UNIVERSIDADE FEDERAL DE PERNAMBUCO
CENTRO DE CIÊNCIAS BIOLÓGICAS
DEPARTAMENTO DE BIOQUÍMICA
DISCIPLINA BIOQUÍMICA 4 – BQ 004
MANUAL DE AULAS PRÁTICAS
Prof. Levy Dos Santos Guedes
RECIFE
2007
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
2
APRESENTAÇÃO
O objetivo deste manual é tornar o aluno consciente da importância das aulas práticas
para a sua formação profissional, procurando envolvê-lo de forma mais efetiva, através da
aplicação de questionários sobre cada aula prática executada.
As fichas de estudo trazem informações que ajudam o aluno a entender a relação
existente entre a prática realizada e o tema teórico correspondente. Além das aulas práticas, o
manual contem um texto sobre o tema de aula teórica: “Integração e regulação do
metabolismo”. Este tema é abrangente e será apresentado ao aluno na forma de painel
dirigido. O manual ainda contem textos com exemplos clínicos e temas sobre algumas
doenças relacionadas com o metabolismo celular, que serão apresentados pelos alunos na
forma de seminários.
Considerando que o Curso Farmacêutico é um curso essencialmente prático e que a
Bioquímica é uma disciplina bastante árida em seu conteúdo, esperamos que os alunos
possam, com este manual, compreender melhor os assuntos das aulas teóricas através da
execução das práticas e da apresentação dos seminários sobre temas relacionados. Isso tornase imperioso, uma vez que a Bioquímica é uma disciplina fundamental na formação do
Farmacêutico.
Nesta edição estamos introduzindo algumas modificações na metodologia das práticas
e algumas correções no texto.
Recife, 11 de maio de 2007
Prof. Levy dos Santos Guedes
3
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
ÍNDICE
APRESENTAÇÃO ..........................................................................................
2
1. Cinética de Reações Catalisadas por Enzimas .............................................
4
2. Inibição de Reações Catalisadas por Enzimas .............................................
8
3. Consumo de Glicose por Células de Sacharomyces cerevisiae ...................
10
4. Dosagem de Peróxidos de Lipídeos .............................................................
13
5. Efeito da Glicose sobre a Peroxidação de Lipídeos .....................................
17
6. Hidrólise de Proteínas ...................................................................................
18
7. Dosagem de Uréia no Plasma Sangüíneo .....................................................
20
ESTUDOS DIRIGIDOS ...................................................................................
22
PAINEL DIRIGIDO
1. Integração e Regulação Metabólica ..............................................................
24
2. Questões Sobre o Tema Integração e Regulação Metabólica .......................
33
BIBLIOGRAFIA ................................................................................................ 34
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
4
CINÉTICA DE REAÇÕES CATALISADAS POR ENZIMAS
OBJETIVOS
A realização desta prática permite ao aluno:
1- Uma introdução ao estudo da cinética de reações catalisadas por enzimas
2- Medir a velocidade de uma reação enzimática.
3- Calcular a constante de Michaelis-Menten (KM) e a velocidade máxima (Vmáx) e
destacar a importância destes parâmetros na caracterização das enzimas.
4- Conferir a importância da equação de Lineweaver-Burk no cálculo de KM e Vmáx.
INTRODUÇÃO
Cinética de reações enzimáticas
As enzimas desempenham um papel de destaque no organismo vivo, uma vez que
catalisam reações bioquímicas imprescindíveis ao desenvolvimento e manutenção das células.
A concentração de uma enzima em tecido ou fluido biológico pode ser determinada
medindo-se a velocidade da reação catalisada pela enzima. Para isso são utilizados métodos
analíticos que permitam medir a diminuição na concentração do substrato ou o aumento na
concentração do produto da reação.
Muitos fatores podem afetar a atividade enzimática: concentração do substrato,
temperatura, pH, concentração de ativadores e inibidores, concentração do produto. Todas
estas variáveis podem e devem ser controladas, mas a presença de ativadores e inibidores em
sistemas biológicos nem sempre é detectada e por isso não pode ser controlada. Por isso a
medida da atividade enzimática não corresponde necessariamente à concentração real da
enzima. Para medir com precisão a velocidade de uma reação enzimática é necessário que esta
velocidade se mantenha constante durante o experimento.
Com auxílio da equação de Michaelis-Menten é possível determinar o percentual da
velocidade de uma reação enzimática com uma dada concentração de substrato, em relação à
velocidade máxima da reação, ou seja, à velocidade obtida com uma concentração de
substrato capaz de saturar toda a enzima presente no meio da reação. Assim, se em uma
reação enzimática for utilizada uma concentração de substrato correspondente a 20 vezes o
valor do KM, a velocidade da reação corresponderá a 95% da Vmáx:
v = Vmáx [S] (KM + [S]) v Vmáx100 = 20 KM (KM + 20 KM)
Admitindo que no final da reação 10% do substrato tenha sido consumido, a
concentração agora corresponderá a 18 vezes o valor do KM e a velocidade da reação a 94,7%
da Vmáx. A variação da velocidade é muito pequena e pode ser ignorada.
Na prática esse problema torna-se mais complexo uma vez que a solubilidade do
substrato é limitada e muitas vezes as enzimas apresentam um KM alto. Nestes casos é
aconselhável desenvolver a reação enzimática em um espaço de tempo curto e repetir o
experimento com o material que contem a enzima, diluído, todas as vezes em que mais de
10% do substrato for consumido na reação.
5
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
Aplicações de enzimas nas análises clínicas
Os conhecimentos da enzimologia vêm sendo aplicados no laboratório clínico para
medir a atividade de enzimas e a concentração de substratos em tecidos ou fluidos de
pacientes. Enzimas como trombina e plasmina, associadas ao processo de coagulação
sangüínea, bem como lipoproteína lipase, envolvida com o processamento dos quilomícrons;
são exemplos de enzimas específicas do plasma e por isso são encontradas em concentrações
mais elevadas. Nas doenças de tecidos e órgãos pode haver alterações na permeabilidade da
membrana, ou morte celular e com isso, enzimas dos tecidos difundem-se para o plasma.
Essas enzimas, normalmente estão presentes em baixas concentrações e não têm nenhum
papel funcional no plasma.
No diagnóstico do envolvimento de um órgão específico numa doença, seria ideal se
as enzimas particulares para cada órgão pudessem ser identificadas. Isso é improvável porque
os processos metabólicos de vários órgãos são muito semelhantes. Embora existam poucas
enzimas específicas para um determinado órgão ou tecido, como a álcool desidrogenase do
fígado e a fosfatase ácida da próstata, o estudo da cinética do aparecimento e
desaparecimento de enzimas particulares no plasma, permite que o diagnóstico do
envolvimento de um órgão específico seja feito.
Estudos da cinética de liberação de enzimas cardíacas no soro, após um enfarte do
miocárdio, permitem estabelecer quando o ataque ocorreu e se o tratamento é efetivo. A
concentração plasmática de creatina fosfocinase (CPK), aumenta cerca de seis vezes, entre o
primeiro e o segundo dia após o enfarte; enquanto a lactato desidrogenase (LDH) e a
-hidroxibutírico desidrogenase (HBDH), aumentam cerca de duas vezes, de forma mais
lenta, mas permanecem elevadas por mais tempo.
Entre as enzimas utilizadas com freqüência no diagnóstico de doenças podemos
relacionar, além daquelas citadas acima, fosfatase alcalina, amilase, lipase, aspartato e
alanina amino transferases, estas duas últimas, também conhecidas como transaminase
glutâmico oxaloacética (TGO) e transaminase glutâmico pirúvica (TGP), respectivamente.
O reconhecimento dos metabólitos que se acumulam em fluidos biológicos, tem um
papel importante na identificação de possíveis defeitos na produção ou na atividade de
enzimas. Como exemplo temos a acidúria orótica hereditária, devida à deficiência de duas
enzimas envolvidas com a via de biossíntese de pirimidinas (orotato fosforibosil transferase e
orotidina), gerando um acúmulo de ácido orótico. A hiperuricemia é um outro exemplo de
aumento de metabólito, devido a deficiência na produção da enzima hipoxantina-guaninafosforribosiltransferase.
Enzimas são utilizadas como reagentes químicos em analisadores clínicos portáteis.
Dosagem de colesterol, triacilgliceróis, glicose, podem ser realizados em poucos minutos,
usando 10L de plasma. A química clínica moderna tem se beneficiado da união entre a
química e a imunologia. Anticorpos específicos contra um antígeno protéico são acoplados a
uma enzima indicadora, como peroxidase de raiz forte (horseradish), gerando um ensaio
muito específico e sensível. Esse ensaio é conhecido como ELISA (enzyme-linked
immunoadsorbent assay). Um exemplo de sua utilização é demonstrado por um ensaio para
identificação dos antígenos protéicos da capa do vírus da imunodeficiência humana (HIV),
que gera a síndrome da imunodeficiência adquirida (AIDS).
A identificação de isoenzimas – enzimas que catalisam a mesma reação, mas migram
diferentemente em um campo eletroforético – também tem sido usada para diagnóstico
clínico. Exemplos de isoenzimas que têm ampla aplicação clínica:
1) Creatina fosfocinase – um dímero com dois tipos de subunidades, M (muscular) e
B (cerebral). No músculo esquelético as duas subunidades são do tipo M. No cérebro as duas
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
6
unidades são do tipo B. Somente no miocárdio se encontra a isoenzima contendo as duas
subunidade M e B. Nos outros tecidos são encontradas quantidades variáveis de isoenzimas
MM e BB. De acordo com a mobilidade para o ânodo, na eletroforese, essas isoenzimas são
denominadas CPK1 (BB), CPK2 (MB) e CPK3 (MM).
2) Lactato desidrogenase, uma enzima tetramérica contendo apenas duas
subunidades diferentes: H, para o coração e M, para o músculo esquelético. São identificadas
cinco formas dessas isoenzimas: LDH1 (HHHH) e LDH2 (HHHM), encontradas no miocárdio
e nos eritrócitos; LDH3 (HHMM), no cérebro e rim; LDH4 (HMMM) e por fim LDH5
(MMMM), encontradas no fígado e músculo esquelético.
Estreptocinase, uma mistura de enzimas obtida de um streptococcus, é utilizada para
a remoção de coágulos sangüíneos, através da ativação do plasminogênio, a forma inativa da
plasmina, no plasma. Uma outra aplicação de enzimas como agentes terapêuticos é a
utilização da asparaginase. Algumas células leucêmicas parecem perder a capacidade de
sintetizar asparagina. Dessa forma, a asparaginase inibe o crescimento de células tumorais,
reduzindo os níveis plasmáticos de asparagina do hospedeiro.
Na indústria farmacêutica, enzimas ligadas a matrizes insolúveis (enzimas
imobilizadas) são utilizadas como reatores químicos altamente específicos. Por exemplo, galactosidase é utilizada para reduzir o conteúdo de lactose no leite.
FUNDAMENTO DA PRÁTICA
Nesta prática utilizaremos a enzima urease (uréia amidohidrolase, EC 3.5.1.5). Esta
enzima apresenta uma alta especificidade para o substrato uréia (NH2CO-NH2),
transformando-o em amoníaco (NH4+) e carbonato (HCO3-), em meio ácido:
UREASE
NH2CO-NH2 + 2H2O + H+ =========> 2NH4+ + HCO3A medida da atividade enzimática pode ser determinada medindo-se a diminuição na
concentração de uréia ou o aumento de amoníaco ou carbonato formado.
Neste experimento utilizaremos a reação proposta por Berthelot para determinar os
níveis de amoníaco formado. Em meio alcalino, amoníaco é transformado em amônia (NH3) e
reage com um derivado fenólico (salicilato) na presença de hipoclorito de sódio produzindo
um composto de cor azul, o indofenol, cuja intensidade é proporcional à concentração de
amônia. Nitroprussiato de sódio é utilizado como catalisador desta reação.
7
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
PROCESSO
Identificar sete tubos de ensaio: Branco (B) e mais seis tubos enumerados de 1 a 6 e
seguir a orientação abaixo:
TUBOS
B
1
2
3
4
5
6
ÁGUA
DEIONIZADA
0,4 mL
0,3 mL
0,2 mL
0,3 mL
0,2 mL
-
SUBSTRATO (URÉIA)
8,56%
85,6mg%
0,1 mL
0,2 mL
0,4 mL
0,1 mL
0,2 mL
0,4 mL
ENZIMA (UREASE)
(268 U/ml)
1,0 mL
1,0 mL
1,0 mL
1,0 mL
1,0 mL
1,0 mL
1,0 mL
1- Incubar todos os tubos em banho-maria (BM) a 37°C por 5 minutos (esta é a reação
enzimática).
2- Adicionar a cada tubo 1,0 mL da solução oxidante (hidróxido de sódio 2,8 mM e
hipoclorito de sódio 121 mM), misturar e levar ao BM a 37°C por 5 minutos..
3- Ler as absorções ópticas em 600 nm, utilizando o branco para ajustar o zero do
espectrofotômetro.
CÁLCULO DA ATIVIDADE ENZIMÁTICA
Construir um gráfico contendo no eixo das abcissas as concentrações do substrato [S]
(0,1; 0,2; 0,4; 1,0; 2,0 e 4,0 mM)* e no eixo das ordenadas a leitura das absorções (v).
Construir um novo gráfico contendo nos eixo das abscissas, 1/[S] e nas ordenadas, 1/v.
Calcular KM e Vmáx.
OBSERVAÇÃO
A urease (268 U/ml) está dissolvida com tampão fosfato 10 mM, pH 6,9; salicilato de
sódio 31,2 mM e nitroprussiato de sódio 1,68 mM.
* -Considerando que 0,1ml da solução de uréia, a 8,56%, contem 8,56 μg de uréia.
-Considerando, ainda, que essa massa do substrato está dissolvida em um volume final
(volume dos reagentes) de 1,4ml.
-Em 1000ml da preparação teremos 6,11mg de uréia, cuja massa molecular é 60,06 daltons.
-6,11 mg/60,06mg por litro corresponde, aproximadamente, a uma concentração 0,1mM.
8
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
INIBIÇÃO DE REAÇÕES CATALISADAS POR ENZIMAS
OBJETIVOS
A realização desta prática permite ao aluno:
1- Identificar o efeito de inibidores sobre a velocidade de uma reação enzimática.
2- Conferir a importância do gráfico duplo recíproco, proposto por Lineweaver-Burk,
na identificação do tipo de inibidor de uma reação enzimática.
INTRODUÇÃO
Moléculas diferentes do substrato podem interagir com a enzima levando a uma
redução de sua atividade. O estudo da inibição enzimática é de interesse, porque revela
informações acerca do mecanismo de ação da enzima. Muitas substâncias tóxicas, incluindo
drogas, expressam sua ação no organismo através da inibição de enzimas.
A inibição reversível é descrita pelo equilíbrio da interação enzima inibidor. Os
processos de inibição enzimática podem ser classificados como competitivo e não
competitivo, dependendo da forma como o inibidor age sobre a enzima. O inibidor
competitivo, geralmente tem uma estrutura semelhante à do substrato e pode ligar-se ao sítio
ativo da enzima, mas não pode ser transformado. Um inibidor não competitivo não se liga no
sítio ativo da enzima, mas liga-se em outra região da molécula enzimática e impede que a
enzima reconheça o substrato.
Para distinguir cineticamente, os dois processos de inibição enzimática, aplica-se o
gráfico proposto por Lineweaver-Burk aos resultados obtidos experimentalmente. A enzima,
numa concentração constante, reage com concentrações crescentes do substrato na ausência
do inibidor e em um segundo experimento a enzima reage com o substrato em concentrações
crescentes na presença do inibidor.
FUNDAMENTO DA PRÁTICA
A realização desta prática é muito semelhante à anterior. Utilizaremos a enzima urease
(uréia amidohidrolase, EC 3.5.1.5), em presença do substrato – uréia – mas utilizaremos
cloreto de mercúrio (HgCl2) como inibidor da enzima.
PROCESSO
Identificar sete tubos de ensaio: Branco (B) e mais seis tubos enumerados de 1 a 6 e
seguir a orientação abaixo:
TUBOS
ÁGUA
DEIONIZADA
INIBIDOR
(HgCl2 0,05%)
B
1
2
3
4
5
6
0,4 mL
0,3 mL
0,2 mL
0,3 mL
0,2 mL
-
0,1mL
0,1mL
0,1mL
0,1mL
0,1mL
0,1mL
0,1mL
SUBSTRATO (URÉIA)
8,56mg%
85,6mg%
0,1 mL
0,2 mL
0,4 mL
-
0,1 mL
0,2 mL
0,4 mL
ENZIMA
(UREASE)
(268 U/ml)
1,0 mL
1,0 mL
1,0 mL
1,0 mL
1,0 mL
1,0 mL
1,0 mL
9
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
1- Incubar todos os tubos em banho-maria (BM) a 37°C por 5 minutos (esta é a reação
enzimática).
2- Adicionar a cada tubo 1,0 mL da solução oxidante (hidróxido de sódio 2,8 mM e
hipoclorito de sódio 121 mM), misturar e levar ao BM a 37°C por 5 minutos..
3- Ler as absorções ópticas em 600 nm, utilizando o branco para ajustar o zero do
espectrofotômetro.
CÁLCULO DA ATIVIDADE ENZIMÁTICA
Construir um gráfico contendo no eixo das abcissas as concentrações do substrato [S]
(0,1; 0,2; 0,4; 1,0; 2,0 e 4,0 mM) e no eixo das ordenadas a leitura das absorções (v), deste
experimento e dos valores da prática anterior.
Construir um novo gráfico contendo nos eixo das abcissas 1/[S] e nas ordenadas 1/v.
Calcular KM e Vmáx, com os valores obtidos nesta prática e na prática anterior.
Com os resultados, definir o tipo de inibição do cloreto de mercúrio sobre a urease.
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
10
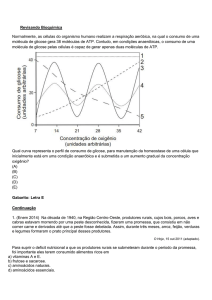
CONSUMO DE GLICOSE POR CÉLULAS DE Sacharomyces cerevisiae
OBJETIVOS
1- Demonstrar o consumo de glicose pelas células.
2- Determinar os níveis de glicose utilizando uma técnica espectrofotométrica.
INTRODUÇÃO
S. cerevisiae são organismos unicelulares, pertencentes à classe dos ascomicetes
(asco= órgão produtor de esporos) e gênero Sacaromicete, utilizados em larga escala na
produção de álcool e bebidas fermentadas e também na panificação. Essas leveduras estão
entre os mais importantes fungos domesticados e metabolizam a glicose gerando etanol e
dióxido de carbono. O pão assado adquire uma textura suave, devido a formação de bolhas de
dióxido de carbono na massa.
Para este experimento poderíamos utilizar as hemácias. Nos mamíferos, a hemácia
madura não contem núcleo, nem mitocôndrias, nem ribossomos e por isso não é capaz de
desempenhar as mesmas atividades metabólicas das células nucleadas. No entanto, o ganho de
energia na hemácia faz-se, fundamentalmente, pela glicólise. Essa energia é utilizada na
manutenção das condições osmóticas, da forma discóide e do ferro no estado ferroso. A
glicose desaparece rapidamente no sangue, quando deixado fora dos vasos sangüíneos e
nessas condições há um aumento nos níveis de ácido láctico.
Na prática do laboratório clínico, o sangue colhido para a dosagem de glicose deve ter
o plasma separado imediatamente das hemácias, ou ao sangue total acrescenta-se fluoreto que
é um inibidor da enolase, enzima que catalisa a seguinte reação na via glicolítica:
2-fosfoglicerato fosfoenolpiruvato + H2O
Este experimento nos permite avaliar o consumo de glicose, in vitro, medindo a
redução nos níveis de glicose com o tempo de incubação.
SUSPENSÃO DE CÉLULAS DE Sacharomyces cerevisiae
Dissolver 15g de células em 100 mL de solução fisiológica tamponada, pH 7,4 (PBS).
PROCESSO
Preparar três tubos de ensaio, identificados com os números 0, 5 e 10, respectivamente e
proceder conforme esquema:
1- Adicionar a cada tubo 0,5mL da suspensão de células + 0,2mL da solução de glicose a
10% + 1,0mL de PBS.
2- Incubar os tubos 5 e 10 em banho-maria a 37o C, por 5 e 10 min, respectivamente.
OBS. Para utilizar o MÉTODO DE DOSAGEM QUÍMICA seguir o item 3. Para
dosagem pelo MÉTODO DE DOSAGEM ENZIMÁTICA seguir a metodologia, adiante.
3- Transferir alíquotas de 0,2 mL (em duplicata) dos tubos 0, 5 e 10 para tubos de centrífuga,
previamente identificados (0a e 0b, 5a e5b, 10a e 10b), contendo 1,8 mL de ácido
tricloroacético (TCA), a 10%. Deixar os tubos em repouso por 10 min, agitando-os
11
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
esporadicamente e centrifugar a 3000 rpm por 5 minutos. Transferir, por decantação, o
sobrenadante para tubos limpos, previamente identificados (0a e 0b, etc), que será
utilizado na dosagem dos níveis de glicose, pelo método de dosagem química.
MÉTODO PARA DOSAGEM DE GLICOSE
1 – Método de dosagem química
Fundamento - O grupamento amino da orto-toluidina reage com o grupamento aldeído da
glicose, em solução de ácido acético glacial a quente e dá origem a um composto de cor
verde, estável. A intensidade de cor é proporcional à concentração de glicose na amostra e sua
estabilidade depende do equilíbrio entre uma glicosilamina (-NH-CHOH-) e a base de Schiff
(-N=CH-) correspondente.
Solução padrão de glicose - 100mg de glicose em 100 mL de solução. Para a dosagem diluir
a solução padrão 1:10 com água destilada.
Dosagem de glicose - Identificar nove tubos (B, P1, P2, 0a, 0b, 5a, 5b, 10a e 10b) - Branco,
Padrões e Testes.
Transferir 1 mL de cada sobrenadante límpido para os respectivos tubos de ensaio
previamente identificados.
Adicionar a cada tubo:
TUBO
SOLUÇÃO
TESTES
ÁGUA
REAGENTE DA
PADRÃO
(0a, 0b, 5a, 5b,
DESTILADA
ORTO-TOLUIDINA
DILUÍDA
10a e 10b)
Padrões
1 mL
3,5 mL
Testes
1 mL
3,5 mL
Branco
1 mL
3,5 mL
Aquecer os tubos em banho-maria fervente por 10 minutos e resfriá-los em água
corrente.
Ler as densidades ópticas (DO) da solução em cada tubo, no espectrofotômetro em
630 nm. Ajustar o zero do aparelho com o Branco.
Calcular a concentração de glicose em cada amostra e expressar os valores em mg%.
Construir um gráfico das DO em função do tempo de incubação.
CONCLUSÕES:
2- Método de dosagem enzimática
Fundamento - Duas enzimas estão envolvidas neste processo: glicose oxidase, que age sobre
a glicose e gera ácido glicônico e peróxido de hidrogênio. E peroxidase, que utiliza o
peróxido de hidrogênio como agente oxidante para transformar 4-aminoantipirina e fenol em
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
12
antipirilquinonimina e água. Antipirilquinonimina é um composto colorido que absorve em
505 nm. A intensidade de cor é proporcional à quantidade de peróxido de hidrogênio gerado
na primeira reação.
Dosagem de glicose - Identificar nove tubos (B, P1, P2, 0a, 0b, 5a, 5b, 10a e 10b) - Branco,
Padrões e Testes.
Transferir alíquotas de 0,05 mL para os respectivos tubos de ensaio previamente
identificados:
TUBO
SOLUÇÃO
TESTES
ÁGUA
REAGENTE
PADRÃO
(0a, 0b, 5a, 5b,
DESTILADA
(100mg%)
10a e 10b)
Padrões
0,05 mL
2,5 mL
Testes
0,05 mL
2,5 mL
Branco
0,05 mL
2,5 mL
Misturar e incubar em banho-maria a 37°C durante 15 minutos.
O nível de água do banho deve ser superior ao nível da mistura em cada tubo de
ensaio.
Ler as densidades ópticas (DO) da solução em cada tubo, no espectrofotômetro em
505 nm. Ajustar o zero do aparelho com o Branco.
Calcular a concentração de glicose em cada amostra e expressar os valores em mg%.
Construir um gráfico das concentrações de glicose em função do tempo de incubação.
CONCLUSÕES:
13
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
DOSAGEM DE PERÓXIDOS DE LIPÍDEOS
OBJETIVOS
A realização desta prática permite ao aluno determinar os níveis de substâncias que
reagem com o ácido tiobarbitúrico em células de Sacharomyces cerevisiae e avaliar o grau da
peroxidação dos lipídeos.
INTRODUÇÃO
A peroxidação de lipídeos é um processo complexo onde ácidos graxos insaturados e
outros lipídeos são oxidados por ação dos radicais livres. Para entender a reatividade destas
espécies químicas, devemos lembrar que os elétrons, nas moléculas, geralmente se reúnem em
pares. Na molécula de água, por exemplo, o oxigênio se apresenta rodeado por quatro pares
de elétrons; dois pares se ligam fracamente em uniões OH e dois pares não estão ligados:
Para descrever completamente um elétron em um átomo devemos considerar, além dos
seus números quânticos n (principal), (secundário) e m, um quarto número quântico ms,
chamado número quântico de spin; que está associado com a rotação do elétron em torno do
seu próprio eixo e pode ter um dos dois valores possíveis: + ½ ou - ½ , dependendo da direção
da rotação do elétron. Dois elétrons em um mesmo orbital com valores de ms + ½ ou - ½ são
ditos emparelhados. Um par eletrônico é mais estável do que os elétrons isolados, uma
vez que, aparentemente, dois elétrons com spins opostos têm seus campos magnéticos
recíprocos anulados.
Radicais livres podem ser definidos como átomos, moléculas ou fragmento molecular
com, pelo menos, um elétron desemparelhado. A presença do elétron desemparelhado, em
geral, confere ao radical livre alta reatividade química e, ainda, certas propriedades como
características paramagnéticas. Para restaurar sua estabilidade estas espécies químicas tendem
a retirar um elétron de outra substância ou doar o seu elétron desemparelhado. Com isso
podem gerar novos radicais livres, numa reação em cadeia.
No organismo vivo a produção de radicais livres é permanente, mas esta produção está
associada ao metabolismo celular do oxigênio e às reações de óxido-redução. Entre as fontes
geradoras de radicais livres destacam-se:
A cadeia respiratória mitocondrial - A mitocôndria é considerada a “casa de força” da
célula, aí se encontra um complexo sistema de transferência de elétrons, conhecido como
Cadeia Transportadora de Elétrons (CTE). A função deste sistema é reduzir o oxigênio ao
mesmo tempo em que gera energia, que é armazenada sob a forma de ATP. Embora os
elétrons sejam transferidos aos pares, os citocromos integrantes da CTE só transferem um
elétron de cada vez. Quando uma molécula de oxigênio recebe um elétron é transformada no
ânion superóxido (O2-.) - um radical livre. Cerca de 5% do ânion superóxido, gerado no
processo de redução do oxigênio, pode ser liberado da CTE e, por dismutação, gera peróxido
de hidrogênio (H2O2).
Os radicais livres derivados da redução do oxigênio são os produtos fisiológicos
potencialmente mais perigosos, gerados a partir da respiração celular.
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
14
A fagocitose - Em estado de repouso os neutrófilos consomem pouco oxigênio. Em
contato com partículas a serem fagocitadas, eles produzem uma invaginação de sua
membrana, encerrando assim o material a destruir (fagossoma), isolando-o do citoplasma.
Essa estimulação dos neutrófilos é acompanhada de uma aceleração no consumo de oxigênio
(“choque respiratório”), com ativação de uma enzima de membrana - a NADPH-oxidase - que
catalisa a redução do oxigênio, gerando o ânion superóxido, que por dismutação gera
peróxido de hidrogênio.
Estes dois agentes oxidantes (O2-. e H2O2) participam da produção de espécies
químicas ativas, como o radical hidroxil (OH.) e na liberação de hipoclorito e de cloraminas,
sob a influência de uma enzima leucocitária, a mieloperoxidase. Este conjunto de reagentes
agressivos (ânion superóxido, peróxido de hidrogênio, hipoclorito e cloraminas) liberados nos
fagossomas são responsáveis pela destruição do material fagocitado. Na ausência de NADPHoxidase, a atividade fagocitária torna-se diminuída ou mesmo abolida.
As reações de desintoxicação - Os
peroxissomos são organelas celulares que
desempenham um papel importante na desintoxicação de numerosas moléculas. Uma
variedade de substâncias tóxicas são absorvidas com freqüência através do trato
gastrointestinal, ou são produzidas a partir do metabolismo celular. Estas substâncias podem
sofrer modificações em sua estrutura química (o processo de desintoxicação) e em seguida
devem ser excretadas, sobre tudo, pela urina.
Os mecanismos de desintoxicação utilizados no fígado são diversificados, envolvendo
oxidação, redução, hidrólise, conjugação, ou uma combinação destes mecanismos. Em geral,
a oxidação é o primeiro dos mecanismos utilizados, seguido às vezes de reação de
conjugação.
A desintoxicação por oxidação, implica na hidroxilação, que é catalisada por um
sistema “oxidase de função mista” presente nos peroxissomos. Este sistema é constituído de
uma hidroxilase, citocromos P450, NADPH, ferredoxina e FAD. Neste sistema o citocromo
P450 transfere um elétron, de cada vez, para o oxigênio molecular, o que favorece a produção
de ânions superóxido e peróxido de hidrogênio. Este último é desdobrado a água e oxigênio
pela catalase, presente nessas organelas.
A síntese de prostaglandinas - As prostaglandinas, como a prostaciclina e o tromboxano
A2, têm sua origem no ácido araqüidônico, liberado dos fosfolipídeos integrantes de
membrana, por ação da fosfolipase A2.
Os radicais hidroxil são gerados no processo de síntese das prostaglandinas, na fase de
transformação do ácido araqüidônico a endoperóxidos, por ação da ciclooxigenase.
Estes radicais livres intervêm secundariamente sobre a cascata do ácido araqüidônico,
inibindo a ciclooxigenase e promovendo a via metabólica pró-agregante do tromboxano A2
sobre a via anti-agregante e vasodilatadora da prostaciclina.
As radiações - Raios gama e raios-X são exemplos de radiações ionizantes. Quando uma
molécula colide com essas radiações absorve energia e pode emitir elétrons com alto nível
energético.
O material celular está fundamentalmente distribuído em uma fase aquosa. A
transferência da energia para a molécula de água excita os elétrons constituintes e gera uma
molécula de água excitada (com um excedente de energia, comparada com sua energia
normal). A conseqüência mais provável deste evento é uma ionização, como segue:
(1)
H2O H2O+ + e+
(2)
H2O + H2O H2O+ + OH.
15
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
ambas as espécies (H2O+ e OH.) têm um elétron desemparelhado, são radicais livres e por isso
são espécies extremamente reativas. Outras espécies reativas podem ser geradas.
O elétron da reação (1) - elétron hidratado - pode reagir com oxigênio presente na
célula e gerar ânion superóxido. Admite-se que cerca de 10-5 segundo é o tempo decorrido
entre a ação inicial dos radicais sobre os constituintes em uma solução e a formação de
produtos. Vale ressaltar, no entanto, que essas espécies reativas têm um tempo de ação muito
curto. Elétrons produzidos na reação (1) perdem sua energia para as espécies químicas do
meio, em uma escala de tempo da ordem de 10-13 a 10-11 segundo.
Quando uma molécula absorve um fóton de luz ultravioleta, cuja energia é inferior à
sua energia de ionização, ela deverá se excitar. Se a energia armazenada for suficiente, pode
haver ruptura de ligações e produção de radicais livres por diversos mecanismos.
Os tecidos mais suscetíveis ao efeito dessas radiações são a pele e sobretudo os olhos,
devido à sua exposição direta e à intensidade de seu metabolismo. De forma geral, os radicais
livres exercem vários efeitos citotóxicos, envolvendo, virtualmente todos os componentes
celulares. São capazes de promover eventual destruição de estruturas e tecidos, causando
doenças genéticas, genético ambientais e ambientais, a exemplo da arteriosclerose,
hipertensão, câncer, osteoartrite, entre outras, além da liberação de uma variedade de produtos
de degradação como cetonas, álcoois, aldeídos e ésteres, propagando-se pela circulação. As
membranas biológicas de um modo geral, oferecem um rico meio para o ataque peroxidativo
que interfere na integridade das células.
Malondialdeído (MDA), formado a partir da quebra de ácidos graxos poliinsaturados,
é identificado como o principal produto da peroxidação de lipídeos e serve como índice
determinante da extensão da reação peroxidativa. O MDA reage com o ácido tiobarbitúrico
(TBA) produzindo um pigmento de cor avermelhada que absorve em comprimento de onda de
535 nm. O pigmento formado é resultado da condensação de 2 moles de TBA com 1 mol de
MDA. Esta reação não é específica para quantificar MDA, uma vez que existem outras
substâncias que reagem com o TBA (TBARS), mas vem sendo amplamente utilizada para
avaliar o grau de peroxidação de lipídeos.
DOSAGEM DE TBARS
Método de Buege JA & Aust SD (Methods in Enzymology 52:302-310, 1978)
REAGENTE: Solução de TBA 0,375% em ácido tricloroacético (TCA) a 15%
PROCESSO:
Identificar quatro tubos de ensaio: 0, 5, 10 e 20.
A cada tubo adicionar 1,0 mL de suspensão de células de S. cerevisiae.
Incubar os tubos respectivos por 5, 10 e 20 minutos a 37o C e em seguida transferir o
conteúdo de cada tubo para tubo de centrífuga contendo 1,0 mL de TCA a 30%.
A suspensão do tubo 0 não será incubada a 37º C, mas deve ser transferida para tubo de
centrífuga contendo 1,0 mL de TCA a 30%.
Depois de transferir as amostras para tubos de centrífuga contendo TCA a 30% Deixa-las em
repouso por um tempo mínimo de10 min, misturando esporadicamente.
Centrifugar todos os tubos a 3000 rpm por 5 min e transferir o sobrenadante límpido para
tubos previamente identificados: 0, 5, 10 e 20.
Preparar mais dois tubos (Branco e Padrão). Ao Branco adicionar 1,0 mL de água destilada e
ao Padrão, 0,1 mL da solução padrão de MDA (32,12nmol/mL) + 0,9 de água destilada.
16
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
Adicionar a todos os tubos 2,0 mL da solução reagente, agitar e levar ao banho fervente por
15 min.
Esfriar os tubos em água corrente e fazer a leitura da absorção óptica em 535nm.
Calcular a concentração (μM) de TBARS em cada experimento, considerando que:
Massa do padrão (3,212nmol)
Massa da amostra ( x nmol )
0,9 ml de suspensão de S. cereviseae
DOpadrão
DOamostra
x nmol de TBARS
Lançar os resultados em um gráfico (concentração vs tempo) e comparar com os resultados
obtidos no próximo experimento.
OBS.: DO – Densidade óptica
17
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
EFEITO DA GLICOSE SOBRE A PEROXIDAÇÃO DE LIPÍDEOS
OBJETIVOS
1- Reconhecer o malondialdeído como produto da peroxidação dos lipídeos.
2- Identificar o efeito da glicose sobre a peroxidação dos lipídeos.
INTRODUÇÃO
Malondialdeído (MDA) é um produto da peroxidação dos lipídeos nos diversos
tecidos e está presente no plasma sangüíneo. Pesquisas têm mostrado que os níveis de MDA
plasmático aumentam com a idade. Esse composto é muito reativo e tem grande afinidade por
grupamentos amino, o que pode favorecer sua ligação com as proteínas da membrana das
células, comprometendo sua perfeita função. Na prática anterior pudemos acompanhar a
produção de peróxidos de lipídeos medindo os níveis de TBARS em células de S. cerevisiae
submetidas ao estresse oxidativo. Nesta prática vamos avaliar os níveis de TBARS em células
de S. cerevisiae com suprimento de glicose, como fonte de energia.
PROCESSO
Identificar quatro tubos de ensaio: 0, 5, 10 e 20.
A cada tubo adicionar 0,9 mL de suspensão de células de S. cerevisiae + 0,1 mL da solução
de glicose a 10%.
Incubar os tubos respectivos por 5, 10 e 20 minutos a 37o C e em seguida transferir o
conteúdo de cada tubo para tubo de centrífuga contendo 1,0 mL de TCA a 30%.
A suspensão do tubo 0 não será incubada a 37º C, mas deve ser transferida para tubo de
centrífuga contendo 1,0 mL de TCA a 10%.
Depois de transferir as amostras para tubos de centrífuga contendo TCA a 10% Deixa-las em
repouso por um tempo mínimo de10 min, misturando esporadicamente.
Centrifugar todos os tubos a 3000 rpm por 5 min e transferir o sobrenadante límpido para
tubos previamente identificados: 0, 5, 10 e 20.
Preparar mais dois tubos (Branco e Padrão). Ao Branco adicionar 1,0 mL de água destilada e
ao Padrão, 0,1 mL da solução padrão de MDA + 0,9 de água destilada.
Adicionar a todos os tubos 2,0 mL da solução reagente, agitar e levar ao banho fervente por
15 min.
Esfriar os tubos em água corrente e fazer a leitura da absorção óptica em 535nm.
Calcular a concentração (μM) de TBARS em cada experimento, considerando que:
Massa do padrão (3,212nmol)
Massa da amostra ( x nmol )
0,9 ml de suspensão de S. cereviseae
DOpadrão
DOamostra
x nmol de TBARS
Lançar os resultados em um gráfico (concentração vs tempo) e comparar com os resultados
obtidos no experimento anterior.
CONCLUSÕES:
18
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
HIDRÓLISE DE PROTEÍNAS
OBJETIVOS
1- Identificar os aminoácidos como unidade fundamental das proteínas
2- Acompanhar a hidrólise de proteínas por aquecimento em meio ácido.
INTRODUÇÃO
Todas as proteínas na natureza têm como unidade fundamental vinte -aminoácidos e
são construídas através de ligações amida entre estes aminoácidos. Quando isolados das
proteínas os aminoácidos apresentam características estruturais comuns, qual seja um grupo
carboxila e um grupo amino ligados ao carbono . O que distingue estes compostos quanto às
suas características físicas, químicas e biológicas é o grupo R, lateral, que é único para cada
aminoácido. É fundamental que se estabeleça métodos práticos usados na separação e
identificação destes compostos, uma vez que a estrutura e funções biológicas de uma proteína
depende do seu conteúdo em aminoácidos.
Quando proteínas são aquecidas em um meio aquoso, ácido ou básico, as ligações
amida sofrem hidrólise liberando os aminoácidos constituintes, cuja identificação implica no
conhecimento de suas propriedades físico-químicas. É importante relembrar que estas
substâncias são anfotéricas, ou seja, podem agir como ácido de Brönsted (doadoras de
prótons) ou como base de Brönsted (aceptoras de prótons) - reações 1 e 2.
H3N+CH(R)COOH <===> H3N+CH(R)COOI
II
H3N+CH(R)COO- <===> H2NCH(R)COOII
III
(Reação 1)
(Reação 2)
A forma zwiteriônica (II) é produzida pela dissociação do próton em I (Reação 1) e essa
forma pode também sofrer dissociação de um próton e gerar o ânion III.
Cada reação iônica é definida por uma constante de ionização, Ka e um pKa (-log Ka).
Todos os grupos -carboxílicos dos 20 aminoácidos têm valores de pKa próximos, mas
diferentes (pKa 2 a 3), o mesmo se observa para os grupos -amino (pKa 9 a 10). O valor
do pKa de um aminoácido representa o pH no qual as duas formas iônicas estão presentes em
concentrações iguais. Alguns aminoácidos apresentam grupos ionizáveis na cadeia R lateral.
Os grupos ácidos ou básicos extras, é lógico, aumentam a complexidade das reações ácidobase dos aminoácidos, o que garante maior resolução na análise destes compostos em uma
mistura.
O primeiro passo na caracterização de uma proteína isolada e purificada é a
determinação da composição de seus aminoácidos. Ela é hidrolisada e a mistura de
aminoácidos livres é submetida à análise qualitativa e quantitativa. A hidrólise de uma
proteína em meio ácido ou básico não leva a resultados ideais, uma vez que ambos os
métodos tendem a destruir alguns aminoácidos. A hidrólise ácida destrói triptofano, causa
alguma perda de serina e treonina e converte asparagina e glutamina em ácido aspártico e
19
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
glutâmico, respectivamente. A hidrólise em meio básico leva à destruição de serina, treonina e
racemização de aminoácidos livres. A hidrólise ácida é mais utilizada, por ser um método que
apresenta menor índice de destruição.
O processo de hidrólise consiste em dissolver a amostra de proteína em uma solução
aquosa ácida (normalmente HCl 6N) e aquecer a solução a 110°C por 12 a 15 h, ou mantê-la à
temperatura ambiente por uma semana. O intervalo de tempo necessário para a hidrólise
depende da natureza dos resíduos de aminoácidos. Para separação e identificação dos
aminoácidos, as técnicas mais versáteis, econômicas e convenientes são baseadas em métodos
cromatográficos. A análise qualitativa dos aminoácidos pode ser feita por cromatografia em
papel, em camada fina de sílica gel ou celulose, no entanto muitas técnicas mais sensíveis são
utilizadas atualmente. Os aminoácidos livres são detectados na placa ou papel, em que foi
desenvolvido o cromatograma, através da reação com ninhidrina. O aminoácido prolina
desenvolve uma cor amarela, enquanto os demais aminoácidos desenvolvem uma cor violeta.
FUNDAMENTO DA PRÁTICA
Em nosso experimento iremos acompanhar o processo de hidrólise protéica, medindo
os níveis de aminoácidos liberados através da reação com hidróxido de sódio. As proteínas
serão precipitadas com ácido tricloroacético (TCA) enquanto os aminoácidos livres, no
sobrenadante, reagem com NaOH, formando um composto de coloração castanha, que
absorve em 440 nm (Leigton et al. Journal Molecular Biology, 76:103-122, 1973).
PROCESSO
A- Preparação da solução de ovoalbumina (solução A):
Dissolver 3 ml de clara de ovo com 17 ml de água destilada, em um Erlenmeyer e
adicionar 5,0 ml de HCl concentrado. Misturar.
B- Hidrólise ácida com aquecimento:
1- Transferir alíquotas de 3 ml da solução de ovoalbumina para cinco tubos de
ensaio. Levar os tubos ao banho-maria fervente e após 20, 30, 40 e 50 min, transferir o
conteúdo dos tubos de ensaio (um a cada período de tempo) para tubos de centrífuga,
previamente identificados (20, 30, 40 e 50), contendo 0,5 ml de TCA a 30%. Agitar e deixar
em repouso por um mínimo de10 minutos.
2- Centrifugar o conteúdo de cada tubo, transferir 1,5 ml do sobrenadante para tubos
de ensaio, previamente identificados, e ao tubo branco adicionar 1,5 ml de água destilada.
3- No final adicionar ao conteúdo de cada tubo de ensaio, 3 ml da solução de
hidróxido de sódio 1 M.
4- Ler a densidade óptica (DO) em 440 nm, nos diversos tubos. Ajustando o zero do
espectrofotômetro com o branco.
5- Construir um gráfico da DO em função do tempo.
20
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
DOSAGEM DE URÉIA
OBJETIVOS
1- Identificar a uréia como o principal produto de excreção de amônia.
2- Reconhecer a enzima urease como um reagente para análises bioquímicas.
3- Descrever os métodos mais comumente utilizados para a determinação de uréia.
4- Descrever o fundamento do método utilizado neste experimento.
5- Determinar os níveis de uréia no soro sangüíneo.
6- Identificar as causas que geram o aumento nos níveis plasmáticos de uréia.
INTRODUÇÃO
A uréia é formada no fígado a partir de amônia produzida em todos os tecidos, como
produto final do catabolismo protéico. A uréia representa a maior fração de substâncias
orgânicas presentes na urina e cerca de 80% do nitrogênio não protéico excretado na urina,
em condições normais.
Existem vários métodos utilizados na determinação de uréia. Entre eles destacamos:
a) A utilização de xantidrol em métodos gravimétricos, colorimétricos, oxidimétricos.
b) A determinação de nitrogênio pelo método de Kjeldahl.
c) Determinação colorimétrica utilizando reagentes como diacetil-monoxima, dimetilglioxima ou p-dimetil-amino-benzaldeído.
d) Determinação enzimática utilizando urease (uréia amidohidrolase, EC 3.5.1.5). A
hidrólise enzimática da uréia gera amônia (NH3) e bióxido de carbono (CO2). Este pode ser
dosado por gasometria, enquanto a amônia pode ser quantificada: i) por titulometria, com o
reagente de Nessler; ii) enzimaticamente, com glutamato desidrogenase; iii) através da reação
de Berthelot.
FUNDAMENTO DA PRÁTICA
Utilizaremos a reação de Berthelot para quantificar a amônia formada por ação da
urease sobre a uréia, presente no soro sangüíneo. Cada molécula de amônia reage com duas
moléculas do derivado fenólico (salicilato) na presença de hipoclorito de sódio, em meio
alcalino, para formar indofenol, composto de cor azul. A intensidade de cor da reação é
proporcional à concentração de amônia.
PROCESSO
Em um tubo de ensaio diluir 0,1 mL de soro com 0,9 mL de solução salina.
Marcar cinco tubos de ensaio: Branco, Padrão(2) e Teste(2) e adicionar a cada um:
TUBOS
Branco
Padrão (2)
Teste (2)
URÉIA (70mg%)
SORO
0,05 mL
-
0,05 mL
ENZIMA (UREASE)
(268 U/ml)
1,0 mL
1,0 mL
1,0 mL
1- Incubar todos os tubos em banho-maria (BM) a 37°C por 5 minutos (esta é a reação
enzimática).
2- Adicionar a cada tubo 1,0 mL da solução oxidante (hidróxido de sódio 2,8 mM e
hipoclorito de sódio 121 mM), misturar e levar ao BM a 37°C por 5 minutos..
21
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
3- Ler as absorções ópticas em 600 nm, utilizando o branco para ajustar o zero do
espectrofotômetro.
CÁLCULO
Podemos calcular a concentração de uréia no soro - expressa em mg% - através da
fórmula:
Concentração de uréia (mg%) = LT / LP x 70
LT - Leitura do Teste
LP - Leitura do Padrão
CONSIDERAÇÕES SOBRE O MÉTODO
1- O método enzimático para dosagem de uréia apresenta um alto índice de confiança,
uma vez que a urease reage especificamente com a uréia.
2- A reação de Berthelot é um método de dosagem de amônia sensível e acurado.
3- A amostra de soro ou plasma deve ser utilizada tão logo tenha sido coletada. Para
obtenção do plasma não devem ser utilizados anticoagulantes contendo sais de amônia, por
motivos óbvios e fluoretos, porque inibem a urease.
IMPORTÂNCIA CLÍNICA
Os níveis plasmáticos de uréia são utilizados em conjunto com os níveis de creatinina
como auxiliar no diagnóstico diferencial de hiperuremia pré-renal e pós-renal.
Entre as causas pré-renais responsáveis pelo aumento de uréia no plasma estão a
descompensação cardíaca, desidratação, aumento no catabolismo de proteínas. Entre as causas
renais destacam-se nefrite crônica, necrose tubular, glomerulonefrite aguda ou crônica. As
causas pós-renais são qualquer tipo de obstrução do trato urinário (cálculos, dilatação da
glândula prostática, tumores).
Os níveis de uréia se encontram reduzidos na insuficiência hepática aguda, com lesões
muito extensas. No último trimestre da gravidez também se observa uma diminuição nos
níveis de uréia.
Valores normais: 10 a 50 mg de uréia por 100 mL de plasma.
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
22
ESTUDOS DIRIGIDOS
Cinética de Reações Catalisadas por Enzimas
01- Qual a importância das enzimas nos organismos vivos?
02- Como é possível avaliar a concentração de uma enzima em tecido ou fluido biológico?
03- Explique porque a medida da atividade enzimática não corresponde necessariamente à
concentração real da enzima.
04- Qual a importância dos parâmetros Vmáx e KM, na caracterização de uma enzima?
05- Dê exemplos da aplicação clínica das enzimas.
06- Explique como foi possível acompanhar a velocidade da reação enzimática, na prática.
07- Com os dados obtidos na prática sobre enzimas, demonstre como cresce a velocidade de
uma reação enzimática com o aumento da concentração do substrato e calcule KM e Vmáx
da enzima.
Inibição de Reações Catalisadas por Enzimas
01- Qual a importância de analisarmos o efeito de inibidores sobre reações enzimáticas?
02- O que distingue o efeito de um inibidor competitivo daquele não competitivo?
03- Como é possível distinguir, experimentalmente, um inibidor competitivo de um não
competitivo?
04- Com os dados obtidos na prática, demonstre o tipo de inibição em que se enquadra o
cloreto de mercúrio, em relação à urease.
Consumo De Glicose por células de Sacharomyces cerevisiae
05- Explique porque a via glicolítica é importante para as hemácias.
02- Porque é importante separar o plasma dos elementos figurados do sangue, o mais rápido
possível, quando se pretende dosar os níveis plasmáticos de glicose?
03- Qual a opção, quando não há condições de separar o plasma dos elementos figurados de
imediato?
06- Explique como foi possível acompanhar o consumo de glicose pelas células de
Sacharomyces cerevisiae.
07- Com os resultados obtidos na prática calcule a concentração de glicose, em mg%,
construa um gráfico das concentrações em função do tempo de incubação e explique
porque os resultados foram diferentes.
Dosagem de Peróxidos de Lipídeos
01- O que são radicais livres e qual a ação dessas espécies químicas no organismo vivo?
02- Fale sobre três fontes geradoras de radicais livres no organismo vivo.
23
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
03- Como explicar a ação danosa da luz ultravioleta sobre a pele?
04- Explique como foi possível determinar os níveis de peróxidos de lipídeos no plasma.
05- Com os resultados obtidos na prática calcule a concentração de peróxidos de lipídeos
plasmáticos, referidos como níveis de TBARS em M e construa um gráfico da
concentração em função do tempo de incubação das células de S. cerevisiae.
Efeito da Glicose Sobre a Peroxidação de Lipídeos
01- Como é gerado, no organismo vivo, o malondialdeído (MDA)?
02- Por que um aumento de MDA no plasma pode comprometer a atividade biológica das
proteínas?
03- Qual o efeito prático na determinação do efeito da glicose sobre a peroxidação de
lipídeos?
04- Com os resultados obtidos nesta prática e aqueles obtidos na prática anterior demonstre,
graficamente, o efeito da glicose sobre os níveis de TBARS medidos nas células.
Hidrólise de Proteínas
01- Porque é importante conhecermos a seqüência dos aminoácidos constituintes de uma
proteína?
02- O que acontece com as proteínas quando são aquecidas em um meio aquoso, ácido ou
básico?
03- Conceitue pKa e fale sobre a importância desta constante na análise dos aminoácidos em
uma mistura.
04- Explique como foi possível acompanhar o processo de hidrólise protéica.
05- Com os resultados obtidos na prática demonstre, através de um gráfico, o aumento na
concentração dos aminoácidos livres com o tempo.
Dosagem de Uréia no Soro Sangüíneo
01- Fale sobre a origem da uréia no organismo e sobre sua importância.
02- Relacione três métodos utilizados na dosagem de uréia, destacando o mais preciso entre
eles.
03- Qual o fundamento da reação de Berthelot e qual a sua aplicação?
04- Com os dados obtidos na prática calcule a concentração de uréia e comente o resultado.
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
05- Fale sobre a importância clínica da dosagem plasmática de uréia.
24
PAINEL DIRIGIDO
INTEGRAÇÃO E REGULAÇÃO METABÓLICA
Metabolismo é o conjunto total das transformações das moléculas dos nutrientes
orgânicos nas células vivas. Através dessas transformações, catalisadas por enzimas, é
extraída a energia química das moléculas dos nutrientes, que é utilizada na realização do
trabalho celular.
Os organismos vivos podem ser divididos em dois grandes grupos: Os seres
autotróficos (que se alimentam por si mesmos) podem utilizar o dióxido de carbono da
atmosfera como única fonte de carbono e produzir todas as biomoléculas essenciais à vida. Os
seres heterótrofos (que se alimentam às custas de outros) necessitam obter os átomos de
carbono do meio ambiente na forma de moléculas orgânicas relativamente complexas.
Além das fontes de carbono, oxigênio e energia, todos os organismos vivos necessitam
de uma fonte de nitrogênio, necessário para a síntese dos compostos nitrogenados, como os
aminoácidos, as bases púricas e pirimídicas, etc.
O metabolismo pode ser entendido, ainda, como o conjunto das diversas vias
metabólicas, resultado da ação de seqüências multienzimáticas que, individualmente,
catalisam os passos sucessivos dessas vias. As seqüências específicas de intermediários
envolvidos nas vias do metabolismo celular é designado freqüentemente como metabolismo
intermediário. A transformação dos nutrientes, como carboidratos, proteínas e lipídeos; em
estruturas mais simples, como CO2 e H2O é chamada de catabolismo. Este é um processo de
degradação dos nutrientes, acompanhado pela liberação da energia livre inerente à estrutura
complexa das moléculas orgânicas. As vias catabólicas geram energia e convergem para
poucos produtos finais.
O processo de síntese de macromoléculas, como proteínas e ácidos nucleicos, a partir
de pequenas moléculas precursoras (as unidades fundamentais) é conhecido como
anabolismo ou biossíntese. As vias anabólicas envolvem consumo de energia e divergem
para a formação de muitos produtos diferentes.
Figura 1 – Produção de energia no catabolismo e sua
utilização no anabolismo celular
25
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
A Figura 1 mostra que há, na célula, um ciclo energético em que o ATP serve como
elo de transporte de energia química, favorecendo uma união entre o catabolismo e o
anabolismo. Um outro elo importante entre estes dois processos metabólicos é o NADPH, que
transporta energia na forma de força redutora. Com isso é possível entender que a estratégia
do metabolismo é formar ATP, poder redutor e unidades fundamentais para os processos de
biossíntese.
No metabolismo celular existe um intrincado sistema de regulação, de tal forma que a
liberação de energia nos processos catabólicos é controlada pelas necessidades celulares de
energia na forma de ATP e de NADPH, e não pela simples disponibilidade dos nutrientes.
A regulação do metabolismo ocorre de várias formas:
1 – Compartimentação – A célula está dividida em compartimentos, separados uns dos
outros por membranas. Alguns processos metabólicos como a glicólise, a via pentose fosfato,
a síntese dos ácidos graxos ocorrem no citossol. Outros processos ocorrem na mitocôndria,
como acontece com oxidação dos ácidos graxos, as reações do ciclo do ácido cítrico e a
fosforilação oxidativa. Existem ainda aqueles processos que dependem da interação de
reações que ocorrem em ambos os compartimentos, como a gliconeogênese, a síntese de
uréia. Com isso a utilização de um determinado nutriente pela célula vai depender do
compartimento em que essa molécula se encontra. A separação entre as vias de síntese e as
vias de degradação é de fundamental importância para a regulação do metabolismo.
2 – Disponibilidade de substrato – Com freqüência, a concentração do substrato
comanda direta ou indiretamente a velocidade de determinados processos. Embora muitas
vezes seja ignorada a importância do suprimento de substrato como um fator de regulação do
metabolismo, a concentração de ácidos graxos do sangue que entra no fígado é um fator
determinante da velocidade da cetogênese. Gordura é sintetizada em excesso quando ocorre o
consumo excessivo de substratos que contribuem para a lipogênese. Por outro lado, a ingestão
deficiente de substratos glicogênicos (principalmente alanina) podem gerar hipoglicemia na
gravidez ou no jejum prolongado.
3 – Efetores alostéricos – A primeira reação essencialmente irreversível em uma via
metabólica é quase sempre fortemente regulada. As enzimas que catalisam estas reações são
reguladas alostericamente. A quantidade e a atividade dessas enzimas vão determinar o fluxo
de moléculas na maioria das vias metabólicas. Na via glicolítica, frutose 2,6-bisfostato
estimula a fosfofrutocinase e inibe a frutose 1,6-bisfosfatase, dessa forma estimula a glicólise
ao mesmo tempo em que inibe a gliconeogênese. Níveis elevados de ADP estimulam a
atividade da isocitrato desidrogenase, no ciclo do ácido cítrico. O citrato ativa a acetil CoA
carboxilase, estimulando a síntese de malonil CoA, na via de síntese de ácidos graxos. O
aumento nos níveis de malonil CoA inibe a carnitina aciltransferase I, interrompendo a
oxidação de ácidos graxos.
4 – Modificação covalente – Algumas enzimas quando são fosforiladas mudam sua
conformação e sua atividade. A glicogênio fosforilase, por exemplo, quando fosforilada tem a
sua atividade catalítica aumentada, enquanto a glicogênio sintase fosforilada tem a sua
atividade diminuída. Essas modificações covalentes são catalisadas por enzimas específicas.
Entre as enzimas que sofrem regulação pela fosforilação reversível, podemos citar ainda:
piruvato cinase, lipase sensível a hormônio, fenilalanina hidroxilase, frutose 2,6-bisfosfatase.
5 – Níveis de enzimas – A mudança nos níveis enzimáticos é um mecanismo de
regulação que envolve mudanças da velocidade de síntese ou de degradação de enzimas
chaves. Entre essas enzimas está a glicocinase, fosfrutocinase, piruvato cinase, glicose 6fosfatase, frutose 1,6-bisfosfatase.
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
26
6 – Especialização metabólica dos órgãos – A existência de órgãos com diferentes
características, afeta consideravelmente a regulação do metabolismo. Nem todas as principais
vias metabólicas operam em todos os tecidos em um determinado momento.
Estas formas de regulação do metabolismo evidenciam o papel preponderante das
enzimas. Mas vale ressaltar a relevância dos hormônios, como moduladores da velocidade de
uma grande variedade de reações bioquímicas e vias metabólicas.
A Figura 2 apresenta um esquema relacionando às principais vias metabólicas e o local
onde elas ocorrem na célula. Os fatores que dirigem o fluxo das moléculas no metabolismo
podem ser melhor compreendidos pelo exame de três importantes junções metabólicas:
glicose-6-fosfato, piruvato e acetil CoA. Cada uma dessas moléculas têm diversos destinos
mas estão interrelacionadas:
A Glicose-6-fosfato pode ser armazenada como glicogênio, degradada a
piruvato, gerando ATP, ou transformada em ribose 5-fosfato, para síntese de nucleotídeos,
com a produção concomitante de NADPH. Por outro lado, glicose 6-fosfato pode ser formada
pela mobilização do glicogênio ou pode ser sintetizada a partir de piruvato e aminoácidos
glicogênicos pela via da gliconeogênese.
Figura 2 – Uma visão esquemática da compartimentação das
principais vias metabólicas
O Piruvato é primariamente derivado da glicose 6-fosfato, alanina e lactato. A
fácil redução do piruvato, catalisada pela lactato desidrogenase, serve para regenerar NAD+,
permitindo que a glicólise prossiga temporariamente em condições anaeróbicas. O lactato
acumulado nos músculos é transferido para o fígado e sofre oxidação. Uma outra reação
reversível no citossol é a reação de transaminação do piruvato gerando alanina, o que garante
um elo de ligação entre o metabolismo de aminoácidos e de carboidratos. Além da alanina,
outros aminoácidos podem ser convertidos em piruvato. Oxaloacetato, produto da
carboxilação do piruvato, pode ser transformado em fosfoenolpiruvato e, por isso, permite que
a glicose seja sintetizada a partir de piruvato.
Acetil CoA é produzido em maior quantidade pela descarboxilação oxidativa do
piruvato e pela -oxidação dos ácidos graxos. Esse composto pode ser totalmente oxidado a
CO2 e H2O, através das reações do ciclo do ácido cítrico e cadeia transportadora de elétrons,
27
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
ou pode gerar 3-hidroxi-3-metil glutaril CoA, precursor para a via de síntese do colesterol
como também dos corpos cetônicos. Um outro destino do acetil CoA é a sua utilização para a
síntese de ácidos graxos. O acetil CoA serve como elo de integração entre o metabolismo dos
carboidratos, dos lipídeos e das proteínas e a integração destas vias metabólicas com o ciclo
do ácido cítrico.
Integração Metabólica entre os Diversos Tecidos
A integração das diversas funções dos órgãos é um processo muito complexo, uma vez
que os padrões metabólicos de cérebro, músculo, tecido adiposo, trato digestivo e fígado são
muito diferentes. Para um perfeito entendimento destas inter-relações há necessidade, além da
abordagem bioquímica, de uma abordagem fisiológica. Neste texto abordaremos somente
alguns aspectos bioquímicos significativos.
Tabela 1 – Reserva energética em um homem adulto (~70kg)
Energia disponível (kcal)
ÓRGÃO
Glicogênio
Triacilgliceróis
Proteínas
ou glicose
Mobilizáveis*
Sangue
60
45
0
Fígado
400
450
400
Cérebro
8
0
0
Músculo
1.200
450
24.000
Tecido adiposo
80
135.000
40
*Embora proteína não constitua material de reserva, na inanição podem ser consumidos 50%
do seu total.
A distribuição das reservas energéticas nos diversos tecidos (Tabela 1) mostra como
esses tecidos diferem no uso dos alimentos para satisfazer as suas necessidades energéticas:
Cérebro – Não tem reservas energéticas e, por isso, necessita de um
suprimento contínuo de glicose. Ele consome cerca de 120 g diariamente, o que representa
cerca de 60% do consumo de glicose pelo organismo inteiro em repouso. Durante a inanição,
o cérebro utiliza os corpos cetônicos (acetoacetato e -hidroxibutirato), gerados pelo fígado,
como fonte de energia, substituindo parcialmente a glicose. Essa mudança no uso de
combustível de glicose para corpos cetônicos tem por objetivo minimizar a degradação de
proteínas durante a inanição. Os ácidos graxos não servem como alimento para o cérebro, por
não atravessarem a barreira hematoencefálica, mas os corpos cetônicos, derivados dos ácidos
graxos, podem ser utilizados como fonte de energia pelo cérebro.
Músculo – Difere do cérebro por ter grande reserva de glicogênio e além de
glicose utiliza ácidos graxos e corpos cetônicos como fontes de energia. O músculo, como o
cérebro, não tem a enzima glicose-6-fosfatase, por isso não pode exportar glicose, que é seu
alimento preferido para surtos de atividade. A velocidade da glicólise no músculo esquelético
em contração ativa, excede aquela do ciclo do ácido cítrico. Muito do piruvato formado nesse
processo é reduzido a lactato, que flui para o fígado onde é transformado em glicose. A
alanina, como o lactato, podem ser transformada em glicose pelo fígado. No músculo em
repouso os ácidos graxos são a principal fonte de energia, enquanto os corpos cetônicos
podem também servir de alimento para o músculo cardíaco.
Tecido adiposo – Tem como função armazenar triacilgliceróis – produto da
esterificação dos ácido graxos, sintetizados no fígado, com glicerol. O glicerol 3-fosfato,
intermediário nessa reação de esterificação, é um produto da degradação da glicose na via
glicolítica. Como os adipócitos não podem fosforilar o glicerol, necessitam de glicose para a
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
28
síntese de triacilgliceróis. As lipases do tecido adiposo são enzimas envolvidas com a
liberação da maior parte de energia armazenada. A lipase que remove o primeiro ácido graxo
de um triacilglicerol é sensível a vários hormônios circulantes (Figura 3). O controle da
hidrólise de triacilgliceróis deve ser balanceado com o processo de síntese destes compostos,
para assegurar um armazenamento de energia adequado e evitar a obesidade. Os níveis de
glicerol 3-fostato, produzidos a partir de glicose, nos adipócitos desempenham um papel
importante sobre destino dos ácido graxos. Níveis elevados de glicerol 3-fostato estimulam a
esterificação, enquanto baixos níveis deste composto nos adipócitos favorecem a liberação
dos ácidos graxos para a corrente sangüínea.
Fígado - Suas atividades metabólicas são essenciais para o provimento de
material energético para o cérebro, músculo e outros órgãos periféricos. A maioria dos
compostos absorvidos pelos intestinos passa através do fígado, o que permite que ele regule o
nível de muitos destes compostos no sangue. O fígado pode captar grandes quantidades de
glicose e convertê-las em glicogênio, como pode liberar glicose para o sangue a partir das
reservas de glicogênio ou através da gliconeogênese. Lactato e alanina, a partir dos músculos
e glicerol, do tecido adiposo, bem como os aminoácidos glicogênicos, são os principais
precursores de glicose na gliconeogênese. Assim como o fígado regula o metabolismo dos
carboidratos, também desempenha papel central na regulação do metabolismo dos lipídeos.
As lipoproteínas de muito baixa densidade (VLDL) são sintetizadas no fígado e transportam
os ácidos graxos para o tecido adiposo para a síntese de triacilgliceróis. E é o fígado que, no
jejum, transforma os ácidos graxos em corpos cetônicos. Malonil-CoA, precursor para a
síntese de ácidos graxos, inibe a enzima carnitina aciltransferase I, envolvida com o transporte
de ácidos graxos para a mitocôndria. Dessa forma, quando os níveis de malonil CoA estão
altos a síntese de ácidos graxos é favorecida e a -oxidação, inibida. Quando os alimentos são
escassos, os níveis de malonil CoA baixam e os ácidos graxos liberados do tecido adiposo são
transformados em corpos cetônicos.
Figura 3 – Mobilização de ácidos graxos dos adipócitos
induzida por hormônio. PKA
– Proteína
cinase, MAG, DAG, TAG – Mono, di e
triacilglicerol
29
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
Entre os hormônios envolvidos com a integração metabólica, insulina, glucagon
epinefrina e norepinefrina são de particular importância no armazenamento e na mobilização
de alimentos:
Insulina – É secretada pelas células do pâncreas estimulada pela presença de
glicose e pelo sistema nervoso parassimpático. Esse hormônio atua nas cascatas de proteínas
cinase. Estimula a síntese de glicogênio no músculo e no fígado. Estimula a glicólise hepática,
favorecendo a síntese de ácidos graxos. Como já foi discutido, o aumento de ácidos graxos e
glicose no tecido adiposo estimula a síntese e armazenamento de triacilglicerol. A insulina
também promove a captação de aminoácidos ramificados (valina, leucina e isoleucina) pelo
músculo e apresenta um efeito estimulador geral sobre a síntese proteica e inibe a degradação
intracelular de proteínas.
Glucagon – É secretado pelas células do pâncreas em resposta aos baixos
níveis de glicose no sangue. O glucagon inibe a síntese de glicogênio e estimula a
glicogenólise, porque dispara a cascata de cAMP. Além disso estimula a gliconeogênese e
inibe a glicólise por reduzir os níveis de frutose 2,6-bisfosfato. O aumento de cAMP nos
adipócitos ativa a lipase que mobiliza os triacilgliceróis.
Figura 4 – Integração do metabolismo entre os principais tecidos no
estado bem alimentado
Epinefrina e norepinefrina – São secretadas pela medula adrenal e terminações
nervosas simpáticas em resposta ao baixo nível de glicose circulante. Têm efeito semelhante
àquele do glucagon por disparar a cascata de cAMP. Esses hormônios diferem do glucagon
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
30
porque seu efeito glicogenolítico é mais acentuado no músculo do que no fígado. Na presença
desses hormônios a captação de glicose pelos músculos é inibida e os ácidos graxos, liberados
do tecido adiposo, são utilizados como fonte de energia. A epinefrina também estimula a
secreção de glucagon e inibe a secreção de insulina. Assim, as catecolaminas aumentam a
liberação de glicose hepática no sangue e diminuem a utilização de glicose pelo músculo.
A Figura 4 mostra um panorama da integração metabólica entre diversos tecidos. O
transporte dos diversos compostos entre os órgãos é geralmente assumida pelo sangue. Os
lipídeos, a princípio constituintes dos quilomícrons, são removidos do intestino pelo sistema
linfático, enquanto glicose passa das células epteliais intestinais para o fígado através da veia
porta. Ainda no intestino ocorre o metabolismo parcial dos aminoácidos, antes de serem
liberados no sistema porta.
Adaptações Metabólicas no Jejum Prolongado
A energia necessária para um período de 24 horas varia de aproximadamente 1.600
kcal no estado basal a 6.000 kcal, dependendo do grau de atividade. Considerando os dados
da Tabela 1, as reservas energéticas em um indivíduo com peso em torno de 70 kg são
suficientes para satisfazer as necessidades calóricas na inanição por cerca de três meses. Mas
as reservas de glicose se esgotam em apenas um dia. Mesmo assim, o nível de glicose
circulante é mantido acima de 40 mg/dl. O cérebro não suporta níveis mais baixos, mesmo por
períodos curtos. Além disso existem tecidos que só utilizam glicose como fonte de energia
(Tabela 2). No entanto, os precursores de glicose não se encontram em grande quantidade. A
maior parte da energia está armazenada nos ácidos graxos dos triacilgliceróis, mas esses
compostos não podem ser convertidos em glicose, porque o acetil CoA não pode ser
convertido em piruvato. O glicerol, que pode ser utilizado como precursor da glicose, é
liberado em pequenas quantidades dos triacilgliceróis. Resta uma única fonte de precursores
da glicose, os aminoácidos derivados da degradação proteica. Mas como o músculo é a maior
fonte de proteínas e tem papel fundamental na estrutura do organismo, precisa ser
preservado. Como é preciso prover os níveis de glicose acima de 40 mg/dl e preservar as
proteínas do músculo, os ácido graxos e corpos cetônicos passam a serem utilizados como
combustível.
Tabela 2 – Combustíveis Usados pelos Diversos Tecidos
TECIDOS
GLICOSE
A. GRAXOS
C. CETÔNICOS
Eritrócitos
+
Leucócitos
+
Medula renal
+
Córtex renal
+
+
+
Cérebro
+
+
Músculo esquelético
+(exercício intenso)
+(repouso)
+
Músculo cardíaco
+
+
+
Retina
+
Fígado
+
+
Mucosa intestinal
+
As alterações metabólicas durante o primeiro dia de jejum são como aquelas após o
jejum de uma noite. Com a redução nos níveis de carboidratos, aumenta a secreção de
glucagon, estimulando a mobilização de triacilgliceróis do tecido adiposo e a gliconeogênese
pelo fígado. A captação de glicose pelo músculo é reduzida com os baixos níveis de insulina
circulante, enquanto os ácidos graxos entram livremente e a -oxidação destes compostos no
31
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
músculo geram aumento na concentração de acetil CoA e de citrato, inibindo a via glicolítica
e impedindo a conversão de piruvato em acetil CoA.. Após várias semanas de inanição, os
corpos cetônicos tornam-se o principal combustível para o cérebro, reduzindo a utilização de
glicose e garantindo a preservação das proteínas do músculo (Tabela 3).
Tabela 3 – Regulação do Metabolismo no Jejum Prolongado
QUANTIDADE GERADA OU CONSUMIDA EM 24
FONTES DE ENERGIA
HORAS (GRAMAS)
o
3 DIA
40o DIA
Consumo pelo cérebro
Glicose
100
40
Corpos cetônicos
50
100
Utilização de glicose por
50
40
outros tecidos
Liberação pelo fígado
Glicose
150
80
Corpos cetônicos
150
150
Lipólise no tecido adiposo
180
180
Degradação
da
proteína
75
20
muscular
Adaptações Metabólicas no Exercício
A conversão da energia química em trabalho mecânico pelo músculo implica na
necessidade instantânea do consumo de glicose. No exercício anaeróbico (como corrida de
velocidade ou levantamento de peso) o músculo depende de suas reservas de glicogênio e
creatina fosfato, para a produção de ATP. A creatina fosfato serve como fonte de fosfato de
alta energia para síntese de ATP, até que a glicogenólise e a glicólise sejam estimuladas. A
via glicolítica torna-se a fonte primária de ATP para o músculo, devido a falta de oxigênio.
Durante uma corrida de velocidade (100 m em ~10 s) o nível de ATP do músculo cai de 5,2
mM para 3,7 mM, enquanto o nível de creatina fosfato cai de 9,1 mM para 2,6 mM. Com a
glicólise anaeróbica aumenta o nível sangüíneo de lactato de 1,6 mM para 8,3 mM,
favorecendo o abaixamento do pH sangüíneo de 7,42 para 7,24. Por isso essa marcha não
pode ser mantida em uma corrida de 1000 m em ~132 s, porque as reservas de creatina fosfato
seriam consumidas em poucos segundos e a glicólise seria interrompida por falta de NAD+;
sem falar na quantidade de ácido formado. Parte do ATP consumido será gerado pela
fosforilação oxidativa.
A corrida de uma maratona (42.200 m em ~2 h) implica na utilização de ácidos graxos
para a produção de ATP, uma vez que os depósitos de glicogênio são insuficientes para
prover os 150 mol de ATP necessários para essa competição. No entanto, se a oxidação dos
ácidos graxos fossem a única fonte de ATP a maratona deveria durar 6 h, uma vez que essa
via metabólica é muito mais lenta do que a oxidação do glicogênio. Os melhores corredores
consomem quantidades aproximadamente iguais de glicogênio e ácidos graxos, de tal forma
que a glicose seja poupada para o final da maratona.
A taxa de oxidação dos ácidos graxos de cadeia longa aumenta de cinco a oito vezes,
quando humanos são submetidos a um esforço físico que exige menos de 85% do consumo de
oxigênio. Dois fatores determinam a utilização de ácidos graxos: A duração e a intensidade do
exercício. Dessa forma, exercício intenso por 5 a 10 min, implica na utilização de ATP gerado
a partir da oxidação de carboidratos. Em um esforço com menor intensidade por um período
de 30 min ou mais, gera um aumento na taxa de oxidação de ácidos graxos. Essa taxa é
32
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
máxima, quando o esforço físico requer cerca de 60% do consumo máximo de oxigênio.
Indivíduos bem treinados utilizam um percentual maior de ácidos graxos como fonte de
energia, do que indivíduos sedentários, não treinados.
Transtornos do Metabolismo no Diabetes
No diabetes mellitus há uma superprodução de glicose pelo fígado que é subutilizada
pelos outros órgãos. Em um indivíduo não tratado, o nível sangüíneo de insulina é muito
baixo e o de glucagon é muito alto em relação às necessidades do paciente. A alta relação
glucagon/insulina no diabetes estimula a glicogenólise e gliconeogênese e inibe a glicólise,
contribuindo para o aumento dos níveis de glicose circulante. A glicose é excretada na urina
junto com a água, o que leva o diabético na fase aguda da doença sentir fome e sede.
A utilização reduzida de carboidratos no diabético leva a um estímulo na quebra de
lipídeos e proteínas. A oxidação de ácidos graxos promove a síntese de glicose através do
aumento da concentração de Acetil-CoA, um efetor alostérico positivo da piruvato
carboxilase. O aumento nos níveis de Acetil-CoA estimula a produção de corpos cetônicos,
que podem suplantar a capacidade dos rins de manter o equilíbrio ácido-base e levar o
diabético não tratado a entrar em coma, por causa do abaixamento do pH sangüíneo e da
desidratação.
São conhecidas duas formas do diabetes mellitus: O tipo I, ou insulino-dependente,
que em geral começa antes dos 20 anos de idade. Devido à produção defeituosa de insulina
pelas células do pâncreas, os níveis sangüíneos de insulina não aumentam em resposta aos
níveis elevados de glicose no sangue. No tipo II, ou não insulino-dependente, a insulina não
está ausente. Níveis elevados de insulina podem ser observados nesse tipo de diabetes, mas há
resistência dos tecidos à ação do hormônio. O pâncreas do paciente diabético não insulinodependente não produz insulina suficiente para superar a resistência à insulina, induzida por
sua obesidade. Embora não esteja bem entendido o fenômeno da resistência à insulina, a
maioria dos indivíduos com diabetes do tipo II são obesos.
Metabolismo do Etanol e a Relação NAD+/NADH
Etanol é uma substância hidrossolúvel e de baixo peso molecular. Por isso é
rapidamente absorvido, especialmente quando o estômago se encontra vazio. A concentração
alcoólica no sangue pode atingir, em média, cerca de 0,1% uma hora após a ingestão de um
copo de álcool a 50%, quatro copos de vinho, ou quatro garrafas de cerveja. O etanol chega
ao fígado e é transformado em acetato, evolvendo duas etapas:
CH3CH2OH +
Etanol
NAD+
CH3CHO
Acetaldeído
+
NADH + H+ (1)
CH3CHO
Acetaldeído
NAD+
CH3COOAcetato
+
NADH + H+ (2)
+
A etapa (1) – etapa lenta – é catalisada pela enzima álcool desidrogenase, no citossol
enquanto a etapa (2) ocorre na matriz mitocondrial e é catalisada pela enzima aldeído
desidrogenase. Em ambas as etapas ocorre a produção de NADH. Por esta razão, as vias
metabólicas da gliconeogênese e da -oxidação dos ácidos graxos, que têm enzimas cuja
atividade é sensível à presença de NADH, sofrem inibição durante o metabolismo do etanol.
No jejum, esta situação favorece à hipoglicemia e ao acúmulo de triacilgliceróis no fígado
33
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
(fígado gordo). Pode, também, haver um acúmulo de ácido láctico, gerando acidose
metabólica.
O consumo de álcool por uma pessoa subnutrida ou após exercício extenuante pode
causar hipoglicemia. Os equivalentes de NADH em excesso bloqueiam a conversão de lactato
em glicose e promovem a conversão de alanina em lactato, que acumula na corrente
sangüínea. Com a redução da glicemia ocorre queda no desempenho motor e intelectual. Altas
doses de etanol apresenta um efeito depressor, que pode levar a estupor e anestesia.
O etanol apresenta um alto conteúdo energético, produzindo cerca de 7,1 kcal/g,
quando oxidado. O acetato derivado do metabolismo do etanol pode ser transformado em
acetil-CoA, que é oxidado a CO2 e H2O, através das reações do ciclo do ácido cítrico e da
cadeia transportadora de elétrons. No fígado as mitocôndrias têm uma capacidade limitada
para oxidar o acetato, porque a ativação do acetato a acetil-CoA requer GTP. Com o aumento
nos níveis de NADH, as reações do ciclo sofrem inibição. Isto leva a uma redução nos níveis
de GTP, gerado na transformação de sucnil-CoA a succinato, e oxidação do acetil-CoA
também fica comprometida. Grande parte do acetato é exportado do fígado para o sangue e
pode ser oxidado, na forma de acetil-CoA, em outros tecidos.
Como acetato, acetaldeído pode ser exportado do fígado para a circulação sangüínea.
Este último composto reage, rapidamente, com grupos funcionais importantes, como as
proteínas das células sangüíneas e dos demais tecidos, comprometendo a sua atividade
biológica.
Questões Sobre o Tema Integração e Regulação Metabólica
01- Conceitue metabolismo, e explique o que são seres autotróficos e seres heterótrofos e
como esses seres sobrevivem.
02- Fale sobre a importância das enzimas no metabolismo e conceitue anabolismo e
catabolismo.
03- Qual a importância do ATP e NADPH na regulação do metabolismo celular?
04- Relacione as formas pelas quais ocorre a regulação do metabolismo e explique como elas
funcionam.
05- Relacione as principais vias metabólicas e fale sobre a importância das junções
metabólicas: glicose-6-fosfato, piruvato e acetil CoA.
06- Como é possível explicar a integração entre os diversos órgãos, quando existe diferentes
padrões metabólicos entre cérebro, músculo, tecido adiposo, trato digestivo e fígado? Fale
sobre essas diferenças
07- Fale sobre os hormônios envolvidos com a regulação do armazenamento e da mobilização
dos alimento e sobre seu papel na integração do metabolismo.
08- Explique como o organismo vivo se adapta a uma situação de jejum prolongado.
09- As fontes de energia são as mesmas em qualquer atividade muscular? Explique.
10- Quais os transtornos do metabolismo causados pelo diabetes mellitus e o que entende por
diabéticos insulino-dependentes e não insulino-dependentes?
11- Independente dos efeitos prejudiciais, sociais e econômicos do alcoolismo, explique
porque o etanol não deve ser consumido como fonte de energia, apesar do seu alto teor
calórico.
Manual de Aulas Práticas – Prof. Levy dos Santos Guedes
34
BIBLIOGRAFIA
01. BOYER, RF, Modern Experimental Biochemistry. Addison-Wesley Publishing Company,
Massachusetts, 1986.
02. DEVLIN, TM, Manual de Bioquímica com Correlações Clínicas. 6ª Ed., Editora Edgar
Blücher Ltda. São Paulo, 2007.
03. GARRET & GRISHAN, Biochemistry, 2a Ed. Saunders College Publishing, New York,
1999.
04. LEHNINGER, A; NELSON, DL; COX, MM, Princípios de Bioquímica, 3a Ed. Sarvier,
São Paulo, 2002.
05. LIMA, AO, et al., Métodos de Laboratório Aplicados à Clínica, 5a Ed. Guanabara
Koogan, Rio de Janeiro, 1977.
06. MARZZOCO, A; TORRES, BB, Bioquímica Básica. Editora Guanabara Koogan, Rio de
Janeiro, 1990.
07. MONTGOMERY, R; CONWAY, TW; SPECTOR, AA, Bioquímica uma Abordagem
Dirigida por Casos. Artes Médicas, São Paulo, 1994.
08. PURVES, WK, et al, Vida a Ciência da Biologia, 6ª Ed. Artmed, Porto Alegra, 2002.
09. SMITH, EL, et al., Bioquímica Aspectos Gerais, 7a Ed. Guanabara Koogan, Rio de
Janeiro, 1985.
10. SMITH, EL, et al., Bioquímica Mamíferos, 7a Ed. Guanabara Koogan, Rio de Janeiro,
1985.
11. STRYER, L, Bioquímica, 4a Ed. Guanabara Koogan, Rio de Janeiro, 1996.
12. TIETZ NW, Quimica Clinica Moderna. Interamericana, México,1972.
13. UCKO, DA, Química para as Ciências da Saúde: Uma Introdução à Química Geral,
Orgânica e Biológica. Editora Manole Ltda., São Paulo, 1992.
14. UFPR, SCB, DEPARTAMENTO DE BIOQUÍMICA, Bioquímica: Aulas Práticas.
Scientia et Labor, Editora da Universidade Federal do Paraná, Curitiba, 1987.
15. VOET, D, et al., Fundamentos de Bioquímica, Artmed Editora, 2002.