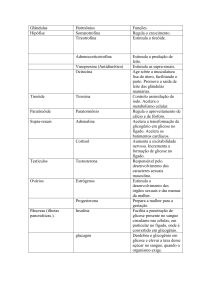
INTRODUÇÃO
A maior parte da glicose catabolizada nos tecidos animais sofre o
processo glicolítico e origina moléculas de piruvato. Por sua vez, a
maior parte do piruvato é oxidada.
Existe uma via secundária que também participa deste processo. A via
das pentoses, também chamada de via do fofogliconato produz dois
produtos especiais nos tecidos dos animais. Esta via é metabólica e
constituída por uma seqüência enzimática com várias etapas,
começando na glicose 6-fosfato, presente no início da glicólise, e
levando a produção do açúcar ribose, que é uma pentose. Este açúcar é
importante, pois é utilizado na síntese da molécula de ácidos nucléicos
indispensáveis na síntese de proteínas.
Ao longo dessa via ocorre também a redução da coenzima NADP para
NADPH. Em etapa posterior, o hidrogênio que reduz o NADP é
cedido e utilizado na síntese intracelular de lipídeos, sendo, portanto, a
via das pentoses também importante para este processo.
A glicogenossíntese é o processo pelo qual a glicose é polimerizada a
glicogênio, que é acumulado nas células em quantidades variáveis de
acordo com o tipo celular, funcionando aí como depósito de energia
acessível à célula. Em determinadas células, como nas do fígado e
músculo, este processo pode ser intenso e ocorrem extensos depósitos
de glicogênio. O glicogênio hepático que chega a 150g é degradado no
intervalo das refeições mantendo constante o nível de glicose no
sangue ao mesmo tempo que fornece este metabólito as outras células
do organismo. O glicogênio muscular, ao contrário, só forma glicose
para a contração muscular.
As vias metabólicas citadas acima, ocorrem de modo geral na maioria
das células, havendo, porém, maior ênfase em determinadas vias, de
acordo com a função de cada tipo celular.
Como já foi dito, a glicose passa por um catabolismo glicólitico, o
mesmo acontece com os aminoácidos que passam por um processo de
oxidação, através de uma pequena fração de energia provida pelo
homem. Depois da remoção dos grupos amino pela transaminação até o
α- cetoglutarato, os esqueletos carbônicos dos aminoácidos sofrem
degradação oxidativa em compostos que entram no ciclo do ácido cítrico
para oxidação a CO2 e H2O. Há cinco vias pelas quais os esqueletos
carbônicos dos aminoácidos entram no ciclo do ácido cítrico: via acetilCoA, via α-cetoglutarato, via succinato, via fumarato, via oxalacetato. Os
aminoácidos que entram na via da acetil-CoA, são divididos em dois
grupos. O primeiro ( alanina, cisteína, glicina, serina e treonina ) produz
acetil-CoA via piruvato, e o segundo ( leucina, lisina, tirosina, triptofano )
produz acetil-CoA via acetoacetil-CoA. Os aminoácidos prolina,
histidina, arginina, glutamina, e valina entram via succinato; quatro
átomos de carbono da fenilalanina e da tirosina entram via fumarato e a
aspargina e o ácido aspártico entram via oxaloacetato. Vários defeitos
genéticos humanos ocorrem nas vias catabólicas dos aminoácidos. A
fenilcetonúria é particularmente séria e um defeito relativamente
comum.
Os animais amonotélicos (peixes ósseos e girinos) excretam o
nitrogênio do grupo amino através de suas guelras como amônia,
obtida pela hidrólise da glutamina. Os animais ureotélicos(a maioria
dos animais terrestres) excretam o nitrogênio do grupo amino como
uréia. A uréia é formada no fígado pelo ciclo da uréia, descoberto por
Hans Krebs. A arginina é o percursor imediato da uréia, pela ação da
arginase, que produz uréia e ornitina. A arginina é ressintetizada a
partir da ornitina pela carbamoilação desta última até a citrulina as
expensas do carbamoil fosfato, seguido pela adição de um grupo
amino a citrulina, as expensas do aspartato. A ornitina é regenerada
em cada volta do ciclo. Os animais uricotélicos(pássaros, serpentes,
lagartos) excretam o nitrogênio do grupo amino numa forma semisólida, como ácido úrico, um derivado das purinas. A formação da
uréia, não-tóxica, e do ácido úrico, sólido, tem um alto gasto de ATP.
As vias biossintetizantes que levam aos aminoácidos são sujeitas à
inibição alostérica pelo produto final; a enzima reguladora geralmente
é a primeira na seqüência. Os aminoácidos são percursores de muitas
outras moléculas importantes.
O anel da porfirina das proteínas hemínicas é derivado da glicina e do
succinil-CoA.
O sistema do anel purínico dos nucleotídeos purínicos é constituído
passo-a-passo a partir do carbono 1 da 5-fosforribosilamina. Os
aminoácidos fornecem todos os átomos de nitrogênio das purinas. O
fechamento dos dois anéis garante a formação do núcleo das purinas.
As pirimidinas são formadas a partir do ácido aspártico, CO2 e amônia.
A ribose 5-fosfato é então ligada produzindo os ribonucleotídeos das
pirimidinas.
As purinas livres são recuperadas e transformadas em nucleotídeos por
uma via separada. A deficiência genética numa das enzimas de
recuperação resulta na doença de Lesc Nyham, caracterizada por
sintomas comportamentais bizarros. A gota, outra doença genética,
resulta do acúmulo de cristais de ácido úrico nas articulações.
Artigo completo: “GAINING ON FAT”, do qual foi extraído uma
parte, que discute o armazenamento de glicogênio e sua utilização pelo
corpo humano, ressaltando a obesidade dependente de fatores
genéticos, mais fatores ambientais, como os hábitos alimentares e o
acesso à comida.
Artigo completo “Gaining on Fat”, Scentifc American Agust 1996, do
qual foi extraído um tópico relativo ao armazenamento do glicogênio,
discute a razão do porque alguns indivíduos são obesos toda a vida,
inferindo uma hipótese que considera os genes determinantes de 80%
ao fato de o indivíduo ganhar peso com facilidade, adicionados aos
fatores do ambiente familiar; ou seja, se desde bebê foi acostumado a
ingerir alimentos calorícos; ou o tipo de comida de refrigeradores,
hábito de atividades esportivas etc...
Quais genes influenciam nossa alimentação, metabolismo, atividades
físicas, e como eles exercem esse poder permanece um mistério. Apesar
dos cientistas terem localizado versões de 5 genes no DNA humano,
dos genes que já foram identificados em animais, correlacionados
diretamente com o aumento de peso, os genes propriamente ditos não
foram
encontrados,
nestas
pesquisas
porém,
os
cientistas
estabeleceram que devem haver múltiplos genes que interagem e
selecionam uma “suscetibilidade individual”, para ganhar peso.
Eventualmente esse genes poderiam ser encontrados, mas o principal é
saber como eles agem no corpo humano.
Seguindo a tradução do tópico do artigo:
A GORDURA NA BALANÇA.
Em uma conferência no ano passado, pesquisadores reviram a
evidência e julgaram que apesar da hipótese sob a suscetibilidade para
ganhar peso não ter sido descartada, existe mais um “mérito biológico”
do que evidências estabelecidas. A mais recente teoria afirma que nós
mantemos o peso, quando a nossa variação do metabolismo de
feedback
mantém-se
sintonizada
com
qualquer
que
seja
a
suscetibilidade genética que carregamos.
Mudanças econômicas e culturais estão desarranjando este equilíbrio e
propelindo mais pessoas; aquelas com maiores fatores de riscos
genéticos para obesidade.
Os primeiros suspeitos são precedentes das gorduras que pingam dos
hambúrguer, as que amaciam os sorvetes e as que fritam carnes. Mas
os bioquímicos pelo menos estão resolvendo o porque destas gorduras
serem ruins. Por alguns anos, eles sabiam que as pessoas que
alimentavam-se com refeições altamente calóricas em gorduras, iriam
consumir cerca da mesma quantidade das que se alimentavam-se com
uma refeição a base de muitos carboidratos. Porque as gorduras têm
mais calorias por mordida? Contudo pessoas obesas tendem a
consumir mais energia do que podem queimar; um fenômeno
conhecido como super consumo passivo. Uma razão para isto, de
acordo com o biopsicólogo John E. Blundell da Universidade de Lerds,
parece ser que o sistema de controle da fome e saciedade responde
rápido para as proteínas e carboidratos, mas vagarosamente para as
gorduras, e vagarosamente demais para parar o alto consumo de
gorduras, antes que o corpo já tenha muito. O sistema metabólico
parece ser a favor de carboidratos (os quais incluem açúcares e amido).
Enfrente um prato de massa e uma soda, e seu organismo irá acelerar a
combustão dos carboidratos; devore uma porção de torresmo e sua
taxa de oxidação de gorduras mal mover-se-á, aponta Jean Pierre Flatt,
bioquímico da Escola de Medicina da Universidade de Massachustehs.
A maioria das gorduras que chega é transportada para diretamente
para o estoque, e queimada depois, somente se a reserva de
carboidratos cair abaixo do limite, o qual varia de pessoa para pessoa.
A uma outra maneira de aumentar a taxa na qual a gordura é
queimada, para obtenção de energia: perder peso. Mais gordura no
corpo, mais ácidos graxos circulando na corrente sangüínea, o que é
um impulso para a oxidação das gorduras, para que seja
eventualmente o “balanço de gordura” seja alcançado, onde toda a
gordura ingerida é consumida, e o peso estabiliza. Muitos fatores
genéticos e biológicos podem influenciar na taxa de oxidação e podem
afetar a sua sintonia com ambiente.
Olestra uma gordura artificial aprovada recentemente este ano, pelo
Foof and Drug Adminstration, pode mudar estas taxas. Olestra tem
valor parecido com uma gordura normal, mas não é digerível quando
flui através do corpo. Um estudo preliminar feito por George A. Bray,
diretor executivo da Penmington’s, sugere que o ingrediente cause um
curto circuito no consumo passivo exagerado. Por duas semanas, Bray
substitui a gordura natural em suas principais refeições por Olestra.
“Não
compensamos
comendo
maior
quantidade”,
diz
acrescentando que: “são necessários estudos mais apurados”.
Bray,
O balanço de gordura explica em parte porque o ajuste varia entre as
pessoas que comem gordura demasiadamente, algumas oxidam
gordura eficientemente para pesar normalmente, outras queimam
muito pouco , até que o excesso force que a taxa de oxidação suba. Mas
o modelo não explica sozinho porque alguns de nós não comem além
do normal. Para responder isto, Flatt propôs a “hipótese do
glicogênio”.
O corpo humano pode estocar glicogênio por um suprimento diário de
carboidratos, na forma de glicogênio ou simplesmente amido. As
reservas de glicogênio funcionam como um tanque de combustível,
nós particularmente completamos o estoque com cada refeição, mas
raramente o lotamos. Na verdade, a diferença entre o “cheio e o vazio”
parece ser um caso de preferência individual, influenciado por fatores
como diversidade e paladar da comida a disposição, pressões sociais e
hábitos alimentares. Pessoas que contentam-se com baixas taxas de
glicogênio ou que freqüentemente reduzem o nível através de
exercícios queimam gordura mais rapidamente, do que aquelas que
gostam de manter seus tanques cheios, sugeriu Flatt.. Mas, ele acredita
que o principal fator que liga a estocagem de glicogênio com o apetite
permanece ainda não aprovada.
Pesquisadores precisam de mais evidências antes que possam provar
esse ajuste de pontos, essa sintonia entre vários fatores como correta.
James O. Hill, da Universidade do Colorado, no centro de pesquisas de
Ciências e Saúde começou coletando alguns dados críticos. Ele esta
reunindo em um registro as mais preciosas fontes na pesquisa de
obesidade: pessoas têm perdido grandes quantidades de peso e
mantendo isto por vários anos sem recaídas. Hill já tem identificados
cerca de mil destes indivíduos e começou a examinar uma grama de
pistas biológicas para o sucesso destes indivíduos.
Infelizmente no decorrer da explanação da regulação de peso, muito é
deixado para o controle voluntário, porque todos os ciclos metabólicos
envolvidos são governados subconcientemente. A teoria da sintonia
de ajustes de pontos até agora não sugeriu mudanças drásticas no
estilo de vida que pudessem produzir no corpo a volta de um novo
peso. Mas sem assistência, mudanças radicais são o suficiente para
fazer uma diferença evidente, desconfortável e impraticável para
milhões de seguidores de dieta que tem tentado estratégias e falhado.
ESTUDO DIRIGIDO I
VIA DAS PENTOSES E GLIGONEOGÊNESE
1-Qual a diferença entre NADH e NADPH? Qual é o papel de cada
uma destas moléculas?
O NADH é oxidado pela cadeia respiratória para gerar ATP, enquanto o
NADPH serve como doador de um hidrogênio e elétrons (serve como cofator)
nas biossínteses redutoras. O que destingue o NADH do NADPH é o grupo
fosforila no carbono 2 (C2) de uma das unidades de ribose do NADPH.
2-Quando o NADPH é formado? Mostre a equação da reação em que o
NADPH é formado.
Na via metabólica de pentose fosfato, composta por uma seqüência enzimática
composta por várias etapas, com início na glicose 6-P (também presente na via
glicolítica), leva a formação da ribose concomitantemente o NADPH é gerado
pela redução da coenzima NADP+ para NADPH, quando a glicose 6-P é
oxidada a ribose 5-P.
Glicose 6P+2 NADP+H2O
Ribose5P+2NADPH+2H+CO2.
3-Na célula onde ocorre a via das pentoses?
A via das pentoses ocorre no citosol, parte solúvel do citoplasma. Em vegetais,
a via das pentoses ocorre a partir do CO2 na fotossíntese.
4-Como a via das pentoses está relacionada com a glicólise? Por quais
enzimas?
A via das pentoses está relacionada com a glicólise pelas seguintes enzimas:
1º Transcetolase:
Xilulose5P+Ribose5P-(transcetolase) Gliceraldeido3P+Sedo-heptulose7P.
2ºTransaldose:
Sedo-heptulose7P+Gliceraldeido3P-(transaldose) Eritrose4P+Frutose6P
3ºReação:
Xilose5P+Eritrose4P-(tanscetolase) Gliceraldeido3P+Frutose6P.
O composto comum as duas vias é formado à partir da Ribose 5P, formada na
via da pentose-fosfato que é transformado em intermediário da glicólise, ou
seja a Frutose 6P.
5-Nas reações de transferência de três carbonos, catalisadas pela
tanscetolase, quais os tipos de açúcar considerados receptores e
doadores?
Receptores: Aldose
Doadores: Cetoses
Aldeídos receptores(aldoses): D Ribose 5P; D Gliceraldeido 3P; D Eritrose 4P.
Cetonas doadoras(cetoses): D Xilulose 5P; D Frutose 6P; D Sedoheptulose 7P.
6-Explique como na via das pentoses, a reação de epimerização é
importante na obtenção do Gliceraldeido 3P.
A reação de epimerização é importante pois forma a Ribulose em seu epímero,
que servirá de substrato à reação com a transcetolase e, ocorrendo
posteriormente a formação de Gliceraldeido 3P através da reação do epímero
com outra pentose.
Ribulose 5P-(fosfopentoseepimerase) Xilulose 5P
Xilulose 5P+Eritrose 4P-(transcetolase) Gliceraldeido 3P+Frutose 6P
7-Quais os produtos formados na primeira etapa da via das pentoses?
A primeira etapa da via das pentoses é a etapa oxidativa. Os produtos
formados são: Gliceraldeido 3P e Sedoheptulose 7P.
8-Na via das pentoses quem determina o fluxo da glicose 6P? Quais as
condições que podem ser estudadas?
É determinado pela necessidades de NADPH e de Ribose 5P. As condições são:
1º É exigido muito mais Ribose 5P do que NADPH;
2º A necessidade de NADPH e de Ribose 5P estão em equilíbrio;
3º É exigido muito mais NADPH do que Ribose 5P.
9-Por que a via das pentoses é muito mais ativa no tecido adiposo do
que no muscular?
Porque a missão da via das pentoses é gerar NADPH para as biossínteses
redutoras. No tecido adiposo são consumidas grandes quantidades de NADPH
na síntese redutora de ácidos graxos à partir da acetil CoA. Experiências com
marcações radioativas mostram que a atividade da via das pentoses é muito
baixo no músculo esquelético e muito alta no tecido adiposo.
10-Explique como a deficiência de glicose 6P desidrogenase causa a
anemia hemolítica induzida por drogas.
A via das pentoses é a única fonte de NADPH em hemácias pois elas não têm
mitocôndrias. A Deficiência de Glicose 6P desidrogenase causa hemólise dos
glóbulos vermelhos, pois diminui a quantidade de glutatião reduzido que é
essencial para a manutenção normal das hemácias e manter a hemoglobina no
estado ferroso.
11-O que significa gliconeogênese? Onde ocorre?
A gliconeogênese inclui todos os mecanismos e vias responsáveis pela
conversão de compostos aglicanos(não carboidratos ou não açúcares) a glicose
e glicogênio, ocorrendo no fígado(principal local) e córtex do rim(menor
quantidade).
12-Quais os pontos de entrada para a gliconeogênese?
Os principais percursores não glicídios são: Lactato; Aminoácidos e glicerol.
O Lactato é formado pelo músculo esquelético ativo, quando a velocidade da
glicólise excede a taxa metabólica do ciclo do ácido cítrico (ciclo de Krebs) e a
cadeia respiratória (fosforilação oxidativa). Entra pelo piruvato.
Os Aminoácidos são derivados de proteínas em dietas e jejum, da degradação
de proteínas no músculo esquelético. Entra pelo oxalacetato e piruvato.
O Glicerol é produzido pelas células adiposas através da hidrólise (degradação)
dos trigliceróis. Entra pelo dihidroxiacetona-P.
13-Quais os percursores da glicose através da gliconeogênese?
Lactato, aminoácidos e glicerol.
14-Que tipos de reações da glicólise devem ser contornadas na
gliconeogênese?
São as reações irreversíveis da via:
Glicose+ATP-(hexoquinase) Glicose 6P+ATP
Frutose 6P-(fosfofrutoquinase) Frutose 1,6 di P
Fosfoenolpiruvato+ATP-(piruvatoquinase) Piruvato+ATP
15-Qual é o tipo de controle utilizado pelo organismo para a atividade
da piruvato carboxilase? Qual é a reação que ela participa?
A atividade da piruvato descarboxilase depende da presença de acetil CoA, que
é uma enzima mitocondrial, enquanto que as demais enzimas da
gliconeogênese são citoplasmáticas. Sendo assim a ativação da enzima pela
acetil CoA é um importante mecanismo de controle fisiológico. A enzima
participa da gliconeogênese, fornecendo o oxalacetato à partir do piruvato e
regula o nível de intermediários do ciclo de Krebs ( ciclo do ácido cítrico).
Piruvato+CO2+ATP+H2O-(piruvato carboxilase) Oxalacetato Acetil
CoA+ATP+P
Oxalacetato+GTP-(piruvato carboxilase) Fosfoenolpiruvato+GDP+
CO2.
16-Qual é o cofator necessário para a transaldose?
Os cofatores necessários para a transaldose são produtos da reação da
transcetolase que tem como coenzima a tiamina di-P. Os produtos são: Cetose
de sete carbonos; aldose: gliceraldeido 3P.
17-Como a dihidroxiacetona ativada é transportada pela transaldose?
A dihidroxiacetona ativada é transportada pela transaldose como uma base de
Schiff, que se forma entre a carbonila da cetose substrato e o grupamento
amina de uma lisina no centro ativo da enzima.
18-Qual a importância da gliconeogênese?
Através da gliconeogênese as necessidades de glicose no organismo são
supridas à partir de fontes não glicídicas, em ocasiões de um período de maior
jejum, ou durante períodos de exercícios intensos, onde as reservas de glicose
disponíveis à partir do glicogênio não são suficientes.
BIBLIOGRAFIA:
- Conn & Stumpf
“Introdução à Bioquímica”
São Paulo, 1980
Edgard Blücher Ltda.
- Junqueira & Carneiro
“Biologia Celular e Molecular”
Rio de Janeiro, 1991
Guanabara Koogan.
- Lenhninger,Albert L.
“Princípios de Bioquímica”
São Paulo, 1989
Sarvier.
- Stryer, Lubert
“Bioquímica”
Rio de Janeiro, 1995
Guanabara koogan.
- Revista: Scientif American, Julh 1996.