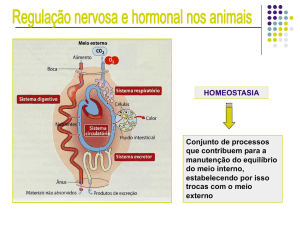
T26 – Tecido Nervoso
O sistema nervoso é constituído por neurónios (em número igual às estrelas da via Láctea) e por células
da glia. Podem ser astrócitos (de suporte metabólico), oligodendrócitos (podem estar associadas vários
axónios), células de Schawn (estão associadas a apenas um neurónio), microglia e células ependimárias.
Como é que os neurónios passam a informação?
Quando o neurónio é estimulado, permite a abertura dos canais de Na+ sensíveis à voltagem, pelo que o
interior da célula fica positiva, dá-se então o potencial de acção e a célula encontra-se despolarizada. Após o
pico máximo, é necessário voltar ao estado basal para ocorrer novo estímulo, aí a célula abre os canais de
potássio, e fica repolarizada, voltando ao seu estado electroquímico de repouso. Durante o potencial de
repouso é activada a bomba Na+-K+ em que há saída de 3Na+ e entrada de 2K+, de modo a manter o
potencial eléctrico negativo. Esta bomba ATPásica é responsável pelo consumo de 25-50% de ATP em
repouso.
Graças às células de Schwan, que contém mielina (isolador da passagem à corrente eléctrica), a
despolarização do axónio acontece apenas nos Nódulos de Ranvier. Diz-se, portanto, que a passagem do
impulso nervoso é de forma saltatória. Este facto vai aumentar em larga escala a rapidez da transmissão do
impulso nervoso, permitindo uma reacção rápida ao estímulo.
O impulso nervoso é também unidireccional, pois o neurónio pré-sináptico conte vesículas que contêm
neurotransmissores que, quando é atingido, liberta essas vesículas, por fusão com a membrana, e os
neurotransmissores são libertados para a fenda sináptica. Assim, apenas o neurónio pós-sinático é estimulado
e os neurotransmissores que estão na fenda sináptica são rapidamente ou recapatados (reciclados) ou
degradaos por enzimas, para que não haja uma hiperestimulação.
O cérebro tem duas funções essenciais, a função eléctrica que permite o fluxo eléctrico em cada
neurónio, permitindo a propagação do impulso nervoso e a transmissão da informação em cada neurónio. E,
ainda a função química, que permite a comunicação entre os neurónios, por libertação de compostos
químicos (neurotransmissores) para a fenda sináptica.
Algumas toxinas, venenos ou até medicamentos têm como alvo os canais de Na+ sensíveis à voltagem.
Assim, como não há entrada de NA+ para dentro da célula, o impulso nervoso não é passado, pois não ocorre
a despolarização da célula.
Há vários tipos de neurotransmissores. O neurotransmissor que por excelência que é libertado na junção
neuromuscular é a acetilcolina, que é degradada por acetilcolinesterases. O glutamato é um neurotransmissor
excitatório, enquanto que o GABA é inibitório. Além destes, há ainda outros neurotransmissores, a
dopamina, a serotonina.
Metabolismo neuronal e interacção neurónio-glia
Nas células da glia há conversão de glutamato em glutamina pela glutamina sintetase, o que permite
incorporar amónia livre. Por isso, diz-se que esta conversão glutamato-glutamina é um importante
mecanismo de protecção contra à toxicidade da amónia.
Para que haja formação de neurónio GABAérgicos inibitórios, é necessário a entrada de glutamato para
o neurónio, proveniente dos astrócitos. O glutamato já dentro do neurónio é convertido a GABA pela
glutamato descarboxilase (GAD).
A Barreira Hematoencefálica (BHE ou BBB- blood-brain barrier) é composta por uma junção apertada
de células endoteliais que estão intimamente aderentes à membrana basal, o que permite que haja uma
selectividade muito apertada das moléculas que passam para o cérebro. Como esta barreira é lipofílica,
formada por membranas biológicas, permite a passagem de moléculas também lipofílicas, enquanto que as
moléculas hidrofílicas apenas atravessam por mecanismos de alta selectividade (GLUT1 – entrada de
glucose).
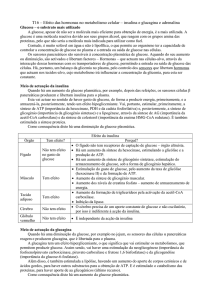
Astrócitos
Fonte de energia
Glucose
Glicogénio
Metabolismo
Anaeróbio
Vias de obtenção de energia
Reservas de Glicogénio
Isoforma dos GLUT’s
Enzimas específicas citosólicas
Enzimas específicas
mitocondriais
Glicólise
Muita da glucose é armazenada sob
a forma de glicogénio
GLUT1
Glutamina sintetase
Enzima málica
LDH5 – transforma
preferencialmente priruvato a
lactato
Piruvato carboxilase
Neuroesteróide sintetase
Glutaminase activada por fosfato
(pouco)
Neurónios
Glucose
Lactato
Alanina (estes últimos 2 provém do
piruvato em excesso dos astrócitos)
Aeróbio
Glicólise, mas sobre tudo ciclo de
Krebs e fosforalição oxidativa
Toda a glucose é consumida e nã há
formação de glicogénio
GLUT3
Colina acetiltransferase
Glutamato descarboxilase
Neuronal enolase
LDH1– transforma
preferencialmente lactato em
piruvato. Esse piruvato é depois
conduzido para o ciclo TCA
Glutaminase activada por fosfato
(muita)
Enzima málica mitocondrial
Creatina cinase de ubiquitina
mitocondrial
Jejum: apesar da hipoglicémia, ocorre transporte de glucose pela BHE, pois o cérebro não é sensível
à glucagina
Se ↓↓ [glucose]plasma o cérebro pode utilizar reservas glicogénio, contidas nos astrócitos
Se persiste ↓↓ [glucose]plasma o cérebro utiliza corpos cetónicos (β-Ho-butirato e acetoacetato)
Utilização de corpos cetónicos pelo cérebro ocorre em:
o Jejum prolongado
o Dieta rica em gorduras ou corpos cetónicos (cetogénica – método de tratamento de doenças
como epilepsia ou défices de GLUT’s)
o Diabetes mellitus
Métodos de diagnóstico de doenças cerebrais, baseados em neuroanatomia, neurometabolómica e
imagiologia funcional
Ressonância magnética
PET (positrons emisson transmisson) – permite ver o metabolismo da glucose