Capítulo 4
Importância da Fauna de Solo
para a Ciclagem de Nutrientes
Maria Elizabeth Fernandes Correia
Luís Cláudio Marques de Oliveira
Introdução
A degradação dos detritos vegetais e animais no solo é um
processo biológico fundamental para o ecossistema, onde o carbono é
reciclado para a atmosfera como dióxido de carbono, o nitrogênio se
torna disponível como amônia e nitrato, e outros elementos associados
(fósforo, enxofre e vários micronutrientes) assumem formas inorgânicas
e podem então ser assimilados pelas plantas (STEVENSON; COLE, 1999).
A abundância e diversidade dos organismos que compõem a teia
alimentar decompositora determinam a velocidade e a magnitude de
processos como a mineralização e imobilização desses nutrientes.
Conseqüentemente, a assimilação de nutrientes pelas plantas e a produtividade das culturas podem ser fortemente afetadas pelos organismos
do solo, mesmo quando é feita uma adubação mineral.
Miolo_Biota.pmd
77
1/12/2006, 12:32
78
Capítulo 4
Apesar de os microrganismos serem os principais responsáveis
pelo processo de mineralização dos nutrientes, é a fauna de solo que
exerce um papel de regulação das populações microbianas. A predação
seletiva de fungos e bactérias, feita especialmente pela microfauna;
a estimulação, digestão e disseminação de microrganismos ingeridos
pela macrofauna e a fragmentação dos detritos realizada pelas meso e
macrofauna interferem na decomposição da matéria orgânica e alteram a
disponibilidade de nutrientes para as plantas (CRAGG; BARDGETT, 2001).
Por sua vez, a vegetação determina em grande parte o estabelecimento da comunidade da fauna de solo. A qualidade e a quantidade do
material decíduo, especialmente em ambientes tropicais, podem afetar
as populações da macrofauna do solo, resultando em uma mobilização
diferencial dos nutrientes, com conseqüências para a ciclagem de
nutrientes e a fertilidade do solo (WARREN; ZOU, 2002).
As interações da fauna de solo com os microrganismos e a sua
ação sobre a decomposição e ciclagem de nutrientes variam entre os
diferentes grupos, sendo resultantes de características intrínsecas de
cada grupo e, por vezes, de cada espécie da fauna de solo.
A morfologia, aliada a características fisiológicas e comportamentais,
é fundamental na determinação de onde e como a fauna de solo irá
intervir na decomposição e na ciclagem de nutrientes. O tipo de
deslocamento apresentado por um animal do solo, se preponderantemente horizontal, promove a redistribuição de nutrientes e matéria
orgânica em uma determinada área, como no caso dos macroartrópodes
que vivem na serapilheira, tais como isópodes e diplópodes. Se esse
deslocamento for preferencialmente vertical, essa redistribuição ocorre
no perfil do solo, enquanto ocorre um aumento da heterogeneidade na
área como um todo, com a formação de um mosaico de pequenas
manchas de solo mais ou menos fértil. Um bom exemplo disso são os
corós (larvas de besouros escarabeídeos), que se enterram no solo e
levam consigo uma grande quantidade de matéria orgânica (BROWN
et al., 2002).
Além disso, o tamanho do organismo e a sua capacidade de
deslocamento determinam qual o alcance da sua influência sobre a
ciclagem de nutrientes. Um invertebrado que seja capaz de deslocarse apenas poucos centímetros terá a sua área de influência mais restrita.
Miolo_Biota.pmd
78
1/12/2006, 12:32
Importância da Fauna de Solo para a Ciclagem de Nutrientes
79
Esse é o caso da maioria dos animais da microfauna (diâmetro corporal
< 0,2 mm) e mesofauna (diâmetro corporal entre 0,2 e 2,0 mm), que, no
entanto, compensam essa restrição com populações muito numerosas.
No entanto, muitos invertebrados da macrofauna do solo (diâmetro
> 2,0 mm) são capazes de deslocamentos consideravelmente maiores,
de poucas dezenas de metros, como no caso de alguns diplópodes. As
minhocas geófagas muitas vezes apresentam uma grande capacidade
de deslocamento tanto horizontal quanto vertical e são capazes de
modificar todo um horizonte do solo. É o caso da minhoca geófaga
Millsonia anomala, encontrada em savanas da Costa do Marfim, que
em apenas um ano é capaz de processar 70% da matéria orgânica do
solo de 0-10 cm (LAVELLE, 1978 apud FRAGOSO et al., 1997).
Esse tipo de ação apresentada por algumas minhocas promove
uma homogeneização do solo, já que praticamente todo o solo de um
determinado estrato é modificado. No caso dos térmitas de montículo,
há também um grande deslocamento horizontal e vertical dos indivíduos,
mas como a matéria orgânica é concentrada no interior dos ninhos, há
a formação de mosaicos de solo, com pontos de elevada concentração
de nutrientes. Arshad (1982), em estudo realizado na África, constatou
que em solos com presença de térmitas, a razão C/N tende a ser mais
baixa, entre 5 e 12, nos ninhos e solos adjacentes, enquanto em solos
sem térmitas essa razão aumenta e alcança valores de 15 e até maiores
que 20.
A movimentação de um animal no interior do solo pode ocorrer
de quatro formas:
• Pela ingestão do solo, que atravessa todo o tubo digestivo e é
eliminado na forma de fezes, tendo como resultado a formação
de galerias. Essa é a estratégia utilizada pelas minhocas
geófagas e pelos térmitas humívoros e, neste caso, a fonte
alimentar é a matéria orgânica do solo.
• Pela retirada de partículas ou pequenos agregados, como
ocorre com os cupins de terra solta (e.g. Syntermes sp.) e
formigas saúvas (Atta sp.). O tamanho das partículas é determinado pelo tamanho das mandíbulas desses insetos, já que é
por meio delas que ocorre o transporte desse material. Essa
atividade é capaz de modificar pontualmente o horizonte
Miolo_Biota.pmd
79
1/12/2006, 12:32
80
Capítulo 4
superficial do solo, alterando suas características, tais como a
classe textural, a quantidade de matéria orgânica e a umidade.
• Pela pressão de perfuração, que é bem observada em diplópodes
da subclasse Chilognatha. Esses invertebrados, vulgarmente
conhecidos como gongolos ou piolhos-de-cobra, possuem uma
cápsula cefálica bastante calcificada que é pressionada para
o interior do solo com a ajuda dos duplos pares de pernas
presentes em cada segmento (HOPKIN; READ, 1992). Esse tipo
de escavação é bastante dispendiosa em termos energéticos e
se torna inviável para grandes deslocamentos. No entanto,
esses animais habitam preferencialmente a serapilheira ou
troncos em decomposição e enterram-se no solo apenas para
fugir de condições adversas ou para a troca do exoesqueleto.
• Por escavação, muito utilizada por artrópodes de um modo
geral, e que consiste na utilização das pernas, que podem ser
ou não especializadas para essa atividade. Esse tipo de
locomoção é facilitado em solos arenosos, que não oferecem
muita resistência ao deslocamento.
Ação das diferentes associações da fauna
de solo sobre a ciclagem de nutrientes
Os microrganismos são os principais agentes da atividade
bioquímica do solo, estando envolvidos diretamente em todos os
processos biológicos e influenciando processos físicos e químicos.
A fauna do solo, no entanto, pode influenciar os processos do solo por
mio de duas vias principais: diretamente, pela modificação física da
serapilheira e do ambiente do solo, e indiretamente, pelas interações
com a comunidade microbiana (GONZÁLEZ et al., 2001).
Seus efeitos diretos na ciclagem biogeoquímica ocorrem por meio
da fragmentação e incorporação ao solo de detritos vegetais,
promovendo um aumento na disponibilidade de recursos para os
microrganismos e mediando a transferência de solutos e particulados
profundamente no perfil do solo. Eles também afetam a ciclagem
biogeoquímica pelo rearranjo físico das partículas do solo, mudando a
Miolo_Biota.pmd
80
1/12/2006, 12:32
Importância da Fauna de Solo para a Ciclagem de Nutrientes
81
distribuição de tamanho de poros e, como resultado, os padrões de
infiltração e emissão de gases (BEARE et al., 1995). Os efeitos das
minhocas são universalmente conhecidos, mas o turn over do solo
promovido por formigas e cupins pode ser igualmente importante (LOBRY
DE BRUYN; CONACHER, 1990).
A fauna do solo, particularmente a macrofauna, exerce um papel
fundamental na fragmentação dos resíduos vegetais e na regulação
indireta dos processos biológicos do solo, estabelecendo interações em
diferentes níveis com os microrganismos (SWIFT et al., 1979). Em poucos
casos, essas simbioses são do tipo mutualista, sendo mais freqüentes as
simbioses acidentais, onde os microrganismos são ingeridos junto com
o solo ou serapilheira e encontram no tubo digestivo e/ou nas fezes um
hábitat favorável. O efeito dos animais sobre os microrganismos podem
tanto ser estimuladores quanto inibidores (THEENHAUS; SCHEU, 1996).
É esse balanço que promove uma regulação dos processos de
decomposição e conseqüentemente da liberação de nutrientes.
Os invertebrados do solo, por sua vez, possuem uma capacidade
enzimática limitada, restringindo-se à digestão de proteínas, lipídios e
glicídios simples. Como outras classes, a fauna de solo não é capaz de
produzir enzimas que degradem compostos como a celulose ou a lignina.
No entanto, as associações da fauna com microrganismos, tanto
decorrentes da ingestão simultânea com o alimento, ou de simbioses
mutualísticas, promovem um sinergismo no sistema de decomposição.
Se, por um lado, os microrganismos, ao serem transportados pelos
invertebrados do solo, obtêm uma maior dispersão no ambiente, os
invertebrados do solo, ao utilizarem as enzimas produzidas pelos
microrganismos, ampliam a gama de substratos energéticos a serem
explorados. Além de atuarem como reguladores da atividade
microbiana, os invertebrados do solo agem como fragmentadores do
material vegetal e engenheiros do ecossistema, modificando-o
estruturalmente (LAVELLE, 1997). De acordo com Lavelle et al.(1992),
a interação da fauna de solo com microrganismos e plantas é capaz de
modificar funcional e estruturalmente o sistema de solo, exercendo uma
regulação sobre os processos de decomposição e ciclagem de nutrientes.
Sulkkava et al. (2001) obtiveram resultados que comprovam essa
Miolo_Biota.pmd
81
1/12/2006, 12:32
82
Capítulo 4
afirmação, pois, em experimentos em áreas plantadas com Betula sp.,
os autores compararam áreas com e sem a fauna de solo e verificaram
que na presença da fauna a vegetação cresceu até três vezes mais do
que nas áreas defaunadas. Interessante também que, no mesmo
experimento, Sulkava et al. (2001) verificaram também que tanto o
C como o N foram influenciados pela presença da fauna, permitindo
inclusive uma assimilação mais rápida deste último nutriente pela
vegetação, principalmente na primeira fase de crescimento.
A influência da fauna de solo está fundamentalmente ligada ao
tipo de interação que os invertebrados estabelecem com os
microrganismos, com a conseqüente digestão de compostos orgânicos
complexos e a importância dos dejetos da fauna para a formação de
estruturas do solo. Lavelle (1997), na tentativa de identificar a magnitude
dessas interações e influências, identificou três associações de
invertebrados do solo reguladoras desses processos em diferentes escalas
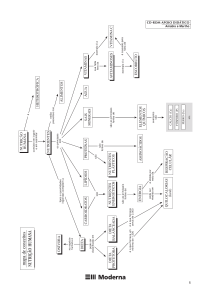
de tempo e espaço (Fig. 1). Uma associação representa um conjunto de
Fig. 1. Regulação das atividades microbianas pelas três associações de
invertebrados do solo, operando em escalas crescentes de tempo e espaço.
Fonte: Modificado a partir de Lavelle (1997).
Miolo_Biota.pmd
82
1/12/2006, 12:32
Importância da Fauna de Solo para a Ciclagem de Nutrientes
83
animais que exploram o mesmo recurso, mas o fazem de modo a
minimizar a sobreposição de nichos (COLINVAUX, 1993).
Tal conceito apresenta semelhanças com o grupo funcional, mais
amplamente difundido. No caso das associações propostas por Lavelle
(1997), esses três grupos podem ser distinguidos com base na relação
que estabelecem com os microrganismos do solo e o tipo de excremento
que produzem. Os excrementos dos invertebrados são de fundamental
importância na evolução da matéria orgânica (MARTIN; MARINISSEN,
1993), na formação e manutenção da estrutura do solo e, em longo
prazo, influenciam em processos pedológicos. As guildas propostas são:
Microteias alimentares
Compreendem a microfauna predadora de bactérias e fungos e
seus predadores. Em virtude do seu tamanho reduzido, a microfauna
não é capaz de produzir excrementos sólidos que gerem estruturas
duradouras e estáveis após a sua deposição. No entanto, esses organismos
possuem um impacto considerável na dinâmica populacional de
microrganismos e na liberação de nutrientes imobilizados na biomassa
microbiana. Esse processo é especialmente importante na rizosfera.
Colêmbolos, ácaros predadores, e até mesmo invertebrados maiores
como as minhocas podem estender essa teia alimentar por muitos outros
níveis tróficos (LAVELLE, 1997).
Transformadores de serapilheira
Compreendem a mesofauna e os grandes artrópodes, que
normalmente ingerem material orgânico puro e desenvolvem um
mutualismo externo com a microflora baseado em um “rúmen externo”
(SWIFT et al., 1979). Os artrópodes da serapilheira podem digerir parte
da biomassa microbiana ou desenvolver interações mutualísticas nos
seus excrementos. Nessas estruturas, os recursos orgânicos que foram
fragmentados e umedecidos durante a passagem pelo tubo digestivo
são ativamente digeridos pela microflora. Após alguns dias de incubação,
Miolo_Biota.pmd
83
1/12/2006, 12:32
84
Capítulo 4
os artrópodes freqüentemente reingerem esses excrementos e absorvem
os compostos orgânicos assimiláveis que foram disponibilizados pela
atividade microbiana, e ocasionalmente parte da biomassa microbiana
(HASSAL; RUSHTON, 1982). Outra importante característica dos
transformadores de serapilheira é sua habilidade de estimular a
mineralização do nitrogênio, liberando amônia, podendo ocorrer três
possíveis cenários: pode ser nitrificada e capturada pelas plantas;
temporariamente acumulada antes de sua volatilização; imobilizada
na biomassa microbiana (LAVELLE, 1997). Como exemplos de transformadores de serapilheira, serão apresentados a seguir alguns dados da
atividade de diplópodes e de larvas de besouros escarabeídeos (corós)
na decomposição de serapilheira e liberação de nutrientes para o solo.
Os diplópodes, conhecidos vulgarmente como gongolos, são um grupo
saprófago importante encontrado em todos os tipos de ecossistemas,
sejam florestais, agrícolas ou de pastagens. Os corós são bem
conhecidos como pragas agrícolas em cultivos de grãos. No entanto,
nem todas as espécies atuam como pragas, tendo, pelo contrário, um
efeito benéfico sobre a fertilidade do solo.
Diplópodes
Experimentos de laboratório demonstram que estes “transformadores de serapilheira” exibem uma preferência alimentar pela serapilheira de determinadas espécies de plantas (KHEIRALLAH, 1990).
Tal capacidade de seleção está relacionada diretamente à palatabilidade
do material e, indiretamente, à sua qualidade nutricional. A Tabela 1
mostra que a serapilheira de Eucalyptus grandis foi muito menos palatável
para duas espécies de diplópodes que a de Mimosa caesalpiniifolia,
conhecida vulgarmente como sabiá, uma leguminosa arbórea de
elevada qualidade nutricional e rápida decomposição (CORREIA;
ANDRADE, 1999). Em muitos casos, o animal morre de fome, mas
não consome materiais que apresentem elevados teores de lignina ou
polifenóis. Esse comportamento também foi observado para serapilheiras
contaminadas com elevados teores de zinco, cádmio e chumbo (READ;
MARTIN, 1990).
Miolo_Biota.pmd
84
1/12/2006, 12:32
Importância da Fauna de Solo para a Ciclagem de Nutrientes
85
Tabela 1. Consumo de serapilheira (em mg de serapilheira ingerida/g
de diplópode/dia) por duas espécies de diplópodes estudadas em
laboratório.
Serapilheira
Eucalyptus grandis (camada L1)
Eucalyptus grandis (camada F2)
Acacia mangium (camada L)
Acacia mangium (camada F)
Mimosa caesalpiniifolia (camada L)
Mimosa caesalpiniifolia (camada F)
Rhinocricus
padbergi
Trigoniulus
corallinus
13,52B
9,01B
10,46 B
16,10B
45,22A
55,47A
34,57B
27,61B
48,32B
53,31B
136,45A
237,86A
Médias seguidas de mesma letra em uma mesma coluna não diferem significativamente (p <
0,05), pelo teste de Tukey.
1
Camada superior da serapilheira composta por folhas recém-caídas e ainda sem sinais de
ataque por microrganismos e fauna de solo.
2
Camada da serapilheira subjacente à camada L e que apresenta sinais de ataque por
microrganismos e fauna de solo.
Fonte: Correia; Andrade (1999).
Existem poucas estimativas da quantidade de serapilheira consumida pelos “transformadores de serapilheira”. Em geral, esses dados
são relativos a poucas espécies e são oriundos de experimentos de
laboratório. Correia (1994) estimou que o consumo de serapilheira de
quatro espécies de diplópodes em uma área de Mata Atlântica no norte
do Espírito Santo era da ordem de 27g/ha/dia. No entanto, acredita-se
que para esse ecossistema existam mais de 20 espécies de diplópodes,
além de todos os demais grupos saprófagos da fauna de solo. Dangerfield
& Milner (1996) estimaram que o consumo de serapilheira também de
quatro espécies de diplópodes em florestas e savanas do sul da África
era respectivamente de 6% e 39% da serapilheira depositada. Os
mesmos autores verificaram que para plantações o consumo era de
menos de 1% da serapilheira depositada.
Os diplópodes mobilizam nutrientes presos na serapilheira e
enriquecem o solo com N, C, Ca, Mg, P e K, em situações de microcosmos (SMITT; VAN AARDE, 2001). Esse enriquecimento é fruto de uma
elevada capacidade de consumo de serapilheira, aliada a uma elevada
atividade microbiana nas fezes dos diplópodes. Durante a passagem
Miolo_Biota.pmd
85
1/12/2006, 12:32
86
Capítulo 4
da serapilheira pelo tubo digestivo dos diplópodes, esse material é
triturado, o que aumenta a sua superfície específica, umedecido e
enriquecido com microrganismos. Esse efeito catalisador é comparado
ao que acontece no rúmen de alguns mamíferos, com a particularidade
de estar fora do corpo do animal, mais precisamente nas suas fezes,
sendo chamado de “rúmen externo” (SWIFT et al., 1979).
Corós e fertilidade do solo
Os corós, também conhecidos como bicho-bolo, capitão ou pãode-galinha, são larvas de besouros escarabeídeos, que se caracterizam
como insetos de solo subterrâneos. Em áreas agrícolas, algumas espécies
podem atingir o status de praga, por alimentarem-se das raízes de
culturas como o trigo e a soja (GASSEN, 2000). A prática do plantio direto
promove um ambiente favorável, pois assegura alimento, microclima
propício e, como não há preparo do solo, as galerias construídas pelas
larvas não são destruídas.
O Diloboderus abderus ou coró-da-pastagem é uma dessas
espécies que podem ser pragas de lavouras e pastagens, mas que
também ganhou a denominação de praga útil, pelos benefícios que traz
à fertilidade e à física do solo (GASSEN, 2000). Amostras de solo coletadas
no perfil de solo até 25 cm e dos resíduos encontrados nas câmaras de
larvas de D. abderus, em lavouras de Santa Rosa e Giruá, RS,
demonstraram que o pH e os teores de nutrientes, matéria orgânica e
argila são semelhantes aos da camada superficial (Tabela 2).
As galerias construídas por essa e outras espécies de corós podem
atingir mais de 1 m de profundidade. Em áreas de plantio direto no
Paraná, constatou-se a existência de galerias com diâmetro de 33,5 mm
e profundidade de 117 cm (BROWN et al., 2002). Essas galerias constituem-se em canais abertos para a infiltração da água da chuva,
diminuindo o escoamento superficial que pode levar a um processo
erosivo intenso. Outras espécies de corós, como o Bothynus sp.
(conhecido como coró-da-palha) e Cyclocephala flavipennis, não
causam danos às plantas cultivadas, mesmo em densidades elevadas,
mas apresentam os mesmos efeitos benéficos de construção de galerias
e incorporação da matéria orgânica (GASSEN, 2000) (Fig. 2).
Miolo_Biota.pmd
86
1/12/2006, 12:32
87
Importância da Fauna de Solo para a Ciclagem de Nutrientes
Tabela 2. Teores de acidez (pH), alumínio (Al), cálcio (Ca), magnésio
(Mg), fósforo (P), potássio (K), matéria orgânica (M.O.) e argila no perfil
do solo e nas câmaras de D. abderus.
Profundidade
cm
0-5
5 - 10
10 - 15
15 - 20
20 - 25
Câmara
C.V.%
pH
5,6 a
5,5 a
5,4 a
4,9 b
4,7 b
5,5 a
7,1
Al
Ca
Mg
me/100 g me/100 g me/100 g
0,13
0,26
0,41
1,41
2,24
0,20
6,2
c
c
c
b
a
c
5,7 ab
5,1 b
4,4 b
2,6 c
1,8 c
6,6 a
4,9
1,9 ab
1,6 b
1,6 b
1,1 c
0,9 c
2,1 a
15,6
P
ppm
55,4
26,5
17,7
7,8
3,5
46,0
3,6
a
bc
cd
de
e
a
K
ppm
M.O.
%
194 a
3,4 b
126 ab 2,5 c
79 bc 2,3 cd
50 c
2,1 cd
33 c
2,0 cd
172 a
4,7 a
7,5 19,6
Argila
%
30,8
34,6
41,6
46,0
47,0
35,5
8,9
c
c
ab
a
a
bc
Médias seguidas da mesma letra na coluna são equivalentes por meio do teste de Tukey 95%.
Fonte: Gassen e Kochhann (1993).
Fig. 2. Larva de
Bothynus sp.
com palha
armazenada no
solo.
Fonte: Gassen
(2000).
Miolo_Biota.pmd
87
1/12/2006, 12:32
88
Capítulo 4
Engenheiros do ecossistema
Nesta guilda, encontram-se organismos da macrofauna, principalmente, minhocas e térmitas, que são grandes o bastante para
desenvolver relações mutualísticas com os microrganismos, no interior
do seu próprio tubo digestivo. Essas interações podem ser obrigatórias
(como no caso dos protozoários flagelados e dos térmitas inferiores) ou
simbioses facultativas, como as que ocorrem no tubo digestivo dos
térmitas superiores e minhocas (BREZNAK, 1984). Esses animais
geralmente ingerem uma mistura de materiais orgânicos e minerais. Os
ácidos orgânicos produzidos pela digestão, e a subseqüente incubação da
matéria orgânica nos coprólitos, são normalmente floculados na
presença de minerais de argila e elevada atividade microbiana
(LAVELLE, 1997). Os excrementos produzidos, que são grandes (na faixa
de 0,1-2 cm ou mais), podem fazer parte da estrutura dos
macroagregados e participar proeminentemente da formação de
estruturas estáveis por meio da regulação da porosidade, densidade e
outras propriedades do solo (BAL, 1982).
A atividade de cada organismo modifica o ambiente em que ele
vive, de tal forma que, em alguns casos, essas interações podem ser
extremamente importantes para a manutenção de certas espécies em
um dado ambiente, bem como o funcionamento do ecossistema
(BEGON et al., 1997). Os organismos que possuem essa capacidade
modificadora e essencial são denominados engenheiros do ecossistema.
As modificações exercidas pelos engenheiros é de tal monta que
Dawkins (1982) considerou sua ação como uma extensão do seu
fenótipo. Esses organismos contribuem de maneira essencial para a
diversidade de uma dada comunidade, criando uma gama de nichos
utilizáveis que podem ser explorados por determinadas espécies. Dessa
maneira, seu efeito primário é permitir o aumento da diversidade na
comunidade-alvo.
Jones et al. (1994) definiram os engenheiros do ecossistema como
organismos que, direta ou indiretamente, modulam a disponibilidade
de recursos para outras espécies por meio de modificações no estado
físico nos materiais bióticos e abióticos, criando e/ou mantendo hábitats
passíveis de utilização pelas espécies beneficiadas. Dois tipos básicos
Miolo_Biota.pmd
88
1/12/2006, 12:32
Importância da Fauna de Solo para a Ciclagem de Nutrientes
89
de engenheiros do ecossistema podem ser identificados de acordo com
seu modo de atuação e interação com as espécies presentes: os
engenheiros autogênicos, que modificam o hábitat com suas próprias
estruturas físicas, como por exemplo as árvores e os corais, e os
engenheiros alogênicos, que modificam o ambiente pela transformação
de materiais vivos ou não, de um estado físico em outro (JONES et al.,
1994). Pode-se dizer que esse é o caso de dois dos principais grupos da
fauna edáfica: térmitas e oligoquetos. As atividades exercidas por esses
táxons, tanto internamente, na construção de ninhos e galerias
subterrâneas e no transporte de materiais, como externamente, na
construção de ninhos epígeos que servem de abrigo e local de
reprodução para algumas espécies, alteram a estrutura edáfica de tal
forma que criam artefatos modificadores do ambiente no âmbito da
paisagem, servindo inclusive para disponibilizar e redistribuir os recursos
às outras espécies (JONES et al., 1994; REDFORD, 1984).
Térmitas
Os térmitas pertencem à ordem Isoptera dos insetos, com
aproximadamente 2.800 espécies. Esse grupo é especializado em uma
dieta à base de materiais lignocelulósicos, fato esse que os colocou
evolutivamente em contato com um recurso trófico extremamente
produtivo (MYLES, 1988). Os térmitas são capazes de fixar o nitrogênio
atmosférico a partir das bactérias anaeróbicas que habitam seu trato
digestivo (TAYASU et al., 1994), simbiose que, junto com sua superabundância, permite aos térmitas representar o papel de espécie-chave como
superdecompositor e “estabilizador do balanço carbono-nitrogênio”
(HIGASHI; ABE, 1997).
Sobre esse aspecto, os térmitas, ao se alimentarem de matéria
orgânica morta, utilizam recursos que apresentam uma relação carbononitrogênio (C/N) muito maior do que a encontrada em seus próprios
tecidos, tendo dessa forma que balancear esses inputs (HIGASHI et al.,
1992). Isso ocorre porque tanto a celulose como os constituintes da
parede celular são abundantes na matéria orgânica morta, enquanto as
substâncias citoplasmáticas são raras, o que dificulta a decomposição
desse material. Existem dois mecanismos básicos de balanceamento:
Miolo_Biota.pmd
89
1/12/2006, 12:32
90
Capítulo 4
o primeiro em que se adiciona N ao recurso alimentar e o segundo,
favorecido pela eliminação seletiva de C do recurso trófico. Os térmitas
possuem uma série de simbiontes gastrointestinais que possibilitam a
esse grupo resolver a questão da elevada razão C/N de seu recurso
trófico. Tal fato permitiu a um recurso hiperabundante (matéria orgânica
morta) tornar-se um recurso alimentar “bem protegido”, pela sua difícil
utilização e assimilação relacionada ao desbalanceamento da relação
C/N, abrindo assim um nicho que, ao longo do tempo evolutivo, foi
basicamente monopolizado pelos térmitas (HIGASHI et al., 1992).
Os mecanismos utilizados pelos térmitas para a “melhoria” na
qualidade do recurso via simbiontes intestinais podem ser divididos em
dois sistemas básicos: o primeiro, presente em algumas espécies de
térmitas, apresenta simbiontes intestinais, no caso bactérias fixadoras
de nitrogênio ou também recicladoras de N (LEE; WOOD, 1971), e o
segundo, via liberação do carbono em excesso, que pode se dar pela
liberação de gás metano (CH4) por bactérias metanogênicas simbiônticas
ou pela liberação de CO2 produzido por fungos cultivados pelos próprios
térmitas (WOOD; SANDS, 1978). As espécies utilizam de diferentes
formas esses mecanismos descritos, algumas delas utilizam vários
mecanismos de forma conjunta. Tais mecanismos têm profunda influência
na manutenção e no sucesso das colônias de térmitas, e quanto mais
diversos forem os mecanismos utilizados pela colônia para resolver a
questão do balanço C/N de seu recurso alimentar, maior será essa colônia
(HIGASHI et al., 1992). Essa capacidade de modificar o recurso
disponível a ponto de este tornar-se utilizável é uma das características
dos engenheiros do ecossistema.
Os térmitas contribuem de forma significativa para o enriquecimento do solo adjacente por meio da construção de ninhos, que
acumulam material fecal de importante qualidade, auxiliando na humificação da matéria orgânica do solo, e também pelo aumento da
atividade microbiana tanto dentro como na própria área adjacente ao
ninho, o que permite uma aceleração da ciclagem e a reabsorção dos
nutrientes pelos produtores primários (LAVELLE et al., 1997). Os nutrientes
retidos nas estruturas dos térmitas, ninhos e galerias retornam para o
solo por erosão, possibilitando que ocorra um mosaico de nutrientes na
paisagem, preservando a diversidade ecológica (LAVELLE et al., 1992).
Miolo_Biota.pmd
90
1/12/2006, 12:32
Importância da Fauna de Solo para a Ciclagem de Nutrientes
91
Esse mosaico foi relatado em inúmeros ecossistemas, inclusive o
amazônico, onde Salick et al. (1983) observaram que a capacidade
elevada de retenção e acumulação de nutrientes pelos térmitas faz com
que haja um mosaico de nutrientes no solo tipicamente pobre da região
do Rio Negro.
Como visto anteriormente, térmitas são claramente engenheiros
do ecossistema, em ambientes naturais, e, como frisou Redford (1984),
espécies-chave (keystone species), em ambientes semi-áridos. Dessa
forma, pode-se dizer que os térmitas apresentam um papel principalmente benéfico na promoção dos processos ecológicos em agroecossistemas, em detrimento do seu conhecido papel como praga agrícola.
Apesar da importância exercida pelos térmitas em ecossistemas naturais,
tanto na ciclagem de nutrientes como na modificação das características
físicas e químicas do solo, os estudos em agroecossistemas sempre
privilegiaram o aspecto dos térmitas como pragas, não obstante o fato
de que um número bastante reduzido de espécies deste grupo seja
realmente praga agrícola.
Black e Okwakol (1997) fizeram uma extensiva revisão nesse
sentido, demonstrando claramente que os térmitas podem apresentar
um papel extremamente benéfico em agroecossistemas, se estes forem
manejados de forma sustentável. Os autores afirmam que algumas
práticas agrícolas como aração, manutenção de monoculturas e utilização
de pesticidas podem influenciar negativamente as populações de
térmitas preestabelecidas, fazendo com que a diversidade diminua
drasticamente, o que possibilitaria a entrada de outras espécies, entre
estas as espécies-pragas e, mais importante, alguns processos ecológicos
dos quais os térmitas são mediadores essenciais seriam claramente
prejudicados.
Oligoquetos
Estima-se que existam por volta de 7.254 espécies de Oligoqueta,
e, destas, 3.627 são terrestres, sendo a taxa anual de descrição de novas
espécies próxima de 50, normalmente descobertas em áreas tropicais
(FRAGOSO et al., 1997).
Miolo_Biota.pmd
91
1/12/2006, 12:32
92
Capítulo 4
A ocorrência e a abundância das minhocas variam em um
gradiente termolatitudinal, sendo normalmente excluídas das áreas mais
secas, principalmente com pluviosidade abaixo de 800 mm, e, apesar
do número de espécies não se modificar muito ao longo desse gradiente,
a diversidade funcional aumenta consideravelmente (LAVELLE et al.,
1997). O substrato utilizado como alimento e/ou moradia determina os
tipos funcionais presentes e os mais comuns são as minhocas que se
alimentam de serapilheira, as que se alimentam de matéria orgânica e
as que vivem em galerias.
Quando presentes no ambiente, as minhocas podem apresentar
papel primordial nos processos ecológicos do solo. Os mecanismos de
participação nesses processos podem ocorrer a partir da ingestão de
uma mistura de partículas orgânicas e minerais, eliminando coprólitos
que contêm matéria orgânica estabilizada, por meio de um sistema de
simbiose no trato digestivo em associação com a microflora do solo,
permitindo que as minhocas se alimentem de substratos complexos, além
da criação de estruturas que acarretam modificações físicas e na
pedogênese do solo (LAVELLE, 1997).
As minhocas apresentam uma gama extensa de preferências
alimentares, mas alguns estudos realizados em diferentes plantios comprovaram que boa parte das espécies presentes em agroecossistemas é
mais atraída por substratos na forma de matéria orgânica parcialmente
decomposta (DOUBE et al., 1994).
A digestão da matéria orgânica do solo é feita pelas minhocas por
enzimas produzidas parcialmente por elas e, em parte, por associação
mutualística com a microflora ingerida. Adicionando 80% a 120% de
água ao material e 5% a 30% de muco, a atividade microbiana é estimulada
e, junto com enzimas não produzidas pelas minhocas, no caso celulase
e mananase, essas enzimas são liberadas no trato intestinal pelos
microrganismos, o que facilita a digestão de uma importante fração do
material ingerido (LAVELLE, 1997).
O resultado da digestão da matéria orgânica do solo resulta na
liberação de quantidades significativas de nutrientes assimiláveis (MARTIN
et al., 1992). Lee (1985) delineou três possibilidades básicas: a primeira,
pela liberação de coprólitos, já que o material, quando passa pelo trato
intestinal, é enriquecido por compostos nitrogenados do metabolismo
Miolo_Biota.pmd
92
1/12/2006, 12:32
Importância da Fauna de Solo para a Ciclagem de Nutrientes
93
dos oligoquetos, por meio da excreção de urina e pelos tecidos de
oligoquetos mortos, já que o peso seco do tecido desses animais é
composto de 60% a 70% de proteína com 12% de nitrogênio desse
total. Coprólitos frescos de Pontoscolex corethrurus continham 18 vezes
mais amônia do que o solo não ingerido e 1,5 vez a quantidade de nitrato.
A redução nas concentrações dos coprólitos diminui de forma
diferenciada, de acordo com o ambiente analisado, e deve estar
relacionada com a reorganização microbiana, que pode ocorrer em
diferentes velocidades, já que outras formas de liberação de nutrientes
como a lixiviação e a própria reabsorção pela vegetação foram evitadas
(LAVELLE et al., 1992). O fósforo (P), outro nutriente especialmente crítico
para o crescimento das plantas, também apresentou concentrações
altíssimas se comparadas com o solo controle, chegando até a oito vezes
a concentração no solo não ingerido.
Pashanasi et al. (1992) verificaram que a presença de minhocas
influenciou o crescimento de algumas espécies vegetais, enquanto outras
espécies não foram influenciadas, o que se supõe estar relacionado
com o tamanho e a distribuição do sistema radicular de cada planta,
pois existe uma relação direta entre tamanho do sistema radicular e a
absorção dos nutrientes liberados via coprólitos.
Jégou et al. (1998), analisando os efeitos da presença de minhocas
na transferência de carbono no solo, encontraram nas estruturas
construídas pelas minhocas e no solo adjacente aos coprólitos um
enriquecimento de C de até 40% em comparação com o solo utilizado
como controle, comprovando a importância das minhocas na
transferência de matéria orgânica do solo.
A diversidade de minhocas em agroecossistemas, se comparada
a sistemas não perturbados, é bem inferior, com a predominância de
espécies endogeicas, o que torna essas espécies extremamente
importantes para a regulação das funções edáficas, pois as mesmas
atuam como engenheiras do ecossistema, por meio de suas interações
mutualísticas com a microflora presente, ingestão seletiva de partículas
do solo e produção de coprólitos e galerias, afetando de forma
significativa e positiva a liberação de nutrientes antes indisponíveis para
a utilização da vegetação, assim como na dinâmica da matéria orgânica
e outros processos pedológicos (FRAGOSO et al., 1997).
Miolo_Biota.pmd
93
1/12/2006, 12:32
94
Capítulo 4
Referências
ARSHAD, M. A. Influence of termite Macrotermes michaelseni on soil
fertility and vegetation on semi-arid savannah ecosystem. AgroEcosystems, Amsterdam, v. 8, p. 47-58, 1982.
BAL, L. Zoological ripening of soils. Wageningen: Pudoc, 1982.
BEARE, M. H.; COLEMAN, D. C.; CROSSLEY Jr., D. A.; HENDRIX, P. F.;
ODUM, E. P. A hierarchical approach to evaluating the significance of
soil biodiversity to biogeochemical cycling. Plant and Soil, Dordrecht,
v. 170, p. 5-22, 1995.
BEGON, M.; HARPER, J. L.; TOWNSEND, C. R. Ecología: individuos,
poblaciones e comunidades. Barcelona: Ediciones Omega, 1997.
BLACK, H. I. J.; OKWAKOL, M. J. N. Agricultural intensification, soil
biodiversity and agroecosystem function in the tropics: the role of
termites. Applied Soil Ecology, Amsterdam, v. 6, p. 37-53, 1997.
BREZNAK, J. A. Biochemical aspects of symbiosis between termites and
their intestinal microbiota. In: ANDERSON, J. M.; RAYNER, A. D. M.;
WALTON, D. W. H., (Ed.). Invetebrate-microbial interactions.
Cambridge: Cambridge University Press, 1984. p. 173-224.
BROWN, G. G.; ALBERTON, O.; BRANDÃO Jr., O.; SARIDAKIS, G. P.;
TORRES, E. Scarab beetle-grub holes in various tillage and crop systems
at Embrapa Soybean, Londrina, Brazil. In: INTERNATIONAL
TECHNICAL WORKSHOP ON BIOLOGICAL MANAGEMENT OF SOIL
ECOSYSTEMS FOR SUSTAINABLE AGRICULTURE, 2002, Londrina, PR.
Program, abstracts and related documents. Londrina: Embrapa Soja,
2002. p. 91. (Embrapa Soja. Documentos, 182).
COLINVAUX, P. Ecology 2. New York: John Wiley & Sons, 1993.
CORREIA, M. E. F. Organização da comunidade de macroartrópodos
edáficos em um ecossistema de Mata Atlântica de Tabuleiros, Linhares
(ES). 1994. 76 p. Dissertação (Mestrado em Ecologia) – Universidade
Federal do Rio de Janeiro, Rio de Janeiro.
Miolo_Biota.pmd
94
1/12/2006, 12:32
Importância da Fauna de Solo para a Ciclagem de Nutrientes
95
CORREIA, M. E. F.; ANDRADE, A. G. Formação de serapilheira e
ciclagem de nutrientes. In: SANTOS, G. A. & CAMARGO, F. A. O., (Ed.).
Fundamentos da matéria orgânica do solo: Ecossistemas tropicais e
subtropicais. Porto Alegre: Gênesis, 1999. p. 197-225.
CRAGG, R. G.; BARDGETT, R. How changes in soil faunal diversity
and composition within a trophic group influence decomposition
processes. Soil Biology and Biochemistry, Oxford, v. 33, p. 2073-2081,
2001.
DANGERFIELD, J. M.; MILNER, A. E. Millipede fecal pellet production
in selected natural and managed habitats of southern Africa: implications
for litter dynamics. Biotropica, Lawrencem v. 28, p. 113-120, 1996.
DAWKINS, R. The extended phenotype. Oxford: Oxford University
Press, 1982.
DOUBE, B. M.; STEPHENS, P. M.; DAVOREN, C. W.; RYDER, M. H.
Earthworms and the introduction and management of beneficial soil
microorganisms. In: Pankhurst, C. E.; Doube, B. M.; GUPTA, V. S. S. R.;
GRACE, P. R., (Ed.). Soil biota: Management in sustainable farming
systems. Western: CSIRO, 1994. p. 32-41.
FRAGOSO, C.; BROWN, G.; BLANCHAERT, E.; LAVELLE, P.;
PASHANASI, B.; SENAPATI, B.; KUMAR, T. Agricultural intensification,
soil biodiversity and agroecosystem function in the tropics: the role of
earthworms. Applied Soil Ecology, Amsterdam, v. 6, p. 17-35, 1997.
GASSEN, D. Os escarabeídeos na fertilidade de solo sob plantio direto.
In: REUNIÃO BRASILEIRA DE FERTILIDADE DO SOLO E NUTRIÇÃO
DE PLANTAS, 24., REUNIÃO BRASILEIRA SOBRE MICORRIZAS, 8.,
SIMPÓSIO BRASILEIRO DE MICROBIOLOGIA DO SOLO, 6., REUNIÃO
BRASILEIRA DE BIOLOGIA DO SOLO, 3., oct. 2000, Santa Maria-RS.
Biodinâmica do solo... Santa Maria: SBCS, SBM, 2000. 216 p. FERTBIO
2000. CD ROM.
GASSEN, D.; KOCHHANN, R. A. Diloboderus abderus: benefícios de
uma praga subterrânea no sistema de plantio direto. In: ENCONTRO
Miolo_Biota.pmd
95
1/12/2006, 12:32
96
Capítulo 4
LATINO AMERICANO SOBRE PLANTIO DIRETO NA PEQUENA
PROPRIEDADE, 1., Ponta Grossa, PR, 1993. Anais... Ponta Grossa:
IAPAR, 1993. p.101-107.
GONZÁLEZ, G.; LEY, R. E.; SCHMIDT, S. K.; ZOU, X.; SEASTEDT, T. R.
Soil ecological interactions: comparisons between tropical and subalpine
forests. Oecologia, New York, v. 128, p. 549-556, 2001.
HASSAL, M.; RUSHTON, S. P. The role of coprophagy in the feeding
strategies of terrestrial isopods. Oecologia, New York, v. 53, p. 374-381,
1982.
HIGASHI, M.; ABE, T. Global diversification of termites driven by the
evolution of symbiosis and sociality. In: ABE, T.; LEVIN, S. A.; HIGASHI,
M., (Ed.). Biodiversity, an ecological perspective. New York: SpringerVerlag, 1997. p. 83-112.
HIGASHI, M.; ABE, T.; BURNS, T. P. Carbon-nitrogen balance and termite
ecology. Proceedings of the Royal Society of London, Série B, Biological
Sciences, London, v. 249, p. 303-308, 1992.
HOPKIN, S. P.; READ, H. J. The biology of millipedes. New York: Oxford
University Press, 1992. 233 p.
JÉGOU, D.; CLUZEAU, D.; BALESDENT, J.; TRÉHEN, P. Effects of four
ecological categories of earthworms on carbon transfer in soil. Applied
Soil Ecology, Amsterdam, v. 9, p. 249-255, 1998.
JONES, C. G., LAWTON, J. H.; SHACHAK, M. Organisms as ecosystems
engineers. Oikos, Copenhagen, v. 69, p. 373-386, 1994.
KHEIRALLAH, A. M. Fragmentation of leaf litter by a natural population
of the millipede Julus scandinavus (Latzel 1884). Biology and Fertility
of Soils, Berlin, v. 10, p. 202-206, 1990.
LAVELLE, P. Faunal activities and soil process: strategies that determine
ecosystem function. Advances in Ecological Research, London, v. 37,
p. 93-132, 1997.
Miolo_Biota.pmd
96
1/12/2006, 12:32
Importância da Fauna de Solo para a Ciclagem de Nutrientes
97
LAVELLE, P.; BIGNELL, D.; LEPAGE, M.; WOLTERS, V.; ROGER, P.;
INESON, P.; HEAL, O. W.; DHILLION, S. Soil function in a changing
world: the role of invertebrate ecosystems engineers. European Journal
of Soil Biology, Oxford, v. 33, n. 4, p. 159-193, 1997.
LAVELLE, P.; BLANCHART, E.; MARTIN, A.; SPAIN, A. V.; MARTIN, S. V.
Impact of soil fauna on the properties of soils in the humid tropics. In: LAL,
R.; SANCHEZ, P. A., (Ed.). Myths and science of soils of the tropics.
Madison: Soil Science Society of America/American Society of
Agronomy, 1992. p. 157-185. (SSSA Special Publication, 29).
LEE, K. E. Earthworms: their relationships with soils and land use. London:
Academic, 1985. 411 p.
LEE, K. E.; WOOD, T. G. Termites and soils. London: Academic, 1971.
LOBRY DE BRUYN, L. A.; CONACHER, A. J. The role of termites and
ants in soil modification: A review. Australian Journal of Soil Research,
Rockville, v. 28, p. 55-93, 1990.
MARTIN, A.; MARINISSEN, J. C. Y. Biological and physico-chemical
processes in excrements of soil animals. Geoderma, Amsterdam, v. 56,
p. 331-347, 1993.
MARTIN, A.; MARIOTTI, A.; BALESDENT, J.; LAVELLE, P. Soil organic
matter assimilation by a geophagus tropical earthworm based on d13 C
measurements. Ecology, Washington, v. 73, p. 118-128, 1992.
MYLES, T. G. Resource inheritance in social evolution from termites to
man. In: C.N. SLOBODCHIKOFF, C. N., (Ed.). The ecology of social
behaviour. San Diego: Academic, 1988. p. 379-423.
PASHANASI, B.; MELENDEZ, G.; SZOTT, L.; LAVELLE, P. Effect of
inoculation with the endogeic earthworm Pontoscolex corenthrurus
(Glossoscolecidae) on N availability, soil microbial biomass and the
growth of three tropical fruit tree seedlings in a pot experiment. Soil
Biology and Biochemistry, Oxford, v. 24, p. 1655-1660, 1992.
Miolo_Biota.pmd
97
1/12/2006, 12:32
98
Capítulo 4
READ, H. J.; MARTIN, M. H. A study of millipede communities in
woodlands contaminated with heavy metals. In: PROCEEDINGS OF
THE 7TH INTERNATIONAL CONGRESS OF MYRIAPODOLOGY, 7.,
Leiden, 1990. Proceedings... Leiden: E. J. Brill, 1990. p. 289-298.
REDFORD, K. H. The termitaria of Cornitermes cumulans (Isoptera:
Termitidae) and their role in determining a potential keystone species.
Biotropica, Lawrence, v. 16, p. 112-119, 1984.
SALICK, J.; HERRERA, R.; JORDAN, C. R. Termitaria: nutrient patchiness
in nutrient deficient rain forests. Biotropica, Lawrence, v. 15, p. 1-7,1983.
SMITT, A. M.; VAN AARDE, R. J. The influence of millipedes on selected
soil elements: a microcoesm study on three species occurring on coastal
sand dunes. Functional Ecology, Oxford, v. 15, p. 51-59, 2001.
STEVENSON, F. J.; COLE, M. A. Cycles of soil: Carbon, nitrogen,
phosphorus, sulfur, micronutrients. New York: John Wiley & Sons, 1999.
427 p.
SULKKAVA, P.; HUHTA, V.; LAAKSO, J.; GYLÉN, E. R. Influence of soil
fauna and habitat patchiness on plant (Betula pendula) growth and carbon
dynamics in a microcosm experiment. Oecologia, New York, v. 129,
p. 133-138, 2001.
SWIFT, M. J.; HEAL, O. W.; ANDERSON, J. M. Decomposition in
terrestrial ecosystems. Oxford: Blackwell, 1979. 372 p. (Studies in
Ecology, 5).
TAYASU, I.; SUGIMOTO, A.; WADA, E.; ABE, T. Xylophagus termites
depending on atmospheric nitrogen. Naturwissenschaften, New York,
v. 81, p. 229-231, 1994.
THEENHAUS, A.; SCHEU, S. Successional changes in microbial biomass
activity and nutrient status in faecal material of the slug Arion rufus
(Gastropoda) deposited after feeding on different plant materials. Soil
Biology and Biochemistry, Oxford, v. 28, p. 569-577, 1996.
Miolo_Biota.pmd
98
1/12/2006, 12:32
Importância da Fauna de Solo para a Ciclagem de Nutrientes
99
WARREN, M. W.; ZOU, X. Soil macrofauna and litter nutrients in three
tropical tree plantations on a disturbed site in Puerto Rico. Forest Ecology
and Management, Amsterdam, v. 170, p. 161-171, 2002.
WOOD, T. G.; SANDS, W. A. The role of termite in ecosystems.
In: BRIAN, M. V., (Ed.). Production ecology of ants and termites.
Cambridge: Cambridge University Press, 1978. p. 245-292.
Miolo_Biota.pmd
99
1/12/2006, 12:32
Miolo_Biota.pmd
100
1/12/2006, 12:32