RESUMÃO DE BIOLOGIA
METABOLISMO ENERGÉTICO
Metabolismo celular é um conjunto de reações químicas, sejam de
síntese [anabolismo] ou de quebra [catabolismo], voltadas para a
transformação de energia em uma célula. A fonte de energia mais
importante para os seres vivos é a luz solar, que é captada pelos seres
clorofilados e transformada em energia química, sendo armazenada em
moléculas orgânicas como a glicose.
•
•
Fotossíntese: síntese de glicose a partir de gás carbônico [
]e
água [
] com presença de luz.
Respiração aeróbia: quebra de moléculas orgânicas como a
glicose a partir de
.
Os seres autótrofos conseguem sintetizar o seu alimento a partir de
moléculas inorgânicas simples, por isso também são chamados de
produtores [produzem energia química], constituindo a base da cadeia
alimentar.
Os seres heterótrofos não conseguem sintetizar o próprio alimento,
precisando alimentar-se dos produtores para adquirir energia, sendo,
por isso, também chamados de consumidores [seja primário,
secundário, terciário etc.].
Apesar de a fotossíntese e a respiração serem os processos mais
importantes de transformação de energia nos seres vivos, eles não são
os únicos.
•
•
Quimiossíntese: síntese de moléculas orgânicas a partir de
e
na ausência de luz.
Fermentação: quebra parcial de moléculas orgânicas como a
glicose na ausência de
.
Os processos de fotossíntese e de quimiossíntese são denominados
processos de incorporação de energia.
Os processos de respiração e de fermentação são denominados
processos de liberação de energia.
A energia é assimilada, armazenada e liberada através de reações
químicas, processos de transformação de moléculas [reagentes] em
outras [produtos], em meio aquoso, podendo ou não ocorrer
espontaneamente.
As reações químicas que liberam energia são chamadas exergônicas.
Nessas reações, os reagentes possuem mais energia que os produtos, e
parte dessa energia é liberada no decorrer da reação na forma de calor
[reações exotérmicas].
Em geral as reações exergônicas são espontâneas. Porém, as reações
exergônicas que ocorrem nas células não são tão espontâneas assim.
Para ocorrer, elas precisam de um estímulo chamado de energia de
ativação. A maioria dessas reações é mediada por enzimas, que
reduzem a energia de ativação necessária para a ocorrência da reação.
+ 6
→ 6
+ 6
+
[
Í
]
Os principais processos celulares de liberação de energia dos
alimentos, a respiração e a fermentação, ocorrem através de
reações exergônicas.
Todavia, existe outra categoria de reações químicas que precisam
receber muita energia para ocorrer. Essas reações são chamadas
endergônicas e, nesses casos, os reagentes têm menos energia que os
produtos. Assim, há necessidade de incorporação de energia.
6
+ 6
+
[
]→
+ 6
Os principais processos celulares de incorporação de energia,
representados pela fotossíntese e pela quimiossíntese, são
endergônicos.
NOTA: reações exergônicas e endergônicas diferem quanto ao
significado de reações exotérmicas e endotérmicas, respectivamente. As
reações exergônicas e endergônicas dependem da variação da energia
livre de Gibbs.
A energia livre de Gibbs [∆ ] é uma grandeza termodinâmica definida
como o balanço entre variação de entalpia [∆ ] e a temperatura [ ]
vezes a variação de entropia [∆ ] em uma reação.
∆ =∆ −#∙∆
Onde:
∆ é uma função de estado chamada de variação de entalpia, que
informa a variação de energia em pressão constante.
# é a temperatura, uma grandeza física intensiva que é influenciada ou
sofre influência das variações energéticas durante a movimentação das
partículas.
∆ é a variação de entropia, uma função de estado que informa a
variação de energia em função do grau de liberdade das partículas.
O ATP é um nucleotídeo em que a adenina é a base, e o açúcar é a
ribose. O nucleosídeo adenina + ribose é chamado de adenosina. A
união de adenosina com três radicais fosfato leva ao composto
adenosina trifosfato, ATP, responsável por transportar energia em
reações exergônicas e endergônicas. As ligações que mantêm o segundo
e o terceiro radical fosfato presos no ATP são altamente energéticas
[liberam cerca de 7 Kcal/mol de molécula orgânica].
Durante os processos de síntese e degradação de moléculas orgânicas
nas células, existem muitas reações em que há liberação de hidrogênio,
que é transportado principalmente por duas substâncias: o %&'( e o
)&', produzidos a partir de vitaminas do complexo B.
Nessas reações, entregam para outras substâncias os átomos de
hidrogênio que carregam e voltam a ficar disponíveis na célula. As
reações em que ocorrem transferências de elétrons são conhecidas
como reações de oxirredução.
Os elementos que ganham elétrons ficam reduzidos e os que
perdem, oxidados.
%&'( [) *+& ,-'&'&] ↔ %&' + %&'2( [) *+& ,-'&'&] ↔ %&'2 +
)&'[) *+& ,-'&'&] ↔ )&'
(
[) *+&*/'01-'&]
(
[) *+&*/'01-'&]
[) *+&*/'01-'&]
O NAD+ [nicotinamida adenina dinucleotídeo] e o FAD [flavina
adenina dinucleotídeo] são nucleotídeos associados respectivamente
às vitaminas nicotinamida [B3] e riboflavina [B2]. Tanto o NAD+ quanto
o FAD participam da respiração celular, mas somente o NAD+ está
presente nos processos de fermentação.
FOTOSSÍNTESE
A fotossíntese é um processo autotrófico realizado por seres
eucariontes e procariontes clorofilados: plantas, bactérias e
cianobactérias. Nos eucariontes, a fotossíntese ocorre no interior da
célula, no cloroplasto.
Acredita-se que os cloroplastos eram cianobactérias primitivas
englobadas por células eucariontes ao longo da evolução [organela com
DNA].
O cloroplasto é formado pelo envelope [uma membrana externa e
outra interna], cujo exterior é liso e o interior dobra-se formando
vesículas internas, os tilacóides, que armazenam clorofila nos
complexos de antena. Chama-se de granum o conjunto de tilacóides
empilhados, e de grana o conjunto de granuns de um cloroplasto. O
estroma é a região compreendida entre os tilacóides e o envelope, sendo
onde se encontra os ribossomos e o material genético.
Todas as células fotossintetizantes, exceto as bacterianas, contêm 2
tipos de clorofila, e um deles sempre é a clorofila A. O segundo tipo de
clorofila geralmente é a clorofila B [nos vegetais superiores] ou
a clorofila C [nas algas].
Os diversos tipos de clorofila diferem quanto à faixa do espectro da
luz visível na qual cada uma delas capta luz com mais eficiência.
É possível verificar, analisando o gráfico, que ambas as clorofilas
possuem dois picos de absorção: um mais elevado, na faixa do violeta, e
outro menor, na faixa do vermelho.
Todos os cloroplastos das plantas atuais são formados a partir de
organelas indiferenciadas, chamadas problastídeos. Existem três tipos
de plastos formados:
PLASTOS
CLOROPLASTO
FUNÇÃO
Possuem
clorofila;
realizam
fotossíntese.
Com carotenoides; conferem cor
aos frutos.
Apigmentados; armazenam amido.
CROMOPLASTO
LEUCOPLASTO
A fotossíntese obedece a seguinte equação geral:
+ 2
+
4+
5
→
+ 2
Em que A pode ser enxofre [no caso das bactérias púrpuras] ou
oxigênio [no caso dos demais clorofilados]. Apesar de ser representada
por uma equação geral, a fotossíntese ocorre a partir de uma série de
reações químicas organizadas tradicionalmente em duas etapas
interligadas:
•
•
Etapa fotoquímica ou fase clara, em que há participação direta
da luz, ocorrendo nos tilacóides do cloroplasto.
Etapa química ou fase escura, em que não há participação
direta da luz, ocorrendo no estroma do cloroplasto.
Na etapa fotoquímica, dois conjuntos básicos de rações acontecem em
presença de luz: fotofosforilação e fotólise da água. Esta fase ocorre
na membrana dos tilacóides e dela participam um complexo de
pigmentos existente nos grana [fotossistemas I e II], aceptores de
elétrons, moléculas de água e luz. Como resultado desta fase tem-se a
produção de oxigênio, ATP [a partir de ADP + Fosfato (P)] e também a
formação de NADPH2.
Cada complexo de antena é capaz de captar a energia luminosa por
meio dos pigmentos fotossintetizantes e conduzi-la até um centro de
reação, onde essa energia pode ser aproveitada. Esse centro é formado
por um par especial de clorofilas A, cujas propriedades diferem das
demais clorofilas.
Nota: no fotossistema I, esse par de clorofilas recebe o nome de P700,
pois absorvem mais eficientemente feixes de luz com 700 nm de
comprimento de onda. No fotossistema II, esse par de clorofilas recebe
o nome de P680, pois absorve mais eficientemente feixes de luz com 680
nm de comprimento de onda.
Quando clorofilas P700 são excitadas
saem da molécula e são transferidos
repassam os elétrons de um para
transportadora de elétrons. Dentre
estão os citocromos.
pela energia luminosa, elétrons
para aceptores. Estes aceptores
outro, formando uma cadeia
as substâncias transportadoras
Nestas transferências os elétrons vão liberando energia, que é
aproveitada para a síntese de ATP a partir de ADP e fosfato. Ao chegar
ao último aceptor da cadeia, os elétrons têm nível energético
suficientemente baixo e retornam ao par de clorofilas P700. Como os
elétrons retornam para o par de clorofilas do fotossistema, fala-se em
fotofosforilação cíclica.
Entretanto, existe outro tipo de fotofosforilação, em que os elétrons das
moléculas de clorofila P700 do fotossistema I, excitados pela luz, são
captados por um aceptor, mas em vez de passarem pela cadeia, são
captados pelo NADP+, formando NADPH, e não retornam para o centro
de ração. Esses elétrons são repostos por outros provenientes do
fotossistema II [o par de clorofilas P680].
Excitado pela energia luminosa, esse par libera elétrons para uma
cadeia de aceptores [citocromos], onde a energia liberada é usada para
a síntese de ATP. Ao final dessa sequência, os elétrons desgastados ao
invés de retornarem ao par P680, vão para o par P700, que se recompõe.
Agora, o par de clorofilas deficiente corresponde ao P680, que é
restabelecido pelos elétrons provenientes da quebra da molécula de
água em presença de luz [fotólise da água]. Como os elétrons não
retornam aos centros de reação dos quais saíram, fala-se em
fotofosforilação acíclica.
Na fotólise da água, os elétrons são captados pelo P680 ao mesmo tempo
em que os hidrogênios são captados pelo NADP+, formando NADPH2, e o
O2 é expelido da célula para o meio externo, sustentando toda forma de
vida aeróbia do planeta.
Na etapa química, a energia contida nos ATP e os hidrogênios dos
NADPH2, serão utilizados para a construção de moléculas de glicose. A
síntese de glicose ocorre durante um ciclo de reações chamado ciclo
das pentoses ou ciclo de Calvin-Benson. Durante o ciclo, moléculas
de CO2 unem-se umas as outras formando cadeias carbônicas que
levam à produção de glicose. A energia necessária para o
estabelecimento das ligações químicas ricas em energia é proveniente
do ATP e os hidrogênios que promoverão a redução dos CO2 são
fornecidos pelos NADPH2.
O CO2 entra no ciclo de Calvin e se une a um composto de 5 carbonos,
a ribulose bifosfato [RuBP], que já existe no estroma do cloroplasto. A
esse processo dá-se o nome de fixação do carbono. O produto de seis
carbonos é instável e imediatamente hidrolisado pela ação de uma
enzima chamada rubisco, a mais abundante na natureza. Pela ação do
rubisco, formam-se duas moléculas de 3 carbonos, o ácido
fosfoglicérico ou fosfoglicerato [PGA].
A seguir, com o fornecimento de energia de moléculas de ATP e a ação
do NADPH, forma-se a partir do PGA o gliceraldeído fosfato ou PGAL,
que também tem 3 carbonos. Parte das moléculas de PGAL formadas
irão recompor a RuBP, reiniciando o ciclo, e a outra parte irá para o
citosol ou hialoplasma da célula, onde será sintetizada em glicose e
outros carboidratos utilizados na respiração celular.
Por depender dos produtos da fase clara, que por sua vez depende
da energia luminosa, diz-se que a fase escura depende
indiretamente da luz solar.
QUIMIOSSÍNTESE
A quimiossíntese é um processo restrito a certas bactérias autótrofas
em que a energia utilizada na formação de compostos orgânicos, a
partir de gás carbônico [CO2] e água [H2O], provém da oxidação de
substâncias inorgânicas. Dentre essas substâncias destacam-se:
hidrogênio, amônio, nitrito, sulfeto de hidrogênio, tiossulfato, ferro
e manganês.
A quimiossíntese é realizada por:
Ferrobactérias: utilizam a energia química proveniente da oxidação de
compostos de ferro para a síntese de matérias orgânica.
Nitrobactérias: utilizam a energia química proveniente da oxidação de
7
íons amônio [% (
] para a síntese de matéria
6 ] ou íons nitrito [%
orgânica. As nitrobactérias ou bactérias nitrificantes existem livres no
solo e são de grande importância no ciclo do nitrogênio. As
nitrobactérias Nitrosomonas transformam o amônio em nitrito [% 7 ], e
as nitrobacter transformam o nitrito em nitrato [% 7
8 ], que pode ser
aproveitado pelas plantas como fonte de nitrogênio para a síntese de
proteínas, ácidos nucleicos e ATP.
Sulfobactérias: utilizam a energia química gerada na oxidação de
compostos inorgânicos de enxofre para a síntese de matéria orgânica.
Esses organismos vivem em ambientes que apresentam altas
concentrações de sulfeto de hidrogênio [H2S] ou outros compostos de
enxofre oxidáveis, como o sulfito [ 9 7 ].
RESPIRAÇÃO
A respiração celular baseasse na reação de combustão completa da
glicose, na presença de oxigênio como comburente, resultando água,
gás carbônico e energia. A respiração aeróbia é realizada por muitos
procariontes, protistas, fungos e pelas plantas e animais. O processo é
comumente dividido em duas fases, uma anaeróbia [onda não há
dependência de , constituída pela glicólise] e outra aeróbia [onde há
dependência de
, constituída pelo ciclo de Krebs e pela cadeia
respiratória].
Nos seres eucariontes, a fase anaeróbica da respiração ocorre no citosol,
enquanto que a fase aeróbica ocorre no interior da mitocôndria. A
mitocôndria é delimitada por uma dupla camada lipoproteica, de face
externa lisa, e face interna com invaginações, formando as cristas
mitocondriais, que constituem a matriz mitocondrial.
A glicólise da respiração é igual à da fermentação e ocorre no
citoplasma ou hialoplasma. Na glicólise, a glicose [
] é quebrada
em duas moléculas de piruvato [ácido pirúvico], cada uma com três
carbonos [ 9
9 ], com saldo de duas moléculas de ATP. Todavia, na
degradação da glicose em piruvato, perde-se quatro átomos de
hidrogênio, que se combinam dois a dois, com moléculas de
nicotinamida adenina dinucleotídeo [ :( ], formando duas
moléculas de %&' .
A glicólise é um processo exergônico, no qual há liberação de energia
que é utilizada na síntese de ATP a partir de ADP [adenosina difosfato]:
cada molécula de glicose quebrada libera energia para formar quatro
moléculas de ATP. Entretanto, para a própria ativação da glicose no
início do processo gasta-se duas moléculas de ATP. Portanto, diz-se que
o saldo enérgico da glicólise é de dois ATP. As duas moléculas de
piruvato, juntas, formam 6 carbonos e 6 oxigênios, como na glicose,
mas formam somente 8 hidrogênio, contrastando com os 12 hidrogênios
da glicose. Os 4 hidrogênios que sobram são captados então por duas
moléculas de %&'( , formando duas moléculas de %&' .
As moléculas de piruvato [ácido pirúvico] resultantes da degradação da
glicose penetram no interior das mitocôndrias, onde ocorrerá a
respiração propriamente dita, iniciando a fase aeróbia da respiração.
Ao entrar na mitocôndria, antes mesmo de começar o ciclo de Krebs,
cada piruvato reage com uma substância chamada coenzima A e sofre
uma primeira descarboxilação [perde
para o meio] e uma primeira
desidrogenação [perde
para o meio, formando
: ], originando o
acetil – coenzima A [2C] que entrará no ciclo de Krebs ou ciclo do
ácido cítrico.
O ciclo de Krebs nada mais é do que um conjunto cíclico de reações
químicas que ocorrem na matriz mitocondrial, onde o acetil –
coenzima A será transformado em citrato [6C], cetoglutarato [5C],
succinato [4C], fumarato [4C], malato [4C] e, finalmente, ácido
oxalacético [4C]. Todavia, o importante é compreender que o ciclo vai
moer a molécula de piruvato, liberando carbono que se ligará ao
oxigênio, formando o gás carbônico, e liberando hidrogênios, que se
ligarão ao %&' e ao )&', formando %&'
e )&' .
Logo no início do ciclo, cada molécula de acetil - CoA reage com uma
molécula de ácido oxalacético, resultando em citrato [ácido cítrico – daí
o nome do ciclo] e coenzima A.
Nota: por se tratar de uma enzima, a coenzima A não se desgasta no
decorrer da reação, começando-a e terminando-a intacta.
O citrato então será degradado, perdendo carbonos no decorrer do ciclo
de Krebs, até terminar na forma de ácido oxalacético, na qual reagirá
com uma molécula acetil-CoA, reiniciando o ciclo. Para cada volta no
ciclo de Krebs, são liberados uma molécula de ATP, duas moléculas de
e 4 hidrogênios [que são captados pelo
: e pelo 5 :, formando
:
e 5 : ]. Mas, como na glicólise são produzidos dois piruvato,
em cada ciclo de Krebs são formadas duas moléculas de ATP, três
moléculas de
: , uma molécula de 5 :
e quatro moléculas de
.
O destino dos hidrogênios liberados na glicólise e no ciclo de Krebs
é um ponto crucial no processo de obtenção de energia na
respiração aeróbica.
Até chegar a terceira fase da respiração, foram liberados quatro
hidrogênios durante a glicólise, que foram capturados sintetizados em
duas moléculas de NADH2. Na reação de cada ácido pirúvico com a
coenzima A formam-se mais duas moléculas de NADH2. No ciclo de
Krebs, dos oito hidrogênios liberados, seis se combinam com três
moléculas de NAD, formando três moléculas de NADH2, e dois se
combinam com outro aceptor, o FAD, formando uma molécula de
FADH2.
O destino final dos hidrogênios liberados pela respiração é a
combinação com átomos do
atmosférico [sendo assim, o oxigênio é
chamado de aceptor final de hidrogênios], formando água e evitando a
acidose [diminuição do pH] da célula.
Antes de reagirem com o O2, porém, os hidrogênios, percorrem uma
longa trajetória, na qual se combinam sucessivamente com diversas
substâncias aceptoras intermediárias. Somente ao final dessa trajetória,
os hidrogênios se encontram com seus parceiros definitivos, os átomos
de oxigênio. Esse conjunto de substâncias transportadoras de
hidrogênio constitui a cadeia respiratória [também chamada de cadeia
transportadora de elétrons ou fosforilação oxidativa], que ocorre nas
cristas mitocondriais.
Na transferência de hidrogênios ao longo da cadeia respiratória, há
liberação de elétrons excitados, que vão sendo captados por
transportadores intermediários, dentre eles os citocromos. Desgastados,
ao chegarem no final da cadeia respiratória, os elétrons são captados
pelo oxigênio [aceptor final de hidrogênios e elétrons]. As moléculas
de NAD, de FAD e de citocromos que participam da cadeia respiratória
captam hidrogênios e os transferem, através de reações que liberam
energia, para um aceptor seguinte. Os aceptores de hidrogênio que
fazem parte da cadeia respiratória estão dispostos em sequência na
parede interna da mitocôndria.
Com o último aceptor de hidrogênios na cadeia respiratória ocorre
a formação de moléculas de ATP, processo chamado de fosforilação
oxidativa. Cada molécula de NADH2 que inicia a cadeia respiratória
leva à formação de três moléculas de ATP a partir de três moléculas de
ADP e três grupos fosfato.
1
:
1
+ 2
+ 3 := + 3= → 1
+ 3 >= + 1
:
Já a FADH2 formado no ciclo de Krebs leva à formação de apenas duas
moléculas de ATP.
15 :
1
+ 2
+ 2 := + 2= → 1
+ 2 >= + 15 :
Desse modo, tradicionalmente [há novas pesquisas que contestam os
resultados, mas por serem muito recentes ainda não são admitidas]
considera-se que a cadeia respiratória gera 34 moléculas de ATP.
Somando esse número ao saldo de dois ATP formados na glicólise e de
dois ATP formados no ciclo de Krebs, o rendimento energético total da
respiração é de 38 ATP.
O oxigênio participa diretamente apenas da última etapa da cadeia
respiratória. Apesar disso, é um reagente fundamental para que a
respiração ocorra, pois todas as demais reações da fase aeróbia cessam
na sua ausência. Sem o gás oxigênio, alguns organismos e mesmo
células do tecido muscular esquelético humano continuam a realizar
glicólise [por se tratar de uma etapa anaeróbia da respiração aeróbia],
mas como o ciclo de Krebs e a cadeia respiratória necessitam dele para
funcionar, não ocorrem, e a célula desvia o metabolismo para a
fermentação.
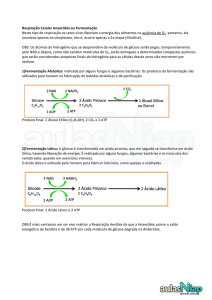
FERMENTAÇÃO
Por meio da fermentação, a glicose é parcialmente degradada na
ausência de oxigênio, originando substâncias mais simples, como o
ácido lático [na fermentação lática] e o álcool etílico [na
fermentação alcoólica]. Nesses processos, há saldo de apenas duas
moléculas de ATP.
Um dos produtos da glicólise é o NADH2 que se forma a partir do %&'( .
Esta substância é essencial para a quebra da molécula de glicose, mas
existe em quantidades muito inferiores às necessárias para que a
glicólise ocorra. Assim, o
: ( deve ser recuperado.
Para isso existem dois mecanismos:
•
•
Usando oxigênio para oxidar o NADH2: caracterizando a
respiração aeróbia, que obviamente depende do oxigênio.
Na ausência de oxigênio, usando o próprio piruvato formado na
glicólise: caracterizando a fermentação.
Todos os tipos de fermentação ocorrem exclusivamente no citosol ou
hialoplasma e são processos autossuficientes, diferenciando-se somente
quanto ao produto final da reação. Entre os tipos de fermentação
destacam-se a fermentação lática e a fermentação alcoólica.
Os lactobacilos [bactérias presentes no leite] executam fermentação
lática, em que o produto final é o ácido lático. Para isso, eles utilizam
como ponto de partida, a lactose, o açúcar do leite, que é desdobrado,
por ação enzimática que ocorre fora das células bacterianas, em glicose
e galactose. A seguir, os monossacarídeos entram nas células, onde
ocorre a fermentação.
Cada molécula do ácido pirúvico é convertida em ácido lático, que
também contém três átomos de carbono.
O sabor azedo do leite fermentado se deve ao ácido lático formado e
eliminado pelos lactobacilos. A diminuição do pH causado pelo ácido
lático provoca a coagulação das proteínas do leite e a formação do
coalho, usado na fabricação de iogurtes e queijos.
Às vezes a quantidade de oxigênio que as células musculares recebem
para a respiração aeróbia é insuficiente para a liberação da energia
necessária para a atividade muscular intensa.
Nessas condições, ao mesmo tempo em que as células musculares
continuam respirando, elas começam a fermentar uma parte da glicose,
na tentativa de liberar energia extra.
O ácido láctico acumula-se no interior da fibra muscular produzindo
dores, cansaço e cãibras. Depois, uma parte desse ácido é conduzida
pela corrente sanguínea ao fígado, onde é convertido em ácido
pirúvico, e o restante é metabolizado pelas células musculares quando
o teor de oxigênio é restabelecido.
As leveduras e algumas bactérias fermentam açúcares, produzindo
álcool etílico e gás carbônico [CO2], processo denominado fermentação
alcoólica. Na fermentação alcoólica, as duas moléculas de ácido
pirúvico produzidas são convertidas em álcool etílico [também
chamado de etanol], com a liberação de duas moléculas de CO2 e a
formação de duas moléculas de ATP.
Esse tipo de fermentação é realizado por diversos microrganismos,
destacando-se
os
chamados
“fungos
de
cerveja”,
da
espécie Saccharomyces cerevisiae. O homem utiliza os dois produtos
dessa fermentação: o álcool etílico, empregado há milênios na
fabricação de bebidas alcoólicas [vinhos, cervejas, cachaças etc.], e
o gás carbônico, importante na fermentação de massas como os pães.