ajuste Induzido (Daniel E. Koshland, Jr em 1958)
.................................................................................................15
Catálise Ácido-Base ................................................................17
Torção do Substrato................................................................17
Catálise Covalente ..................................................................18
Efeitos de Diminuição da Entropia .......................................18
REGULAÇÃO DA ACTIVIDADE ENZIMÁTICA .......19
1.
MODULAÇÃO ALOSTÉRICA ........................................ 19
1.a.
1.b.
1.c.
Enzimas alostéricas apresentam cinética sigmoidal .............20
Modelo de transição concertada ............................................21
Modelo sequencial ..................................................................21
2.
MODULAÇÃO COVALENTE ......................................... 23
3.
ACTIVAÇÃO PROTEOLÍTICA....................................... 23
E.
ENZIMAS NA CLÍNICA...........................................24
1.
1.a.
1.b.
AS ISOENZIMAS ........................................................... 24
Creatina fosfocinase (CPK)....................................................24
Lactato desidrogenase (LDH) ................................................25
Enzimas página 2 de 25
Carlos Capela
A. INTRODUÇÃO
1.CONCEITO
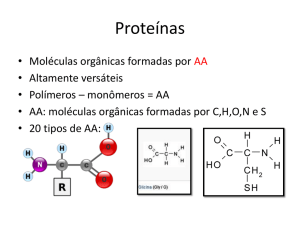
Enzimas são proteínas globulares com actividade catalítica.
Praticamente todas as reacções que caracterizam o metabolismo celular são
catalisadas por enzimas.
Como catalisadores celulares ou biológicos extremamente poderosos, as enzimas
aceleram a velocidade de uma reacção, sem no entanto participar dela como reagente ou
produto. As enzimas ainda actuam como reguladoras deste conjunto complexo de
reacções.
As enzimas são, portanto, consideradas as unidades funcionais do metabolismo
celular.
2.PROPRIEDADES
•
São catalisadores biológicos extremamente eficientes – Aceleram em média 109 a
1012 vezes a velocidade da reacção, transformando de 100 a 1000 moléculas de
substrato em produto, por minuto de reacção.
•
Diminuem a energia de activação necessária à reacção, permitindo que esta ocorra a
uma velocidade mais elevada;
•
Encontram-se intactas no final da reacção;
•
Actuam em concentrações muito baixas;
•
Actuam em condições suaves de temperatura e pH;
•
Possuem todas as características das proteínas;
•
Podem ter a sua actividade regulada;
•
Estão quase sempre dentro das células, compartimentalizadas.
Enzimas página 3 de 25
Carlos Capela
3.NOMENCLATURA E CLASSIFICAÇÃO
Existem 3 métodos para nomenclatura enzimática:
•
Nome Usual: Consagrados pelo uso; Exs: Tripsina, Pepsina, Ptialina.
•
Nome Recomendado: Mais curto e utilizado no dia-a-dia de quem trabalha com
enzimas; em muitos casos trata-se de um nome já correntemente utilizado. Este
consiste em 2 partes (a 1ª contêm o nome do substrato e a 2ª terminada em “ase”
indica a natureza da acção). Exs: Lactato-desidrogenase, Hexoquinase, etc.
•
Nome Sistemático: formado de acordo com regras definitivas, revela a acção de
uma enzima tão completamente quanto possível, identificando-a com precisão.
Não é nada mais de que um código atribuído às enzimas. Estes números,
precedidos por EC (Enzyme Classification), contêm 4 algarismos separados por
pontos:
O 1º algarismo indica a qual das seis categorias ou classes pertence a enzima;
O 2º algarismo e o 3º algarismo indicam a subclasse e a subsubclasse;
O 4º algarismo é o número de série na respectiva subsubclasse.
Ex: a Lactato-desidrogenase é classificada como EC.1.1.1.27
As classes das enzimas são as seguintes:
1. Oxidorredutases: São enzimas que catalisam reacções de transferência de
electrões, ou seja: reacções de oxi-redução. Ex: as Desidrogenases e as Oxidases
2. Transferases: Enzimas que catalisam reacções de transferência de grupos
funcionais como grupos amina, fosfato, acil, carboxil, etc. Como exemplo temos
as Quinases e as Transaminases
3. Hidrolases: Catalisam reacções de hidrólise de ligações covalentes. Ex: As
peptidases
4. Liases: Catalisam a quebra de ligações covalentes e a remoção de moléculas de
água, amónia e dióxido de carbono. As Desidratases e os Descarboxilases são
bons exemplos.
5. Isomerases: Catalisam reacções de interconversão entre isómeros ópticos ou
geométricos. As Epimerases e as mutases são exemplos.
6. Ligases (ou Sintetases): Catalisam reacções de formação de novas moléculas a
partir da ligação entre duas já existentes, à custa de energia (ATP). Ex:
aminoacil-tRNA sintetases.
Enzimas página 4 de 25
Carlos Capela
4.COFACTORES
Um grande número de enzimas necessita, para a sua acção catalítica de um cofactor.
Cofactores são pequenas moléculas orgânicas ou inorgânicas que podem ser
necessárias para a função de uma enzima.
Estes cofactores podem ou não estar ligados permanentemente à molécula da
enzima, mas na ausência deles, a enzima é inactiva.
A fracção proteica de uma enzima, na ausência do seu cofactor, é denominada de
Apoenzima. Enzima + Cofactor, chamamos de Holoenzima.
Os cofactores são divididos em 3 grupos: as coenzimas, os iões metálicos e os
grupos prostéticos.
4.a. Coenzimas
São compostos orgânicos, quase sempre derivados de vitaminas, que actuam em
conjunto com as enzimas. Podem actuar segundo 3 modelos:
•
Ligando-se à enzima com afinidade semelhante à do substrato;
•
Ligando-se covalentemente num local próximo ou no próprio sítio catalítico da
apoenzima;
•
Actuando de maneira intermediária aos dois extremos acima citados.
Ex: NAD+ e NADP+, FMN e FAD, Pirofosfato de Tiamina, Fosfato de Piridoxal, etc.
4.b. Iões Metálicos
Metal
Tipos de Enzima
Fe2+/Fe3+
Citocromos, Peroxidases, Catalases
Cu+/Cu2+
Citocromo-oxidase, Tirosinase
Mg2+
Fosfotransferases
Mn2+
Fosfotransferases, Arginase
Zn2+
Álcool-desidrogenase, Anidrase Carbónica
+
K
Piruvato-cinase
Na+
ATPase membranária
Enzimas página 5 de 25
Carlos Capela
4.c. Grupos Prostéticos
São cofactores de natureza não proteica, mas fortemente ligados à apoenzima. É
exemplo o grupo porfirina nas catalases, peroxidases e citocromos.
Enzimas página 6 de 25
Carlos Capela
B. CINÉTICA ENZIMÁTICA
É a parte da enzimologia que estuda a velocidade das reacções enzimáticas, e os
factores que influenciam esta velocidade.
A cinética de uma enzima é estudada avaliando-se a quantidade de produto formado
ou a quantidade de substrato consumido por unidade de tempo de reacção (velocidade
de reacção). Uma reacção enzimática pode ser expressa pela seguinte equação:
E + S ⇔ [ES] ⇒ E + P
O complexo enzima/substrato (ES) tem uma energia de activação ligeiramente
menor que a do substrato isolado, e a sua formação leva ao aparecimento do estado de
transição “Ts”. A formação de “P” a partir de ES é a etapa limitante da velocidade da
reacção. A velocidade de uma reacção enzimática depende das concentrações da Enzima
e do Substrato.
Quando tratamos da cinética de reacções enzimáticas há que ter em atenção:
1) Velocidade inicial da reacção;
2) Influência da concentração da enzima na cinética;
3) Influência da concentração do substrato na cinética;
4) Influência de Factores Externos.
Enzimas página 7 de 25
Carlos Capela
1.VELOCIDADE INICIAL DA REACÇÃO
A velocidade da reacção enzimática não se mantêm constante ao longo do tempo,
decrescendo progressivamente. Assim, é necessário indicar a que tempo se refere uma
certa velocidade. Considera-se a velocidade inicial v0, numericamente igual ao declive
da recta tangente ao gráfico de produto formado em função do tempo. A velocidade
inicial só depende da:
•
Concentração da enzima
•
Concentração do substrato
•
Afinidade da enzima pelo substrato
2.INFLUÊNCIA DA [E] NA CINÉTICA
Estudos mostraram que a velocidade inicial varia na razão directa da quantidade de
enzima.
Enzimas página 8 de 25
Carlos Capela
3.INFLUÊNCIA DA [S] NA CINÉTICA
Para baixas concentrações de substrato, a velocidade é aproximadamente
proporcional à sua concentração (reacção dita de 1ª ordem em relação ao substrato),
mas, para altas concentrações, a velocidade tende para um valor assímptótico, designado
por velocidade máxima, Vmáx. Nesta zona, v é praticamente invariável com a
concentração de substrato (reacção dita de ordem zero).
Ordem mista
Ordem zero
1ª ordem
3.a. Equação de Michaelis-Menten (1913)
Michaelis e Menten foram 2 pesquisadoras que propuseram o modelo acima citado
como modelo de reacção enzimática para apenas um substrato. A partir deste modelo,
estes pesquisadores criaram uma equação, que nos permite demonstrar como a
velocidade de uma reacção varia com a variação da concentração do substrato. Esta
equação pode ser expressa graficamente, e representa o efeito da concentração de
substrato sobre a velocidade de reacção enzimática.
Vmáx × [S ]
v=
Km + [S ]
Desta expressão podemos retirar que Km corresponde à concentração de substrato em
que a velocidade é ½ da velocidade máxima. Isto significa que, para esta concentração, a
enzima está 50% saturada, (pois a Vmáx verifica-se quando a totalidade da enzima está
saturada, ou seja, na forma ES).
Enzimas página 9 de 25
Carlos Capela
O Km de um substrato para uma enzima específica é característico, e fornece-nos um
parâmetro de especificidade desse substrato em relação à enzima. Essa especificidade ou
afinidade é dada por 1/Km, ou seja, quanto menor o Km, maior a especificidade, e viceversa.
3.b. A representação de Lineweaver-Burk
Como o cálculo do Km não é rigoroso pois trata-se de uma aproximação e, por outro
lado o traçado da curva de Michaelis exige um número elevado de determinações, surgiu
a alternativa linear: Lineweaver-Burk, de 1/v em função de 1/[S].
Invertendo ambos os membros da equação de Michaelis-Menten e isolando 1/v no
primeiro membro, pode transformar-se essa equação em:
1 Km
1
1
=
× +
v Vmáx [S ] Vmáx
Sob esta forma temos uma equação do tipo y = ax + b, onde 1/v e 1/[S] são as
variáveis y e x.
Se na equação, fizer y = 0, isto é, 1/v = 0, obtêm o valor -1/Km. Se fizer 1/[S] = 0 ou
x = 0, obtêm 1/Vmáx.
O valor do Km é lido com rigor, pois é simétrico do inverso da intersecção com o
eixo das abcissas.
Enzimas página 10 de 25
Carlos Capela
3.c. Inibição Enzimática
Os inibidores enzimáticos são compostos que ligando-se a uma enzima, diminuem a
sua actividade. A inibição enzimática pode ser reversível (quando o inibidor se liga à
enzima de um modo não covalente, sendo a actividade biológica da enzima recuperada
após a dissociação) e irreversível (há modificação covalente e definitiva no sítio de
ligação ou no sítio catalítico da enzima).
Existem 3 tipos de inibição enzimática reversível:
3.c.i. INIBIÇÃO ENZIMÁTICA REVERSÍVEL COMPETITIVA
O inibidor é estruturalmente semelhante ao substrato e liga-se reversivelmente ao
mesmo sítio de ligação do substrato. O efeito é revertido aumentando-se a concentração
de substrato. Este tipo de inibição depende das concentrações do substrato e do inibidor.
O inibidor compete pelo sítio activo com o substrato, diminuindo por isso a afinidade
da enzima pelo substrato (Km aumenta), mas a Vmáx mantém-se inalterada. Isto significa
que torna-se necessária maior quantidade de substrato para atingir a mesma Vmáx .
2
3.c.ii. INIBIÇÃO ENZIMÁTICA REVERSÍVEL NÃO-COMPETITIVA
São inibidores que se fixam reversivelmente na enzima noutro local que não o centro
activo, não diminuindo a sua acção por adição de um excesso de substrato. A enzima
pode ligar-se ao mesmo tempo ao inibidor não competitivo e ao substrato.
Dado que o inibidor se liga num local diferente do centro activo, a enzima pode
continuar a ligar a si o substrato. No entanto, como a conformação do centro activo é de
algum modo afectada pela presença do inibidor, essa ligação efectua-se com menos
facilidade e a velocidade da reacção diminui. Um inibidor não competitivo comporta-se
como se removesse a enzima activa da solução, resultando na diminuição da velocidade
máxima, enquanto a afinidade não se altera.
Este tipo de inibição depende apenas da concentração do inibidor.
3.c.iii. INIBIÇÃO ENZIMÁTICA REVERSÍVEL ANTICOMPETITIVA
Neste tipo de inibição, o inibidor liga-se ao complexo ES, e não à enzima na forma
livre. Neste caso, tanto o Km como a Vmáx diminuem.
Enzimas página 11 de 25
Carlos Capela
4.FACTORES EXTERNOS QUE
INFLUENCIAM A VELOCIDADE DE UMA
REACÇÃO ENZIMÁTICA
4.a. Temperatura
A velocidade de todas as reacções químicas aumenta com a temperatura (maior
número de choques), mas isto só se aplica até determinado valor de temperatura –
Temperatura Óptima. Isto é, até ser atingida a temperatura a partir da qual a proteína
enzimática, por começar a desnaturar, vê também diminuir a sua actividade.
4.b. pH
Idem à temperatura; existe um pH Óptimo, onde a distribuição de cargas eléctricas
da molécula da enzima e, em especial do centro catalítico, é ideal para a catálise. A
ionização dos aminoácidos do centro catalítico pode inibir a catálise; bem como a
ionização de outros aminoácidos fora do centro catalítico, pode induzir uma alteração
conformacional, alterando assim a afinidade entre a enzima e o substrato. O pH pode
ainda modificar o grau de ionização do substrato.
Enzimas página 12 de 25
Carlos Capela
Verifica-se pois que cada enzima tem um pH óptimo de actuação, havendo portanto
enzimas que actuam em meio ácido, como as enzimas gástricas, outras em meio neutro
como as da boca e outras em meio alcalino como as do duodeno.
Enzimas página 13 de 25
Carlos Capela
C. MECANISMOS DE ACÇÃO
ENZIMÁTICA
1.O CENTRO ACTIVO
As enzimas são muito específicas para os seus substratos.
Esta especificidade pode ser relativa a apenas um substrato ou a vários substratos ao
mesmo tempo. Esta especificidade deve-se à existência, na superfície da enzima de um
local denominado Centro de ligação ao substrato ou Centro activo.
O centro de ligação ao substrato de uma enzima é dado por um arranjo
tridimensional especial dos aminoácidos de uma determinada região da molécula,
geralmente complementar à molécula do substrato, espacial e electricamente ideal para a
ligação do mesmo.
A ligação entre a enzima e o substrato pode envolver interacções iónicas, ligações
por pontes de hidrogénio e interacções van der Waals.
O centro de ligação do substrato é capaz de reconhecer inclusive isómeros ópticos
“D” e “L” de um mesmo composto.
O centro activo compreende o local de fixação (que se combina com o substrato por
ligações fracas), e o centro catalítico (que pode ou não estar próximo dele, mas é onde
ocorre a reacção enzimática).
Enzimas página 14 de 25
Carlos Capela
2.MODELOS
Alguns modelos procuram explicar a especificidade substrato/enzima:
2.a.Modelo Chave/Fechadura (Emil Fischer, 1890)
Prevê um encaixe perfeito do substrato no local de ligação, que seria rígido como
uma fechadura. Ou seja, o substrato deveria possuir uma determinada forma de modo a
poder-se “encaixar” na enzima.
2.b. Modelo do Ajuste
Koshland, Jr em 1958)
Induzido
(Daniel
E.
Prevê um local de ligação não totalmente pré-formado, mas sim moldável à molécula
do substrato; a enzima ajustar-se-ia na presença da molécula do substrato.
Enzimas página 15 de 25
Carlos Capela
Evidências experimentais sugerem um terceiro modelo que combina o ajuste
induzido a uma “torção” da molécula do substrato, que o “activaria” e o prepararia para
a sua transformação em produto.
Enzimas página 16 de 25
Carlos Capela
3.MECANISMOS DE ACÇÃO
As enzimas aceleram a velocidade de uma reacção por diminuir a Energia Livre de
Activação da mesma, sem alterar a termodinâmica da reacção, ou seja, a energia dos
reagentes e produtos da reacção enzimática e da sua equivalente não enzimática são
idênticas.
Para se superar a energia de activação de uma reacção, passa-se pela formação de um
estado intermediário chamado “Estado de Transição”, sempre um composto instável e de
alta energia, representado por “Ts”, ligado com altíssima afinidade ao centro catalítico.
Nas reacções enzimáticas, este composto de transição “Ts” não pode ser isolado ou
mesmo considerado um intermediário, uma vez que não é libertado para o meio da
reacção.
Como a afinidade do “Ts” ao centro catalítico é muito maior que a afinidade do
substrato ao mesmo, a pequena quantidade de moléculas em “Ts” será rapidamente
convertida em produto. Assim, todo o factor que leva a um aumento do número de
moléculas em “Ts” aumenta a velocidade da reacção.
São 4 os mecanismos principais através dos quais as enzimas aceleram uma reacção,
aumentando a formação de moléculas de substrato em “Ts”:
3.a. Catálise Ácido-Base
Ocorre com a participação de AA com cadeias laterais ionizáveis, capazes de doar ou
libertar protões (H+) durante a catálise. A histidina, a cisteína, a tirosina e os AA ácidos
são importantes aminoácidos nestes processos.
3.b. Torção do Substrato
Já citada anteriormente, depende da torção do substrato induzida pela ligação do
mesmo com o centro activo da enzima, alcançando o estado de transição e estimulando a
sua conversão em produto.
Enzimas página 17 de 25
Carlos Capela
3.c. Catálise Covalente
Resulta do ataque nucleofílico ou electrofílico de um radical do centro catalítico
sobre o substrato, ligando-o covalentemente à enzima e induzindo a sua transformação
em produto. Envolve com frequência a participação de coenzimas.
3.d. Efeitos de Diminuição da Entropia
As enzimas ajudam no posicionamento e na definição da estequiometria correcta da
reacção, facilitando os mecanismos anteriores.
Enzimas página 18 de 25
Carlos Capela
D. REGULAÇÃO DA ACTIVIDADE
ENZIMÁTICA
Algumas enzimas podem ter as suas actividades reguladas, actuando assim como
moduladoras do metabolismo celular. Esta modulação é essencial na coordenação de
inúmeros processos metabólicos pela célula.
Além dos mecanismos já citados de modulação de actividade enzimática - por
variação da concentração do substrato, ou por inibição enzimática, por exemplo existem 3 modelos de regulação enzimática mais comuns e conhecidos:
1.MODULAÇÃO ALOSTÉRICA
A regulação da actividade enzimática por ligação de ligandos a centros distintos do
centro activo – regulação alostérica – refere-se, em geral, à ligação de pequenas
moléculas designadas por moduladores ou efectores, produzindo-se, em consequência,
alterações conformacionais na molécula da proteína. Esta possui, em certos locais da sua
estrutura, centros específicos de ligação para os ligandos, designados por centros
alostéricos. Diz-se, então, que se trata de uma proteína alostérica.
Os centros alostéricos podem ser independentes entre si, ou exibirem
cooperatividade, isto é, a ocupação de um centro pelo ligando reflectir-se-á na afinidade
da ligação de outro ligando a um segundo centro. Se os ligandos intervenientes no
fenómeno de cooperatividade são moléculas iguais, diz-se que se trata de
cooperatividade homotrópica; se são diferentes, cooperatividade heterotrópica. A
cooperatividade pode, ainda ser positiva ou negativa, consoante a ligação do ligando
levar ao aumento ou diminuição da afinidade da proteína para posteriores ligações
noutros centros.
Na realidade, muitas enzimas são associações de subunidades (protómeros),
designando-se, então, a molécula de proteína por oligómero. Contudo, parece que esta
não é uma condição necessária para haver regulação alostérica, pois pelo menos duas
enzimas monoméricas com regulação alostérica são conhecidas: ribonucleósido difosfato
redutase e piruvato-UDP-N-acetilglicosamina transferase.
Enzimas página 19 de 25
Carlos Capela
1.a. Enzimas
sigmoidal
alostéricas
apresentam
cinética
Como uma consequência da interacção entre o sítio do substrato, sítio do activador e
sítio do inibidor, uma curva [S] versus v0 em forma de S ou sigmóidal é característico
nas enzimas alostéricas. Efectores alostéricos negativos deslocam a curva na direcção de
concentrações maiores de substrato, aumentando a forma sigmóide da curva, enquanto
efectores alostéricos positivos deslocam a curva na direcção de concentrações menores
de substrato, diminuindo a forma sigmóide da curva.
Como exemplo, temos a hemoglobina. Há cerca de 100 anos que se sabe que a curva
de saturação de hemoglobina, em função da pressão parcial de O2, é sigmóidal. A análise
deste comportamento gráfico sugere que o carácter sigmóidal possa ser resultado de uma
primeira ligação de O2 mais difícil, seguindo-se ligações progressivamente mais
facilitadas.
Como os grupos heme, aos quais se liga o O2, estão localizados em zonas distantes
da proteína, não se trataria de uma influência directa, mas de alterações de conformação
na molécula por ligação do ligando. Esta questão foi desenvolvida em dois modelos
matemáticos – os modelos de Hill (1910) e Adair (1925).
Enzimas página 20 de 25
Carlos Capela
Para descrever matematicamente as curvas de saturação de ligantes observadas
experimentalmente, muitos modelos de cooperatividade têm sido propostas. Os dois
mais proeminentes são o modelo de transição concertada (modelo Monod, Wyman e
Changeux – MWC – 1965) e o modelo sequencial (modelo Koshland, Nemethy e Filmer
– KNF – 1966).
1.b. Modelo de transição concertada
Embora o modelo de transição concertada seja restrito, a maior parte da
nomenclatura associada com o alosterismo e cooperatividade surgiu dele. O modelo
propõe que a enzima existe apenas em dois estados: o T (tense) e o R (relax). Os estados
T e R estão em equilíbrio. Activadores e substratos favorecem o estado R e alteram o
equilíbrio pré-existentes na direcção do estado R. Inibidores favorecem o estado T. Uma
alteração conformacional num protómero causa uma alteração correspondente em todos
os protómeros. Nenhum estado híbrido ocorre. Embora este modelo seja responsável
pelo comportamento cinético de muitas enzimas, não pode ser responsável pela
cooperatividade negativa.
1.c. Modelo sequencial
Este modelo propõe que a ligação do ligante induza uma alteração conformacional
num protómero. Uma alteração correspondente é, então, parcialmente induzida num
protómero adjacente ao protómero contendo o ligante unido. O efeito da união do ligante
é, sequencialmente, transmitido pelo oligómero, produzindo aumento ou diminuição da
afinidade do protómero pelo ligando. Neste modelo, numerosos estados híbridos
ocorrem, surgindo a cooperatividade e a curva [S] versus v0 sigmóide.
Enzimas página 21 de 25
Carlos Capela
Tanto a cooperatividade positiva como a negativa podem ser acomodadas por este
modelo. Um modulador positivo induz a conformação, no protómero, no que resulta a
afinidade aumentada pelo substrato. Um modulador negativo induz uma conformação
diferente no protómero, no que resulta a afinidade diminuída pelo substrato. Os dois
efeitos são transmitidos aos protómeros adjacentes.
Enzimas página 22 de 25
Carlos Capela
2.MODULAÇÃO COVALENTE
Ocorre quando há modificação covalente da molécula da enzima, com conversão
entre formas activa/inactiva. O processo ocorre principalmente por adição/remoção de
grupos fosfato de resíduos específicos de serina.
3.ACTIVAÇÃO PROTEOLÍTICA
Este tipo de activação é, ao contrário dos anteriores, irreversível. Algumas enzimas
passam à forma activa por hidrólise de uma ou mais ligações peptídicas num percursor
inactivado, designado por proenzima ou zimógeno.
Este mecanismo de activação gera as enzimas digestivas, como a pepsina, a tripsina
e a quimiotripsina. O mecanismo de coagulação sanguínea pode ser descrito como uma
cascata de activação de zimogénos, na qual uma forma activada de um factor de
coagulação catalisa a activação do percursor seguinte.
Enzimas página 23 de 25
Carlos Capela
E. ENZIMAS NA CLÍNICA
As enzimas podem ser utilizadas nas Análises Clínicas de 2 formas principais:
•
Como reagentes altamente específicos e sensíveis em reacções colorimétricas
quantitativas;
•
Como indicadoras de lesão celular e tecidual – a evasão de enzimas do meio intra
para o meio extracelular leva a um aumento da actividade destas no sangue. Esta
actividade pode ser medida e fornece importante informação diagnóstica e de
evolução de um quadro clínico. A distribuição órgão-específica de algumas
destas enzimas permite a localização da lesão com bastante precisão.
Exemplos
de
doenças
que
podem
ser
diagnosticadas
e
acompanhadas
enzimaticamente são:
•
O Enfarto Agudo do Miocárdio;
•
As Hepatites;
•
A Pancreatite;
•
As Colestases;
•
Doenças Ósseas, etc.
1.AS ISOENZIMAS
Isoenzimas são enzimas que catalisam a mesma reacção, mas migram diferentemente
em electroforese.
Isoenzimas que têm ampla aplicação clínica são a lactato desidrogenase, a creatina
fosfocinase e a fosfatase ácida.
1.a. Creatina fosfocinase (CPK)
A CPK ocorre como um dímero com dois tipos de subunidades: M (tipo “Muscle”) e
B (tipo “Brain”). No cérebro, as duas subunidades são electroforecticamente iguais e são
designadas por B. No músculo esquelético, as duas subunidades são do tipo M. A
isoenzima contendo tanto a subunidade M como a B (MB) é encontrada apenas no
Enzimas página 24 de 25
Carlos Capela
miocárdio. Outros tecidos contêm quantidades variáveis de isoenzimas MM e BB. As
isoenzimas são numeradas a partir das espécies que migram mais rápido para o ânodo da
electroforese – portanto, CPK1 (BB), CPK2 (MB), CPK3 (MM).
1.b. Lactato desidrogenase (LDH)
É uma enzima tetramérica contendo apenas duas subunidades diferentes: H (“Heart”)
e M (“Muscle”). Essas duas subunidades são combinadas de cinco modos diferentes.
Tipo
Composição
Localização
LDH1
HHHH
Miocárdio
LDH2
HHHM
Miocárdio
LDH3
HHMM
Cérebro e rim
LDH4
HMMM
−
LDH5
MMMM
Fígado e músculo esquelético
O método da electroforese para determinação das enzimas cardíacas num ataque
cardíaco, é lento demais e insensível para ser de valor numa situação de emergência.
Ensaios de ELISA baseados em anticorpos monoclonais contra CPK2 são rápidos (30
minutos) e sensíveis o suficiente para detectarem CPK2 no soro, cerca de uma hora após
o ataque cardíaco.
Enzimas página 25 de 25