2-1
2. C O N C E I T O S D E B I O Q U Í M I C A E
QUÍMICA ORGÂNICA
Para estudos com moléculas orgânicas, principalmente aquelas com
atividade biológica, é necessário o conhecimento de alguns conceitos básicos
químicos e bioquímicos. Neste capítulo iremos apresentar alguns desses
conceitos que serão importantes para a compreensão do nosso trabalho.
2.1. Grupos funcionais
Para o estudo de biomoléculas, é importante conhecer bem as funções
químicas e os grupos funcionais que as determinam. Grupos funcionais são
grupamentos de átomos que conferem aos compostos orgânicos características
químicas específicas, criando um centro reativo e definindo uma função
química nessas moléculas. Portanto cada função química possui um conjunto
de
características
comuns
e
seu
conhecimento
compreensão de como essas moléculas interagem.
é
fundamental
para
a
CONCEITOS
2-2
DE
BIOQUÍMICA
E
QUÍMICA ORGÂNICA
I r e m o s a p r e s e n t a r n e s t a s e ç ã o , d e f o r m a s u c i n t a 1, a l g u n s g r u p a m e n t o s
químico-orgânicos que serão relevantes para nosso estudo. As figuras que
seguem mostram exclusivamente o grupo funcional, onde nos traços de
ligações podem ser conectadas cadeias carbônicas ou átomos de hidrogênio,
conforme o caso.
2.1.1.
Hidrocarboneto
Compostos formados exclusivamente por átomos de carbono e hidrogênio
são conhecidos por hidrocarbonetos. Estes compostos são a base para a
formação de todas as outras substâncias orgânicas.
Os hidrocarbonetos se classificam, quanto ao grau de saturação, em
saturados (alcanos), em que os átomos de carbono fazem apenas ligações
simples, e insaturados, onde um ou mais átomos de carbono podem fazer
ligações duplas (alcenos) ou triplas (alcinos). Os radicais correspondentes a
cada categoria possuem na sua nomenclatura o sufixo –ila, formando assim
grupamentos alcanila, alcenila e alcinila. Podem também ser classificados
quanto ao tipo de cadeia, podendo ser de cadeia normal ou cíclica (formando
anéis estáveis).
C
C
(a)
C
C
(b)
C
(c)
Figura 2.1:Grupamento de (a) alcanila, (b) alcenila e (c) alcinila
2.1.2.
Hidroxila
O grupamento hidroxila é formado por um átomo de oxigênio ligado a um
átomo de hidrogênio. Este grupamento define a função álcool. Os álcoois são
1
Para
informações
adicionais
e
mais
detalhadas
funcionais, consultar as referências [25] e [26]
sobre
química
orgânica
e
grupos
GRUPOS
FUNCIONAIS
2-3
classificados em primários, secundários ou terciários dependendo se o carbono
conectado à hidroxila está ligado a um, dois ou três átomos de carbono.
OH
Figura 2.2: Grupamento hidroxila
2.1.3.
Carbonila
A carbonila é formada por um átomo de oxigênio que realiza uma ligação
dupla com um átomo de carbono. Esse grupamento pode definir as funções
aldeído (se localizado nas extremidades) ou cetona (se localizado no meio)
dependendo somente de sua localização na cadeia carbônica,.
O
C
Figura 2.3: Grupamento carbonila
2.1.4.
Carboxila
A carboxila é uma fusão entre uma carbonila e uma hidroxila. A função
definida por este grupamento é o ácido carboxílico. Esse nome é dado porque a
ligação entre o átomo de oxigênio e o átomo de hidrogênio na carboxila é
extremamente polar, dando um grande caráter ácido ao hidrogênio. Isso faz
c o m q u e q u a n d o c o l o c a d a s e m á g u a , s o b c e r t a s c o n d i ç õ e s d e p H 1, a l g u m a s
substâncias carboxiladas podem se ionizar doando prótons (H+) para o meio e
se tornando negativas.
O
C
OH
Figura 2.4: Grupamento carboxila
1
pH é defindo como sendo −log10[H+]
CONCEITOS
2-4
2.1.5.
DE
BIOQUÍMICA
E
QUÍMICA ORGÂNICA
Éster
O grupo funcional éster é um derivado da carboxila onde o hidrogênio da
hidroxila é substituído por um radical carbônico. Este grupamento define a
função química éster
A reação de formação dos ésteres é uma das mais freqüentes em sistemas
biológicos. A esterificação consiste na reação de ácido carboxílico com álcool
e resulta na produção de um éster e de uma molécula de água.
A ligação simples entre o átomo de carbono e o oxigênio é conhecida
como ester linkage (ligação éster).
O
C
O
Figura 2.5: Grupamento éster
2.1.6.
Amino
Amina é a função química definida pelo grupamento amino. Este é
constituído principalmente pela presença do átomo de nitrogênio efetuando
ligações simples.
A classificação das aminas depende de quantos átomos de carbono estão
ligados diretamente ao nitrogênio, podendo ser primárias (um carbono),
secundárias (dois carbonos) ou terciárias (três carbonos).
O grupo amino possui caráter básico e em solução aquosa este grupo
pode captar prótons (H+) do meio e tornar-se positivo. Como ocorre com a
carboxila, isso acontece sob condições específicas de pH.
N
Figura 2.6: Grupamento amino
GRUPOS
2.1.7.
2-5
FUNCIONAIS
Amida
O grupamento amida é um derivado da carboxila em que a hidroxila é
substituída por um grupo amino. A função química amida é defina pelos grupos
amida.
A ligação entre o carbono e o nitrogênio é extremamente estável e é
denominada amida linkage (ligação amida). Ela é formada quando há a reação
química entre ácido carboxílico e amina, produzindo uma amida e uma
molécula
de
repetidamente
água.
unindo
Esse
tipo
pequenas
de
ligação
moléculas
é
normalmente
(meros),
dando
encontrada
origem
a
macromoléculas poliméricas. As proteínas são um dos principais exemplos de
polímeros onde encontramos esse tipo de ligação. Pequenos aminoácidos se
unem formando peptídeos que se por sua vez unem formando polipeptídios e
proteínas. Devido a esse fato, em bioquímica, esse tipo de ligação amida é
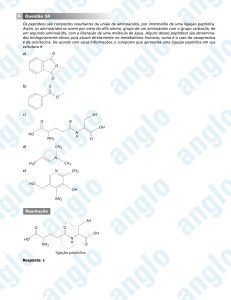
mais comumente conhecido como ligação peptídica.
O
C
N
Figura 2.7: Grupamento amida
2.1.8.
Éter
O grupamento éter é definido quando o átomo de oxigênio possui dois
ligantes carbônicos. Uma conseqüência interessante desse fato é que não
ocorre a formação de ligações de hidrogênio nesse oxigênio [25]. A função
química éter é determinada por este grupamento e seus compostos são pouco
reativos.
O
Figura 2.8: Grupamento éter
CONCEITOS
2-6
2.1.9.
DE
BIOQUÍMICA
E
QUÍMICA ORGÂNICA
Epóxido
O grupamento epóxido é um éter cíclico. Os epóxidos, ou oxiranos, são,
diferentemente dos éteres, bastante reativos, principalmente devido à forte
tensão no anel.
O
C
C
Figura 2.9: Grupamento epóxido
2.1.10. Imino
O átomo de nitrogênio fazendo uma dupla ligação é o que caracteriza o
grupamento imino. Esse grupo define a função química imina, que é um
derivado de compostos que contém carbonila.
C
N
Figura 2.10: Grupamento imino
2.1.11. Sulfidrila
A sulfidrila é caracterizada por um átomo de enxofre ligado a um átomo
de hidrogênio. A função química determinada por esse grupo é a função tiol.
Existe certa semelhança na química dos tióis (também conhecidos como
mercaptanas) e dos álcoois, já que o enxofre e o oxigênio pertencem ao sexto
grupo da tabela periódica e portanto possuem a mesma configuração eletrônica
da camada externa. Entretanto, há também inúmeras diferenças. Dentre elas
podemos citar a formação estável de compostos sulfurados conhecidos como
dissulfetos (R−S−S−R), o que geralmente não ocorre com os peróxidos
(R−O−O−R) pois a ligação entre os átomos de oxigênio é consideravelmente
GRUPOS
2-7
FUNCIONAIS
mais fraca que a ligação entre os átomos de enxofre. Essa estabilidade dos
dissulfetos é um fator muito importante na estrutura de proteínas, podendo
existir
a
formação
de
pontes
dissulfeto
entre
aminoácidos
sulfurados
(cisteínas).
SH
Figura 2.11: Grupamento sulfidrila
2.2. Enzimas: estrutura
Enzimas são proteínas que tem a capacidade de catalisar reações
metabólicas dos seres vivos e por isso são essenciais para a vida. Nesta seção
vamos abordar a constituição e a estrutura das proteínas em geral. Na seção
2.3 iremos discutir a questão de cinética enzimática.
2.2.1.
Aminoácidos e peptídios
Aminoácidos são pequenas moléculas que tem certas características
especiais, dentre as quais a capacidade de polimerização. Isso se deve ao fato
de que essas moléculas possuem nas extremidades dois grupos funcionais: um
grupamento amino (−NH2) e uma carboxila (−COOH), que, como vimos na
seção 2.1.7, podem reagir formando uma ligação C−N do tipo amida e que
nesse caso também é chamada de ligação peptídica. Um peptídeo é a união de
alguns poucos aminoácidos e a união destes formam os polipeptídios e
proteínas. Existem também aminoácidos e derivados que não são constituintes
das proteínas e possuem alguma outra função biológica. Como exemplos
podemos citar alguns antibióticos e hormônios.
A estrutura geral de um aminoácido pode ser vista na Figura 2.12. Existe
um carbono central conhecido como carbono alfa (Cα). Ligados a este carbono
existem um hidrogênio, os grupamentos amínico e carboxílico e uma cadeia
lateral “R”, que diferencia cada aminoácido e confere a eles identidades e
características distintas. A Figura 2.13 mostra os vinte tipos de aminoácidos
usualmente encontrados em proteínas. O carbono central normalmente tem
CONCEITOS
2-8
BIOQUÍMICA
DE
E
QUÍMICA ORGÂNICA
quatro ligantes distintos e portanto também possui atividade ótica. Os
aminoácidos biologicamente ativos possuem configuração ótica levógira (Laminoácidos)
H
C
(a)
H2N
H
R
α
H3N
COOH
R
α
+
C
(b)
CO O
Figura 2.12: Estrutura geral de um aminoácido: (a) em vácuo e (b) zwiteriônico
Apolares
O
O
H3C
H2
C
H
C
C
H
C
CH3
H2
C
OH
H2C
Leucina (Leu)
Prolina (Pro)
CH3
O
H 3C
C
H
OH
C
H
C
H3C
Valine (Val)
H
C
O
H2
C
C
H2
H
C
C
OH
CH
Fenilalanina (Phe)
C
NH2
CH
O
H2
C
C
H
H
C
HC
Triptofano (Trp)
H
C
HC
H2
C
N
H
Metionina (Met)
HC
O
H
C
C
H
NH2
H
C
OH
C
NH2
Alanina (Ala)
S
O
H
C
NH2
H3C
OH
NH
C
H2
NH2
C
H
C
NH2
C
CH3
OH
H3C
C
H2
C
H
O
H
C
C
NH2
Isoleucina (Ile)
Figura 2.13: Os vinte tipos de aminoácidos encontrados em proteínas
OH
OH
ENZIMAS:
2-9
ESTRUTURA
Polares
O
O
H
C
H
C
H2
C
OH
HO
NH2
O
O
H2
C
C
H
C
H2
C
H2N
OH
C
H2
NH2
NH2
OH
O
O
OH
C
H
C
H2
C
HS
H
C
OH
C
NH2
NH2
Treonina (Thr)
Cisteína (Cys)
O
O
H2
C
H
C
HC
C
OH
H2
C
N
H
C
C
OH
HC
CH
HO
C
Glutamina (Gln)
OH
H
C
H
C
NH2
Asparagina (Asn)
C
H
OH
Serina (Ser)
O
H3C
C
NH2
Glicina (Gly)
O
H
C
NH2
C
H
N
H
Tirosina (Tyr)
CH
NH2
Histidina (His)
Básicos
NH
H2N
H2
C
C
H2
O
H2
C
C
H2
H
C
C
H2N
OH
N
H
H2
C
O
H2
C
C
H2
NH2
H
C
NH2
Lisina (Lys)
Arginina (Arg)
C
OH
2-10
CONCEITOS
DE
BIOQUÍMICA
E
QUÍMICA ORGÂNICA
Ácidos
O
O
H2
C
O
OH
O
H
C
C
HO
OH
C
H2
NH2
Ácido Aspártico (Asp)
H2
C
H
C
C
OH
NH2
Ácido Glutâmico (Glu)
Na seção anterior vimos que tanto os grupamentos amínicos como os
carboxílicos podem estar carregados quando em solução aquosa, ou seja, o
grupo amino na forma −NH3+ e a carboxila na forma −COO−. Um aminoácido
isolado possui ambos grupamentos e sendo assim, sob certas condições de pH,
pode apresentar separação de cargas com neutralidade elétrica. Essa forma é
conhecida como forma zwiteriônica (Figura 2.12 (b)). Também pode acontecer
da molécula, mesmo em solução aquosa, permanecer neutra sem separação de
cargas e portanto estar na configuração encontrada em vácuo (Figura 2.12 (a)).
Os aminoácidos possuem então pelo menos dois hidrogênios que podem
se dissociar, podendo haver mais hidrogênios dissociáveis nas cadeias laterais
(grupo R). Com isso estas moléculas podem assumir diversos estados de carga
dependendo do ambiente químico em que se encontram.
Podemos analisar o que ocorre numa solução aquosa de um aminoácido
que não possui hidrogênios dissociáveis nas cadeias laterais. Existem então
apenas dois hidrogênios que podem se dissociar da molécula, localizados nos
terminais amínico e carboxílico. Inicialmente, o pH da solução é bem baixo. O
meio está ácido e ambos grupamentos estão protonados (−COOH e −NH3+).
Nesse estado portanto a molécula possui carga total positiva. À medida que se
eleva o pH da solução (com a adição de hidróxido, por exemplo), percebemos a
dissociação do hidrogênio da carboxila. A molécula de aminoácido resultante
dessa reação é neutra, porém apresentando separação de cargas (forma
zwiteriônica), já que agora a carboxila se encontra na forma −COO− e o grupo
amino na forma −NH3+. Continuando a elevar o pH da solução, temos também a
dissociação do hidrogênio do grupamento amínico, que então se altera para a
ENZIMAS:
2-11
ESTRUTURA
forma neutra −NH2. A molécula agora possui uma carga global negativa. Esses
três estados de carga estão esquematizados na Figura 2.14.
H
COOH
+
H3N
C
H
+
H
CO O
+
H 3N
R
(a)
C
+
H
CO O
H 2N
R
(b)
C
H
R
(c)
Figura 2.14: Os três possíveis estados de carga de um aminoácido sem ionização de
cadeias laterais: (a) forma catiônica, (b) zwiteriônica e (c) aniônica
Podemos escrever essas reações e suas constantes, denotando por AA+,
AA0 e AA− os três estados de carga que o aminoácido pode assumir.
A reação referente à primeira dissociação (carboxila) é
→
+
0
AA + ←
AA + H
e a sua constante K1 é escrita como
K 1=
[ AA 0 ][ H + ]
[ AA + ]
(2.1)
A segunda reação, referente à outra dissociação (grupo amino), é dada
pela expressão
→
−
+
AA 0 ←
AA + H
e a constante K2 pode ser escrita como
K 2=
[ AA − ][ H + ]
[ AA 0 ]
(2.2)
CONCEITOS
2-12
DE
BIOQUÍMICA
E
QUÍMICA ORGÂNICA
É possível obter experimentalmente os valores das constantes de reação1
K1 e K2 e determinar assim os pKa2 dos aminoácidos. Da mesma forma também
podemos determinar as constantes de dissociação de átomos de hidrogênio
presentes em cadeias laterais (R) e por conseguinte o pKa correspondente. Os
valores de pKa dos aminoácidos mais comuns estão mostrados na Tabela 2.1.
Tabela 2.1: Valores de pKa dos aminoácidos mais comuns [27]
pKa
Aminoácido
−COOH
−NH3+
grupo R
Ácido Aspártico
2.1
2.2
2.4
2.2
2.0
1.7
1.8
2.2
2.3
1.8
2.4
2.4
2.2
2.3
2.1
2.2
2.6
2.4
2.2
2.3
9.8
9.7
9.7
9.0
8.8
10.8
9.1
9.1
9.6
9.2
9.7
9.6
9.0
9.2
10.6
9.2
10.4
9.4
9.1
9.6
3.9
4.3
12.5
8.3
6.0
10.5
≈13
≈13
10.1
-
Ácido Glutâmico
Alanina
Arginina
Asparagina
Cisteína
Fenilalanina
Glutamina
Glicina
Histidina
Isoleucina
Leucina
Lisina
Metionina
Prolina
Serina
Treonina
Triptofano
Tirosina
Valina
Como podemos verificar, os aminoácidos que não possuem cadeias
laterais
com
hidrogênio
dissociável
estão
em
grande
parte
na
forma
zwiteriônica em solução aquosa a pH fisiológico (pH 7). Contudo, é bom
lembrar que mesmo nessas condições, pode ocorrer que algumas moléculas
1
Através da construção de uma curva de titulação
2
pKa é definido como sendo −log10Ka
ENZIMAS:
2-13
ESTRUTURA
permaneçam na configuração de vácuo. Existem diversos estudos que discutem
esse tipo de comportamento nos aminoácidos e proteínas [9] [28] [29] [30].
2.2.2.
Proteínas
Proteínas são moléculas compostas por um ou mais polipeptídios. Estes
são polímeros lineares (não ramificados) de aminoácidos unidos por ligações
peptídicas. As proteínas são sintetizadas dentro das células de um modo bem
engenhoso. De maneira bem resumida, podemos descrever como é esse
p r o c e s s o 1. P r i m e i r a m e n t e h á a l e i t u r a d e u m s e g m e n t o d e D N A , m o l é c u l a o n d e
ficam
codificadas
todas
as
i n f o r ma ç õ e s
genéticas
dos
seres
vivos,
e
estabelecida a seqüência de bases de nucleotídeos do gene a ser expresso. Toda
a informação é armazenada num tipo específico de molécula mensageira
chamada mRNA. Essa fase de leitura do DNA e armazenamento de informações
é conhecida como transcrição. Acontece então a fase de leitura da fita de
mRNA e o início da síntese de polipeptídios. Em seres mais complexos, existe
uma fase de preparação da molécula de mRNA antes que esta seja lida. Cada
conjunto de três bases nitrogenadas do mRNA é capaz de identificar um
aminoácido específico. À medida que ocorre essa leitura, uma molécula
especial de transporte de aminoácidos conhecida por tRNA identifica esse
conjunto de três bases e transporta o aminoácido correspondente, conectando-o
através de uma ligação peptídica ao restante da cadeia. Essa fase de construção
dos peptídeos é chamada de tradução.
A conformação final de uma proteína é algo bem específico. Fazer uma
previsão dessa estrutura com base apenas na seqüência de aminoácidos é ainda
um problema muito complexo [32]. Uma proteína pode conter centenas e
centenas de resíduos por cadeia polipeptídica, o que torna realmente muito
difícil prever a conformação biologicamente ativa.
Vamos olhar algumas características da ligação peptídica. O termo
“backbone” de uma proteína consiste na repetição da seqüência −N−Cα−C−,
onde N é o nitrogênio da amida, Cα é o carbono central dos aminoácidos e C é
1
Para maiores informações sobre síntese protéica, consultar as referências [27] e [31]
CONCEITOS
2-14
DE
BIOQUÍMICA
E
QUÍMICA ORGÂNICA
o carbono da carbonila que está conectado ao N da amida do próximo
aminoácido. Em princípio, no backbone, temos três ligações simples que
possuem rotação livre, N−Cα, Cα−C e C−N (Figura 2.15 (a)), observando que
existe uma ligação dupla na carbonila (C=O). Essa representação não está de
acordo com os fatos pois é sabido que não há rotação em torno da ligação
C−N. Existe uma forma ressonante em que a ligação entre C e N possui caráter
de ligação dupla (C=N), deixando um elétron livre no oxigênio (Figura 2.15
(b)) que passa a fazer uma ligação simples com o carbono (C−O). Desse modo
a ligação C=N não teria a possibilidade de rotacionar. Isso também não
acontece de fato pois existiria uma separação de cargas muito grande nessa
região. O que ocorre é um estado intermediário entre as duas formas
ressonantes mencionadas acima (Figura 2.15 (c)) e isso traz conseqüências
importantes no caráter estrutural.
α
C
H
C
O
(a)
α
C
H
C
N
α
C
N
(b)
O
α
C
+
α
C
δ
δ+
N
C
(c)
O
H
α
C
Figura 2.15: Estrutura eletrônica do backbone: ((a) e (b)) formas ressonantes e (c)
forma intermediária
Para tentar facilitar o entendimento do problema conformacional, são
definidos alguns níveis de estrutura que são mostrados abaixo. Estes níveis
dividem em categorias os tipos de organização estrutural das proteínas.
•
Estrutura Primária: É a seqüência de aminoácidos de uma proteína. Por
exemplo: −Lys−Glu−Thr−Ala−Ala−Ala−Lys−Phe−;
•
Estrutura Secundária: Os aminoácidos adjacentes podem interagir e se
conectar por ligações de hidrogênio, formando nas fitas de peptídeos um
certo tipo bem característico de estrutura. Esses padrões são conhecidos
por estruturas regulares e o conjunto dessas estruturas é a estrutura
secundária de uma proteína. Os dois exemplos mais comum de estruturas
regulares são a α-hélice e a folha β, que podem ser vistos em destaque
na Figura 2.16 (b);
ENZIMAS:
•
2-15
ESTRUTURA
Estrutura Terciária: A interação entre as estruturas regulares das cadeias
polipeptídicas leva uma estrutura tridimensional mais bem definida. Esta
estrutura é conhecida como estrutura terciária. Basicamente existem dois
tipos de forma tridimensional que as proteínas podem assumir. As
proteínas fibrosas, que tem um papel estrutural nas células, possuem as
cadeias polipeptídicas organizadas paralelamente ao longo de um eixo,
produzindo tanto longas fibras como largas folhas. Já as proteínas
globulares, que são a grande maioria e desempenham diversas funções,
são aquelas que se empacotam e tem formas aproximadamente esféricas.
Essa forma pode ser entendida já que as proteínas globulares existem na
maioria dos casos em solução aquosa. Exemplos de estrutura terciária de
proteínas fibrosas e globulares podem ser vistos na Figura 2.16;
•
Estrutura Quaternária: Existem muitas proteínas que são constituídas por
duas ou mais cadeias polipeptídicas, cadeias essas que já possuem uma
estrutura terciária definida. A interação entre essas cadeias, também
conhecidas como subunidades, é a estrutura quaternária.
(a)
(b)
Figura 2.16: Exemplo de (a) proteína fibrosa (fibra de colágeno [33]) e (b) globular
(flavodoxina [34]). A tonalidade indica o tipo de estrutura secundária.
CONCEITOS
2-16
DE
BIOQUÍMICA
E
QUÍMICA ORGÂNICA
As interações envolvidas na conformação final das proteínas são de
origem não covalente. Apenas a estrutura primária é definida pela ligação
covalente entre os aminoácidos. As forças envolvidas na organização das
demais
estruturas
são
ligações
de
hidrogênio,
interações
hidrofóbicas,
interações eletrostáticas e forças de van der Waals. Existem contudo alguns
tipos de proteínas que estabilizam a estrutura terciária formando pontes
dissulfeto entre os resíduos de cisteína, como é citado na seção 2.1.11.
As proteínas possuem inúmeras funções diferentes, mas a grande maioria
delas tem a função enzimática.
2.2.3.
Enzimas
Como vimos, enzimas são uma classe de proteínas que desempenham um
papel
fundamental
no
metabolismo.
Elas
possuem
capacidade
catalítica
podendo acelerar muito a velocidade das reações bioquímicas e por isso são
catalisadores biológicos. Em uma reação desse tipo, os reagentes, também
conhecidos por substratos, interagem com a enzima transformando-se em
produtos e assim liberando o catalisador para outra interação. Esta interação
do substrato com a enzima ocorre numa região bem específica, conhecida como
sítio ativo.
O sítio ativo é uma porção relativamente pequena da enzima. Ele tem
uma estrutura bem determinada que é normalmente um pouco mais rígida que o
restante da molécula. Essa estrutura é uma característica fundamental para
determinar o reconhecimento mútuo entre enzima e substratos. Podemos fazer
uma analogia com um sistema de chave e fechadura, em que para funcionar
corretamente deve existir compatibilidade.
As enzimas se classificam essencialmente quanto ao tipo de reação que
catalisam. Existem seis classes principais que depois se dividem em muitas
outras.
Uma enzima pode ser caracterizada basicamente por três fatores: poder
catalítico, regulação e especificidade.
ENZIMAS:
•
2-17
ESTRUTURA
Poder catalítico: é a relação entre as constantes de velocidade da reação
catalisada e da reação não catalisada. Valores do poder catalítico podem
ser números extremamente grandes, indicando que as enzimas podem
amplificar muito as a velocidades das reações.
•
Regulação:
é
o
conjunto
dos
diversos
fatores
que
afetam
o
funcionamento enzimático. Esses fatores são muitos e variam desde a
quantidade enzimática produzida até a interação das enzimas com
moléculas ativadoras ou inibidoras de atividade. Dois fatores muito
importantes na atividade são o pH do meio e a temperatura e serão
discutidos a seguir.
•
Especificidade: representa a seletividade de uma enzima tanto em
relação aos substratos com os quais ela interage quanto em relação a
reação
que
ela
catalisa.
A
estrutura
do
sítio
ativo
é
uma
peça
fundamental na especificidade.
2.3. Enzimas: cinética
Mesmo que uma reação química seja energeticamente favorável, ela pode
não ocorrer ou ser extremamente lenta devido a uma alta barreira de energia de
ativação, sendo assim cineticamente proibida. As enzimas diminuem essa
barreira de energia fazendo com que essas reações se tornem viáveis e muitas
vezes mais rápidas (Figura 2.17).
2.3.1.
Cinética química
Antes de começarmos o estudo de cinética de reações catalisadas por
enzimas, é interessante analisar resumidamente os conceitos básicos de
cinética química geral [35].
Consideremos a reação elementar e irreversível
R
→ P
Podemos expressar a velocidade v dessa reação como a quantidade de
produtos (P) formados ou a quantidade de reagentes (R) consumidos por
2-18
CONCEITOS
DE
BIOQUÍMICA
E
QUÍMICA ORGÂNICA
unidade de tempo e relacionar a velocidade v com a concentração. Essa relação
é determinada experimentalmente e é conhecida como lei de velocidade
(Equação (2.3)). A constante de proporcionalidade k que conecta os dois
membros da equação é chamada de constante de velocidade.
v=
d [ P]
d [R ]
=−
= k[R ]
dt
dt
(2.3)
A ordem de um reagente numa reação é definida como o seu expoente na
lei de velocidade. A ordem global da reação é dada pela soma das ordens de
cada regente envolvido. Um outro conceito é o de molecularidade, que está
relacionado com o número de moléculas de reagente que interagem entre si
durante o processo. No exemplo acima temos uma reação unimolecular de
primeira ordem.
Podemos resolver (2.3) facilmente para este exemplo. A solução da
equação é dada por
[ R ] = [ R ]0 e − kt
(2.4)
com [R]0 sendo a concentração de R no instante t=0.
Olhando para a equação (2.4), vemos que a concentração de R decai
exponencialmente com o tempo a partir do valor inicial [R]0. Essa expressão
pode ser linearizada, tomando a forma
ln[ R ] = ln[ R ]0 − kt
(2.5)
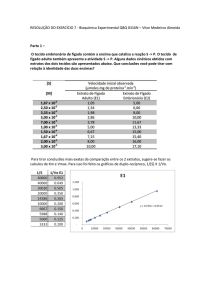
Fazendo medidas de concentração ao logo do tempo, é possível traçar
uma reta utilizando a expressão (2.5) e obter o valor experimental de k.
Podemos construir um diagrama de energia livre para essa reação. O
estado inicial é dado pela energia livre média dos reagentes e o estado final é
definido pela energia livre média dos produtos. Para ocorrer a reação, parte
das moléculas de reagente adquire uma certa quantidade de energia tornandose reativas e atingindo um estado intermediário conhecido como estado de
transição ou complexo ativado. Esse estado corresponde ao máximo da curva
de energia. O diagrama de energia livre é mostrado na Figura 2.17. Quando a
molécula está no estado de transição, ela pode ou decair para formar produtos
ENZIMAS:
CINÉTICA
2-19
ou relaxar de volta para reagentes. A altura do pico de energia, ou seja, a
diferença de energia entre o estado de transição e os reagentes, é conhecida
como energia livre de ativação.
Figura 2.17: Diagrama de energia livre
Os catalisadores são substâncias que participam do processo de reação
alterando seu mecanismo para que haja uma redução na altura da barreira de
energia de ativação (Figura 2.17). Com isso temos um aumento significativo na
velocidade da reação. Apesar de participarem do processo, os catalisadores não
são consumidos, sendo então regenerados após cada ciclo.
2.3.2.
Cinética de reações enzimáticas
Vamos agora analisar a cinética de reações catalisadas por enzimas. Esse
tipo de reação possui uma cinética muito particular. No caso anterior, a lei de
velocidade da reação era uma reta. No caso da cinética enzimática, a lei de
velocidade é mais complexa e sua expressão ainda não é bem conhecida.
Contudo, o comportamento experimental da curva é bem estabelecido.
CONCEITOS
2-20
DE
BIOQUÍMICA
E
QUÍMICA ORGÂNICA
Uma lei de velocidade para reações enzimáticas foi proposta por L.
Michaelis e M. L. Menten [36]. Esta lei era consistente com os resultados
experimentais e era baseada na formação de um complexo ativado entre enzima
e substrato.
k1
→
k2
E + S ← ES →
E+P
k−1
Para se chegar à equação de Michaelis-Menten, é necessário assumir
duas
condições.
A
primeira
é
a
hipótese
de
estado
estacionário.
Essa
proposição assume que a formação e a dissociação do complexo ativado ES
possuem velocidades equivalentes, de forma que
d [ ES]
=0
dt
(2.6)
A segunda é a hipótese de velocidade inicial. Esta simplificação ignora a
reação de formação do complexo ativado ES a partir de E+P. Isso se justifica
já que no início, a concentração de P é muito pequena e a reação de volta é
proporcional a esta concentração.
É interessante definir a constante Km, que é conhecida por constante de
Michaelis. Ela é definida como
Km =
( k −1 + k 2)
k1
(2.7)
Utilizando os conceitos acima, é muito simples chegar a equação de
Michaelis-Menten [27], que é dada pela expressão
v=
Vmax [S]
K m + [S]
(2.8)
em que Vmax é definido como sendo
Vmax = k 2 ([ E] + [ ES]) = k 2 [ E total ]
(2.9)
Vmax pode ser interpretada como a velocidade máxima que uma reação
enzimática pode atingir. Isso acontece quando a concentração de substrato é
grande o suficiente para saturar todas as moléculas de enzimas do sistema.
ENZIMAS:
2-21
CINÉTICA
Podemos interpretar a constante Km de uma forma mais operacional. Ela
pode ser entendida como a concentração de substrato [S] em que a velocidade
da reação é a metade da velocidade máxima, ou seja quando
Vmax
2
(2.10)
[S] = K m
(2.11)
v=
então
Isso pode ser verificado substituindo (2.10) na equação (2.8).
A curva descrita pela equação (2.8) é uma hipérbole e se ajusta muito
bem
aos
resultados
experimentais.
No
entanto,
para
se
determinar
os
parâmetros experimentais Km e Vmax, é interessante realizar a linearização
dessa equação. Existem diversas formas de se efetuar essa operação. Uma
dessas maneiras é a linearização de Lineweaver-Burk [37] (Figura 2.18). Sua
expressão pode ser facilmente obtida tomando o inverso dos dois membros de
(2.8)
1 K m + [S] K m 1
1
=
=
+
v Vmax [S] Vmax [S] Vmax
Figura 2.18: Gráfico de Lineweaver-Burk
(2.12)
CONCEITOS
2-22
DE
BIOQUÍMICA
E
QUÍMICA ORGÂNICA
Também é usual definir outra constante, kcat, chamada de número de
turnover. Essa constante representa uma medida da atividade catalítica máxima
de uma enzima e por isso também é chamada de atividade molecular. Ela é
definida como sendo o número de moléculas de substrato convertidas em
produtos por molécula de enzima por unidade de tempo quando a enzima está
saturada com substrato. Ela pode ser facilmente obtida no caso em que a
equação de Michaelis-Menten é valida. Quando temos saturação de substrato,
pelas equações (2.8) e (2.9), temos
(2.13)
v = Vmax = k 2 [ E total ]
k2 =
Vmax
= kcat
[ E total ]
(2.14)
Contudo, sob condições fisiológicas, as concentrações de substrato são
muito baixas e dificilmente existe a saturação da enzima. Para se obter uma
medida da atividade catalítica enzimática nessas condições, é utilizada a razão
k cat
Km
Existem inúmeros fatores que podem alterar a velocidade de uma reação
enzimática. Como visto anteriormente, o conjunto desses fatores formam a
regulação da enzima. Dois fatores muito importantes que alteram a atividade
são o pH e a temperatura.
A
atividade
enzimática
e
o
reconhecimento
de
substratos
são
extremamente dependentes do pH do meio na maioria dos casos. Como já
vimos, os aminoácidos são sensíveis a variações de pH e isso é determinante
na
estrutura
final
das
proteínas.
Qualquer
modificação
estrutural,
principalmente no sítio ativo pode alterar o funcionamento das enzimas. Outra
questão é que até mesmo os substratos podem possuir grupos ionizáveis e uma
forma iônica pode ter preferência sobre a outra na interação com a enzima. Em
geral as enzimas possuem atividade somente em um intervalo específico de pH,
possuindo ainda um pH ideal onde sua atividade é máxima (Figura 2.19).
ENZIMAS:
2-23
CINÉTICA
Figura 2.19: Efeito do pH na atividade enzimática [27]
A temperatura possui dois efeitos principais sobre as reações catalisadas
por enzimas. O primeiro é o aumento natural da velocidade das reações com a
e l e v a ç ã o d a t e m p e r a t u r a . É e m p i r i c a me n t e c o n h e c i d o q u e c a d a a u m e n t o d e
10ºC na temperatura leva a uma duplicação da velocidade da reação [35]. O
segundo efeito é que quando é atingida uma determinada temperatura, a
proteína se desnatura, ou seja, perde sua estrutura nativa e isso resulta em um
decréscimo de atividade. A comparação entre a estrutura nativa e a estrutura
desnaturada de uma proteína é ilustrada na Figura 2.20. Os efeitos da
temperatura na atividade enzimática podem ser vistos na Figura 2.21.
(a)
(b)
Figura 2.20: Formas (a) nativa e (b) desnaturada de uma proteína [38]
CONCEITOS
2-24
DE
BIOQUÍMICA
E
QUÍMICA ORGÂNICA
Figura 2.21: Efeito da temperatura na atividade enzimática [27]
2.4. Inibidores enzimáticos
Existem diversos tipos de moléculas que podem alterar a atividade das
enzimas e a cinética de suas reações. Inibidores são compostos que diminuem a
atividade enzimática, enquanto que os ativadores a aumentam. Nesta seção
iremos apresentar os efeitos dos inibidores nas reações catalisadas por
enzimas.
Inibidores enzimáticos são moléculas muito estudadas nos dias de hoje,
principalmente por possuírem um potencial muito grande de aplicação como
fármacos.
A
indústria
farmacêutica
investe
muito
na
descoberta
e
no
desenvolvimento de novas moléculas com propriedades inibitórias.
Os inibidores enzimáticos podem ser separados em duas classes de
acordo com a natureza da interação destes com a enzima. Os inibidores
reversíveis são aqueles que formam exclusivamente ligações de caráter nãocovalente (ligações de hidrogênio, por exemplo) enquanto que os inibidores
irreversíveis possuem, além desse tipo de ligação, uma associação covalente
com a enzima.
Os inibidores reversíveis ainda podem ser divididos em competitivos ou
não-competitivos. Os competitivos são aqueles que competem com o substrato
ENZIMAS:
2-25
CINÉTICA
pelo mesmo sítio ativo (chamado de sítio de ligação S). Dessa forma, existe
uma dependência da afinidade da enzima pelo substrato com a concentração
deste. Temos então que
k1
→
k2
E + S ← ES →
E+P
k3
→
E + I ← EI
k− 3
k−1
Como tanto substrato como inibidor interagem no mesmo sítio, não é
p o s s í v e l a f o r m a ç ã o d o c o m p l e x o I E S . U ma o u t r a c o n s e q ü ê n c i a d e s s e f a t o é
que ambos devem ter uma grande similaridade estrutural.
Podemos
também
obter
uma
expressão
para
a
lei
de
velocidade
incorporando o efeito do inibidor ao que foi feito anteriormente. Temos então
que
v=
Vmax [S]
,
[I]
+ [S]
K m 1 +
K I
KI =
(2.15)
k− 3
k3
Notamos que na ausência de inibidor, a equação (2.15) se transforma em
(2.8). O termo adicional que multiplica a constante Km no denominador faz
com que o tempo para se atingir Vmax (que nesse caso não é alterado) seja
maior e pode ser considerado um valor aparente de Km maior que o real.
Os outros tipos inibidores reversíveis são os não-competitivos. Estes
inibidores atuam em sítios diferentes do sítio S dos substratos, podendo então
interagir tanto com a enzima E como com o complexo ES. As reações possíveis
para o inibidor são então
k3
→
E + I ← EI
k− 3
'
k3
→
ES + I ←
IES ,
k −' 3
KI =
k− 3
k3
K I' =
k −' 3
k3'
A inibição não-competitiva pode ser pura ou mista. Na inibição pura, a
ligação do inibidor à enzima não afeta a ligação do substrato, enquanto que na
mista, a ligação do inibidor influencia a ligação do substrato na enzima. No
c a s o p u r o t e m o s q u e K I' = K I e a l e i d e v e l o c i d a d e é
2-26
CONCEITOS
DE
BIOQUÍMICA
v=
E
QUÍMICA ORGÂNICA
Vmax [S]
[I]
1 +
( K m + [S])
K I
(2.16)
N o c a s o m i s t o , K I' ≠ K I . e a l e i d e v e l o c i d a d e t e m a e x p r e s s ã o
v=
Vmax [S]
[ I ]
[I]
K m + 1 + ' [S]
1 +
KI
KI
(2.17)
Uma coisa que é interessante notar é que no caso puro, apenas Vmax é
alterado, não havendo modificação no valor de Km. Isso não ocorre no caso
misto, em que tanto Km como Vmax são modificados em relação ao valor real,
ou seja, existe um valor aparente para essas constantes. Podemos também
o b s e r v a r q u e s e K I' = K I e m ( 2 . 1 7 ) , e s t a s e t r a n s f o r m a n a e q u a ç ã o ( 2 . 1 6 ) .
Existem ainda os inibidores irreversíveis, que realizam uma ligação
covalente estável com a enzima, inutilizando-a. O efeito global desse tipo de
inibição é portanto uma gradual diminuição na quantidade de enzima ativa. O
padrão para a velocidade da reação observado é semelhante ao caso de inibição
reversível não-competitiva pura, mas com uma diminuição da atividade ao
longo do tempo.