Faculdade de Medicina da Universidade do Porto
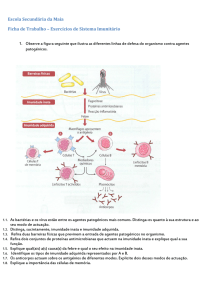
Ontogenia B
Imunologia
Ontogenia B
Aula leccionada por Dra Renata Ramalho a 06/10/2006
8 páginas
Como vocês sabem, todas as células do sistema imune se diferenciam a partir de uma stem cell, com
capacidade pluripotente, e o mesmo se verifica para as células B. No que diz respeito à maturação destas células,
ela ocorre ao nível da medula óssea. É formado um número astronómico de linfócitos B, mas apenas uma
pequena porção destas é que vai conseguir atingir a sua maturidade.
Mas porque é que o sistema imune se dá ao trabalho de produzir essa tão grande quantidade de células B?
Isto tem a ver com a capacidade de adaptação, porque, assim, produzimos uma grande quantidade de
células B, tendo a possibilidade de que, pelo menos uma delas vá reconhecer o antigénio.
Esta selecção que ocorre ao nível da maturação das células B tem a ver com uma série de características
que a célula tem de ter para poder continuar a progredir no processo de maturação.
No que diz respeito à maturação vamos ter uma fase antigénio-independente. Não necessitamos que o
antigénio esteja presente para que a célula B mature. Esta fase inicia-se na stem cell, esta vai proliferar numa
progenitora das células B e a partir de agora continuamos com uma célula pró-B. Para que a célula pró-B passe à
fase seguinte, célula pré-B, é necessário criar um microambiente especial, fornecido pelo estroma da medula
óssea e que tem duas funções importantes: por um lado vai promover a interacção directa entre as células pré-B e
pró-B e, por outro lado, tem a capacidade de produzir IL-7, que vamos ver que é essencial para a continuação da
proliferação dessa célula. Então, o contacto inicial ocorre com a ligação de célula pró-B ao estroma da medula
óssea e isto faz-se com moléculas de adesão, por um lado VLA-4 da célula pró-B e, por outro, VCAM-1 do
estroma da medula óssea.
O passo seguinte é a ligação de c-kit, expresso ao nível das células pró-B, que se vai ligar ao stem cell
factor (SCF) do estroma da medula óssea, o que vai promover uma activação do c-kit e, por sua vez, a
diferenciação em célula pré-B. Esta célula vai expressar na superfície o receptor para a interleucina IL-7, que,
como disse há bocado, é produzida pelo estroma da medula óssea. Esta ligação da interleucina ao seu receptor
vai promover a diferenciação da célula pré-B. Ela vai libertar-se do estroma da medula óssea, não vai mais
necessitar de estar ligada para que haja diferenciação e progressão para célula B imatura.
[ver slide 5, página 3]
Aqui têm uma imagem de microscopia electrónica para verem de forma mais realista este contacto, de que
estivemos a falar, entre a célula pró-B e o estroma. Este contacto vai ser, então, essencial para que haja
progressão para o estadio de célula pré-B. É neste estadio que ocorre o rearranjo ao nível dos genes para a
cadeia pesada (1º ao nível dos segmentos DJ e depois do segmento V para DJ) mas não nos vamos alargar mais
sobre isto….
Como podem verificar este estadio caracteriza-se, por exemplo, além do rearranjo completo ao nível da
cadeia pesada da célula pré-B, pela expressão da cadeia leve e do pré-BCR, do qual vamos falar já a seguir.
Também podemos verificar que as recombinases RAG-1 e RAG-2 estão presentes, o que não é de estranhar já
que há rearranjos genéticos, e estão tanto na célula pró-B como no estadio pré-B. Mas a enzima TdT (terminal
deoxyribonucleotide transferase), que cataliza a inserção de N-nucleotídeos, está activa apenas no estadio pré-B
e é inactivada na célula B imatura. Também há marcadores de superfície característicos, por exemplo, as células
pró-B apresentam à superfície CD-45.
Então a célula pré-B apresenta um pré-BCR. Nesta célula temos uma cadeia pesada µ que vai estar ligada
a uma pseudo-cadeia leve, esta é constituída por um segmento variable-like Vpre-B e outro segmento constant1/8
Ontogenia B
like que é o λ5. A ligação dessa cadeia µ à pseudo-cadeia leve forma o pré-BCR, o pré-receptor da célula B, que
vai estar ligado ao heterodímero Igα/Igβ. Este complexo constitui um passo importantíssimo para o
desenvolvimento da célula pré-B em célula B imatura, porque apenas as que apresentam um complexo pré-BCRIgα/Igβ efectivo é que podem continuar o processo de maturação. E progridem como? Progridem rearranjando os
genes ao nível das cadeias leves e formam um BCR completo. Por exclusão alélica temos apenas um isotipo de
cadeia leve, ou κ ou λ e nunca os dois em simultâneo. Este rearranjo permite uma maior diversidade.
Existem factores de transcrição que têm um papel importante na ontogenia das células B.
Existem dois tipos de células B diferentes: as células B1 e as células B2, que são as que existem em maior
quantidade e expressam BCR. As células B1 expressam CD5 e mIgM e constituem 5% das células B circulantes,
mas ao nível do desenvolvimento existem em maior quantidade. Aparecem durante o desenvolvimento na medula
óssea e têm uma característica importante, como sofrem pouca hipermutação somática e pouca mudança de
classe comparativamente com as células B2, então vão produzir Ig’s membranares de baixa afinidade.
Voltando ao que vos falei no início, temos várias células B potenciais, mas poucas delas acabarão por
maturar. Se eu vos disser que 10% chegam a célula B, então 90% das células fica pelo caminho, porque são
seleccionadas negativamente e sofrem apoptose. E porque é que isto acontece? Imaginem que à superfície da
célula B existe um anticorpo que é específico e reconhece antigénios do próprio, portanto temos auto-anticorpos
reactivos e não é isso que nós queremos. O sistema imune quer-nos proteger de agressões exteriores e não virarse contra nós. Então deve haver uma forma de lidar com estes anticorpos: fazer uma delecção clonal, ou seja,
acabar com eles, fazer apoptose, eliminá-los do organismo ou dar-lhes uma hipótese de serem re-editados ao
nível das cadeias leves ou ainda torná-los anérgicos, eles estão lá, mas não estão funcionais.
Até agora falámos da maturação das células B que ocorre sem contacto com o antigénio e ficámos então
nas células B maduras, células naive. Estas expressam à superfície IgM ou IgD, que reconhecem antigénios
estranhos (que é o que se pretende) e que vão migrar para os órgãos linfóides onde poderão encontrar o
antigénio, ser activadas e originar plasmócitos e células B memória.
De acordo com o tipo de antigénio que nós encontramos vamos ter uma activação diferente. Existem dois
tipos de antigénios: antigénios timo-dependentes, que para activarem a célula B requerem o contacto directo com
a célula T helper e antigénios timo-independentes, que não requerem qualquer tipo de contacto com a célula T
helper. Dentro destes últimos distinguimos 2 tipos: o tipo I e o tipo II. O tipo I é, sobretudo, constituído por
componentes da parede bacteriana e o tipo II por moléculas repetitivas, como a flagelina. O que distingue estes
tipos de antigénios? O tipo de activação e o facto dos antigénios timo-independentes tipo II, apesar de não
requererem contacto directo com a Th, necessitarem de um ambiente de citocinas por elas produzidas. Ao nível
da resposta humoral, a maior diferença entre os dois tipos é que os antigénios do tipo I, ao contrário dos
antigénios do tipo II, têm a capacidade de activar policlonalmente as células B, isto é, são mitogénicos.
Contudo, a maioria dos antigénios são timo-dependentes. Se eles requerem contacto com a célula T é
porque esta tem algum papel importante. De facto, o contacto directo com a célula T permite que ocorra mudança
de classe, maturação de afinidade, memória imunológica, todas fases importantíssimas para uma resposta
humoral eficaz.
Mas esta célula foi activada e vamos precisar de dois sinais específicos de activação e é necessário
existirem os dois tanto para antigénios timo-dependentes como para antigénios timo-independentes. O que
acontece de diferente é que, quando estamos na presença de um antigénio timo-independente, tanto o primeiro
sinal como o segundo são oferecidos quando há ligação cruzada do antigénio com a imunoglobulina de
membrana, certo? No entanto, quando temos um antigénio timo-dependente o crosslinking com a imunoglobulina
de membrana apenas oferece um sinal, o primeiro. Então, é necessário que haja a contribuição do linfócito T
helper para obtermos o segundo sinal da célula. Este segundo sinal é obtido pela ligação do CD40 ligando da
célula T helper activada com a célula B.
Estes dois sinais são extremamente importantes, porque só depois das células B os receberem é que ficam
activadas. Até agora alguma dúvida?
2/8
Ontogenia B
Então vamos avançar um bocadinho para a activação da célula B por antigénios timo-dependentes. Mas
como é feita a transmissão? Nós há pouco falámos do receptor da célula B e vimos que ele teria alguma
importância para a transmissão destes sinais de activação. Vamos então ver qual o seu papel. Como vemos aqui,
temos uma célula B e os heterodímeros Igα e Igβ, que têm os ITAMs, e que vão ser muito importantes na
transdução do sinal.
Começamos, então, com o antigénio que se vai ligar de forma cruzada à imunoglobulina de superfície.
[Esta ligação vai permitir que os ITAMs sejam fosforilados por proteínas cínases da tirosina.] Portanto, isto vai
promover uma afinidade que vai activar directamente o BCR, ou seja, estas cínases da tirosina fosforiladas que
estão ligadas aos ITAMs [na sequência da ligação cruzada do atg] vão activar o BCR. Concomitantemente temos
activação de segundos mensageiros, como por exemplo, o IP3 (trifosfato de inositol), que vai libertar cálcio
intracelular e que por sua vez vai activar e o diacilglicerol. Este vai activar cínases e, tudo isto, vai promover o
sinal de activação e proliferação das células B.
Este BCR pode ter a possibilidade de ser mais activo, de ter uma resposta mais amplificada, se tiver a
participação do co-receptor da célula B. Este co-receptor da célula B, que é constituído por estas proteínas CR2 e
CD19, vai permitir a amplificação da resposta a vários antigénios… Como é que isso vai acontecer? Têm, então, o
antigénio ligado à imunoglobulina de membrana, mas verifica-se aqui a presença de um complemento do sistema
complemento C3d, que está ligado ao antigénio, e que vai também ligar-se ao CR2. Esta ligação do CR2 ao C3d,
que por sua vez está ligado ao antigénio, vai promover a fosforilação de CD19. Isto vai fazer, também, a activação
de cínases e vai amplificar o sinal que o receptor da célula B tem para transmitir o sinal de activação. No fundo,
vamos conseguir fazer com que haja anticorpos com maior afinidade.
Então, voltando àquela parte de activação por antigénios timo-dependentes, vamos explorar um pouco
melhor essa activação. Após o contacto inicial com o antigénio temos a nossa célula B, CD40 e MHC da classe II.
Então, o antigénio ligado à imunoglobulina de membrana, promove o primeiro sinal de activação. Há inserção de
3/8
Ontogenia B
MHC classe II e o antigénio é degradado em peptídeos e apresentado à superfície da célula B. Ora, este sinal de
activação vai promover também a inserção de B7 à sua superfície. Vem então a nossa célula T helper, que vem
ajudar à activação. Porquê? Porque estamos na presença de antigénios timo-dependentes, e esta célula T helper
com o CD28 à sua superfície e o seu TCR vai ligar-se ao complexo MHC-peptídeo. Esta ligação promove a
inserção de CD40 ligando, o qual se vai ligar o CD40 da célula B, e isso constitui o tal segundo sinal de activação
que nós necessitávamos. Para além deste segundo sinal de activação vamos ter a poliestimulação produzida pela
ligação CD28 a B7 e isso vai promover a produção de citocinas por parte da célula T helper, que está agora
activada. A partir deste momento, a célula passou de um estadio de G0 para G1, prepara-se para introduzir à sua
superfície receptores para as citocinas produzidas pela célula T helper, continuando o seu processo de
diferenciação. Chega ao estadio S e, depois de estar activada, ela vai, então, sofrer mitoses, isto é, vai proliferar
em várias células B.
Portanto, até agora cumprimos esta ontogenia dos linfócitos B. Passamos de uma stem cell, para uma
célula progenitora (pro-B), uma célula precursora (pré-B), uma célula B imatura, e uma célula B matura. Depois, a
célula encontrou o antigénio, ficou activada e então temos a ontogenia da célula B completa. Mas vamos continuar
para perceber porque é que há essa ontogenia da célula B e o que ela tem a ver com a resposta humoral, que é
tão necessária para que nós consigamos lidar com antigénios do ambiente. Como vêem aqui, a localização das
células B, bem como das células T, está relacionada com locais anatómicos específicos. Aqui ao nível do córtex
temos uma zona claramente B; ao nível do paracórtex do gânglio linfático uma zona claramente T e depois ao
nível da medula podemos encontrar quer células B, quer células T. A activação das células B ocorre, assim num
folículo primário. Temos aqui nesta imagem uma coisa para a qual vos quero chamar a atenção, que é a chegada
ao centro germinativo. A transformação do centro germinativo vai ser, então, muito importante para o contacto,
para a diferenciação completa das células memória. Temos então o centroblasto, que é uma célula B activada,
recebeu o primeiro sinal de activação, pára…sofre mutação somática, proliferou… A célula B é expressa com
baixa afinidade. Para ter alta afinidade, ela vai ter que, ao migrar para a zona clara do gânglio linfático, competir
com as outras células B para se ligar às células foliculares dendríticas. A selecção vai ser feita para uma alta
afinidade, para os complexos antigénio-anticorpo expressos nas células foliculares dendríticas. Portanto, isto é um
passo muito importante, porque só aqueles que se conseguirem ligar vão poder passar à fase seguinte e contactar
com a célula T helper, certo? Portanto, não têm apenas de competir com as outras células B por um local de
adesão, como também têm de ter alta afinidade. Depois todas aquelas que forem seleccionadas vão entrar em
contacto com a célula T. Há, então, um segundo sinal de activação, que ainda na zona clara vai dar origem a
células que se diferenciam em célula B de memória ou em células produtoras de imunoglobulinas, os plasmócitos.
Isto ocorre pelo processo de hipermutação somática, mudança de classe e maturação da afinidade.
Muito bem, temos então uma célula B activada e para que ela prolifere e dê origem a outras células, é
necessário haver algum ambiente especial. É, então, necessário haver condições especiais para que isso possa
acontecer. Desta forma, um microambiente de citocinas é extremamente importante para o processo que a célula
vai sofrer. Assim, se nós estivermos na presença de interleucina 2 (IL-2), IL-4, IL-5, a célula B activada vai
diferenciar-se, proliferar em centros próprios e depois a partir daqui poderá dar origem a plasmócitos precursores
de imunoglobulinas de classes diferentes. Como é que isto acontece? Isto acontece por um processo de
comutação isotípica, em que há uma mudança na classe da imunoglobulina. Para isso muito contribuem algumas
citocinas, (algumas das quais já ouviram falar), por exemplo o interferão gama (que é importante na formação dos
vários subtipos de imunoglobulinas), o TNF-β (que poderá fazer com que os centrócitos se diferenciem em
plasmócitos produtores de IgA e todos os outros, portanto, IgA, IGE, IgG, IgM, enfim, das classes que nós
necessitamos para termos uma resposta humoral).
Como nós percebemos desde o início da ontogenia, a importância não é só ao nível dos plasmócitos.
Portanto, a importância de termos uma célula B que se consegue diferenciar em plasmócitos e células B de
memória, não está relacionada apenas com a produção de imunoglobulinas em quantidade suficiente. Uma coisa
também importante é nós ficarmos com memória para esses antigénios que encontrámos, e isso é uma
4/8
Ontogenia B
característica especial da nossa resposta secundária a um antigénio. Então, vamos ver qual a diferença entre as
células B de memória e as células B imaturas.
Uma célula B de memória, contrariamente a uma célula B naive, expressa marcadores membranares para
as imunoglobulinas IgM, IgD, IgG, IgA e IgE, ou seja, para diferentes classes de imunoglobulinas, ao contrário do
que acontece com as células B naive, que apenas expressam à sua superfície IgM e IgD.
No que concerne à localização, temos as células B naive essencialmente ao nível do baço e as células B
de memória a inserirem-se nos nódulos linfáticos e também na medula óssea.
No que diz respeito ao tempo de vida é essencial que as células B de memória tenham uma longa vida, não
é? Muito mais do que uma célula B naive. Ambas recirculam (diz ali, e é verdade), mas expressam receptores de
afinidades diferentes. As células B de memória têm uma maior afinidade, porque existiu maturação da afinidade, a
qual resulta de processos de mutação somática quando ocorre proliferação no centroblasto seguida duma
selecção daquelas que têm maior afinidade. Já perceberam isso? Daí que, a afinidade seja maior nas células de
memória que nas células B naive. No que diz respeito a moléculas de adesão celular também temos que as
células B de memória expressam uma maior quantidade de moléculas de adesão, neste caso ICAM-1.
Pergunta de uma colega nossa:
“As células B naive são células B imaturas?”
As células B naive não encontraram o antigénio.
“É igual às células imaturas?”
Não é bem a mesma coisa (células B naive e células B imaturas). As células B imaturas são aquelas que só
produzem IgM e IgD… Imaturas é um processo antes da maturação, antes de estarem maduras. A célula B naive
é uma célula B madura, mas que não encontrou o antigénio (portanto, não está activada). E pode nunca
encontrar, pode morrer no caminho… A maior parte das vezes as células B naive não encontram o antigénio.
Então, a memória imunológica é importante para constituírem, no fundo, uma resposta humoral, uma
resposta secundária, mais eficaz… Vamos ver:
5/8
Ontogenia B
Por exemplo: aqui na resposta primária (no primeiro contacto que temos com o antigénio) temos um lag
bastante largo. A resposta primária não é uma resposta muito amplificada e este primeiro contacto é caracterizado
pela formação de IgM, desenvolvendo-se em pouco tempo.
Precisávamos, de facto, quando encontrássemos o mesmo antigénio pela segunda vez, que essa resposta
fosse: primeiro mais eficaz, mais rápida e mais intensa, porque no fundo vamos encontrar um antigénio que já nos
foi apresentado e temos que saber lidar melhor com ele. Uma vez que já temos as células B de memória, então
(como veêm aqui, naquela parte a amarelo) a resposta vai ter um período lag muito mais curto, vai ter um período
de platô muito maior, vai ser muito mais intensa e, além disso, não conduz apenas a uma classe de
imunoglobulinas. Temos a classe de “IgG característica” e outros subtipos, não é? Porque como vocês viram pela
mudança de classe já vamos ter IgA, IgG, IgD, IgE para responder depois ao antigénio. Então, uma resposta
secundária é muito mais eficaz, muito mais intensa, com um período lag muito mais curto e um período de platô
maior, havendo, portanto, uma maior capacidade de responder ao antigénio de forma muito mais eficaz. Isto é
uma característica muito importante do nosso sistema imune adaptativo.
Aqui temos a comparação entre a nossa resposta primária a um antigénio e a nossa resposta secundária:
Como vêem a resposta primária é caracterizada essencialmente pela presença de células B naive,
enquanto que na resposta secundária, nós já lá temos as células B de memória que já encontraram antigénios. O
período lag é muito mais curto na resposta secundária, o tempo do pico de resposta é mais curto (demora muito
menos tempo a instalar-se uma resposta eficaz), a magnitude da resposta é claramente maior.
Os antigénios numa resposta primária são tanto timo-dependentes como timo-indepedentes e na
secundária os antigénios são timo-dependentes (até porque nós falámos que são os antigénios timo-dependentes
que promovem a memória imunológica - nenhum dos antigénios timo-independentes promove memória
imunológica). No que diz respeito à afinidade do anticorpo é muito mais alta na resposta secundária.
Alguma dúvida em relação a isto? Em relação à ontogenia? Não?
Então, como eu vos falei no início, se nós temos tantas células B (potenciais células B) e tantas morrem
pelo caminho, já percebem como é que isso acontece? Porque é que isso acontece? Por exemplo se existirem
células B que expressam à sua superfície auto-anticorpos contra o antigénio do próprio, isso não é benéfico. Há
alguma situação que vocês conheçam onde o nosso sistema imunológico tenha falhado e isso tenha acontecido?
[Alguém dá um exemplo…]
Exactamente. Deste modo, o nosso sistema imunológico foi, entre aspas, “desenhado” para que nos
pudesse defender das agressões exteriores, mas também tem a possibilidade de falhar. E quando falha reflectese em situações mais de imunopatologia. Assim, é necessário que o nosso sistema imunológico esteja altamente
vigilante para que nenhum mecanismo possa falhar a montagem de uma resposta. Mas também pode falhar, é
certo.
6/8
Ontogenia B
Pergunta de um colega nosso:
“Não percebi a noção de antigénio timo-dependente e timo-independente, por isso, não percebo porque é
que…?”
A diferença entre um e outro?
“Sim”
Os antigénios timo-independentes são antigénios que para activarem a célula B não necessitam de
contacto com a célula T helper, portanto, fazem a activação da célula B por eles próprios. A ligação cruzada
desses antigénios com a célula B promove, automaticamente, os dois sinais de activação e fica a célula B
activada.
“Para uma célula de memória…”
A memória imunológica tem a ver com os antigénios timo-dependentes, porque não há antigénios timoindependentes que promovam memória imunológica. A resposta humoral pelos dois tipos de antigénio é diferente!
A memória imunológica não está presente quando há antigénios timo-independentes a activar a célula B!
[Intervenção do prof. Luís Delgado, a propósito de mais uma pergunta colocada pelo nosso colega, mas
que está imperceptível na gravação.]
“Há antigénios, que, como viram ali, activam directamente as células B, mas esses antigénios (apesar de
parecerem vantajosos porque não precisam de outras células) indo directos às células B levam ao
desenvolvimento de uma resposta que é relativamente fraca, isto é, não é uma resposta muito eficaz. Por outro
lado, há produção só de IgM. Estas células [células B activadas por antigénios timo-independentes] não
conseguem vir a produzir mais tarde outras imunoglobulinas, portanto a resposta é relativamente monótona. E é
geralmente dita uma resposta de baixa afinidade, quer dizer que os anticorpos que vão ser produzidos para esses
antigénios timo-independentes muitas vezes são só IgM. Claro que a IgM também tem algumas vantagens,
porque é uma imunoglobulina grande, mas o facto de ter uma baixa afinidade quer dizer que a resposta também
não é muito eficaz. E isso tem alguma importância porquê? Porque alguns destes antigénios timo-independentes
são, por exemplo, polissacáridos dos microorganismos e este é tipicamente o grupo de antigénios microbianos
para os quais é mais difícil induzir vacinação, por exemplo. Há algumas vacinas que demoram muitos anos a
desenvolver, e algumas delas só apareceram nos últimos 10 ou 15 anos (estou a falar a da meningite, a do
pneumococos), exactamente porque era difícil induzir uma memória imunológica. Como vocês sabem, uma das
vantagens de ter memória de resposta é que nós “damos a volta” utilizando a vacinação para fazer essa memória.
Ora bem, os antigénios timo-independentes são daqueles para quem, tipicamente, criar memória é uma chatice,
de tal maneira que as vacinas demoram mais uns anos a desenvolver e, geralmente, não são tão eficazes. Quer
dizer que é preciso, muitas vezes, combinar antigénios diferentes (não podem ter só um ou dois tipos de
antigénios, têm que ter uma série deles, por exemplo polissacáridos). Às vezes têm que ser conjugados com
proteínas para modificar a resposta e muitas vezes o indivíduo tem que receber várias vacinas, sendo em muitas
delas o esquema vacinal mais complicado (passados 3 ou 4 anos o indivíduo tem que ser revacinado porque a
resposta desapareceu, exactamente por causa desta falta de memória).
Portanto, quando se fala da maturação parece vantajoso [o antigénio ser timo-independente], mas
realmente depois, na eficácia final da resposta, não é. Por isso é que nós dizemos que, de uma maneira geral, a
maior parte dos antigénios são timo-dependentes, isto é, precisam das células T para uma resposta eficaz.”
“Professor, mas então nos timo-independentes termos sinal 1 e sinal 2 não faz muito sentido, pois não?”
“Repara, há um 2º sinal porquê? Muitas vezes estes antigénios actuam através de um receptor. O conceito
de 2º sinal é um conceito mais funcional, isto é, a especificidade é dada essencialmente pelo 1º sinal (ligação à
imunoglobulina de membrana) e este é um sinal crítico para uma célula ser recrutada, porque se ela não
reconhecer o 1º sinal não vai responder àquele antigénio. Mas o que nós sabemos hoje, é que não chega esse
reconhecimento. Nós sabemos que há um 1º sinal, porquê? Porque tem a ver com a mobilização dos anticorpos
que estão na membrana, e isso é importante. Os anticorpos que estão na membrana da célula, desde que seja a
célula naive, são IgMs. E o que é que acontece quando há ligação do antigénio à célula? Esses receptores
7/8
Ontogenia B
começam a mexer-se na membrana e ao mexerem-se na membrana, digamos que, “irritam” aquela célula.
Qualquer coisa está a acontecer. Esse é o 1º sinal e é crítico para a célula ser activada. Mas o que nós sabemos é
que, é preciso mais qualquer coisa. É preciso todas aquelas cínases intracelulares serem mobilizadas. Por isso é
que as células precisam, para uma activação eficaz e para uma proliferação eficaz, de um 2º sinal. Esse 2º sinal
muitas vezes nos antigénios timo-independentes é dado por características próprias do antigénio.
Quando nós falarmos nos antigénios, numa aula mais adiante, vocês vão ver que uma característica destes
antigénios é, por exemplo, serem muito monótonos, isto é, têm constituintes muito repetidos ao longo da sua
constituição. Mas, o facto de terem esta repetição torna-os vantajosos na activação da célula B, porquê? Porque
conseguem cruzar múltiplos daqueles receptores de superfície. Digamos que ao cruzar múltiplos, funciona como
um primeiro sinal, um bocadinho melhor. E depois, alguns deles actuam via outros receptores da célula B,
nomeadamente, os lipopolissacáridos. Estes, por exemplo, podem ligar-se a alguns toll-like receptors (que são
receptores de perigo das células imunológicas e de outras células leucocitárias) e essa ligação a outros receptores
é que funciona como 2º sinal. Portanto, se vocês repararam no diapositivo:
Está ali um 2º sinal, ali a meio, que não está identificado. Enquanto que nós do lado direito temos
identificados os segundos sinais primordiais, do outro lado o 2º sinal não está identificado, mas nós sabemos que
ele muitas vezes é necessário. Isto é, muitas vezes é necessário uma interacção com os receptores, que não são
receptores de especificidade, mas que, de algum modo cooperam na activação. O exemplo típico são os
lipolpolissacáridos, que às vezes conseguem estimular as células até só pelo 2º sinal. Por exemplo, a Renata
chamou atenção que alguns antigénios timo-independentes podem funcionar como mitogénios para as células B.
O que é que isto quer dizer? Eles induzem a mitose das células B independentemente da especificidade. Ora
bem, e isso acontece quando? Quando eles estiverem em concentrações muito altas, porque eles vão actuar
através desses outros receptores, receptores, por exemplo, do tipo toll-like receptors.
O conceito de 2º sinal também é válido para os antigénios timo-independentes, mas é um 2º sinal que é
diferente do 2º sinal dos timo-dependentes, pois é um sinal mais complicado, como vocês viram (já ali [no slide]
estão identificadas várias moléculas) e que funcionalmente tem um outro significado. É que quando a ajuda vem
das células T, estas células não mudam a especificidade, mas melhoram-na um pouco no condicionamento deste
segundo sinal. Quer dizer que as células T vão impor um destino um bocadinho diferente aos linfócitos B e esse
destino é empurrá-los para a memória imunológica, ok?”
Esta é a desgravação integral da aula com apenas uma ou outra correcção. Esperamos que vos seja útil!
Bom estudo e muita sorte para as freqências!
Ana Filipa Castro
Ana Margarida Balsa
Ana Luís
8/8