Pós -Graduação a Dist ância
Bioenergética, Metabolismo de Carboidratos, Lipídios e Proteínas
Bioenergética, Metabolismo de
Carboidratos, Lipídios e Proteínas
Professor Alexandre de Souza Aguiar
www.posugf.com.br
1
Bioenergética, Metabolismo de Carboidratos, Lipídios e Proteínas
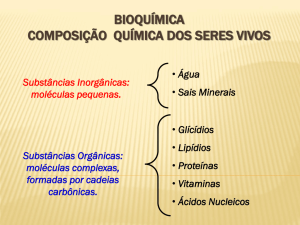
Carboidratos - Conceito e Funções
3
Funções3
SUMÁRIO
Classificação
3
Estruturas Moleculares
3
Digestão e Absorção
4
Aplicação na Prática Clínica do Nutricionista
5
Índice Glicêmico
5
Carga Glicêmica
5
Fibras6
Glicose no Sangue
7
Glicólise na Célula
8
Ciclo de Krebs
9
Glicogênese11
Glicogenólise11
Metabolismo de Lipídios
11
Digestão dos Lipídios
12
Absorção dos Lipídios
12
Biossíntese de Ácidos Graxos - Lipogênese
13
Mobilização de Lipídios - Lipólise
14
Proteínas16
Histórico16
Definição
16
Funções16
Estrutura16
Aminoácidos17
Digestão de Proteínas
18
Destino dos Aminoácidos
19
Papel do Fígado
19
Desaminação Oxidativa
20
Transaminação21
Ciclo da Ureia
21
Descarboxilação de Aminoácidos
22
Transmetilação de Aminoácidos
22
Transulfunação de Aminoácidos
22
Gliconeogênese23
Referências24
www.posugf.com.br
2
Bioenergética, Metabolismo de Carboidratos, Lipídios e Proteínas
crustáceos, também compõem parte das funções
atribuídas a esses compostos.
CARBOIDRATOS - CONCEITO E
FUNÇÕES
• As glicoproteínas e glicolípedes também se
enquadram em algumas funções, fazendo papel
de reconhecimento em exames laboratoriais, como
é o caso das glico-hemoglobinas, e também de
reconhecimento celular, caso dos glicocálices.
O grupo dos carboidratos deve perfazer em
média 55% a 60% da nossa dieta. Em casos mais
específicos, como atletas de alto rendimento, podese superar essa média. Os carboidratos representam
a nossa principal fonte de energia, por isso a
importância de tê-los bem programados em um
plano alimentar. Sabemos que várias dietas da moda
tendem a reduzir a quantidade de carboidratos com
a justificativa de que eles representam os principais
vilões do ganho de peso. A nutrição científica valoriza
a dieta balanceada, por isso devemos entender as
rotas metabólicas a fim de alcançar sucesso no plano
alimentar de nossos pacientes.
Classificação
Como foi citado, os carboidratos dividem-se em:
1)Monossacarídeos, que representam as
unidades elementares para absorção em nosso
intestino: glicose, galactose e frutose, assim como
as pentoses, que possuem funções estruturais de
nossas bases nucleicas: DNA e RNA.
O grupo dos carboidratos é constituído por
estruturas dotadas das funções aldeídos (aldoses)
e cetonas. A denominação sacarídeo vem do grego
sakcharon, que significa açúcar. A fórmula geral
(CH2O)n denota a característica física que esses
nutrientes têm de se conjugarem com moléculas de
água, o que explica em parte algumas particularidades
de seu processo de digestão e absorção. Os
carboidratos são assim então subdivididos de acordo
com a complexidade de suas ligações glicosídicas:
monossacarídeos, oligossacarídeos e polissacarídeos.
2) Polissacarídeos, que constituem a forma mais
complexa e também mais encontrada nos alimentos,
representada por amidos, fibras e glicogênio. Este
último representa a forma armazenada de energia.
3) Oligossacarídeos: possuem tamanho mediano
entre os monossacarídeos e polissacarídeos.
Estruturas Moleculares
Funções
• A função energética é a principal atribuída aos
carboidratos, pois, de fato, trata-se de uma atividade
de suma importância para manter a homeostase
corpórea. Os amidos, ao serem digeridos e absorvidos,
passam pelo processo de fosforilação oxidativa, que
garante o fornecimento de energia para todas as
células de nosso organismo.
• As fibras, compostos protetores da parede
celular de alguns vegetais e dos exoesqueletos de
www.posugf.com.br
3
Bioenergética, Metabolismo de Carboidratos, Lipídios e Proteínas
Digestão e Absorção
basolateral, segunda barreira, com auxílio das gluts
2, que possuem afinidade por todas as espécies de
sacarídeos, tornando a entrada desses compostos
mais facilitada na corrente sanguínea.
O processo de digestão dos carboidratos inicia-se
pelo estímulo da mastigação com atuação da amilase
salivar. Os amidos sofrem a lise por intermédio
dessa enzima, alfa amilase, que transforma os
polissacarídeos em estruturas menores. O processo
tem sua continuidade no duodeno, pela ação da
enzima amilase pancreática, facilitando o processo
de absorção.
A hidrólise é finalizada na superfície das células
epiteliais do intestino delgado, enterócitos, onde
enzimas como sacarase, lactase e maltase estão
preparadas para exercer tal papel, fazendo com que
a formação dos monossacarídeos absorvíveis ocorra
com maior facilidade.
No caso das fibras, não ocorre essa hidrólise,
restando a elas a fermentação, que é realizada
por bactérias probióticas que colonizam o nosso
intestino. Temos como produto dessa fermentação os
ácidos graxos de cadeia curta - acetato, propionato e
butirato -, que serão usados como fração energética
para os enterócitos, além de facilitarem a absorção
de alguns minerais e eliminarem pela redução do pH
as bactérias patógenas.
Finalmente, teremos o processo de absorção
iniciado, porém os carboidratos encontram certa
dificuldade em atravessar as barreiras epiteliais
dos enterócitos. Isso se explica porque eles são
hidrossolúveis e não interagem com a barreira lipídica
dessas células intestinais. Diante do exposto, faz-se
necessário o auxílio das proteínas de transporte:
gluts e SGLT-1.
Nas vilosidades, encontramos duas proteínas
que auxiliam no transporte dos monossacarídeos,
as gluts 5, que auxiliam na absorção da frutose, e
o SGLT-1, que auxilia no transporte de sódio, que
por sua vez beneficia o transporte de água, glicose
e frutose. Após a passagem pelas vilosidades,
os monossacarídeos atravessam a membrana
www.posugf.com.br
4
Bioenergética, Metabolismo de Carboidratos, Lipídios e Proteínas
glicose assume valores baixos após seu consumo.
Veja a tabela.
Aplicação na Prática Clínica do
Nutricionista
Alguns fatores, como a presença de fibras solúveis,
gordura e proteínas e a técnica do fracionamento são
importantes para a redução do índice glicêmico.
Diante do que foi comentado até agora, precisamos
chamar atenção para algumas situações que são
inerentes ao universo do atendimento nutricional.
A primeira é reforçar a importância de uma boa
mastigação para poder otimizar o estímulo da
salivação e melhorar a funcionalidade no processo de
digestão. Sabemos que a falta de tempo faz com que
as pessoas disponibilizem momentos cada vez mais
curtos para suas refeições, prejudicando o processo
de digestão.
Outro ponto importante é valorizar a hidratação,
sempre conjugada com minerais. Daí a importância
das frutas, que possuem em sua estrutura água,
sódio e carboidratos, porque, como vimos, a glicose
somente consegue entrar pela corrente sanguínea
pelo transportador sódio-glicose-acoplado (SGLT-1),
o que mostra a importância dessa combinação para
uma boa hidratação.
Lembre-se da campanha do soro caseiro, em que
essa combinação -água, sal e açúcar - retirou milhares
de crianças da situação de risco da desidratação e
mortalidade infantil.
Índice Glicêmico
Uma forma de monitorar o metabolismo dos
carboidratos no tocante à velocidade de absorção é
entender a dinâmica do índice glicêmico e da carga
glicêmica.
Fonte: http://sol-meu-emagrecimento.blogspot.com.br
Índice glicêmico representa um indicador que
revela a habilidade do alimento em elevar os níveis
de glicose sanguínea após seu consumo, comparado
com os valores relacionados ao consumo de 50 g
de carboidrato na forma de pão branco ou glicose.
Sendo assim, um alimento de alto índice glicêmico é
aquela cuja glicose assume valores altos após o seu
consumo e um de baixo índice glicêmico é aquele cuja
Carga Glicêmica
Representa o efeito da quantidade e qualidade
do carboidrato, expresso por porção de alimento
ingerido.
CG= (porção carboidrato x IG)/100
www.posugf.com.br
5
Bioenergética, Metabolismo de Carboidratos, Lipídios e Proteínas
Classificação
CG do alimento
que confere sua insolubilidade no meio aquoso, pois
a água torna-se incapaz de penetrar no alimento
diante das justaposições das interfaces. Essa fibra
possui, então, uma menor viscosidade que a torna
mais cinética no tubo entérico, permitindo assim uma
aceleração do bolo fecal. Recomenda-se o consumo
das fibras insolúveis aos pacientes que sofrem de
prisão de ventre, geralmente mulheres, pessoas
acamadas ou que fizeram uso de algum medicamento
que resultou em constipação.
CG da dieta
Baixa
Menor ou
igual a 10
Menor ou
igual a 80
Alta
Maior ou
igual a 20
Maior ou
igual a 120
Fibras
As fibras também são carboidratos, porém não
digeríveis.
Estima-se que a quantidade recomendada de
fibras em uma dieta balanceada não é alcançada
por grande parte da população brasileira. Isso
ocorre em função da grande quantidade de produtos
alimentícios refinados fabricados pela indústria que
hoje compõem a mesa do brasileiro.
As fibras diferenciam-se em solúveis e insolúveis,
e essa diferenciação está relacionada com a forma
da estrutura química das cadeias de sacarídeos, que
podem ser lineares ou ramificadas.
As fibras solúveis, representadas por pectinas,
gomas, mucilagens beta glucanas, ligninas e polpas
de frutas, possuem uma estrutura ramificada que
confere pouca superfície de contato entre suas
cadeias, permitindo maior absorção de água entre
suas interfaces. Essa ocorrência permite que
esse tipo de fibra crie uma espécie de “esponja”,
aumentando seu volume no estômago e criando
a sensação de saciedade devido à compressão
gástrica promovida nas células parietais gástricas,
estimulando a produção de hormônios sacietogênicos
como a bombesina. Além disso, também se juntam
às moléculas de colesterol, impedindo o excesso
de absorção, e por fim promovem a produção dos
ácidos graxos de cadeia curta ao serem fermentadas
no intestino. Essa fermentação faz o controle de
colonização das bactérias probióticas.
Fonte:http://academiafw.blogspot.com.br/2011/08/importancia-dasfibras-para-quem-quer.html
As fibras insolúveis, como as celuloses presentes
na estrutura protetoras das cascas e folhas, possuem
uma estrutura linear que confere alta superfície de
contato entre suas moléculas. É justamente isso
www.posugf.com.br
6
Bioenergética, Metabolismo de Carboidratos, Lipídios e Proteínas
Após a absorção, a glicose encontra-se no sangue
e deve seguir para o meio intracelular por intermédio
dos receptores gluts.
e adiposas, necessitando de uma sinalização da
insulina para ser mobilizada para a membrana.
Esse processo ocorre por meio de um mecanismo
de autofosforilação entre algumas enzimas, como
tirosina quinase, IC3K e AKT.
A principal glut do nosso corpo é a glut-4, que se
encontra no meio intracelular de células musculares
Produção de insulina nas células beta
pancreáticas
Glicose no Sangue
Fonte: http://0physiologyonline.phy
s i o l o g y. o r g . l i b r a r y. p c c . e d u /
content/25/3/155/F2.expansion.html
Mobilização de glut-4 pela insulina em células musculares e adipócitas
www.posugf.com.br
7
Bioenergética, Metabolismo de Carboidratos, Lipídios e Proteínas
verde-escuros, que são fontes desses minerais, para
termos assim enzimas com alto nível de atividade.
A inatividade dessas enzimas pela deficiência desses
minerais pode promover a “fuga” da glicose do meio
intracelular de volta para a corrente sanguínea,
promovendo o quadro de hiperglicemia por deficiência
de magnésio.
Glicólise na Célula
A glicose, ao entrar na célula, por meio do
transportador glut-4, pode fazer o caminho inverso
e retornar para corrente sanguínea. Isso porque
ela já possui afinidade pelo receptor, o que confere
a ela maior permeabilidade. Esse processo não é
interessante, visto que o nosso objetivo é converter
esse carboidrato em energia, portanto devemos
mantê-lo no meio intracelular.
É importante destacar que a adição de um átomo
de fósforo à molécula de glicose exige a retirada
de um outro átomo, hidrogênio, porém este sai na
forma de íons H+, um radical livre acidófilo, o que
coloca em risco todo o metabolismo celular. Diante
dessa situação, tem-se que fazer a captura desse
íon hidrogênio. O melhor receptor de hidrogênio é
o oxigênio, pois juntos formam a molécula de água.
Entretanto, nessa posição celular, não há hidrogênio,
pois este se encontra dentro de organelas aeróbicas
denominadas mitocôndrias, por isso a necessidade
de mobilizar a rota metabólica para essa organela.
Nessa etapa, surgem cofatores enzimáticos que
funcionam como transportadores de hidrogênio para
o meio mitocondrial. A niacina, vitamina B3, e a
riboflavina, vitamina B2, são cofatores enzimáticos
dos componentes NAD e FAD. Essas estruturas
têm a função de carrear o hidrogênio para o meio
mitocondrial, daí a mudança de NAD para NADH e
FAD para FADH.
E como a bioquímica resolve esse problema?
Simples: basta alterarmos a fórmula química
da glicose para que ela perca a afinidade pelos
receptores gluts-4, ficando assim “presa” dentro da
célula. É por isso que na bioquímica ouvimos falar
em fosforilação dos carboidratos. Ao fornecer um
átomo de fósforo para a glicose, esta se torna então
uma nova molécula, incapaz de escapar do meio
intracelular.
Dessa forma, o mecanismo de fosforilação tornase um processo que permite obter energia por
intermédio dos nutrientes, uma vez que otimiza sua
metabolização intracelular.
As quinases são enzimas responsáveis por realizar
esse mecanismo. No caso da glicose, a glicose fosfatase
encarrega-se de fazer essa primeira fosforilação. Há
dois tipos de glicose fosfatase: a glicose 1 fosfatase,
também chamda de glicoquinase, e a glicose 6
fosfatase, também chamada de hexoquinase. A
primeira tem maior demanda no fígado, e sua função
é fosforilar a glicose no carbono de posição número
1. Isso significa que essa molécula de glicose será
usada como armazenamento na forma de glicogênio
hepático. A segunda faz a fosforilação na posição
número 6, indicando a rota de oxidação da glicose,
na qual será usada como fonte de energia.
Após a fosforilação, uma série de metabólicos
de seis carbonos é produzida no citoplasma (veja
figura da glicólise) até a conversão em frutose 1,6
difosfato. Porém a entrada na mitocôndria exige uma
partição dessa molécula em um subproduto menor,
para melhorar a permeabilidade na membrana
mitocondrial. Diante dessa situação, teremos o
início de uma reação denominada glicólise, na qual
a molécula de seis carbonos se divide em duas
moléculas de três carbonos, denominadas ácido
pirúvico ou piruvato.
As quinases são enzimas ativadas por cofatores
dependentes de magnésio e/ou manganês, de onde
se evidencia a importância de alimentos vegetais
Após a formação do piruvato, tem-se a entrada
na mitocôndria e em seguida a formação do acetil,
um composto de dois carbonos que dará início a ciclo
www.posugf.com.br
8
Bioenergética, Metabolismo de Carboidratos, Lipídios e Proteínas
de Krebs. A formação do acetil a partir do piruvato
é mediada pela enzima piruvato desidrogenase, que
por sua vez é ativada pelo cofator tiamina, vitamina
B1. Note como as vitaminas do complexo B participam
dessa formação energética a partir dos carboidratos,
reforçando a importância de alimentos integrais para
esse processo.
Ciclo de Krebs
A formação do acetil já foi feita, porém é preciso
sinalizar a sua mobilização para a via energética. Essa
é uma outra etapa, que envolve mais uma enzima,
a coenzima a-CoA. Ela também possui como cofator
uma vitamina do complexo B, o ácido pantotênico,
vitamina B5. Temos agora o composto acetil-CoA, que
representa o primeiro metabólico da via energética.
Os principais destinos do acetil irão depender da
necessidade orgânica: ciclo de Krebs (lactato), ciclo
de Cori (acetil-CoA), síntese de proteínas (alanina) e
gliconeogênese (oxaloacetato).
A partir desse momento, uma série de reações
mediadas por desidrogenases ocorre na matriz
mitocondrial, começando pela conjugação do acetilCoA com o oxalacetato, dando origem a um segundo
metabólico, que se chama citrato. Varias reações
subsequentes levarão à formação de subprodutos:
alfa glutarato, succinato, fumarato e malato. Em todas
essas reações, há a participação dos complexos NAD
e FAD, mostrando a ocorrência das desidrogenações
e a dependência das vitaminas B3 e B2.
No final do ciclo de Krebs, temos um rendimento
de ATP, 3NADH e 1FADH, porém lembrem-se de que
o papel das vitaminas B2 e B3 era carrear hidrogênio
para o oxigênio e que agora é chegada a hora de
fazer essa entrega.
Após a finalização do ciclo de Krebs, as moléculas
de NADH e FADH migram para a membrana da
mitocôndria, região chamada de cristas mitocondriais,
onde será feita a doação de hidrogênio para a
molécula de oxigênio com a consequente formação
de água. Porém esse processo não é tão simples,
pois o hidrogênio é lançado na forma de H+ para a
região intermembranas, o que provoca uma mudança
de polarização entre os meios.
O retorno desses íons de hidrogênio energizados
passando pela membrana interna da mitocôndria a
fim de normalizar a carga da célula é mediado pela
enzima ATP sintetase. Durante essa passagem,
ocorre uma mudança na conformação dessa enzima,
fazendo com que sítios de ligação de ADP se liguem
a sítios de fósforo inorgânico, Pi, promovendo a
www.posugf.com.br
9
Bioenergética, Metabolismo de Carboidratos, Lipídios e Proteínas
formação de moléculas energéticas de ATP. Assim
conseguimos formar as moléculas de ATP a partir dos
carboidratos consumidos.
P—P, uma quantidade de energia que é liberada
para o nosso corpo na forma de energia térmica e
mecânica, promovendo assim as nossas funções
fisiológicas.
Mas, enfim, o que é o ATP?
A quebra dessas ligações entre os átomos de
fósforo é mediada pela enzima ATPase, que também
possui como cofator o mineral magnésio.
ATP é uma molécula denominada adenosina
trifosfato, que armazena entre as ligações de fósforo,
www.posugf.com.br
10
Bioenergética, Metabolismo de Carboidratos, Lipídios e Proteínas
Hormônios como adrenalina, produzida durante
a atividade física, e glucagon, produzido durante
a queda de glicose sanguínea, promovem a
degradação do glicogênio por estímulo da proteína
G, que fica na superfície da célula e que ativará a
proteína quinase, responsável por induzir as enzimas
glicogênio fosforilase e desramificadora de glicogênio,
que irão agir na despolimerização do glicogênio em
moléculas de glicose. A glicose passará então através
do transportador glut-2, que fica na membrana dos
hepatócitos, para o meio extracelular, regressando
à corrente sanguínea e fazendo a manutenção da
glicemia.
Glicogênese
A síntese do glicogênio ocorre a partir da glicose,
podendo ocorrer tanto no músculo quanto no fígado.
O glicogênio é na verdade um polímero de glicose,
um polissacarídeo.
Após as refeições, quando a glicose se encontra
alta no sangue, damos início à glicogênese a partir do
excesso de glicose 6 fosfato, que é então convertida
em glicose 1 fosfato. Esse processo é mediado pela
enzima glicogênio sintetase.
À medida que o polímero de glicogênio aumenta,
ocorre o estímulo de outra enzima, denominada
enzima ramificadora, que irá organizá-lo em estruturas
radicalares. O fato de a cadeia de glicogênio conter
radicais facilita a quebra dessa estrutura no momento
de jejum, pois as estruturas ramificadas possuem
menor superfície de contato entre suas interfaces,
permitindo uma hidrólise mais facilitada.
METABOLISMO DE LIPÍDIOS
Os lipídios representam o grupo de maior energia
da nossa dieta (9 kcal/g, contra 4 kcal/g dos
carboidratos e proteínas). Devem perfazer em média
de 25% a 30% da dieta. Devemos nos atentar para
as diferentes classes desse nutriente, que variam
entre lipídios saturados, insaturados e trans.
Existem duas formas de glicogênio: o hepático,
que é utilizado para manter a glicose sanguínea
durante momentos de jejum, e o muscular, que é
usado pelo próprio músculo para formação de ATP.
As gorduras saturadas são encontradas,
principalmente, nos alimentos de origem animal. Já
as gorduras insaturadas, que se dividem em mono
e poli-insaturadas, são encontradas, principalmente,
nas fontes vegetais.
É importante salientar que o excesso de
carboidratos em nossa alimentação, em um primeiro
momento, contribui para o acúmulo de glicogênio
tanto hepático quanto muscular e que, para cada
grama de glicogênio formado, acumulamos 3 g de
água. Isso explica o fato de estarmos mais pesados
após uma dieta com excesso de carboidratos.
Monoinsaturada = ômega 9 » abacate, linhaça,
gergelim, oliva.
Poli-insaturada = ômega 3 » peixes, canola,
gergelim, chia, linhaça, oliva.
Glicogenólise
Poli-insaturada = ômega6 » soja, milho, girassol.
Denominamos glicogenólise o processo de
degradação do glicogênio que ocorre nos momentos
em que nos encontramos hipoglicêmicos, em função
de atividade física ou jejum curto (três a quatro
horas).
As gorduras trans são formadas a partir do
processo de hidrogenação das gorduras vegetais e
conferem maior durabilidade aos alimentos. Por essa
razão, são usadas com objetivo de aumentar o prazo
de validade destes.
www.posugf.com.br
11
Bioenergética, Metabolismo de Carboidratos, Lipídios e Proteínas
A nossa legislação permite que os alimentos com
até 0,2 g de gordura trans na porção podem anunciálo como isento dela. Não devemos nos esquecer de
que o limite de gordura trans que uma dieta deve
atingir é de 2 g/d.
pâncreas e também na mucosa intestinal. As lípases
pancreáticas possuem um alto potencial de ação lítica
e iniciam o processo de hidrólise do TAG. Moléculas
de ácidos graxos são retiradas, dando origem
aos compostos diacilglicerol e monoacilglicerol e
finalmente aos ácidos graxos livres e ao glicerol livre.
As gorduras do tipo ômega 6, como o ácido
araquidônico, são metabolizadas por enzimas ciclooxigenase de classe 2-COX-2, promovendo ação
pró-inflamatória. Já os óleos do tipo ômega 3 como
linolênico, EPA e DHA, são metabolizados por ciclooxigenase de classe 1- COX-1 e exercem papel antiinflamatório.
Devemos nos atentar para a importância que o
fígado e o pâncreas possuem nesse processo de
digestão dos lipídios. Portanto, alterações patológicas
nesses órgãos, bem como a ausência de bile,
podem surtir problemas que evoluirão para diarreia
gordurosas (esteatorreia) devido a dificuldade no
processo de digestão.
Devemos salientar que o EPA inibe a COX-2, o que
reforça sua ação anti-inflamatória. Já o DHA possui
ação nos receptores de memória, melhorando a
nossa cognição.
Digestão dos Lipídios
Grande parte dos lipídios presentes em nossa
alimentação está na forma de triacilglicerol - TAG.
Essa molécula é composta pela combinação de
três ácidos graxos e um glicerol. Diante disso, é
necessário promover a hidrólise do TAG para que
seus subprodutos consigam atravessar a barreira das
vilosidades intestinais.
Os lipídios são moléculas de baixa solubilidade,
o que dificulta sua exposição enzimática. Diante
disso, o fígado produz um líquido emulsificante
denominado de bile. A bile tem como função tornar
a gordura mais biodisponível, aumentando suas
superfícies de contato com as enzimas lipolíticas.
Trata-se de uma secreção produzida pelo fígado e
armazenada na vesícula biliar. Com a chegada do
alimento no duodeno, um hormônio denominado
colecistoquinina - CCK - estimula a contração da
vesícula e a consequente liberação da bile.
Fonte: http://www.mumbaicancer.com/bile_duct.asp
Absorção dos Lipídios
Agora que os lipídios estão organizados em
moléculas menores, tem início o processo de
absorção. A absorção dos lipídios ocorre na mucosa
intestinal por dois processos:
I) Flip-flop: processo mecânico em que os
lipídios giram em seu próprio eixo passando pelas
bicamadas.
Após emulsificada, a gordura torna-se mais
vulnerável à ação das enzimas lipolíticas (lípases
lipoproteicas - LLP), que são produzidas pelo
www.posugf.com.br
12
Bioenergética, Metabolismo de Carboidratos, Lipídios e Proteínas
II)FATP- proteína transportadora de ácidos
graxos. Essa proteína fixada na membrana das
vilosidades facilita o transporte desses compostos
para o interior do enterócito.
Diante de uma dieta hipercalórica, o citrato
é mobilizado de dentro da mitocôndria para o
citoplasma. Após essa mobilização, a enzima citrato
liase promove a lise desse composto em oxalacetato
e acetil-CoA. Na verdade, esse mecanismo tem
como objetivo a retirada do acetil-CoA de dentro
da mitocôndria, paralisando assim o ciclo de Krebs,
visto que a produção de ATP já está no seu limite.
Os acetis-CoAs que agora estão fora da mitocôndria
sofrem ação de uma enzima citoplasmática chamada
acetil-CoA-carboxilase, que irá promover a união
dos grupos de dois carbonos. Essa soma é feita de
dois a dois em vários ciclos até dar origem a um
metabólico graxo (gorduroso) denominado malonil,
de 14 carbonos. É importante salientar que a acetilCoA-carboxilase é ativada pela insulina e inibida pela
adrenalina e pelo glucagon.
Após a absorção, ácidos graxos e glicerol
encontram-se dentro do enterócito, sendo
reesterificados novamente em TAG e ligando-se a uma
lipoproteína transportadora denominada quilomícron.
Este transporta esse complexo reesterificado para o
sistema linfático e posteriormente para a corrente
sanguínea, concluindo assim o processo de absorção.
Um ponto importante na rota metabólica é a
formação do malonil, que ainda não é a gordura que
nós, seres humanos, armazenamos nos adipócitos.
Sua presença promove algumas reações que
favorecem o acúmulo de gordura, pois, quando o
corpo se vê diante da produção de malonil, interpreta
que estamos diante de uma dieta hipercalórica. Nesse
momento, toda oxidação dos TAGs que estavam
estocados nos adipócitos fica interrompida, o que faz
sentido, pois somente oxidamos gordura diante de
um quadro de hipocalorias ou sob atividade física.
Biossíntese de Ácidos Graxos Lipogênese
Outra ação do malonil é inibir a enzima carnitina
palmitoil transferase I, que facilita a entrada da
gordura dentro da mitocôndria para ser oxidada.
Uma dieta hipercalórica promoverá dentro da célula
o excesso de acetil-CoA, vindo dos carboidratos ou
até mesmo das proteínas. Normalmente esse acetilCoA entraria no ciclo de Krebs e seria convertido em
energia na forma de ATP. Entretanto, ATP é uma
forma de energia que nós, seres humanos, não
conseguimos armazenar, ou seja, gasta-se aquilo que
se produz. Então, diante do excesso de acetil-CoA,
nosso organismo teve que desenvolver habilidades
que nos garantem a reserva de energia na forma de
gordura.
Conclusão: o excesso de carboidratos, pelo visto,
contribui mais com um efeito poupador de gordura
do que necessariamente produtor de gordura, até
porque o próximo passo do malonil é receber mais
dois carbonos e transformar-se em palmitato (16C),
gordura que os seres humanos armazenam. Acontece
que as enzimas que fazem essa conversão estão
ausentes em nós. Assim, concluímos que o palmitato
que nós acumulamos vem da dieta e não do excesso
de carboidratos.
www.posugf.com.br
13
Bioenergética, Metabolismo de Carboidratos, Lipídios e Proteínas
graxos, também chamados de acil, são incorporados
no meio intracelular e devem seguir para a matriz
mitocondrial, dando início ao processo de oxidação.
Porém esse processo exige a demanda de uma outra
proteína que leva os ácidos graxos para a matriz
mitocondrial, a carnitina.
Mobilização de Lipídios - Lipólise
Obter energia a partir dos triacilgliceróis
armazenados requer esforço. É preciso ativar duas
reações: lipólise e beta oxidação.
A lipólise significa a quebra do TAG dentro do
tecido adiposo. Essa reação é feita pela enzima
lípase hormônio-sensível - LHS - sob o comando de
alguns hormônios lipolíticos, como GH, glucagon e
adrenalina. Esses hormônios ligam-se a receptores
beta que ficam na membrana dos adipócitos e
sinalizam a atividade da LHS no processo lipolítico.
A carnitina é uma proteína cuja função é atravessar
o acil pela membrana da mitocôndria no chamado
complexo acil-carnitina, porém esse mecanismo
depende de duas enzimas:
CPTI - carnitina palmitoil transferase I ou CATI carnitina acil transferase I: essa enzima faz a ligação
da carnitina ao acil fora da membrana da mitocôndria,
favorecendo a entrada do complexo acil-carnitina.
Após a hidrólise do TAG, temos então três
ácidos graxos e um glicerol novamente. Os ácidos
graxos dentro dos adipócitos ligam-se a proteínas
transportadoras – FABP -, que os movem para fora
da célula, ou seja, para a corrente sanguínea.
CPTII - carnitina palmitoil transferase II ou CATII
- carnitina acil transferase II: dentro da mitocôndria,
essa enzima remove a carnitina do grupo acil,
fazendo com que retorne para a membrana e que os
ácidos graxos livres (acil) fiquem prontos para serem
oxidados.
No sangue, os ácidos graxos livres ligam-se às
albuminas, que os transportam para outro tecido
com a habilidade de oxidar gorduras - fígado ou
músculo. Após alcançar esses tecidos, os ácidos
www.posugf.com.br
14
Bioenergética, Metabolismo de Carboidratos, Lipídios e Proteínas
A beta oxidação depende de um sinalizador, a
coenzima A, que é dependente da vitamina B5 - ácido
pantotênico. Tem-se então a formação do complexo
acil-CoA, o qual será o protagonista da oxidação.
Uma molécula de palmitato, que é o acil, possui 16
carbonos, mas lembrem que o ciclo de Krebs iniciase com um metabólico de dois carbonos chamado
acetil. Logo, temos que fazer a divisão de um acil
em oito acetis. Essa é a proposta da beta-oxidação,
promover várias moléculas de acetil-CoA a partir
de uma molécula de acil-CoA. Isso explica o fator
energéticos dos ácidos graxos ser maior que o das
proteínas e carboidratos.
Podemos, assim, afirmar que o processo de
eliminação de gordura deve passar por dois processos:
lipólise e beta oxidação.
www.posugf.com.br
15
Bioenergética, Metabolismo de Carboidratos, Lipídios e Proteínas
PROTEÍNAS
Funções
As proteínas diferem dos carboidratos e dos lipídios
pela presença do elemento azoto (nitrogênio). Em
nossa dieta, deve alcançar a recomendação de 10%
a 15%, ou seja, uma quantidade bem menor do que
a que vimos para carboidrato e lipídios. Isso ocorre
porque os aminoácidos que são precursores das
proteínas não têm como função principal a formação
de ATP, ficando com outras atribuições. Sabemos que
a dieta do brasileiro é hiperproteica, o que pode gera
algumas intercorrências indesejáveis.
»»
Hormônios e receptores.
»»
Enzimas.
»»
Neurotransmissores.
»»
Proteínas estruturais.
»»
Proteínas contráteis.
»»
Proteínas motoras.
Histórico
»»
Proteínas transmembranas.
»»
Energia.
1840 - Von Liebig: principal fonte de energia.
1900 - atenção para gorduras e carboidratos.
Estrutura
1977 - Astrand e Rodahl: fonte energética quando
CH estão ausentes.
As proteínas apresentam-se de quatro formas,
conforme sua estrutura e função: primária,
secundária, terciária e quaternária.
1989 - US Board Nutrition considerou o RDA
suficiente, inclusive para esportistas.
Estudos posteriores: concluíram que
necessidades poderiam estar subestimadas.
Colágeno e elastina são exemplo de proteínas
secundárias, a insulina, de terciária, e a hemoglobina,
de quaternária.
as
Definição
São macromoléculas cujos monômeros, moléculas
menores dos polímeros, são aminoácidos.
»»
Dipeptídeos
dois aminoácidos.
»»
Tripeptídeos
três aminoácidos.
»»
Tetrapeptídeos
»»
Polipeptídeos
quatro aminoácidos.
muitos aminoácidos.
www.posugf.com.br
16
Bioenergética, Metabolismo de Carboidratos, Lipídios e Proteínas
Aminoácidos
Os aminoácidos são as estruturas elementares de todas as proteínas.
A estrutura de um aminoácido é composta por cinco partes, que incluem um carbono central (a), um
hidrogênio (H), um grupamento carboxila (COOH), um grupo amino (NH2) e uma cadeia lateral (ou grupo
R). O que diferencia um aminoácido do outro é a cadeia lateral que ele apresenta. A alanina, por exemplo,
tem um grupamento CH3 no grupo R.
www.posugf.com.br
17
Bioenergética, Metabolismo de Carboidratos, Lipídios e Proteínas
Há vários tipos de aminoácidos, que se diferenciam pelo grupo radicalar – R.
Grupos R aromáticos
Grupos R não-polares e alifáticos
Grupos R carregados positivamente
Grupos R não-carregados, mas polares
Grupos R carregados negativamente
Alguns aminoácidos são essenciais, devendo ser
adquirido pela dieta. Outros são não essenciais, pois
são produzidos em nosso organismo.
Os aminoácidos unem-se por ligações peptídicas
para formar as proteínas.
Digestão de Proteínas
A digestão das proteínas inicia-se no estômago,
com quebra de 10-20% das proteínas, pela hidrólise
de proteínas em polipeptídeos a partir da ativação do
pepsinogênio em pepsina em meio ácido (pH 2 a 3).
A maior parte da digestão ocorre no intestino delgado
pela liberação da tripsina, que ativa outras enzimas
Fonte: http://www.ajinomoto.com.br/enciclopedia/amino.html
www.posugf.com.br
18
Bioenergética, Metabolismo de Carboidratos, Lipídios e Proteínas
proteolíticas pancreáticas, carboxipeptidases e quimiotripsina, fazendo com que proteínas intactas e as préhidrolisadas no estômago sejam digeridas em aminoácidos, dipetídeos e polipeptídeos menores. Ainda no
intestino delgado, as células da mucosa intestinal produzem peptidases, que finalmente terminam o processo
de digestão das proteínas pela quebra dos polipeptídeos e dipeptídeos em aminoácidos e dipeptídeos.
Ex: aminopeptidases, dipeptidases, dipeptil aminopeptidases, dipeptil carboxipeptidases.
A absorção dos aminoácidos acontece no jejuno e
no íleo por transporte passivo. Além dos aminoácidos,
dipeptídeos e tripeptídeos também são absorvidos.
»» Oxidação para formação de energia (fígado e
músculo).
»» Músculo: alanina, aspartato,
leucina, isoleucina e valina.
A maioria dos aminoácidos tem um mesmo
transportador dependente de sódio. Os independentes
de sódio são os aminoácidos neutros (Ser, Ter, Cis,
Asp, Gln, Tir) e os básicos (Lis, Arg, His).
Papel do Fígado
O fígado facilita a absorção e a distribuição
dos aminoácidos da dieta para o sangue e outros
tecidos do corpo, inativa e modifica as substâncias
tóxicas presentes no sangue e remove o excesso de
aminoácidos, especialmente os essenciais, mas não
metaboliza BCAAs.
A concentração plasmática de aminoácidos é de 35
mg/dl a 65 mg/dl; valores mais elevados são encontrados
após a refeição, e o excesso será absorvido pelo fígado,
onde serão preferencialmente oxidados.
Destino dos Aminoácidos
»»
glutamato,
Faz a síntese das proteínas do próprio organismo
e de outras substâncias de natureza proteica, realiza
a conversão para glicose (aminoácidos glicogênicos)
e para gorduras (aminoácidos cetogênicos).
Síntese de peptídeos e proteínas.
www.posugf.com.br
19
Bioenergética, Metabolismo de Carboidratos, Lipídios e Proteínas
Aspartato
Desaminação Oxidativa
Alanina
Desaminação é o processo de remoção do
nitrogênio dos aminoácidos com liberação do grupo
amino na forma de amônia livre e formação de um
alfa-cetoácido, que pode ser piruvato, acetil-Coa,
acetoacetil-Coa, a-cetoglutarato, fumarato, succinilCoa ou oxaloacetato.
Glutamato
Oxalacetato
Piruvato
α -cetoglutarato
O que vai determinar o tipo de cetoácido formado
é o aminoácido que foi desaminado. Os cetoácidos
entram no ciclo do ácido cítrico e dessa forma serão
oxidados, formando CO2, H2O e ATP na cadeia de
transporte de elétrons.
A amônia livre (NH4+) liberada pela desaminação
dos aminoácidos será utilizada para síntese de
glutamato e posteriormente convertida em ureia. O
glutamato será formado pela ligação entre a amônia
e o a-cetoglutarato, por ação da enzima glutamato
desidrogenase.
www.posugf.com.br
20
Bioenergética, Metabolismo de Carboidratos, Lipídios e Proteínas
Transaminação
O
glutamato
produzido
será
removido
principalmente no fígado por transaminação
com oxaloacetato e formação de aspartato; ou
será desaminado, produzindo amônia (NH4+) e
a-cetoglutarato. O aspartato e a amônia produzidos
serão utilizados no ciclo da ureia.
No músculo e em outros tecidos, o catabolismo
dos aminoácidos ocorre por transaminação, que é a
transferência do grupo amínico ao a-cetoglutarato,
formando
glutamato
e
o
alfa-cetoácido
correspondente, reação catalisada pelas enzimas
transaminases ou aminotransferases.
Ciclo da Ureia
O aspartato e a amônia produzidos pela
transaminação/desaminação do glutamato e outros
aminoácidos no fígado serão utilizados para síntese
de ureia. Nesse ciclo, primeiramente, a amônia livre
(NH4+) é incorporada à ornitina, formando citrulina.
O aspartato junta-se à citrulina, com formação
do arginosuccinato, que perde uma molécula de
fumarato, produzindo arginina. A arginina recebe
uma molécula de água, formando a ureia (2 NH2),
que será excretada pelos rins.
www.posugf.com.br
21
Bioenergética, Metabolismo de Carboidratos, Lipídios e Proteínas
Descarboxilação de Aminoácidos
Consiste na retirada do grupo carboxílico e na formação do aminoácido correspondente. Essa reação é
mediada pela vitamina B6.
Histidina
Triptofano
Histamina + CO2
Serotonina + CO2
Ácido glutâmico
GABA + CO2
Transmetilação de Aminoácidos
Consiste na transferência do radical metil, sendo mediada por vitaminas do complexo B (9 e 12).
Transulfunação de Aminoácidos
Consiste na transferência do enxofre entre os aminoácidos, sendo mediada também pela B6.
www.posugf.com.br
22
Bioenergética, Metabolismo de Carboidratos, Lipídios e Proteínas
A síntese de glicose a partir dos aminoácidos é
chamada de gliconeogênese e ocorre principalmente
no citoplasma do hepatócito pela formação de
fosfoenolpiruvato, que será metabolizado à glicose.
A formação da glicose irá ocorrer a partir da síntese
mitocondrial de malato, que será transportado ao
citoplasma, onde será reoxidado a oxaloacetato pela
malato desidrogenase.
Gliconeogênese
Além da produção de energia, os cetoácidos
poderão ser utilizados para síntese de glicose
(gliconeogênese) e/ ou ácidos graxos (cetogênese).
O único aminoácido que não é convertido à glicose
é a leucina, precursora somente de ácidos graxos.
Os aminoácidos oxidados a acetil-CoA e acetoacetilCoA são os aminoácidos cetogênicos, que, além de
glicose, formam ácidos graxos (Lis, Fen, Trp, Tir, Iso).
O oxaloacetato será descarboxilado e fosforilado
a fosfoenolpiruvato, reação catalisada pela enzima
fosfoenolpiruvato carboxiquinase (PEPCK), que será
convertido a glicose-6-fosfato, e esta, desfosforilada
pela glicose-6- fosfatase à glicose.
Os aminoácidos desaminados a piruvato,
oxaloacetato, fumarato, a-cetoglutarato e succinilCoa são os glicogênicos e serão utilizados para
síntese de glicose (Ala, Cis, Gli, Ser, Asp, Asg, Met,
Tre, Val, Arg, Glu, Gln, His, Pro).
www.posugf.com.br
23
Bioenergética, Metabolismo de Carboidratos, Lipídios e Proteínas
MCARDLE, W. D.; KATCH, F. I.; KATCH, V. L.
Fisiologia do exercício. 4ª ed. Rio de Janeiro:
Guanabara Koogan, 1998.
REFERÊNCIAS
BUTTERFIELD, G. E. Whole body protein
utilizations in humans. Medicine and Science in
Sports and Exercise, v. 19, supl. 157, 1987.
MURRAY, R. K.; GRANNER, D. K.; MAYES, P. A.;
RODWELL, V. W. Harper: Bioquímica. São Paulo:
Atheneu, 2006.
BUTTERFIELD-HODGEN, G.; CALLOWAY, D. H.
Protein utilization in men under two conditions
of energy balance and work. Federation
Proceedings, n. 39, 1997. p. 377.
PÓVOA, H. Radicais livres em patologia
humana. Rio de Janeiro: Imago, 1995.
COZZOLINO, S. M. F. Biodisponibilidade de
nutrientes. São Paulo: Manole, 2009.
PELLET, P. L. Protein requirements in humans.
American Journal of Clinical Nutrition. n.51, 1990. p.
723.
DURNING, J. V. P. A. Protein requiriments
and physical activity. In: PARIZKOVA, Jana;
ROGOZKIN, V. A. (ed.) Nutrition, physical fitness and
health. Baltimore: University Park Press, 1978.
PINHEIRO, A. B. V. Tabela para avaliação de
consumo alimentar em medidas caseiras. 2ª
ed. Rio de Janeiro: s.n, 1994.
SHILS, M. E.; OLSON, J. A.; SHIKE, M.; ROSS, A.
C. Tratado de nutrição moderna na saúde e na
doença. São Paaulo: Manole, 2003.
HALLIWELL, B.; GUTTERIDGE, J. M. C. Free
radicals in biology and medicine. New York:
Oxford University Press, 2000.
STRYER, Lubert. Bioquímica. Rio de Janeiro:
Guanabara Koogan, 2008.
HENDLER, S.S. Enciclopédia de vitaminas e
minerais. Rio de Janeiro: Campus, 1994.
WALBERG, J. L. et al. Macronutrient content of a
hipoenergy diet affects nitrogen retention and
muscle function in weight lifters. International
Journal of Sports Medicine. n. 9, 1998. p. 261.
HICKSON, J. F. et al. Repeated days of body
building exercise do not enhance urinary
nitrogen excretions from untrained young
males. Nutrition Research. n. 10, 1990. p. 723.
KRAUSE, M. V.; MAHAN, L. K. Alimentos,
nutrição e dietoterapia. São Paulo: Roca, 2010.
LEHNINGER, A. L. Princípios de bioquímica.
São Paulo: Sarvier, 2010.
MARZZOCO, A.; TORRES, B. Bioquímica básica.
2ª ed. Rio de Janeiro: Guanabara Koogan, 1999.
MAUGHAN, R.; GLEESON, M.; GREENHAFF, P. L.
Bioquímica do exercício e do treinamento. São
Paulo: Manole, 2000.
www.posugf.com.br
24