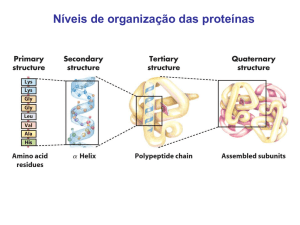
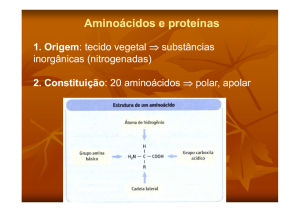
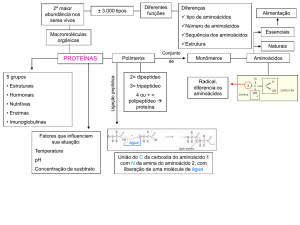
Processos gerais e síntese de aminoácidos; Rui Fontes
Processos gerais no metabolismo proteico e síntese de aminoácidos
1-
Um determinado número de moléculas de cada proteína endógena sofre hidrólise durante um dia mas,
nos indivíduos adultos saudáveis, um número equivalente é sintetizado. A percentagem de moléculas
afectadas por este processo de renovação depende principalmente da proteína em análise sendo muito
baixa no caso do colagénio (cerca de 0,2% de renovação diária), relativamente modesta no caso das
proteínas dos músculos esqueléticos (2%/dia), elevada no caso das proteínas das vísceras (7-15%/dia) e
elevadíssima no caso de enzimas reguladas por transcrição/tradução (renovação total em horas).
Considerando o conjunto das proteínas de um adulto (cerca de 10-12 kg de proteínas num adulto normal
com 70 kg) cerca de 300 g de proteínas sofrem hidrólise por dia e um valor idêntico sofre re-síntese o
que representa uma taxa de renovação de cerca de 3% (em média, 1 cadáver novo por mês). Apesar da
sua modesta taxa de renovação, porque as proteínas dos músculos constituem cerca de metade da massa
total de proteínas do organismo, a sua taxa de renovação (cerca de 120g/dia) contribui com 40% para a
taxa global. Em geral, um indivíduo adulto saudável mantém constante a quantidade total de
proteínas endógenas. De facto, a massa de proteínas endógenas “flutua” ao longo de um dia
aumentando no período pós-prandial e diminuindo durante o jejum. No entanto, tendo em conta a massa
total de proteínas, as variações percentuais são mínimas e, além disso, considerando um período de 24h
(ou mais) pode dizer-se que a velocidade de hidrólise é, no adulto, igual à de síntese. Um indivíduo que
está nestas condições diz-se em equilíbrio azotado (ou que tem um balanço azotado nulo).
2-
A hidrólise das proteínas endógenas é catalisada por protéases e a dos polipeptídeos formados por
peptídases acabando na libertação dos aminoácidos constituintes. No caso das proteínas citoplasmáticas
ou do retículo endoplasmático, nomeadamente as que têm taxas de renovação elevada (caso das enzimas
reguladas por transcrição/tradução) ou alterações estruturais, estes processos hidrolíticos envolvem
protéases do citoplasma que estão integradas numa estrutura proteica designada de proteossoma. As
proteínas que vão ser degradadas no proteossoma são previamente conjugadas com a ubiquitina (um
proteína) numa reacção em que se consome ATP. As proteínas extra-celulares ou associados à membrana
são sobretudo degradadas nos lisossomas por protéases designadas catepsinas. Os componentes
intracelulares que são degradadas nos lisossomas são previamente reunidos em estruturas rodeadas de
uma membrana constituindo os vaculos autofágicos. As proteínas segregadas para o lúmen do tubo
digestivo ou que resultam da descamação do epitélio são, juntamente com as proteínas da dieta,
hidrolisadas pelas protéases e peptídases digestivas.
3-
A esmagadora maioria dos aminoácidos formados durante a hidrólise das proteínas endógenas (cerca de
300g/dia) é reutilizada na síntese de novas moléculas proteicas mas uma parte não é reutilizada. Alguns
dos aminoácidos libertados no catabolismo das proteínas endógenas são transformados de tal forma que
ficam excluídos do ciclo de reutilização. Esta perda obrigatória de aminoácidos endógenos (cerca de
25 g/dia no adulto)1 é, em grande parte, uma consequência da presença, nas células, de enzimas que têm
como substratos aminoácidos e catalisam transformações catabólicas irreversíveis incluindo
desaminações e oxidações. O azoto dos aminoácidos que sofrem catabolismo é maioritariamente
transformado em ureia (que se perde na urina) enquanto o seu esqueleto carbonado (a parte
desprovida de azoto) pode ser oxidado a CO2, em última análise contribuindo para a síntese de ATP. O
azoto das proteínas não se perde apenas na forma de ureia. A urina contém outros compostos azotados
que, em última análise, também provêm do metabolismo dos aminoácidos; dentre estes destacaríamos a
creatinina, o ácido úrico, o ião amónio e, embora em quantidades muito mais pequenas, aminoácidos
(modificados ou não) e catabolitos de hormonas e neurotransmissores que tiveram aminoácidos na sua
génese. Também se perdem aminoácidos endógenos nas fezes pois uma parte das proteínas do epitélio
intestinal que descama, das mucinas secretadas (sobretudo do intestino grosso) ou mesmo das enzimas
digestivas não são completamente digeridas. O facto de, em média, 16% da massa das proteínas ser azoto
permite estabelecer uma relação entre a massa de azoto perdida nas excreções e a massa de proteínas que
essa massa de azoto representa. Assim, para converter a massa do azoto excretado em equivalentes de
1
A perda obrigatória de aminoácidos (obligatory aminoacids losses) pode, na prática, ser determinada avaliando as perdas
de azoto do organismo num indivíduo que tem uma dieta equilibrada sob todos os pontos de vista excepto um: não ingere
proteínas. Porque a ureia é, no rim e no intestino, reabsorvida para ser de novo re-excretada, é necessário esperar vários
dias antes de se tornarem patentes as consequências (diminuição da excreção de ureia) da exclusão das proteínas da dieta.
Página 1 de 8
Processos gerais e síntese de aminoácidos; Rui Fontes
massa de proteínas (ou aminoácidos) multiplica-se a massa do azoto excretado por 6,25 (100/16=6,25).
Cerca de 70% do azoto correspondente às perdas obrigatórias de aminoácidos perde-se na urina (50% na
forma de ureia e 20% na forma de creatinina, amónio e outros compostos) e cerca de 20% nas fezes; os
restante 10% correspondem às perdas de proteínas inteiras na pele que descama, nas unhas e cabelos que
crescem, nas secreções nasais, no fluxo menstrual ou na ejaculação e ureia do suor.
4-
Poderia pensar-se que, para repor as perdas obrigatórias de 25 g de aminoácidos/dia, bastaria ingerir uma
quantidade equivalente de proteínas mas não é isso que acontece. A absorção de aminoácidos no
intestino, leva a um aumento transitório da sua concentração nas células e a um aumento da velocidade
do seu catabolismo: uma parte substancial dos aminoácidos ingeridos fica sujeita à acção das enzimas
catabólicas sofrendo, junto com os libertados na hidrólise das proteínas endógenas, oxidação e
desaminação irreversível. Além disto, uma parte das proteínas ingeridas não chega a ser absorvida e
perde-se nas fezes. Os trabalhos experimentais com seres humanos adultos saudáveis apontam para
valores da ordem dos 50 g/dia como o mínimo de proteínas a ingerir para repor as perdas obrigatórias de
aminoácidos [1]. Nas situações em que a massa de proteínas endógenas está a aumentar diz-se que há um
balanço azotado positivo; na condição contrária diz-se que o balanço azotado é negativo; o balanço
azotado é nulo quando não há aumento nem diminuição da massa proteica. Porque é uma boa
aproximação à realidade considerar que a massa de aminoácidos livres (cerca de 150 g) é estacionária,
quando o balanço azotado é positivo (negativo, nulo) a massa de azoto ingerido é superior (inferior,
igual) à de azoto excretado; caso exista uma diferença entre os valores da síntese e da hidrólise de
proteínas esse valor reflecte-se numa diferença equivalente entre o azoto ingerido e excretado.
5-
Poderia pensar-se que a massa de proteínas ingeridas seria um importante factor na definição da variação
da quantidade de proteínas do organismo. A massa de gordura do organismo aumenta quando o valor
calórico da dieta é superior à despesa energética mas, no caso do azoto, o sistema funciona de forma
diferente. A massa de proteínas endógenas baixa (balanço azotado negativo) se a ingestão for inferior à
quantidade necessária para repor as perdas obrigatórias (25g/dia) e fazer face ao acréscimo de perdas
resultante da ingestão (outros 25g/dia) mas uma ingestão de proteínas acima do montante necessário
para cobrir as necessidades (50g/dia) resulta apenas no catabolismo dos aminoácidos
excedentários e num aumento da produção de ureia. Ao contrário do que acontece com a massa de
gordura, a quantidade de cada uma das proteínas do organismo só depende da dieta na medida em que (i)
esta pode constituir um factor limitador da sua síntese e (ii), acessoriamente, na medida em que o
aumento da massa de gordura é acompanhado pela formação de vasos sanguíneos, de adipócitos e de
tecidos de suporte. A quantidade total de proteínas do organismo aumenta (balanço azotado positivo)
nos indivíduos (i) em fase de crescimento (crianças e adolescentes), (ii) que estão a engordar, (iii) que
estão a recuperar após um período de balanço azotado negativo ou (iv) que, através de exercício físico
(ou ingerindo esteróides anabolizantes), estão a aumentar a sua massa muscular. O contrário (balanço
azotado negativo) acontece normalmente (i) a partir dos 40-50 anos de idade, (ii) quando se diminui a
actividade física ou (iii) quando se emagrece voluntariamente, (iv) em consequência de má nutrição ou
(v) em situações de doença. Contudo, num indivíduo adulto saudável que mantém constante a sua massa
muscular, a quantidade total de proteínas mantém-se também sensivelmente constante porque os
aminoácidos excluídos do ciclo de reutilização são repostos por ingestão e incorporados nas proteínas
sintetizadas.
6-
Poderia pensar-se que cada uma das moléculas de cada um dos aminoácidos que se perde para o ciclo de
reutilização teria de ser substituída por uma molécula igual mas esta ideia, só parcialmente, é verdadeira.
(i) Alguns dos aminoácidos excluídos do ciclo não podem ser sintetizados pelo organismo humano pois
não dispomos das enzimas indispensáveis para o processo e nestes casos os aminoácidos dizem-se
nutricionalmente indispensáveis (ou essenciais). Para substituir um determinado aminoácido
nutricionalmente indispensável que sofreu catabolismo é necessário ingerir esse aminoácido. Ou seja, no
caso dos aminoácidos nutricionalmente indispensáveis, cada molécula perdida tem de ser substituída por
uma igual. (ii) Alguns dos aminoácidos excluídos do ciclo podem ser repostos por síntese endógena a
partir de intermediários do metabolismo da glicose e, nestes casos, os aminoácidos dizem-se
nutricionalmente dispensáveis (ou não essenciais). No entanto, deve notar-se que, embora o esqueleto
carbonado provenha da glicose, o grupo azotado vem de outros aminoácidos que terão de ser ingeridos
em quantidade suficiente para colmatar as perdas de azoto. A “esqueleto carbonado” da alanina, por
exemplo, pode ser sintetizada a partir do piruvato mas o azoto da alanina “tem de vir” doutro
Página 2 de 8
Processos gerais e síntese de aminoácidos; Rui Fontes
aminoácido. (iii) Um terceiro grupo de aminoácidos (cisteína e tirosina) forma-se a partir de aminoácidos
indispensáveis (metionina e fenilalanina, respectivamente) e poderão classificar-se como semiindispensáveis2 [2, 3].
7-
No caso dos aminoácidos sintetizados a partir de intermediários do metabolismo da glicose (serina
[3C,1N,1OH], glicina [2C,1N], alanina [3C,1N], aspartato [4C,1N], asparagina [4C,2N], glutamato
[5C,1N], glutamina [5C,2N], prolina [5C,1N] e arginina [6C,4N]) embora o esqueleto carbonado possa
ser formado a partir da glicose, os grupos azotados (amina, amida ou guanidina) resultam da
transferência directa ou indirecta de grupos amina (ou amida) de aminoácidos para esses intermediários.
Para que um indivíduo adulto tenha a capacidade de manter constante a massa das suas proteínas precisa
de absorver, na forma de aminoácidos, tantos átomos de azoto como os que perde na urina, nas fezes, nos
genitais, nas secreções nasais ou nos tegumentos. Se a quantidade total de azoto ingerido (na forma de
proteínas) não for suficiente para colmatar o azoto excretado o indivíduo fica em balanço azotado
negativo. Em geral, a deficiência de aminoácidos nutricionalmente dispensáveis corresponde a uma
ingestão quantitativamente inadequada de proteínas: na presença de azoto aminoacídico em quantidade
suficiente para formar os grupos azotados o organismo pode sintetizar um aminoácido nutricionalmente
dispensável a partir de intermediários do metabolismo glicídico e, nesta síntese, todos os outros
aminoácidos são, em última análise, potenciais dadores de azoto.
8-
Através da acção catalítica de variadas enzimas os aminoácidos podem libertar o azoto do seu grupo
amina (ou de outros grupos azotados) na forma de amónio (NH4+). O ião amónio é a forma protonada do
amoníaco (NH3); o seu pKa é cerca de 9,3, predominando, por isso, a forma protonada quer no meio
interno quer na urina. A maioria do amónio (azoto inorgânico) formado dá origem a ureia que se perde
na urina, mas uma parte pode ser recuperado para o metabolismo por acção catalítica (i) da
desidrogénase do glutamato (ver equação 1) e (ii) da sintétase da glutamina (ver equação 2). Por
acção destas enzimas o azoto inorgânico do amónio pode ser convertido em azoto aminoacídico. O
glutamato [5C,1N] é um aminoácido dicarboxílico com 5 carbonos e difere do -cetoglutarato por ter,
em vez do grupo cetónico, um grupo amina no carbono 2. A glutamina [5C,2N] difere do glutamato
porque, em vez do grupo carboxílico em C5, tem um grupo amida nesse carbono.
-cetoglutarato + NH4+ + NADPH glutamato + NADP+ + H2O
glutamato + NH4+ + ATP glutamina + ADP + Pi
9-
Para além de poder ter origem na acção da desidrogénase do glutamato (ver equação 1), a síntese de
glutamato também tem lugar em reacções de transaminação (ver equação 3) em que diversos
aminoácidos cedem o grupo amina (azoto orgânico) ao -cetoglutarato gerando glutamato e os cetoácidos correspondentes. Assim, o glutamato e a glutamina (via sintétase da glutamina; ver equação
2) podem formar-se endogenamente a partir de um intermediário do ciclo de Krebs (o -cetoglutarato);
sabendo-se que os intermediários do ciclo de Krebs se podem formar a partir da glicose (via glicólise e
carboxílase do piruvato) conclui-se que o glutamato e a glutamina são aminoácidos nutricionalmente
dispensáveis.
-aminoácido X + -cetoglutarato glutamato + -cetoácido X
10-
(1)
(2)
(3)
A alanina [3C,1N] difere do piruvato porque, em vez do grupo cetónico no carbono 2, tem um grupo
amina; o aspartato [4C,1N] difere do oxalacetato pela mesma razão. A síntese de alanina e aspartato é o
resultado da transferência do grupo amina do glutamato para os -cetoácidos correspondentes: o
piruvato e o oxalacetato, respectivamente. A transamínase da alanina (ver equação 4) e a
transamínase do aspartato (ver equação 5) catalisam, respectivamente, a formação de alanina e
aspartato mas, como estas reacções são fisiologicamente reversíveis, também intervêm nos processos em
que estes aminoácidos perdem o grupo -amina para o -cetoglutarato. Existem muitas transamínases
com especificidades distintas relativamente a um dos substratos mas o outro substrato é (quase) sempre o
glutamato/-cetoglutarato (ver equação 3). Dependendo do sentido em que a reacção esteja a ocorrer
2
Quem classifica faz um exercício de organização dos conhecimentos da forma que lhe dá mais jeito. Também é frequente
chamarem à cisteína e à tirosina “condicionalmente indispensáveis” porque só são indispensáveis se a dieta for pobre em
metionina e fenilalanina, respectivamente.
Página 3 de 8
Processos gerais e síntese de aminoácidos; Rui Fontes
uma reacção de transaminação pode servir para formar um determinado aminoácido à custa da conversão
do glutamato em -cetoglutarato ou para formar glutamato à custa da conversão de um determinado
aminoácido no seu -cetoácido correspondente. Uma característica comum a todas as transamínases (e a
muitas outras enzimas envolvidas no metabolismo aminoacídico) é a presença de fosfato de piridoxal
(derivado da vitamina B6) como grupo prostético3.
glutamato + piruvato -cetoglutarato + alanina
glutamato + oxalacetato -cetoglutarato + aspartato
11-
A serina [3C,1N,1OH] é um aminoácido que contém 3 carbonos e um grupo hidroxilo em C3. A glicina
[2C,1N] é o aminoácido mais simples e contém apenas 2 carbonos. Transamínases com diferentes
especificidades intervém no processo de síntese da serina a partir de 3-fosfoglicerato (um
intermediário da glicólise) e da glicina a partir de glioxilato (contém um grupo aldeído em vez do
grupo amina no carbono ). No processo de síntese da serina a partir do 3-fosfoglicerato intervém
primeiro uma desidrogénase que converte o grupo hidroxilo do carbono 2 num grupo cetónico levando à
formação do 3-fosfohidroxipiruvato (ver equação 6) que é substrato da transamínase da fosfoserina (ver
equação 7). A fosfoserina (formada após a reacção de transaminação) é hidrolisada por uma fosfátase
com a consequente formação da serina (ver equação 8). O glioxilato (aceitador de grupos amina em
reacções de transaminação em que a alanina é o dador da amina; ver equação 9) pode resultar da
oxidação do glicolato (que existe em muitas plantas comestíveis) por acção da oxídase do glicolato (ver
equação 10)4.
3-fosfoglicerato + NAD+ 3-fosfohidroxipiruvato + NADH
glutamato + 3-fosfohidroxipiruvato -cetoglutarato + fosfoserina
fosfoserina + H2O serina + Pi
alanina + glioxilato piruvato + glicina
glicolato + O2 glioxilato + H2O2
12-
(6)
(7)
(8)
(9)
(10)
A reacção catalisada pela hidroximetiltransférase da serina (ver equação 11) para além de permitir a
síntese de glicina a partir de serina (e o inverso) também permite a metilação do tetrahidro-folato (H4folato): o N5,N10-metileno-H4-folato formado nesta reacção é indispensável na síntese de timina e
portanto do DNA. O facto de a glicina se poder formar a partir da serina (ver equação 11) e de esta poder
gerar-se a partir de um intermediário da glicólise (3-fosfoglicerato; ver equações 6-8) permite
compreender que quer a serina quer a glicina sejam aminoácidos nutricionalmente dispensáveis.
serina + H4-folato glicina + N5,N10-metileno H4-folato
13-
(4)
(5)
(11)
A prolina [5C,1N] é o único aminoácido em que o grupo amina é uma amina secundária (que liga os
carbonos 2 e 5). A arginina [6C,4N] contém 6 carbonos mas um deles faz parte da estrutura do grupo
guanidina [1C;3N] que se liga ao carbono 5. Quer a prolina quer a arginina podem ser sintetizadas a
partir do glutamato. O glutamato pode, por redução do grupo carboxílico C5, originar o semialdeído do
glutamato e este composto pode seguir dois destinos distintos: (i) num deles (por redução dependente do
NADPH) dá origem à prolina e (ii) no outro origina a ornitina e, posteriormente, a arginina. A
conversão do semialdeído do glutamato em ornitina é catalisada por uma transamínase (ver equação 12).
glutamato + semialdeído do glutamato -cetoglutarato + ornitina
3
(12)
No decurso do ciclo catalítico o piridoxal-fosfato que está, no início do ciclo, ligado ao grupo 6-amina de um resíduo de
lisina da transamínase, converte-se em piridoxamina-fosfato, mas, no final do ciclo, regenera-se a forma original.
4
A glicina também pode formar-se a partir da colina. Nesta via metabólica a colina é oxidada no grupo hidroxilo
formando-se betaína (trimetilglicina). A betaína é dadora de um metilo à homocisteína formando-se dimetilglicina (betaína
+ homocisteína dimetilglicina + metionina) que por sua vez pode ceder os dois restantes metilos ao tetrahidrofolato
(H4-folato) gerando-se a glicina (dimetilglicina + H4-folato sarcosina + N5,N10-metileno-H4-folato; sarcosina + H4folato glicina + N5,N10-metileno-H4-folato).
Página 4 de 8
Processos gerais e síntese de aminoácidos; Rui Fontes
14-
A arginina é sintetizada no ciclo da ureia [1C,2N] a partir de ornitina [5C,2N] e esta pode formar-se
(via semialdeído do glutamato) a partir do glutamato. A ureia é apenas sintetizada no fígado pois é neste
órgão que existe a argínase, uma hidrólase que catalisa a formação de ureia a partir da arginina. No
entanto, as enzimas que levam, a partir da glutamina (via glutamato) à formação de ornitina e à
conversão desta em citrulina [6C,3N] também existem nos enterócitos. A citrulina formada nos
enterócitos passa para o plasma sanguíneo e pode ser captada pelo fígado mas também pelo rim. As
enzimas “do ciclo da ureia” que catalisam a conversão sequenciada de citrulina em arginino-succinato e
deste em arginina (sintétase do arginino-succinato e arginino-succínase) existem nestes dois órgãos e
levam à formação de arginina. As enzimas do ciclo da ureia, para além do seu papel no catabolismo de
todos os aminoácidos também têm um papel anabólico: a síntese de arginina. A velocidade de
formação líquida de arginina (massa formada subtraída da parte que se converte em ureia e ornitina) é,
contudo, inadequada nos indivíduos em crescimento ou em situações em que, após um período em que o
indivíduo perdeu proteínas endógenas (por doença ou/e por má nutrição), este se encontra em fase de
convalescença a recuperar as proteínas perdidas. Do ponto de vista nutricional, Stipanuk [2, 3] classifica
a arginina como um aminoácido condicionalmente indispensável.
15-
A asparagina [4C,2N] difere do aspartato [4C,1N] porque, em vez do grupo carboxílico em C4, tem um
grupo amida nesse carbono. De forma semelhante ao que acontece no caso da glutamina e do glutamato,
a asparagina forma-se a partir do aspartato por acção catalítica da sintétase da asparagina (ver equação
13). No entanto, ao contrário do caso da síntese da glutamina em que o azoto incorporado é azoto
inorgânico, na síntese da asparagina, o dador do azoto é a glutamina. Além disso, na reacção catalisada
pela sintétase da asparagina, forma-se AMP e PPi e não ADP e Pi como no caso da sintétase da
glutamina (ver equação 2).
aspartato + glutamina + ATP asparagina + glutamato + AMP + PPi
16-
A fenilalanina [9C,1N] contém um anel benzénico; a tirosina [9C,1N,1OH] deriva da fenilalanina por
hidroxilação desse anel benzénico. É frequente classificar-se a tirosina como semi-indispensável
porque é sintetizada a partir da fenilalanina, um aminoácido nutricionalmente indispensável. Uma
deficiência nutricional de tirosina pode ser colmatada desde que ocorra a ingestão de fenilalanina em
quantidade adequada para satisfazer as necessidades dos dois aminoácidos. A reacção de formação da
tirosina é catalisada pela hidroxílase da fenilalanina, uma oxigénase de função mista (ver equação 14).
Para que o processo possa continuar a dihidrobiopterina formada é reduzida pelo NADPH numa reacção
catalisada por uma redútase (ver equação 15).
fenilalanina + tetrahidrobiopterina + O2 tirosina + dihidrobiopterina + H2O
dihidrobiopterina + NADPH tetrahidrobiopterina + NADP+
17-
(13)
(14)
(15)
O átomo de enxofre da cisteína [3C,1N,1S] tem origem na metionina [5C,1N,1S], um aminoácido
indispensável. Tal como no caso da tirosina, também a cisteína pode ser classificada como semiindispensável: as necessidades nutricionais de cisteína podem ser colmatadas desde que ocorra a
ingestão de metionina em quantidade adequada para satisfazer as necessidades dos dois aminoácidos. Os
carbonos da cisteína têm origem na serina. O processo de síntese da cisteína é complexo porque se
relaciona com a complexa via metabólica da degradação da metionina (ver equações 16-21). Durante o
catabolismo da metionina forma-se um intermediário (homocisteína) que contém ainda 4 carbonos da
metionina mas que, em vez do grupo metilo ligado ao carbono 4 por uma ligação sulfureto, contém um
grupo tiol. Este intermediário reage com a serina formando-se um composto (cistationina) que contém o
átomo de enxofre entre os carbonos que derivaram da homocisteína e da serina (ver equação 19). A
clivagem da cistationina (ver equação 20) origina cisteína (3 carbonos e azoto derivados da serina e o
enxofre da homocisteína) assim como NH3 e -cetobutirato (derivados da homocisteína).
ATP + metionina S-adenosil-metionina + Pi + PPi
S-adenosil-metionina + aceitador S-adenosil-homocisteína + aceitador metilado
S-adenosil-homocisteína + H2O homocisteína + adenosina
homocisteína + serina cistationina
cistationina cisteína + NH3 + -cetobutirato
Página 5 de 8
(16)
(17)
(18)
(19)
(20)
Processos gerais e síntese de aminoácidos; Rui Fontes
-cetobutirato + NAD+ + CoA propionil-CoA + NADH + CO2
(21)
Embora a metionina seja um aminoácido nutricionalmente indispensável existe um mecanismo que
permite "salvar" metionina em processo catabólico: a homocisteína é aceitadora do grupo metilo do N5metil-H4-folato regenerando-se metionina (síntase da metionina; ver equação 22). O N5-metil-H4-folato
forma-se por redução (dependente do NADPH; acção da redútase do N5,N10-metileno-H4-folato; ver
equação 23).
N5-metil-H4-folato + homocisteína H4-folato + metionina
N5,N10-metileno-H4-folato + NADPH N5-metil-H4-folato + NADP+
18-
Oito (valina, leucina, isoleucina, treonina, metionina, lisina, fenilalanina, triptofano) dos 20
aminoácidos5 que são incorporados nas proteínas aquando da sua síntese são, classicamente, classificados
como nutricionalmente indispensáveis. Com três excepções (treonina, lisina e triptofano) existem,
contudo, transamínases que (com maior ou menor eficácia) são capazes de catalisar a troca entre o
grupo cetónico dos -cetoácidos correspondentes e o grupo amina do glutamato [3]. Embora absurdo do
ponto de vista económico, seria possível usar os -cetoácidos correspondentes para substituir na dieta
uma grande parte dos aminoácidos nutricionalmente indispensáveis. As equações 24-26 mostram as
reacções de transaminação que envolvem os aminoácidos ramificados. Na prática, porque os cetoácidos referidos não fazem parte de uma dieta normal as reacções 24-26 são fisiologicamente
irreversíveis no sentido em que os aminoácidos essenciais (presentes na dieta ou que resultam da
hidrólise das proteínas endógenas) são consumidos. No caso da histidina também não existem, nos
mamíferos, vias metabólicas de síntese, mas a deficiência deste aminoácido só se torna aparente após
relativamente longos períodos de dietas sem histidina [4]. É possível que na origem desta resistência
esteja a capacidade de formar histidina a partir de carnosina, um dipeptídeo (-alanil-histidina)
abundante no tecido muscular. Embora alguns livros de texto classifiquem a histidina num grupo à parte,
de acordo com Kopple e Swendseid [4], a histidina é um aminoácido nutricionalmente indispensável.
-ceto-isocaproato + glutamato leucina + -cetoglutarato
-ceto--metil-valerato + glutamato isoleucina + -cetoglutarato
-ceto-isovalerato + glutamato valina + -cetoglutarato
19-
5
(22)
(23)
(24)
(25)
(26)
Tal como os aminoácidos dispensáveis também os aminoácidos indispensáveis sofrem catabolismo a
uma velocidade que depende da actividade intrínseca das enzimas envolvidas e da concentração do
aminoácido em causa. Para assegurar a manutenção da massa de proteínas do organismo há, não só que
ingerir uma quantidade total de aminoácidos adequada (em média 50 g/dia no adulto saudável com 70
kg), mas também que repor todos e cada um dos aminoácidos indispensáveis que se perderam. Tendo em
conta as necessidades mínimas de cada um dos aminoácidos indispensáveis foram inventadas proteínas
padrão: uma proteína padrão é uma proteína que, ingerida na quantidade mínima indispensável para
repor as perdas obrigatórias de aminoácidos totais, contém a quantidade mínima de cada aminoácido
indispensável para repor a perda individual de cada um destes aminoácidos [5]. Se uma dieta contiver
como único constituinte proteico uma proteína que não contém um aminoácido indispensável (caso da
gelatina que não contém triptofano) a capacidade dessa dieta para colmatar as necessidades
aminoacídicas é nula. Todas as proteínas endógenas contém pelo menos um resíduo de triptofano e, por
isso, nenhuma proteína pode ser sintetizada na ausência de triptofano e o mesmo poderia ser dito
relativamente a cada um dos outros aminoácidos indispensáveis. Quando se ingere como única proteína
gelatina nenhum dos aminoácidos que resultam da sua hidrólise intestinal pode ser usado na síntese
proteica porque falta o triptofano. Nestas circunstâncias, com a excepção do triptofano, todos os
aminoácidos aumentam de concentração aumentando a velocidade da sua oxidação. Quando se ingere
como única proteína gelatina a quantidade de azoto perdido é igual à perda obrigatória somada a toda a
gelatina ingerida cujos aminoácidos são também perdidos. No caso da gelatina o aminoácido limitante da
sua qualidade dietética é o triptofano mas no caso de outras proteínas como, por exemplo, nas proteínas
do trigo e outros cereais o aminoácido limitante é a lisina. No caso das proteínas do trigo a lisina não está
ausente mas existe numa quantidade menor que a prevista nas proteínas padrão. A percentagem de lisina
Ou 21, se considerarmos também o caso da selenocisteína.
Página 6 de 8
Processos gerais e síntese de aminoácidos; Rui Fontes
nas proteínas de trigo é cerca de metade da percentagem de lisina numa proteína padrão: assim, para
colmatar as necessidades de lisina usando exclusivamente proteínas de trigo haveria que ingerir não 50 g
de proteína de trigo mas o dobro deste valor [2, 6].
20-
Em algumas proteínas (como a peroxídase do glutatião) existem resíduos de selenocisteína, um
aminoácido semelhante à cisteína e à serina. Na selenocisteína em vez do átomo de enxofre do grupo tiol
(caso da cisteína) ou do átomo de oxigénio do grupo hidroxilo (caso da serina) existe um átomo de
selénio. A síntese da selenocisteína ocorre a partir da serina quando esta está ligada a um tRNA
específico que tem como anticodão a sequência ACU e se denomina tRNASec (Sec é a abreviatura de
selenocisteína). A reacção é catalisada por uma transférase (ver equação 27) em que o dador de selénio é
o seleno-fosfato (“selénio activado”). O codão correspondente ao tRNASec (UGA) é normalmente um
codão de terminação mas em determinados RNA mensageiros contendo sequências específicas (como é o
caso do RNAm codificador da peroxídase do glutatião) este codão liga-se ao anticodão do
selenocisteinil-tRNASec ocorrendo a incorporação do aminoácido selenocisteína na estrutura da proteína
em processo de síntese.
seleno-fosfato + seril-tRNASec selenocisteinil-tRNASec + Pi
21-
Os aminoácidos hidroxiprolina e hidroxilisina constituem casos especiais pois existem na estrutura do
colagénio (a proteína mais abundante dos mamíferos) mas não existem no RNA codificador do colagénio
codões para estes aminoácidos. A síntese da hidroxiprolina e da hidroxilisina ocorre por acção de
oxigénases do retículo endoplasmático (hidroxílases da prolina e da lisina) que catalisam a hidroxilação
de resíduos de prolina e lisina do colagénio durante o processo de acabamento pós-tradução (ver equação
28). A vitamina C é um cofactor das hidroxílases da prolina e da lisina e a deficiência de vitamina C leva
à formação de colagénio anormal.
resíduo prolil ou lisil + O2 + -cetoglutarato
resíduo hidroxiprolil ou hidroxilisil + succinato + CO2
22-
(27)
(28)
O caso do aminoácido carboxiglutamato (constituinte de várias proteínas como a protrombina e outras
proteínas envolvidas no processo de coagulação sanguínea) tem algumas semelhanças com os casos da
hidroxiprolina e hidroxilisina já que a sua formação resulta da transformação de resíduos de glutamato
após a síntese da proteína. A transformação envolve a actividade de uma oxigénase (ver equação 29) e
uma reacção não enzímica (ver equação 30). A regeneração da forma hidroquinona da vitamina K a
partir da forma epóxido envolve a acção de oxiredútases.
resíduo de glutamato + O2 + vitamina K (forma hidroquinona)
resíduo de glutamato na forma de carbanião + vitamina K (forma epóxido)
resíduo de glutamato na forma de carbanião + CO2 → resíduo de carboxiglutamato
(29)
(30)
1. Rand, W. M., Uauy, R. & S., S. N. (1984) Protein-Energy-Requirement Studies in Developing Countries: Results of
International Research in FOOD AND NUTRITION BULLETIN SUPPLEMENT (UNIVERSITY, T. U. N., ed),
http://www.unu.edu/Unupress/unupbooks/80481e/80481E00.htm, Tokyo 150, Japan.
2. Fuller, M. F. (2000) Protein and amino acid requirements in Biochemical and physiological aspects of human nutrition
(Stipanuk, M. H., ed) pp. 287-04, W.B. Saunders Company, Philadelphia.
3. Stipanuk, M. H. (2006) Biochemical, Physiological, Molecular Aspects of Human Nutrition, 2nd edn, Sunders,
Elsevier., St. Louis.
4. Kopple, J. D. & Swendseid, M. E. (1975) Evidence that histidine is an essential amino acid in normal and chronically
uremic man, J Clin Invest. 55, 881-91.
5. Young, V. R. & el-Khoury, A. E. (1995) Can amino acid requirements for nutritional maintenance in adult humans be
approximated from the amino acid composition of body mixed proteins?, Proc Natl Acad Sci U S A. 92, 300-4.
6. Schaafsma, G. (2000) The protein digestibility-corrected amino acid score, J Nutr. 130, 1865S-7S.
Página 7 de 8
Processos gerais e síntese de aminoácidos; Rui Fontes
colina
cisteína
glicose
betaína
cistationina
dimetil-glicina
homocisteína
sarcosina
metionina
3-fosfoglicerato
3-fosfohidroxipiruvato
3-fosfoserina
serina
glicina
glicolato
alanina
piruvato
oxalacetato
treonina
asparagina
aspartato
arginina
ureia
prolina
ornitina
Ciclo de
Krebs
semiladeído do
glutamato
NH3 + NAD(P)H
glioxilato
Carbamilfosfato
Ciclo
da ureia
citrulina
NAD(P)+
-ceto-glutarato
glutamato
fenilalanina
glutamina
tirosina
prolina
hidroxiprolina
lisina
hidroxilisina
glutamato
carboxiglutamato
Seril-t-RNASec
Selenocisteinil-t-RNASec
Página 8 de 8
argininosuccinato