Digestão e absorção dos glicídeos; Rui Fontes
Estrutura, digestão e absorção dos hidratos de carbono
1.
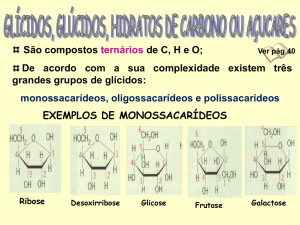
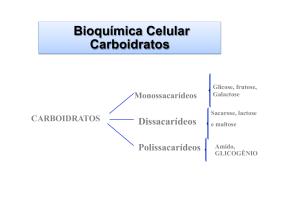
Os hidratos de carbono (ou glicídeos) costumam ser definidos como derivados aldeídicos
ou cetónicos de poliálcoois. Os mais simples são os monossacarídeos de que são
exemplos a glicose, a galactose, a frutose, a manose, a ribose, a desoxiribose, a xilulose,
a ribulose, a eritrose, a sedoheptulose, o ácido glicurónico, o ácido L-idurónico, o
ácido N-acetil-neuramínico, a L-fucose, etc. Nos seres vivos, excetuando os casos do
ácido L-idurónico e da L-fucose, todos os outros monossacarídeos só existem na forma D.
Com raríssimas exceções só um dos enantiómeros das moléculas quirais é sintetizado pelos
seres vivos. Assim, nos seres vivos todo o ácido idurónico é o enantiómero L e toda a
fucose é fucose L; em todos os outros monossacarídeos acima referidos, o enantiómero
existente nos seres vivos é o enantiómero D. Por isso quando, em biologia, nos referimos,
por exemplo, à glicose (ou a outros monossacarídeos) omite-se que se trata do enantiómero
D; toda a glicose existente nos seres vivos é glicose D, toda a galactose é galactose D, etc.
Um determinado enantiómero de um monossacarídeo é o enantiómero D, quando se
observa que, na molécula orientada de acordo com a convenção de Fischer, o grupo
hidroxilo do último carbono assimétrico fica voltado para a direita. A estrutura molecular
dos monossacarídeos, assim como a de todas as substâncias, está na base da sua capacidade
para se ligar e interagir com as enzimas e os transportadores presentes nas células dos seres
vivos. As enzimas e os transportadores membranares dos seres vivos são de natureza
proteica e são incapazes de se ligar à glicose L e a outros monossacarídeos de tipo L que,
de facto, só existem porque podem ser sintetizadas laboratorialmente.
2.
No ser humano saudável, exceto a seguir a refeições que contenham frutose ou galactose
(ou dissacarídeos que lhe possam dar origem), o único monossacarídeo que existe (em
concentrações detetáveis) no plasma sanguíneo é a glicose. O outro monossacarídeo que
também existe na forma livre nos seres humanos é a frutose que está presente no esperma.
Os outros monossacarídeos referidos acima só existem no ser humano como resíduos
componentes de moléculas mais complexas. Estas moléculas mais complexas podem ser
intermediários do metabolismo onde são frequentes as formas fosforiladas (como a
ribulose-5-fosfato ou a xilulose-5-fosfato) ou ligadas de forma covalente a nucleotídeos
(como a UDP-galactose ou a GDP-manose; respetivamente, o uridino-difosfato de glicose
e a guanosino-difosfato de manose). Noutros casos estas moléculas mais complexas são
substâncias de reserva energética ou componentes estruturais e funcionais das células ou da
matriz extracelular: o glicogénio, os ácidos nucleicos, os glicolipídeos, as glicoproteínas
e os glicosaminoglicanos contêm resíduos de monossacarídeos incorporados nas suas
estruturas moleculares. O glicogénio é um polímero ramificado em que as unidades
estruturais são resíduos de glicose ligados entre si por ligações glicosídicas α-1,4 e, nos
locais das ramificações, por ligações glicosídicas α-1,6. Nos glicolipídeos, glicoproteínas e
glicosaminoglicanos é frequente que, pelo menos, alguns dos monómeros de glicose e de
galactose tenham um dos grupos hidroxilo substituído por um grupo amina (glicosamina e
galactosamina). Na maioria dos casos, estes grupos amina estão ligados por ligações amida
a grupos acetilo formando monómeros de N-acetilglicosamina ou N-acetilgalactosamina. Os glicosaminoglicanos são longos polímeros lineares existentes na matriz
extracelular em que, com uma exceção (o ácido hialurónico), o monómero de uma
extremidade da cadeia se liga, através de uma ligação glicosídica, a um resíduo
aminoacídico de uma proteína: assim, com a exceção referida, os glicosaminoglicanos são
se facto proteoglicanos. Nos glicosaminoglicanos há uma unidade estrutural que se repete
formando cadeias lineares (não ramificadas); essa unidade é um dissacarídeo que difere
nos diferentes tipos de glicosaminoglicanos. Com uma exceção (sulfato de queratano)
todos os glicosaminoglicanos podem conter resíduos de ácido glicurónico que se ligam a
resíduos de um aminoaçúcar. Alguns glicosaminoglicanos também contêm outro ácido
urónico, o ácido L-idurónico que (nos casos da heparina, do sulfato de heparano e do
sulfato de dermatano) substitui o ácido glicurónico na maioria das unidades dissacarídicas.
Página 1 de 5
Digestão e absorção dos glicídeos; Rui Fontes
3.
A glicose, a galactose e a frutose são hexoses, ou seja, contêm 6 carbonos; um desses
carbonos contém um grupo carbonilo e os outros carbonos grupos hidroxilo. Na forma
cíclica (a mais abundante, mas em equilíbrio com a forma linear) o grupo carbonilo e um
grupo hidroxilo (geralmente o do carbono 5) estão “escondidos”: na forma cíclica há um
rearranjo molecular designado por ligação semiacetal (ou semicetal) e o carbono que
continha o grupo carbonilo passa a designar-se de anomérico. A glicose e a galactose são
aldoses porque o grupo carbonilo é um aldeído. A frutose é uma cetose porque o grupo
carbonilo (presente em C2) é uma cetona.
4.
O amido, a sacarose e a lactose são hidratos de carbono importantes na dieta dos seres
humanos. O amido e a sacarose são de origem vegetal enquanto a lactose é o açúcar do
leite. Todos os outros hidratos de carbono podem estar presentes nos alimentos, mas não
têm, em geral, relevância do ponto de vista nutricional.
5.
O amido é um polímero formado exclusivamente por resíduos de glicose ligados entre si
por ligações glicosídicas de tipo O (ou acetal). O amido natural é formado por dois tipos
de estruturas distintas: a amilose e a amilopectina. Na amilose as ligações são
exclusivamente de tipo α-1,4, enquanto na amilopectina também existem ligações do tipo
α-1,6. Por isso a molécula de amilose é uma cadeia linear de resíduos de glicose e a
amilopectina uma cadeia ramificada com uma estrutura que lembra uma árvore. Quer na
amilose, quer na amilopectina, o número de resíduos de glicose por molécula, sendo
variável, é de vários milhares. Nos dois tipos de ligações (α-1,4 ou α-1,6) um dos carbonos
envolvidos é sempre o carbono anomérico de um resíduo de glicose que está na forma α
(ou seja, é sempre o anómero α da glicose); o outro carbono envolvido pode ser o carbono
4 ou o 6 de outro resíduo de glicose. (O glicogénio só existe nas células animais e a sua
estrutura é muito semelhante à da amilopectina: a diferença é que o intervalo entre
ramificações é de 5-7 unidades de glicose no glicogénio e esse intervalo é muito maior
(cerca de vinte ou mais) no caso da amilopectina.)
6.
A lactose é um dissacarídeo formado por um resíduo de galactose ligado por uma ligação
glicosídica β-1,4 a um resíduo de glicose. A ligação é também glicosídica de tipo O
envolvendo o carbono anomérico de um resíduo de galactose (sempre o anómero β) e o
carbono 4 de um resíduo de glicose. O carbono anomérico do resíduo de glicose está livre.
Para expressar a ideia de que apenas um dos carbonos anoméricos dos resíduos do
dissacarídeo está envolvido na ligação glicosídica usa-se, às vezes, a expressão “ligação
osídeo-ose”.
7.
A sacarose é um dissacarídeo formado por um resíduo de glicose ligado por uma ligação
glicosídica α-1,2 a um resíduo de frutose. Os carbonos 1 na glicose e o carbono 2 na
frutose são os carbonos anoméricos em cada um dos açúcares. Ou seja, ambos os
carbonos anoméricos estão envolvidos na ligação e a ligação é glicosídica de tipo O quer
“quando se olha do lado” da glicose quer “quando se olha do lado” da frutose. Às vezes
usa-se a expressão “ligação osídeo-osídeo” para expressar esta ideia.
8.
Durante a digestão dos glicídeos as hidrólases digestivas catalisam a hidrólise do amido e
dos dissacarídeos da dieta. No processo catalítico rompem-se as ligações glicosídicas e
libertam-se os monómeros constituintes. As reações catalisadas por enzimas da classe das
hidrólases podem ser entendidas como processos em que a água é um dos reagentes que,
ao reagir com o substrato da enzima em questão, rompe uma ligação levando à libertação
dos resíduos (ou grupos de resíduos) que faziam parte desse substrato (ver Equação 1).
Equação 1
AB + H2O → A + B
As hidrólases que, no tubo digestivo, catalisam a hidrólise dos glicídeos têm pHs ótimos
próximos de 7 o que coincide com o pH existente na boca e no lúmen do intestino (mas
não com o pH do lúmen gástrico que é ácido).
Página 2 de 5
Digestão e absorção dos glicídeos; Rui Fontes
9.
Na hidrólise do amido participam duas isoenzimas1 (as amílases salivar e pancreática) que
catalisam a rotura de ligações glicosídicas α-1,4, mas poupam as ligações α-1,6 assim
como as α-1,4 das extremidades das cadeias ou as que se situam na vizinhança das ligações
α-1,6. Assim, da ação das amílases digestivas sobre o amido, resulta a formação de
maltose (α-D-glicopiranosil-(1→4)-D-glicose), maltotriose (trímero com 3 resíduos
glicose ligados por ligações glicosídicas α-1,4) e dextrinas limite (oligossacarídeo
formado por vários resíduos glicose sendo que dois deles estão ligados por uma ligação α1,6).
10. A hidrólise da maltose, da maltotriose e das dextrinas limite resulta na formação de glicose
e é catalisada por duas dissacarídases: a maltase (α-1,4 glicosídase) e a isomaltase (α-1,6
glicosídase). A expressão “dissacarídases” aplicada à maltase e à isomaltase pode ser
enganadora porque, na verdade, estas enzimas podem atuar em oligossacarídeos com mais
de dois resíduos de glicose. Em ambos os casos liberta-se o resíduo glicose que contribuía
com o carbono anomérico para a ligação glicosídica que foi hidrolisada. O resto da
molécula original contendo um ou mais resíduos de glicose é o outro produto da reação. Só
quando os substratos são os dissacarídeos maltose (caso da maltase; ver Equação 2) ou a
isomaltose (caso da isomaltase; ver Equação 3) é que se libertam duas moléculas de
glicose. As dissacarídases digestivas também incluem a lactase (β-1,4 galactosídase; ver
Equação 4) e a sacarase (ver Equação 5) que, respetivamente, catalisam a hidrólise da
lactose e da sacarose. As dissacarídases estão ancoradas no polo apical da membrana
citoplasmática dos enterócitos2, mas têm o centro ativo mergulhado no lúmen do intestino
delgado. Porque são enzimas da membrana citoplasmática que atuam em substratos que se
situam fora da célula, as dissacarídases são denominadas por ectoenzimas.
Equação 2
Equação 3
Equação 4
Equação 5
maltose + H2O → 2 glicose
isomaltose + H2O → 2 glicose
lactose + H2O → galactose + glicose
sacarose + H2O → frutose + glicose
11. Alguns glicídeos complexos que são componentes de plantas não são digeridos pelas
enzimas presentes na saliva, no estômago ou no intestino delgado e não são absorvidos.
Passam para o intestino grosso (cólon) onde podem ser parcialmente digeridos pelas
bactérias aí presentes e são importantes componentes das fezes. Estes glicídeos são
coletivamente designados por fibras.
12. A síntese de lactase e a quantidade de lactase no polo apical da membrana dos enterócitos
decresce ao longo da vida. Em algumas pessoas essa diminuição da síntese da lactase
começa a manifestar-se ainda na infância (pelos 4-10 anos de idade). A consequente
incapacidade para digerir a lactose designa-se de “intolerância à lactose” e é muito
frequente sobretudo em determinados grupos étnicos3. Os indivíduos com “intolerância à
lactose” têm, após a ingestão de substâncias que contenham lactose, mal-estar abdominal,
aumento da produção de gases e, eventualmente, diarreia. Os gases são produzidos
aquando da fermentação da lactose que é levada a cabo pelas bactérias do intestino grosso.
1
Duas enzimas são isoenzimas quando catalisam a mesma reação mas são, na mesma espécie (o homem,
por exemplo), codificadas por genes distintos sendo, por isso, estruturalmente diferentes.
2
Os enterócitos constituem a maioria das células que forram o intestino delgado. Para além do papel
apontado também é nos enterócitos que ocorre a absorção dos nutrientes. Na membrana do seu polo
apical existem microvilosidades (2000 a 4000 por célula) o que aumenta em dezenas de vezes a superfície
de contacto com o lúmen. Devido ao seu aspeto quando observadas por microscopia eletrónica a
membrana apical dos enterócitos é, frequentemente denominada por “bordadura em escova”.
3
Em Portugal afetará cerca de 1/3 da população adulta; na China mais de 95%.
Página 3 de 5
Digestão e absorção dos glicídeos; Rui Fontes
A diarreia é uma consequência do efeito osmótico da lactose e de produtos resultantes da
fermentação bacteriana4.
13. Da digestão completa do amido, da lactose e da sacarose resulta a formação das oses ou
monossacarídeos (glicose, galactose e frutose) que são absorvidos pelos enterócitos.
14. As moléculas dos monossacarídeos são demasiado hidrofílicas e demasiado grandes para
se poderem dissolver no duplo folheto lipídico das membranas biológicas; assim, a sua
capacidade para atravessar membranas biológicas por mecanismos que não envolvam
transportadores (transporte não mediado ou simples) é praticamente nula. Tal como
acontece em todas as células, nos enterócitos, o transporte transmembranar dos
monossacarídeos é catalisado por moléculas proteicas da membrana que se designam por
transportadores. Quando uma substância atravessa uma membrana e esse transporte é
catalisado por transportadores, o transporte diz-se mediado (mediado por transportadores
de natureza proteica, subentende-se). O transporte de uma substância pode ocorrer a favor
do gradiente (passagem do lado onde está mais concentrada para o lado onde está menos
concentrada – transporte passivo), mas a glicose e a galactose também podem, no polo
apical dos enterócitos (o que está voltado para o lúmen), ser absorvidas contra gradiente
(transporte ativo).
15. No polo apical dos enterócitos, a absorção da glicose e da galactose pode ocorrer contra
gradiente de concentrações e, neste caso, o transporte é indiretamente dependente da
hidrólise de ATP (ATP + H2O → ADP + Pi) e diretamente dependente da existência de
um gradiente de concentração de Na+ (maior concentração de ião Na+ no lúmen
intestinal que no citoplasma).
16. A bomba de sódio-potássio (ou ATPase do Na+-K+) é uma proteína da membrana
citoplasmática de todas as células do organismo que, usando a energia libertada na
hidrólise do ATP, promove a entrada de iões K+ do meio extracelular para o citoplasma e
a saída de iões Na+ do citoplasma para o meio extracelular (ver Equação 6). A bomba de
sódio-potássio é uma máquina biológica que faz a acoplagem de um processo exergónico
(a hidrólise do ATP) com processos endergónicos (o transporte de iões Na+ e K+ contra
gradiente). Porque a energia usada no processo de transporte destes iões provém
diretamente de uma reação química (no caso, a hidrólise do ATP) diz-se que este transporte
ativo é de tipo primário. A existência da bomba de sódio-potássio em todas as células do
organismo permite compreender que a concentração de Na+ seja muito mais baixa dentro
das células que nos líquidos extracelulares (e que o contrário aconteça no caso do K+). A
ação da bomba de sódio-potássio cria gradientes de concentração que explicam a tendência
para a entrada do Na+ para dentro das células.
Equação 6
ATP + H2O + 3 Na+ (citoplasma) + 2 K+ (extracelular)
→ ADP + Pi + 3 Na+ (extracelular) + 2 K+ (citoplasma)
17. Alguns transportadores das membranas (designados por simportes) só funcionam se
transportarem duas substâncias diferentes na mesma direção, mas o transporte só é possível
se, pelo menos, no caso duma das substâncias transportadas o processo for exergónico (ou
seja, ocorrer a favor de gradiente). No polo apical dos enterócitos do duodeno e do jejuno
existe um simporte que transporta conjuntamente Na+ e glicose (ou Na+ e galactose) na
mesma direção; porque só transporta glicose se também transportar Na+, designa-se de
SGLT1 (da expressão inglesa “Sodium dependent GLucose Transporter 1”). O SGLT1 faz
a acoplagem entre um processo exergónico (o transporte de Na+ a favor do gradiente) e
outro endergónico (o transporte de glicose contra gradiente); ver Equação 7. A existência
4
A fermentação bacteriana de glicídeos também produz ácidos carboxílicos de cadeia curta, sobretudo
acetato (2 carbonos; C2), propionato (C3) e butirato (C4). Alguns (como o acetato e o propionato) são,
em grande parte, absorvidos pelos colonócitos e, depois de passarem para o sangue, são metabolizados
noutras células do organismo. O butirato é um nutriente dos colonócitos, as células epiteliais que forram o
intestino grosso.
Página 4 de 5
Digestão e absorção dos glicídeos; Rui Fontes
do gradiente de Na+ que tende a mover este ião para dentro da célula fornece a energia
que permite o transporte de glicose contra gradiente. Este tipo de transporte ativo
(contra gradiente no caso da glicose) diz-se secundário porque o gradiente do Na+ (que
diretamente fornece a energia necessária para o transporte) foi criado pela ação da bomba
de sódio-potássio, esta sim, diretamente dependente de uma reação química exergónica, no
caso a hidrólise do ATP.
Equação 7
glicose (lúmen) + 2 Na+ (lúmen) → glicose (citoplasma) + 2 Na+ (citoplasma)
18. No polo basal dos enterócitos o transportador da glicose é um uniporte (designado por
GLUT2; da expressão inglesa “GLUcose Transporter 2”) e a energia envolvida no
transporte da glicose é a corresponde ao seu gradiente de concentrações. O transporte
catalisado pelo GLUT2 diz-se passivo (ou, significando o mesmo, ocorre por difusão), isto
é, dá-se sempre a favor do gradiente: dos enterócitos para o sangue durante o período
absortivo e em sentido inverso quando a glicose escasseia nos enterócitos. (Embora a
glicose não seja o principal combustível dos enterócitos, estas células também podem
oxidar glicose.) O GLUT2 também catalisa o transporte de galactose e frutose (das células
para o sangue) no polo basal dos enterócitos durante a absorção intestinal destes
monossacarídeos. Embora o sentido líquido do movimento das moléculas dos
monossacarídeos através do GLUT2 seja determinado pelo gradiente de concentrações, em
indivíduos saudáveis e fora do período absortivo, as concentrações de galactose e frutose
no sangue são praticamente nulas. Por isso, não existe transporte de frutose ou de galactose
no sentido sangue → enterócito.
19. O transporte da frutose ocorre sempre a favor do gradiente. No polo apical dos enterócitos
existe um uniporte que catalisa a entrada de frutose e se designa por GLUT5. Como já
referido, no polo basal o transportador da frutose é o GLUT2.
20. A transcrição do gene que codifica o SGLT1 aumenta quando a concentração de glicose é
elevada no lúmen intestinal. Ou seja, a síntese de SGLT1 aumenta após uma refeição que
contenha glicose ou glicídeos que a possam gerar, mas não é afetada pela concentração de
glicose no sangue. O mesmo acontece no caso do GLUT5 quando, neste caso, a refeição
inclui frutose [1].
21. A existência do SGLT1 tem importantes consequências na terapêutica de diarreias agudas.
Nos países pobres, as diarreias agudas são uma importante causa de mortalidade infantil e a
morte pode ser prevenida mantendo as crianças hidratadas e alimentadas. A atividade da
SGLT1 ao permitir a absorção de Na+ e glicose para os enterócitos (e, em última análise
para o sangue) diminui a osmolaridade do conteúdo do lúmen intestinal aumentando-a no
citoplasma dos enterócitos. Tal como a glicose, também a água não é absorvida por difusão
simples, mas sim através da ação de canais específicos situados na membrana denominados
aquaporinas. A atividade da SGLT1 ao permitir a entrada de substâncias osmoticamente
ativas para dentro dos enterócitos resulta também na absorção de água [1]. Um processo
semelhante, neste caso no sentido enterócitos → sangue e envolvendo o GLUT2 e as
aquaporinas, ocorre no polo basal dos enterócitos. A introdução do uso de soluções
contendo simultaneamente glicose e iões sódio para reidratação oral permitiu baixar a
mortalidade infantil causada por diarreias [2].
1. Frayn, K. N. (2012) Regulação Metabólica. Uma perspetiva focada no organismo humano., U.P.
Editorial, Porto.
2. Drozdowski, L. A. & Thomson, A. B. (2006) Intestinal sugar transport, World J Gastroenterol. 12,
1657-70.
Página 5 de 5