Sistema imunitário
Imunidade
capacidade de resistir a agressões de
substâncias estranhas, como microrganismos
e substâncias químicas nocivas.
Tecido epitelial
Tecidos epitelial
são constituídos por células poliédricas, justapostas, entre as
quais existe pouca substância intercelular.
As suas principais funções são as de
revestir e proteger superfícies (ex: epiderme)
a absorção e secreção (ex: epitélios intestinais),
excreção (ex: glândulas)
funções sensitivas (neuroepitélios).
Tecido epitelial
Caracteristicas dos epitélios
Estão apoiadas sobre uma camada de tecido conjuntivo, que é
designado por lâmina própria.
As células epiteliais são geralmente polarizadas.
Apresentam uma porção ou polo basal e uma porção ou
polo apical.
Tecido epitelial
A forma do núcleo das células acompanha a forma das células
Na superfície de contacto entre as células epiteliais e o tecido
conjuntivo há uma estrutura chamada lâmina basal
A lâmina basal é constituída por colagénio tipo IV, por
glicoproteínas e proteoglícanas e liga-se ao tecido conjuntivo por
fibrilas de ancoragem.
duas lâminas basais originam uma membrana basal
Tecido epitelial
a nutrição das células epiteliais é feita por difusão, a partir do
tecido conjuntivo.
os epitélios possuem terminações nervosas.
os tecidos epiteliais são continuamente renovadas por
actividade mitótica.
as mitoses ocorrem na camada basal do epitélio, a camada
mais interna próxima à lâmina basal.
Tecido epitelial
As células epiteliais apresentam uma grande adesão uma grande
força mecânica para separa-las.
A adesão entre as células é em parte devida à acção coesiva de
glicoproteínas, chamadas canderinas (necessitam de Ca2+).
Apresentam especializações que constituem as junções intercalares.
Estas junções funcionam como locais:
adesão (zônulas de adesão, hemidesmossomas e os desmossomas),
impermeavéis (zônulas de oclusão)
comunicação (junções comunicantes ou junções gap).
Tecido epitelial
As junções de adesão e oclusão, são as junções mais apicais.
Ao conjunto da zônula de oclusão e a zônula de adesão que circunda
toda a parede lateral da região lateral denomina-se complexo unitivo.
As proteínas das junções comunicantes, chamadas conexinas,
organizam-se em hexameros em torno de um poro hidrófilo. Este
conjunto é designado por conexon, é a unidade estrutural da junção
formada por centenas dessas estruturas.
Os conexons de células vizinhas unem-se de modo a formar canais
hidrófilos entre as 2 células. Estas junções permitem o intercâmbio de
moléculas, permitindo que as células de muitos tecidos trabalhem de
maneira coordenada (exemplo: discos intercalares das células dos
tecidos musculares cardíacos).
Tecido epitelial
Tecido epitelial
Desmossoma ou mácula densa
estrutura complexa, em forma de disco, presente na superfície de
uma célula, que é sobreposta a uma outra estrutura idêntica presente
na outra célula. Em cada célula há uma placa circular denominada
placa de ancoragem.
As canderinas participam na adesão dos desmossomas.
Hemidesmossomas
Estas junções têm uma estrutura de meio desmossoma e prendem a
célula epitelial à membrana basal.
No hemidesmossomas em vez de canderinas, participam as
integrinas.
Tecido epitelial
Tecido epitelial
Especializações da superfície livre das células epiteliais
A superfície livre de algumas células epiteliais possui modificações
que permitem aumentar a sua superfície ou mover partículas.
Microvilos
São extensões cilíndricas da membrana plasmática.
A função é aumentar a área de superfície de contacto.
Apresentam apenas 1/10 a 1/20 do tamanho dos cílios.
Não se deslocam e são sustentadas por filamentos de actina.
Podem modificar-se e funcionar como receptores sensoriais, por
exemplo, no ouvido interno, reagem ao som.
Tecido epitelial
Estereocílios
São microvilos longos, imóveis e ramificados.
Aumentam a área de superfície da célula facilitando o movimento das
moléculas para dentro e fora das células.
Encontram-se nas células do epidídimio e do ducto deferente.
.
Tecido epitelial
Cílios e flagelos
Os cílios são apêndices que se projectam da superfície das células com
capacidade de se movimentar.
Apresentam uma forma cilíndrica com 2 túbulos centrais e 9 microtúbulos
periféricos fundidos, padrão 9+2.
O movimento dos microtúbulos requer ATP e é responsável pelo
movimento ciliar.
Os braços de dineína, proteínas que ligam entre si os pares adjacentes de
microtúbulos, provocam o deslizamento dos microtúbulos uns sobre os
outros.
Os cílios movem-se de uma forma coordenada.
Tecido epitelial
Tecido epitelial
Os flagelos apresentam uma estrutura idêntica à dos cílios mas são mais
compridos.
Existe um flagelo por espermatozóide.
Ao contrário dos cílios, os flagelos não deslocam pequenas partículas ao
longo das superfícies celulares, sendo responsáveis pelos movimentos das
próprias células.
Ao contrário dos cílios que apresentam um movimento pendular ou em
chicote, os flagelos deslocam-se em movimentos ondiformes.
Tecido epitelial
Deficiências nos genes que codificam as proteínas ciliares provocam o
síndroma da imobilidade ciliar que estruturalmente origina a ausência ou
anomalia dos cílios provocando:
na embriogenese que os órgãos não assumem a posição anatómica
normal (ex: coração no lado direito);
falha da remoção do muco dos pulmões originando recorrentes infecções
que podem provocar a morte
infertilidade, porque o transporte dos óvulos na trompa de falópio e a
mobilidade
da
cauda
funcionamento dos cílios.
dos
espermatozóides
depende
do
normal
Imunidade inata
O organismo reconhece e destrói algumas substâncias estranhas,
mas a resposta é sempre a mesma em cada exposição, apresenta
uma resistência não específica
Está presente nos invertebrados e também nos vertebrados
Fornece a 1ª linha de defesa contra as infecções
Imunidade adaptativa
O organismo reconhece e destrói substâncias estranhas, mas a
resposta aumenta cada vez que se dá uma exposição a um
agente agressor, apresenta uma resistência específica
Nos vertebrados a imunidade inata é complementada pela
imunidade adquirida
A imunidade adquirida embora mais lenta é muito mais específica
que a inata
A especificidade e a memória são características da
Imunidade adaptativa, e estão ausentes na imunidade
inata.
A imunidade inata apresenta:
1- factores mecânicos;
2- mediadores químicos;
3- Células envolvidas na fagocitose.
1. Factores mecânicos
Formam barreiras protectoras que evitam a entrada
e que removem da superfície corporal microrganismos.
2. Mediadores químicos
São moléculas que impedem a entrada de microrganismos
nas células (ex. lisozimas e o muco), que favorecem o
estabelecimento da inflamação (histamina, o complemento,
as prostaglandinas) e que protegem as células das infecções
virais (ex. interferões)
Citocinas
São moléculas semelhantes a hormonas que participam na
regulção de respostas imunes
Citocinas
São moléculas semelhantes a hormonas que participam na
regulação de respostas imunes.
São secretadas por linfócitos, macrófagos, células endoteliais,
neurónios, células da glia e outras células do nosso organismo.
São normalmente designadas com base nas suas funções, por
exemplo, factor de diferenciação das células B. No entanto, existe
uma convenção de que, uma vez definida a sequência de
aminoácidos desses factores, ele passa a ser designada por
interleucina
Algumas das funções clínicas mais importantes das interleucinas:
Patogenia do choque séptico
Artrite reumatóide
Aterosclorose
Participam no tratamento do carcinoma das células renais
Participam no processos de alergia
Participam no processo inflamatórios causadas por parasitas
Estão associadas ao combate de glomerulonefrite
Estão associas aos diabetes
Implicadas na patologia da esclorose múltipla
Regeneração da medula óssea
Associadas ao combate na doença granulomatosa crónica.
Podem ser boas coadjuvantes de vacinas
Outro tipo de citocinas é a família das quimiocinas
São substâncias que atraem os neutrófilos e outros
leucócitos até áreas de inflamação ou de resposta
imune.
Participam ainda na regulação do crescimento celular e
na angiogênese.
Os receptores das quimiocinas são receptores em
serpentina, que actuam através de proteínas G.
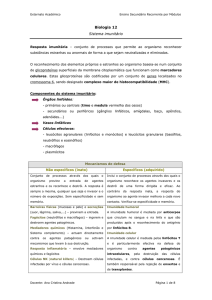
Os efeitos da imunidade inata e adquirida na destruição de
células são mediados, em, parte por um sistema de
enzimas plasmáticas denominado sistema complemento.
Este sistema é activado por 3 vias:
1. Via clássica – iniciada por acção de Imunocomplexos;
2. Via da lectina de ligação de manose – desencadeada
quando a lectina se liga a grupos de manose nas bactérias
3. Via alternativa ou da properdina – activada pelo contacto
com diversas bactérias, vírus, fungos e células tumorais
As proteínas do complexo podem:
Formar o complexo de ataque à membrana
(MAC)provocando a lise das paredes das bactérias;
aderir à superfície das bactérias e estimular os
macrófagos a fagocitar as bactérias;
atrair células do sistema imunitário para os locais de
infecção e induzirem a inflamação
A via clássica é usada na imunidade adaptativa.
Interferão
São proteínas que protegem os organismos contra as infecções virais e
algumas formas de neoplasias.
Vírus
célula
replicação viral
infecção de novas células
Os vírus e outras substâncias também vão estimular as células a produzir
interferões
Estes interferões vão se ligar à superfície das células vizinhas, estimulandoas a produzir proteínas anti-virais que impedem a replicação viral nestas
células
A resistência pelos interferões não é específica, os mesmos interferões
actuam contra diversos tipos de vírus
Alguns dos interferões também têm funções na activação de células
imunitárias
3- Células envolvidas na fagocitose (leucócitos)
Apresentam quimiotaxia
Neutrófilos
. São as primeiras células a entrar nos tecidos infectados.
. Libertam enzimas lisossómicas, que matam microrganismos e que
lesam e inflamam os tecidos.
Macrófagos
. São monócitos que deixam o sangue e entram nos tecidos onde
aumentam em cerca de 5 vezes o seu tamanho.
. São responsáveis pela fagocitose e pela remoção dos neutrófilos
mortos.
. Produzem diversas substâncias como proteínas de complemento e
interferões, reforçando a resposta imunitária.
. Podem ser denominados por células de Kupfer no fígado, por
macrofagos alveolares nos pulmões, e células de microglia no
sistema nervoso central.
Basófilos
. São leucócitos móveis que podem deixar o sangue e penetrar no
tecido conjuntivo;
Mastócitos
. São imóveis e residem no tecido conjuntivo, principalmente perto
dos capilares
Os basófilos e os mastócitos libertam substâncias quimícas tais como
a estamina e por isso estão associados a reacções alergicas
Eosinófilos
. São leucócitos móveis que podem deixar o sangue e penetrar no
tecido conjuntivo;
. Nas infecções parasitárias o número de eosinófilos é geralmente
muito elevado
Células natural Killer (NK)
. São um tipo de linfócito que reconhecem vários tipos de células,
como células neoplásicas ou células infectadas por vírus.
. Não apresentam especificidade
. Não têm memória, por isso são classificadas como imunidade inata.
. Matam as células alvo de diversas formo, libertando substâncias
que destroem a membrana e provoca a lise celular.
Resposta inflamatória
Sequência complexa que envolve
muitos mediadores quimícos e células
da imunidade inata
A inflamação local é confinada a uma
área específica do organismo. Provoca
edema, rubor, calor e dor.
A inflamação sistémica é generalizada
(ocorre em diversos locais do nosso
organismo). 1. A médula óssea
vermelha produz e liberta muitos
neutrófilos que promovem a fagocitose;
2. libertação dos pirogénios, substâncias
químicas que estimulam a produção da
febre; 3. Aumento muito acentuado da
permabilidade vascular
Imunidade adaptativa
As substâncias que estimulam uma resposta imunitária são designadas por
antigénios.
A imunidade adaptativa está dividida em duas partes:
. Imunidade humoral ou imunidade mediada por anticorpos
. Imunidade celular
Na imunidade humoral, as células B estimulam a produção de proteínas
específicas, os anticorpos, que circulam no plasma.
Na imunidade celular, as células T são as responsáveis pela imunidade mediada
pelas células. Existem diversos tipos de células T e cada um deles é responsável
por um aspecto da imunidade mediada pelas células.
Origem e maturação dos linfócitos
Medula vermelha óssea (células estaminais)
Circulação
Células pré-T
Células pré-B
Células B
Células pré-T
Timosina
Células T
Circulação
Timo
Sistema
linfático
Órgãos linfáticos primários, a medula óssea vermelha e o timo, são os locais
onde os linfócitos amadurecem, transformando-se em células funcionantes
Órgãos linfáticos secundários, tecido linfático, nódulos linfáticos, amígdalas,
gânglios linfáticos e baço, são os locais onde os linfócitos interagem entre si e
com os antigénios para produzirem uma resposta imunitária.
Figure 18.08
Activação dos linfócitos
1.
Os linfócitos têm de ser capazes de reconhecer o antigénio
2.
Após o reconhecimento, o número de linfócitos tem de aumentar
para que ocorra uma destruição eficaz do antigénio
Determinantes e receptores antigénicos
Os linfócitos vão interagir somente com determinadas regiões específicas
de cada antigénio, os determinantes antigénicos ou epitopos.
Um determinante antigénico só se pode combinar com um receptor
antigénico específico no linfócito.
Antigénios do complexo maior de histocompatibilidade (MHC)
A activação dos linfócitos pode ser directa ou com a intervenção de glicoproteínas
da superfície das células, os antigénios do MHC.
Estes antigénios fixam-se na membrana celular e têm uma região variável que
reconhece os antigénios.
Antigénio MHC I
Os antigénios MHC I, encontram-se nas células nucleadas e têm como função
expressar os antigénios produzidos dentro das células.
Na superfície da célula, o complexo antigénio - antigénios do MHC I pode ligar-se a
um receptor das células T, activando a célula T
O complexo antigénio - antigénios do MHC I induz o sistema imunitário à
destruição da célula em causa.
Antigénio MHC II
Os antigénios MHC II, encontram-se nas células apresentadoras de antigénios,
onde se incluem as células B, os macrófagos, os monócitos e as células
dendríticas.
As células apresentadoras de antigénios, estão especializadas em captar
antigénios estranhos, em processar esses antigénios e em levar os antigénios MHC
II a apresentar os antigénios estranhos a outras células do sistema imunitário. Ex.
o complexo antigénio – antigénios MHC II pode-se ligar aos receptores das células
T.
O complexo antigénio – antigénios MHC II funciona como um sinal que estimula
outras células do sistema imunitário a regir contra o antigénio.
Figure 18.12
O complexo antigénio – antigénios MHC II funciona como o primeiro sinal
necessário para induzir uma resposta imunitária. Mas, em muitos casos, é
também necessária a co-estimulação por outros sinais.
Essa co-estimulação pode ser efectuada por moléculas libertadas pelas células e
por moléculas ligadas à superfície das células.
As citocinas, tais como as interleucinas e a linfotoxina, são exemplos de
moléculas libertadas pelas células e que actuam como co-estimuladores. Estão
implicadas na regulação da imunidade, da reparação dos tecidos e ainda pelo
crescimento celular
As moléculas de superficie, como os CD (cluster differentiation), também podem
estar envolvidas no processo de co-estimulação.As células T helper apresentam
uma glicoproteína denominada CD4 e as células T citotóxicas apresentam uma
glicoproteína denominada CD8
Proliferação dos anticorpos
A exposição a um antigénio vai provocar um aumento de
linfócitos.
1. Proliferação das células T helper
2. Proliferação e activação das células B ou das células T
efectoras
Inibição dos linfócitos
A tolerância é um estado no qual os linfócitos não respondem
a um antigénio específico
É necessário manter a tolerância contra os auto-antigénios
para evitar doenças auto-imunes.
A tolerância pode ser induzida através da:
1. Elimição dos linfócitos auto-reactivos
2. Prevenção da activação dos linfócitos
3. Activação das células T supressoras
Imunidade mediada por anticorpos
A exposição do organismo a uma antigénio, pode conduzir à activação das
células B e a produção de anticorpos responsáveis pela destruição dos
antigénios.
É eficaz contra os antigénios extra-celulares, como bactérias, vírus,
protozoários, fungos, toxinas e parasitas
Anticorpos ou imunoglobulinas
Existem cinco classes de imunoglobulinas: igG, IgM, IgA, IgE e IgD.
Todos os anticorpos apresentam quatro cadeias: 2 cadeias pesadas idênticas e duas
cadeias leves também idênticas.
Apresentam uma região variável, que é o local que se combina com o determinate
antigénico do antigénio, e uma região constante que é responsável pelas funções de
anticorpo
Figure 18.15
Linha germinal
Dn
Vn
Jn
RNA mensageiro
Vn
Dn Jn
Cn
Cn
Diversidade dos anticorpos
1.
Processo de rearranjo
•
Diversidade de combinações na linha germinal
•
Diversidade na altura da junção dos genes
2. Diversidade após o processo de rearranjo
•
Hipermutação somática
•
Conversão génica
Produção de anticorpos
Resposta primária
. Surge na primeira exposição de uma célula B específica
para um determinado antigénio, inclui múltiplas divisões celulares,
diferenciação celular e produção de anticorpos.
. Durante a estimulação as células B originam linfócitos de grande tamanho
. Na divisão celular formam-se células plasmáticas que produzem anticorpos,
enquanto outras voltam a transformar-se em pequenos linfócitos e tornam-se
células B de mémoria.
. A 1ª classe de anticorpos a ser formada como resposta a um antigénio é a IgM,
mais tarde por um fenómeno designado por swiching class são produzidos
outros
tipos de anticorpos
. Em média a resposta primária demora 3 a 14 dias para produzir anticorpos
suficientes para serem eficazes contra um antigénio
Produção de anticorpos
Resposta secundária
. Ocorre quando o sistema imunitário é exposto a um antigénio contra o qual ele
já tinha produzido uma resposta primária.
. Resulta da acção da células B de mémoria que, quando expostas ao antigénio,
rapidamente se dividem , para produzir células plasmáticas e uma grande
quantidade de anticorpos.
. A resposta secundária é muito mais eficiente:
. tempo necessário para começar a produzir anticorpos é muito menor;
. A quantidade de anticorpos produzidos é muito maior.
. As células B de memória são a base do sistema de imunidade adaptativa.
Imunidade mediada por células
. Constitui uma das funções das células T e é mais eficaz contra os
microrganismos intracelulares como vírus, fungos, bactérias intracelulares e
parasitas.
. A activação das células T contra os antigénios é regulada pelas células
apresentadoras de antigénio e pelas células T helper.
. Após a activação as células T dividem-se e formam as células T efectoras e as
células T de memória.
. As células T efectoras e as células T citotóxicas são responsáveis pelas reacções
imunitárias mediadas por células.
. As células T de memória podem fornecer uma resposta secundária e imunidade de
longa duração.
Imunidade mediada por células
. As células T citotóxicas têm duas acções:
. Lisar as células
. Produzir citocinas
. As células infectadas por vírus têm antigénios virais, as células tumoraise os
tecidos transplantados têm, à superfície, antigénios que podem estimular a
produção das células T citotóxicas
. as células T citotóxicas ligam-se às células-alvo e libertam mediadores químicos que
provocam a sua lise. Esta lise envolve uma proteína, a perforina, que faz um oríficio
na membrana da célula-alvo.
. As células T de hipersensibilidade retardada reagem aos antigénios através da
libertação de citocinas que participam na fagocitose e na inflamação, especialmente
nas reacções alergicas.