MEDICINA – UFPE – 119
PROFº: EDUARDO BELTRÃO
ALUNO: JOSÉ RAMOS DA COSTA JÚNIOR
OBJETIVOS – A OXIDAÇÃO DE ÁCIDOS GRAXOS
1. ESQUEMATIZE A DIGESTÃO, MOBILIZAÇÃO E TRANSPORTE DE GORDURAS NO ORGANISMO. DESTAQUE A
AÇÃO DE SAIS BILIARES, FORMAÇÃO DE QUILOMICRONS, DISTRIBUIÇÃO DE ÁCIDOS GRAXOS,
TRIACILGLICERÓIS, COLESTEROL E GLICEROL.
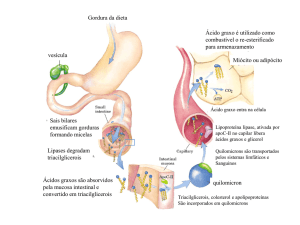
As células podem obter ácidos graxos combustíveis de três fontes: gorduras ingeridas na alimentação, gorduras armazenadas nas
células na forma de gotículas gordurosas e gorduras sintetizadas em um órgão para serem exportadas para outro.
Para serem absorvidos, através da parede intestinal, os triacilgliceróis ingeridos precisam ser convertidos de partículas gordurosas
macroscópicas insolúveis em micelas microscópicas finamente dispersas.
As gorduras ingeridas são emulsificadas pelos sais biliares no intestino delgado formando miscelas mistas de sais biliares e
triacilgliceróis.
A formação de miscelas aumenta enormemente a fração de moléculas lipídicas acessíveis à ação das lipases intestinais, que
hidrolisam os triacilgliceróis em monoacilgliceróis, diacilgliceróis, ácidos graxos livres e glicerol;
Logo em seguida eles são reconvertidos em triacilgliceróis e agrupados com o colesterol da dieta e com proteínas específicas,
formando agregados protéicos chamados quilomícrons;
Na captação de lipídios no intestino, os quilomícrons que contem a apolipoproteína CII, movem-se da mucosa intestinal para o
sistema linfático, de onde saem para a corrente sanguínea e são transportados para os músculos e para o tecido adiposo;
Nos capilares desses tecidos, a enzima extracelular lipoproteína lípase é ativada pela apo-CII. Essa enzima hidrolisa os
triacilgliceróis em ácidos graxos mais glicerol;
Em seguida eles são captados pelas células dos tecidos-alvo.
Nos músculos, os ácidos graxos são oxidados para obtenção de energia; no tecido adiposo, eles são reesterificados e armazenados
como triacilgliceróis;
2. CONCEITUE APOLIPOPROTEÍNAS.
São proteínas existentes no sangue que se ligam aos lipídios; elas são responsáveis pelo transporte dos triacilgliceróis, fosfolipídios,
colesterol e ésteres de colesterol. Elas podem se combinar com vários tipos de lipídios para formar varias classes de partículas
lipoprotéicas, estas são agregados esféricos com lipídios hidrofóbicos no centro e na superfície cadeias laterais protéicas hidrofílicas.
As várias combinações possíveis produzem partículas de densidades diferentes como os quilomícrons, as proteínas de densidade muito
baixa (VLDL) e as de densidade muito alta (VHDL).
3.
EXPLIQUE A AÇÃO DO GLUCAGON E DA EPINEFRINA NO PROCESSO DE MOBILIZAÇÃO DE
TRIACILGLICERÓIS ARMAZENADOS.
Os hormônios glucagon e epinefrina, secretados em resposta a níveis baixos de glicose no sangue, ativam a adenilato ciclase na
membrana plasmática do adipócito aumentando a concentração intracelular de um segundo mensageiro o cAMP. Por sua vez, uma
proteína quinase, dependente de cAMP fosforila e, assim, ativa a lipase de triacilgliceróis hormônio-sensível, a qual catalisa a
hidrolise de ligações de ésteres dos triacilgliceróis. Os ácidos graxos assim liberados passam do interior do adipócito para o sangue,
onde se ligam a proteína albumina ou soroalbumina. Ligados a essa proteína solúvel, eles são transportados para os tecidos como o
músculo esquelético, o coração e o córtex renal. Neles os ácidos graxos dissociam-se da albumina e difundem-se para o citosol das
células nas quais servirão como combustível. O glicerol liberado pela ação da lípase é fosforilado pela glicerol quinase e resulta em
glicerol-3-fosfato, que é oxidado em diidroxiacetona fosfato. A enzima glicolítica triose fosfato isomerase converte esse composto em
gliceraldeído-3-fosfato.
4. EXPLIQUE A ATIVAÇÃO E TRANSPORTE DOS ÁCIDOS GRAXOS PARA A MITOCONDRIA.
Os ácidos graxos livres que, provindos do sangue, entram no citosol das células não podem passar diretamente para o interior das
mitocôndrias, através de suas membranas. Eles para chegarem lá, passam por três reações enzimáticas.
: a primeira é catalisada pela família de isoenzimas presentes na membrana mitocondrial externa, as acil-CoA sintetases, que
promovem a reação geral:
ácido graxo + CoA + ATP acil-CoA graxo + AMP + 2Pi
A acil-CoA sintetase catalisa a formação de um ligação tioéster entre o grupo carboxila do ácido graxo e o grupo tiol da coenzima A
para liberar um acil-CoAgraxo (ao mesmo tempo o ATP sofre clivagem em AMP e PPi);
Os acil-CoA graxos da mesma forma como os acetil-CoA são compostos de alta energia. O pirofosfato formado na reação de ativação
é imediatamente hidrolisado por uma segunda enzima, uma pirofosfatase inorgânica, que força a reação de ativação precedente na
direção de formação do acil-CoA graxo.
: Os ésteres dos acil-CoA graxos formados na membrana mitocondrial externa não cruzam a membrana mitocondrial interna intacta.
Assim, o grupo acil-graxo é transientemente ligado ao grupo hidroxila da carnitina. Essa transesterificação é catalisada pela carnitina
aciltransferase I, presente na face externa da membrana interna. O éster acil-carnitina graxo cruza a membrana mitocondrial interna e
chega à matriz por difusão facilitada por meio do transportador acil-carnitina.
No terceiro e último passo do processo de entrada, o grupo acil-graxo é transferido enzimaticamente da carnitina para a coenzima A
intramitocondrial pela carnitina aciltransferase II. Ela está na face interna da membrana interna mitocondrial e regenera o acil-CoA
graxo e o libera, juntamente com a carnitina livre, na matriz mitocondrial.
5. ONDE OCORRE E QUAL A FUNÇÃO DA -OXIDAÇÃO DOS ÁCIDOS GRAXOS.
Na -oxidação os ácidos graxos sofrem a remoção oxidativa de sucessivas unidades de dois átomos de carbono na forma de acetilCoA, começando pela extremidade carboxila da cadeia do ácido graxo. A formação de cada molécula de acetil-CoA requer a ação de
desidrogenases para a remoção de 4 átomos de hidrogênio (dois pares de elétrons e 4H+). A ligação simples carbono-carbono que
conecta os grupos metileno nos ácidos graxos é relativamente estável. A seqüência de reações da -oxidação representa uma solução
para o problema de romper essas ligações.
EXPLIQUE O PROCESSO DE -OXIDAÇÃO QUANTO AOS TIPOS DE ÁCIDOS GRAXOS (saturados, insaturados,
com número par e impar de carbonos);
A -oxidação dos ácidos graxos saturados possui quatro passos básicos:
Primeiro uma desidrogenação produz uma dupla ligação entre os átomos de carbono e liberando um trans-enoil-CoA. É
catalisado por três isozimas da acil-CoA desidrogenase, e todas elas têm o FAD como grupo prostético;
No segundo passo uma molécula de água é adicionada à dupla ligação do trans-enoil-CoA para formar o estereoisômero L do hidroxiacil-CoA. Essa reação é catalisada pela enoil-CoA hidratase;
No terceiro passo o L--hidroxiacil-CoA é desidrogenado para formar -cetoacil-CoA pela ação da -hidroxiacil-CoA
desidrogenase. O NAD é o receptor de elétrons;
O quarto e último passo da oxidação dos ácidos graxos é catalisado pela acil-CoA acetiltransferase (tiolase) que promove a reação
do -cetoacil-CoA com uma molécula de coenzima A livre para romper o fragmento carboxiterminal de dois átomos de carbono do
acido graxo original na forma de acetil-CoA;
6.
A -oxidação dos ácidos graxos insaturados requer duas reações adicionais:
A maioria dos ácidos graxos dos nos triacilgliceróis e nos fosfolipídios são insaturados; essas ligações estão na configuração cis e não
podem sofrer a ação da enoil-CoA hidratase, a enzima que catalisa a adição de H2O, entretanto, por meio da ação de duas enzimas
auxiliares, a oxidação dos ácidos graxos pela seqüência descrita anteriormente pode também quebrar os ácidos graxos insaturados
comuns.
O ácido graxo vai formando acetil-CoA até chegar junto de onde está a dupla ligação, então entra a enoil-CoA-isomerase que passa o
composto da forma cis para a trans e daí por diante segue normalmente a rota de -oxidação;
A -oxidação dos ácidos graxos poliinsaturados usa uma enzima redutase:
Da mesma forma, o ácido graxo vai formando acetil-CoA até chegar na dupla ligação, daí através de uma ação combinada da enoilCoA isomerase e 2,4-dienoil-CoA redutase, é permitida a reentrada desse intermediário na via normal de -oxidação.
A -oxidação dos ácidos graxos de número ímpar requer três reações extras:
O ácido graxo vai sendo oxidado normalmente na mesma via, só que quando se chega a um composto de cinco átomos de carbono,
quando ele é clivado mais uma vez nós temos: acetil-CoA e proprionil-CoA. O acetil-CoA é oxidado pelo Ciclo de Krebs, mas o
propionil-coA toma uma via enzimática incomum.
Primeiro ele é carboxilado para formar o estereoisômero D do metilmalonil-CoA pela propionil-CoA carboxilase, que requer a
biotina. Esse produto formado é enzimaticamente epimerizado formando seu estereoisômero L pela ação da metil-malonil-CoA
epimerase. Ele sofre então um rearranjo intramolecular e forma o succinil-CoA, que pode entrar no ciclo de Krebs. Esse rearranjo é
catalisado pela metilmalonil-CoA-mutase, que requer como coenzima a vitB12.
7.
APRESENTE O RENDIMENTO ENERGÉTICO COMPLETO (ATP) OBTIDOS PELA OXIDAÇÃO DE UM ÁCIDO
GRAXO E COMPARE COM A OXIDAÇÃO DE UMA MOLÉCULA DE GLICOSE.
A oxidação completa do ácido graxo palmitoil, que possui 16 átomos de carbono, gera 108 ATP:
Palmitoil-Coa + 23 O2 + 108Pi + 108ADP Côa + 108ATP + 16CO2 + 23H2O
Uma molécula de glicose ao ser oxidada completamente em CO2 e H2O é 38 ATP.
8. COMO SE FAZ O CONTROLE DA -OXIDAÇÃO?
A regulação da oxidação dos ácidos graxos é feita de maneira estrita. Ela é regulada para ocorrer apenas quando a necessidade por
energia requeira. No fígado, o acil-CoA graxo formado no citosol tem duas grandes vias abertas: 1) A -oxidação pelas enzimas da
mitocôndria ou 2) a conversão em triacilgliceróis e fosfolipídios pelas enzimas do citosol. A velocidade de transferência para o interior
das mitocôndrias dos acil-CoA graxos de cadeia longa decide qual via será tomada. O processo em três passos, por meio do qual os
grupos acil-graxos são transferidos do citosol para a matriz mitocondrial é o limitante da velocidade de oxidação dos ácidos graxos.
Uma vez que os grupos acil-graxos entram na mitocôndria, eles estão definitivamente destinados à oxidação até acetil-CoA.
A concentração do malonil-CoA, o primeiro intermediário na biossíntese citosólica dos ácidos graxos de cadeia longa a partir do
acetil-CoA, aumenta sempre que o animal é bem suprido com carboidratos, qualquer excesso de glicose é convertido em ácidos graxos
citosólicos para estocagem, na forma de triacilgliceróis. A inibição da carnitina aciltransferase I pelo malonil-CoA assegura que a
oxidação dos ácidos graxos seja diminuída sempre que o fígado tenha amplo suprimento de glicose para usar como combustível e, ao
mesmo tempo, esteja fabricando ativamente triacilgliceróis a partir dessa glicose em excesso. Duas das enzimas da -oxidação
também são reguladas por metabólicos que sinalizam o suprimento suficiente de energia. Quando a relação NADH/NAD+ está alta, a
hidroxiacil-CoA desidrogenase é inibida, além disso, altas concentrações de acetil-CoA inibem a tiolase.
9. QUANTO À SINTESE DOS ÁCIDOS GRAXOS SITUE A ETAPA PREPARATÓRIA E AS REAÇÕES REPETITIVAS.
Produção de acetil-CoA citosólico
A primeira etapa da síntese dos ácidos graxos é a transferência de unidades acetato da acetil-CoA mitocondrial, a qual é produzida por
oxidação do piruvato, degradação de ácidos graxos, corpos cetônicos, até o citosol, formando acetil CoA citosólico. A porção de
coenzima A da acetil CoA não pode atravessar a membrana mitocondrial, e somente a porção acetil é transportada ao citosol em forma
de citrato (acetil + oxalacetato). No citosol, é clivado pela citrato liase, que separa os dois. Esse processo ocorre quando a
concentração de citrato mitocondrial está elevada. Isto ocorre quando a isocitrato desidrogenase é inibida pela forte presença de ATP,
resultando num aumento da concentração de isocitrato e citrato. Uma vez que uma grande quantidade de ATP é gasta para sintetizar
os ácidos graxos, o aumento relacionado de ATP e citrato aumenta a ocorrência desta rota.
Carboxilação do acetil-CoA para formar malonil CoA
A carboxilação de acetil-CoA para formar malonil-CoA é catalisada pela acetil-CoA carboxilase e requer ATP. A coenzima é a
biotina.
As séries restantes de reações da síntese dos ácidos graxos em eucariontes consiste de um dímero, cujos monômeros possuem cada
um sete diferentes atividades enzimáticas e mais um domínio que liga-se covalentemente a uma molécula de 4´fosfopanteteína.(que transporta as unidades acetil e acil em seu grupo tiol terminal (-SH) na síntese dos ácidos graxos).
[1] uma molécula de acetato é transferida do acetil-CoA ao grupo SH do ACP;
[2] a seguir esse fragmento de dois carbonos é transferido para um resíduo de cisteina da enzima, o qual age como um sítio de
captação temporário;
[3] o ACP agora vago aceita uma unidade de malonato de três carbonos do malonil CoA;
[4] o grupo acetil ataca o grupo malonil, que perde o CO2 originalmente adicionado pela acetil CoA carboxilase. A perda de energia
livre desta descarboxilacao dirige esta reação.
[5] o grupo acetona é convertido em um álcool;
[6] uma molécula de água é removida para introduzir uma ligação dupla;
[7] ocorre uma segunda etapa de redução;
O resultado destas sete etapas é a produção de um composto de quatro carbonos cujos três carbonos terminais são completamente
saturados, permanecendo aderidos ao ACP. Estas sete etapas são repetidas, iniciando com a transferência da cadeia de quatro carbonos
do ACP à cadeia lateral periférica de cisteína [8], a ligação de uma molécula de malonato ao ACP [9] e a condensação de duas
moléculas, liberando CO2 [10]. O grupo carbonila no carbono beta é reduzido. Este ciclo se repete sete vezes, a cada vez incorporando
uma unidade de dois carbonos derivada do malonil CoA na cadeia de ácido graxo em formação. Uma vez que o ácido graxo atinja um
comprimento de 16 carbonos, o processo sintético é encerrado, produzindo uma molécula completamente saturada de palmitato.
Todos os carbonos no ácido palmítico passaram pelo malonil CoA, exceto os dois doados pela acetil CoA original, os quais são
encontrados na extremidade do grupo metila do ácido graxo.
10. EXPLIQUE A FONTE DE NADPH E ACETIL-CoA NA SÍNTESE DOS ÁCIDOS GRAXOS.
Fontes de acetil-CoA: oxidação do piruvato, degradação de ácidos graxos, corpos cetônicos ou aminoácidos.
Fonte de NADPH: a rota da hexose monofosfato é a principal fonte de NADPH para a síntese dos ácidos graxos. Dois NADPH são
produzidos para cada molécula de glicose que entra nesta rota. A conversão citosólica de malato em piruvato também produz uma
quantidade significativa de NADPH citosólico.
11. COMO SE REGULA A SÍNTESE DOS ÁCIDOS GRAXOS.
12. ESQUEMATIZE A INTERAÇÃO DA -OXIDAÇÃO COM O CICLO DE KREBS, A CADEIA TRANSPORTADORA
DE ELÉTRONS E A FOSFORILAÇÃO OXIDATIVA.