UNIVERSIDADE VILA VELHA
PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA DE
ECOSSISTEMAS
Dissertação de Mestrado
RAZÃO SEXUAL DE Podocnemis unifilis NO PARQUE INDÍGENA
DO XINGU, MATO GROSSO, BRASIL.
ALINE SILVA GOMES
VILA VELHA
DEZEMBRO DE 2011
UNIVERSIDADE VILA VELHA
PROGRAMA DE PÓS-GRADUAÇÃO EM ECOLOGIA DE
ECOSSISTEMAS
Dissertação de Mestrado
RAZÃO SEXUAL DE Podocnemis unifilis NO PARQUE INDÍGENA
DO XINGU, MATO GROSSO, BRASIL.
Dissertação apresentada a Universidade
Vila Velha, como pré-requisito do
Programa de Pós-graduação em Ecologia
de Ecossistemas, para a obtenção do
título de Mestre em Ecologia.
ALINE SILVA GOMES
Orientador:
Prof. Dr. Paulo Dias Ferreira Júnior
VILA VELHA
DEZEMBRO DE 2011
Dissertação de Mestrado
RAZÃO SEXUAL DE Podocnemis unifilis NO PARQUE INDÍGENA
DO XINGU, MATO GROSSO, BRASIL.
ALINE SILVA GOMES
Aprovada em 16 de dezembro de 2011,
Banca Examinadora:
________________________________________________________
Dra. Cecília Baptistotte – Projeto TAMAR/ICMBio
________________________________________________________
Prof. Dr. Ary Gomes da Silva – UVV
________________________________________________________
Prof. Dr. Paulo Dias Ferreira Júnior– UVV
Dedico este trabalho à minha mãe, que
nunca se importou em sonhar meus sonhos. Com
todo amor e gratidão que uma filha pode ter na vida.
AGRADECIMENTOS
Agradeço primeiramente ao CNPq, pela concessão da bolsa de mestrado que
possibilitou a realização deste trabalho. Agradeço à Universidade Vila Velha e,
especialmente, ao Programa de Pós Graduação em Ecologia de Ecossistemas pela
estrutura e formação concedidos durante essa jornada.
Agradeço à equipe do Projeto Tarekaja’a Xingu, que possibilitou a realização do
trabalho. Ao RAN/ ICMBio, que sempre nos apoiou quando precisamos. A
EMBRAPA/CENARGEN que possibilitou à execução de todas as etapas do trabalho.
À comunidade indígena Kamayurá-Morená, que além de possibilitar a realização do
trabalho, nos acolheu muito carinhosamente, permitindo que eu conhecesse a magia
da vida em uma aldeia indígena (e não só a magia!).
Agradeço carinhosamente ao meu orientador, Paulo Dias Ferreira Júnior, pela
amizade e dedicação ao longo dos anos de trabalho juntos. Agradeço também pelos
puxões de orelha, quando vieram a calhar, e por todas as oportunidades que tive em
função da confiança depositada, principalmente a oportunidade de conhecer o
“mundo novo” das tartarugas de água doce!
A Alessandro Coutinho Ramos que, muito mais do um coordenador de pósgraduação, foi um grande amigo e me adotou como se fosse de sua equipe de
trabalho e fez da Ecologia uma família para mim. Muito obrigada!
Ao Laboratório de Histopatologia da UVV, em especial ao Carlos Roberto Marcos,
que fez o preparo de todas as lâminas histológicas. Aos Professores do Curso de
Medicina Veterinária Marcelo Renan e Taísse.
A todos os colegas do Laboratório de Ecologia Aquática e Terrestre (LETA) que
sempre acompanharam e apoiaram o trabalho, em especial à Alexandra Frossard,
que desde o início do trabalho ajudou no foi preciso. À Aline Dassoler e Paulo
Roberto de Jesus, que também prestaram grande auxílio. À Jordana Borini Freire,
colega que sempre esteve disposta a fazer o intercâmbio entre campo e laboratório,
ajudando no que fosse necessário. A Adélio Lubiana que, além de transmitir muito
conhecimento nos meus tempos de estagiária, retribuiu a ajuda quando precisei.
Agradeço especialmente à minha família, pelo apoio e compreensão nas minhas
ausências em casa (nem sempre foi fácil!).
A Felipe Tessarolo, que ultrapassou os limites do companheirismo, foi amigo,
colega, auxiliar de laboratório (quando foi necessário), e sempre me impulsionou
para frente, me puxou as orelhas e me manteve seguindo, muitas vezes abrindo
mão dos seus próprios interesses em função dos meus deveres. Agradeço mais
uma vez à minha mãe, que sempre “me cobriu” quando as coisas apertavam e
possibilitou que eu seguisse em frente, mostrando que eu estava no caminho certo.
Agradeço a Deus por ter mantido minha força de vontade e perseverança para
chegar até aqui.
SUMÁRIO
1. Introdução .............................................................................................................3
2. Fundamentação Teórica ...................................................................................... 7
Referências Bibliográficas I ................................................................................ 15
3. Capítulos .............................................................................................................24
3.1. Capítulo 1: Razão sexual de Podocnemis unifilis (Troschel 1848) (Testudines:
Podocnemididae) no Parque Indígena do Xingu, Mato Grosso, Brasil...............23
Referências Bibliográficas II...............................................................................38
3.2. Capítulo 2: Influência do local da desova no desenvolvimento embrionário de
Podocnemis unifilis (Troschel 1848) (Testudines: Podocnemididae) no Parque
Indígena do Xingu, Mato Grosso, Brasil.............................................................41
Referências Bibliográficas III..............................................................................56
4. Considerações Finais .........................................................................................61
1
RESUMO
GOMES. A.S. RAZÃO SEXUAL DE Podocnemis unifilis NO PARQUE INDÍGENA DO
XINGU, MATO GROSSO, BRASIL. Orientador: Paulo Dias Ferreira Júnior.
A determinação sexual dependente da temperatura é um dos aspectos mais
importantes da biologia reprodutiva dos quelônios, fazendo do ambiente de
incubação um fator limitante da reprodução, uma vez que as características do meio
serão responsáveis pela definição da temperatura no interior do ninho e,
conseqüentemente do sexo dos indivíduos. Entender a influência das características
do meio físico no desenvolvimento dos embriões de tartarugas ainda é um desafio,
uma vez que uma ampla gama de variáveis envolvidas dificulta a interpretação dos
resultados obtidos em condições naturais. Visando preencher as lacunas acerca da
temperatura responsável pela geração de machos e de fêmeas de Podocnemis
unifilis foi monitorada a temperatura de 120 ninhos naturais em conjunto com a
realização de incubação sob temperaturas controladas em laboratório. Foi realizada
a sexagem dos filhotes, a fim de definir a temperatura pivotal e estabelecer a razão
sexual dos filhotes gerados nas praias para a população do rio Xingu, na área do
Parque Indígena do Xingu. A partir da sexagem dos filhotes eclodidos no laboratório
foi possível estabelecer a temperatura pivotal, responsável pela produção de
machos e fêmeas em proporções iguais e então foi estimada a razão sexual dos
filhotes eclodidos nos ninhos situados nas praias do rio Xingu, no Parque Indígena
do Xingu (MT). Com base no equivalente de temperatura constante a razão sexual
dos filhotes na temporada de 2010 foi de 47% de fêmeas. O ambiente de desova é
um importante fator para os ninhos sujeitos ao alagamento, mas não interferiu na
temperatura dos ninhos. O manejo adotado pela comunidade indígena e que
envolve a transferência de ninhos sujeitos ao alagamento e à predação não
influenciou o sucesso da eclosão e a temperatura dos ninhos indicando que ele é
adequado.
Palavras chaves: tracajá, determinação sexual, incubação, comunidade indígena,
manejo
2
ABSTRACT
GOMES. A.S. RAZÃO SEXUAL DE Podocnemis unifilis NO PARQUE INDÍGENA DO
XINGU, MATO GROSSO, BRASIL. Orientador: Paulo Dias Ferreira Júnior.
Temperature-dependent sex determination is one of the most important aspects of
turtle reproductive biology. The incubation environment is a limiting factor for
reproduction, because environmental characteristics are responsible for the
temperature in the interior of the nest and consequently for the sex ratio of the
hatchlings. To understand the influence of the characteristics of the physical
environment on embryo development is still a challenge, because the wide range of
variables involved makes it difficult to interpret results obtained in natural conditions.
Aiming to fill the gaps in understanding the role of temperature in the differentiation of
male and female Podocnemis unifilis, the temperature of 120 natural nests was
monitored, concomitantly with incubation of eggs under controlled temperatures in
the laboratory. Based on the sexes of hatchlings incubated in the laboratory, it was
possible to establish the pivotal temperature. Then, the sex ratio of clutches in
natural nests located on the beaches of the Xingu River, in the Xingu Indigenous
Park (MT), was estimated. According to the constant-temperature equivalent, the sex
ratio of hatchlings in the 2010 nesting season was 47% females. Nest placement is
important for hatching success, principally for nests subject to flooding; but did not
affect the nest temperatures. The management protocol adopted by the indigenous
community and which involves the transfer of nests that are subject to flooding and to
predation, did not affect the hatching success or nest temperatures, showing that it is
the most appropriate method.
Keywords: incubation, management, sex determination, yellow spotted turtle, CTE.
3
1. INTRODUÇÃO
O meio exerce papel fundamental no processo reprodutivo das tartarugas. As
características do local de desova irão definir as condições da incubação e a
viabilidade do ninho, garantindo o calor e o abrigo necessários para a eclosão dos
filhotes (FERREIRA JÚNIOR et al. 2008, FERREIRA JÚNIOR 2009 a, b). O gênero
Podocnemis, no Brasil, possui quatro espécies que ocupam as bacias dos rios
Amazonas e Tocantins-Araguaia (RUEDA ALMOCINADE et al. 2007). O gênero,
considerado como um dos melhor estudado do Brasil, conta com a tartaruga-daamazônia (P. expansa), o tracajá (P. unifilis), o iaçá (P. sextuberculata) e a irapuca
(P. erythrocephala) amplamente distribuídos pelos rios amazônicos. Essas espécies
são continuamente exploradas como fonte de alimento, tanto pelos seus ovos,
quanto pela sua carne (PEZZUTI et al. 2005, KEMENES & PEZZUTI 2007, PEZZUTI et al.
2010).
Todas as espécies o gênero Podocnemis testadas apresentam determinação sexual
dependente da temperatura de incubação, o que significa que as características
ambientais afetam a sua reprodução (VALENZUELA & LANCE 2004). Nestas espécies a
incubação é um dos momentos mais críticos do ciclo de vida. Apesar de figurarem
entre as espécies brasileiras mais estudadas, ainda existem grandes lacunas no que
diz respeito à temperatura pivotal, uma das principais características reprodutivas
para as espécies que apresentam determinação sexual dependente da temperatura
da incubação. A temperatura pivotal é aquela que define a produção de machos e de
fêmeas em proporções iguais e ainda não é muito bem conhecida para as tartarugas
de água doce brasileiras.
4
O mecanismo da determinação sexual dependente da temperatura da incubação já
foi comprovado para P. expansa (ALHO et al. 1984, MALVASIO 2001), P. unifilis
(SOUZA & VOGT 1994) e P. sextuberculata (PEZZUTI & VOGT 1999), entretanto todos
esses estudos foram conduzidos em campo onde as condições de temperatura são
muito variadas. LUBIANA & FERREIRA JÚNIOR (2009) determinaram a temperatura
pivotal para P. expansa, do rio Javaés (TO), analisando exclusivamente a influência
da temperatura, através da incubação dos ovos em temperaturas constantes, assim
como VALENZUELA (2001) que comparou dados de campo com dados obtidos em
laboratório (VALENZUELA et al. 1997). É desejável que a temperatura pivotal seja
avaliada considerando-se as mesmas condições de contorno e de experimentação.
Em campo, sob condições naturais, as variações da temperatura são muito grandes
(VALENZUELA et al. 1997) e dificultam a interpretação dos resultados (MALVASIO et al.
2002). Outros fatores como a umidade (PACKARD et al. 1991) a profundidade do
ninho (FERREIRA JÚNIOR & CASTRO 2003), e o sombreamento (GOMES & FERREIRA
JÚNIOR 2011), por exemplo, afetam o ambiente termal do ninho tornando difícil
apontar a influência exclusiva da temperatura (GEORGES et al. 2004). No laboratório
as temperaturas são mantidas constantes permitindo avaliar sua influência na
diferenciação das gônadas dos embriões.
Via de regra, os filhotes de quelônios não apresentam dimorfismo sexual nos
primeiros anos de vida, o que se torna evidente apenas quando atingem a
maturidade sexual (PRITCHARD & TREBBAU 1984). A despeito dos avanços nas
técnicas de identificação do sexo de filhotes recém-eclodidos, e de animais juvenis
por métodos não invasivos como a quantificação de hormônios esteróides (LANCE et
al. 1992), ou da aplicação dos princípios da morfometria geométrica (VALENZUELA et
al. 2004, LUBIANA & FERREIRA JÚNIOR 2009), a maneira mais segura e eficiente ainda
é a análise histológica das gônadas que requer, além do sacrifício dos animais,
5
procedimentos relativamente complexos e onerosos (W YNEKEN et al. 1988, MALVASIO
2001).
A proposta de investigação desse projeto visa preencher parte dessa lacuna, relativa
aos primeiros momentos do ciclo de vida, determinando a temperatura pivotal de P.
unifilis sob temperaturas constantes em laboratório e estimar a razão sexual dos
ninhos naturais. As informações sobre a temperatura pivotal serão correlacionadas
às características fisiográficas do local da desova, associando os locais de
nidificação à faixa de temperatura na qual os filhotes são gerados na natureza. As
características do meio físico podem influenciar o sucesso da eclosão (relação entre
o número de filhotes vivos sobre o número total de ovos viáveis), a razão sexual
(relação entre o número de fêmeas e o número total de filhotes de um ninho), e a
duração da incubação (período transcorrido entre a desova e a eclosão dos filhotes)
(FERREIRA JÚNIOR et al. 2008, FERREIRA JÚNIOR 2009a, b), apontando a importância
do local escolhido pela fêmea na reprodução. A temperatura da incubação guarda
uma relação direta com a taxa de desenvolvimento embrionário e nas espécies com
determinação sexual dependente da temperatura de incubação a duração da
incubação pode ser utilizada na predição do sexo dos filhotes e, por isso, pode ser
importante na definição de áreas prioritárias de preservação (MARCOVALDI et al.
1997, MROSOVSKY et al. 1999).
O presente trabalho foi desenvolvido sob a hipótese de que a razão sexual e o
sucesso de eclosão são dependentes das condições do contorno do ninho.
Testamos também a hipótese de que a temperatura média de incubação é
dependente dos aspectos sedimentológicos dos ninhos e que, portanto, o local
escolhido pela fêmea desempenha um papel crucial na reprodução de P. unifilis.
6
O objetivo do trabalho é determinar a razão sexual de P. unifilis baseado na
temperatura pivotal obtida em condições controladas de temperatura. Os objetivos
específicos do trabalho visam associar a duração de incubação e o sucesso de
eclosão dos ovos às características sedimentológicas das áreas de desova e
verificar se a duração de incubação pode ser utilizada como ferramenta preditiva da
razão sexual dos ninhos mantidos no ambiente natural.
7
2. FUNDAMENTAÇÃO TEÓRICA
Podocnemis unifilis é considerada uma espécie de tartaruga que ocupa pequenos
rios e lagos pouco profundos, sendo encontrada também em rios de maior vazão
(SANTOS 1994). Com o início do período chuvoso os rios transbordam e é comum a
migração de P. unifilis para ambientes calmos como lagos e florestas inundadas
(PRITCHARD & TREBBAU 1984).
Podocnemis unifilis apresenta cerca de 8 kg quando adulto, podendo o comprimento
do casco, que é ligeiramente côncavo, chegar a 68 cm de comprimento. Apresenta
manchas amareladas bem evidentes na parte dorsal da cabeça e nas bordas das
placas marginais da carapaça principalmente em filhotes. As manchas amarelas
permanecem vivas nos machos, mas tendem a desaparecer nas fêmeas adultas,
sendo uma característica que auxilia na identificação do sexo dos adultos. A cabeça
é alongada com focinho proeminente; não possuindo sulco interorbital. Possui uma
ou duas barbelas e quilha medial acentuada nos filhotes e restrita na região central
(2ª e 3ª vértebras nos adultos) (ERNST & BARBOUR 1989).
Segundo PRITCHARD & TREBBAU (1984), P. unifilis é primariamente herbívora.
Entretanto LEGLER (1993) aponta a maioria das espécies de Testudines como
onívoras oportunistas, apresentando o comportamento típico de onívoros aquáticos,
rondando perto do leito do rio, investigando a vegetação e as frutas caídas das
árvores, e aproveitando-se da disponibilidade de pequenos invertebrados. MALVASIO
(2001), em estudos no Estado do Tocantins, confirma a dieta predominantemente
herbívora e a ingestão de pequenas quantidades de insetos e crustáceos, para a
espécie.
8
Dados importantes no que diz respeito à reprodução de P. unifilis são relacionadas
às características do ninho. A fêmea desova entre 15 a 25 ovos por postura, sendo
estes alongados e de casca rígida, em um ninho de profundidade máxima de 30 cm
(ERNST & BARBOUR 1989). Diferentemente de outras espécies de tartarugas, cuja
profundidade dos ninhos pode variar entre 45 e 100 cm. Dessa forma os ninhos de
P. unifilis se tornam mais expostos à temperatura do meio externo e sujeitos às
variações de temperatura ao longo do dia.
Em um momento em que as mudanças climáticas são o foco de inúmeros debates
políticos, econômicos e, principalmente, ambientais é fundamental que se entenda a
influência da temperatura na reprodução de um dos mais importantes grupos de
répteis brasileiros e no qual estão inseridas espécies ameaçadas de extinção (IUCN
2011) juntamente com espécies de grande relevância sócio-cultural e econômica
(PEZZUTI et al. 2010). Somente assim será possível argumentar sobre as possíveis
conseqüências das mudanças do clima global na ecologia reprodutiva, e no
processo evolutivo desses animais (HAYS et al. 2003; GIRONDOT et al. 2004).
A determinação da temperatura pivotal sob condições controladas permite a
comparação de resultados entre espécies e populações, possibilitando a análise das
diferenças entre os hábitos reprodutivos e, conseqüentemente, entre os fatores
evolutivos de cada grupo (GIRONDOT et al. 2004). O conhecimento dessas
informações também é de grande utilidade para os projetos de conservação que
utilizam o manejo dos ovos, como alternativa para ampliar o sucesso de eclosão
(MARCOVALDI & LAURENT 1996, GOMES & FERREIRA JÚNIOR 2011). No caso de
transferências de ninhos, as diferenças no meio físico podem ocasionar mudanças
na temperatura, podendo afetar o desenvolvimento embrionário e causar alterações
na razão sexual dos ninhos transferidos (MILTON et al. 1999, NARO-MACIEL et al.
9
1999), o que pode afetar a estrutura populacional. Devido a este fator, o
conhecimento da razão sexual das espécies que apresentam determinação sexual
dependente da temperatura de incubação dos ovos é de extrema importância para o
sucesso das ações conservacionistas que utilizam do manejo dos ovos (MORREALE
et al. 1982, SPOTILA & STANDORA 1986, MROSOVSKY & PROVANCHA 1992, MROSOVSKY
1994).
A duração da incubação é sugerida como ferramenta preditiva da razão sexual para
tartarugas marinhas (MARCOVALDI et al. 1997, MROSOVSKY et al. 1999, GODFREY et al.
1999, BRODERICK et al. 2000, GODLEY et al. 2001). O método baseia-se na relação
entre a temperatura de incubação na taxa de desenvolvimento embrionário. Em
temperaturas elevadas o desenvolvimento embrionário é acelerado reduzindo a
duração de incubação. Alternativamente, quando as temperaturas são mais baixas o
desenvolvimento embrionário é mais lento levando a uma duração de incubação
mais longa. Nesse contexto surgiu o conceito de duração pivotal de incubação, que
determina a duração de incubação (em dias), responsável pela produção de cada
sexo. Por isso a duração de incubação pode apresentar-se como ferramenta viável
de predição da razão sexual também para quelônios continentais, uma vez que o
clima das regiões centrais dos continentes apresenta certa estabilidade quando
comparamos temporadas reprodutivas distintas. Contudo, esta metodologia vem
sendo contestada, tendo em vista que, variações climáticas durante o período termosensitivo podem gerar um sexo não esperado para o período de incubação dos
embriões (PIEAU 1982, GEORGES 1989, GEORGES 1992). Por exemplo, em um ninho
de Podocnemis expansa com o período de incubação de 70 dias espera-se a
prevalência de machos. No entanto, se durante o segundo terço da incubação
ocorrer um aumento repentino na temperatura, mesmo que por poucos dias, as
condições serão favoráveis ao desenvolvimento de fêmeas (BULL & VOGT 1981). Se
10
após esse período a temperatura voltar a cair, o desenvolvimento embrionário será
prolongado levando a uma duração pivotal de incubação condizente com uma razão
sexual favorável aos machos (PIEAU & DORIZZI 1981, GEORGES et al. 1994).
Comprovar e testar a uma relação entre a duração de incubação e a determinação
sexual de quelônios continentais é importante principalmente por se tratar de um
método de baixo custo, não invasivo e de aplicação relativamente simples. Em um
cenário onde os recursos e o conhecimento são escassos, esse método pode ser
uma importante ferramenta para estimar a razão sexual das populações e ampliar o
conhecimento sobre a ecologia das tartarugas (LUBIANA & FERREIRA JÚNIOR 2009).
A importância da temperatura na reprodução de quelônios foi comprovada por
MROSOVSKY & YNTEMA (1980), quando foi demonstrado experimentalmente que, em
laboratório, a diminuição de apenas 1 °C na temperatura de incubação causou um
aumento de cinco dias da duração de incubação e alterou significativamente a razão
sexual de filhotes de Caretta caretta. No entanto, em ambientes naturais, as
condições ambientais de ninhos rasos como P. unifilis (FERREIRA JUNIOR et al. 2007)
são muito variáveis e oscilam dentro de um ciclo diário, o que inviabiliza a
generalização desse método (GEORGES et al. 1994). Além disso, quando o universo
amostral é pequeno, a margem de erro pode ser elevada e invalidar a utilização da
duração da incubação como ferramenta de predição da razão sexual (MROSOVSKY et
al. 1999). As principais restrições ao uso da duração da incubação da estimativa da
razão sexual dos ninhos rasos das tartarugas continentais se baseiam nas variações
diárias da temperatura (PIEAU 1982). Por outro lado, as variações diárias da
temperatura nos ninhos de tartaruga marinha que são mais profundos é menos,
entre 0,5 e 1,0 para Chelonia mydas (MORREALE et al. 1982), o que possibilita o
estabelecimento de uma correlação entre a taxa de desenvolvimento embrionário e
11
a temperatura média do ninho e a utilização da duração pivotal de incubação na
estimativa da razão sexual dos ninhos.
Estudos que individualizam e quantificam separadamente os fatores ambientais
envolvidos na incubação são fundamentais para esclarecer a maneira como estes
fatores interferem no desenvolvimento embrionário de animais que apresentam
determinação sexual dependente da temperatura. Com um número amostral
ampliado é possível atingirmos a segurança necessária para indicar a duração da
incubação como uma ferramenta preditiva da razão sexual natural, diminuindo-se a
necessidade da utilização de métodos invasivos para a determinação do sexo dos
filhotes. Essa estimativa pode ser de grande utilidade para os projetos de
conservação que trabalham para ampliar ou manter a população de tartarugas,
como o Projeto Tarekaja’a Xingu, que busca restabelecer a população de P. unifilis
na bacia do Rio Xingu, junto à comunidade indígena da etnia Kamayurá.
O principal objetivo do Projeto Tarekaja’a Xingu é recuperar a população e promover
o manejo sustentável de P. unifilis (tracajá) no Parque Indígena do Xingu, no estado
do Mato Grosso (Projeto Tarekaja’a Xingu/EMBRAPA). O tracajá está fortemente
inserido na cultura indígena, fazendo parte dos aspectos culturais, folclóricos,
religiosos e alimentares. Algumas etnias indígenas, como a Kamayurá, acreditam
que o próprio Rio Xingu foi formado através de um tracajá que “deslizou por todo o
caminho trazendo água de volta para todas as tribos que passavam sede” formando
assim o rio e salvando os povos que ali vivem (Cultura Indígena).
As comunidades indígenas da porção superior do rio Xingu sempre utilizaram P.
unifilis como um de seus maiores e mais apreciados recursos alimentares, sendo
igualmente consumidos os ovos, filhotes e animais adultos, principalmente fêmeas
em período de desova (PEZZUTI et al. 2010). Entretanto há algumas décadas o
12
declínio da população de P. unifilis na região foi observado pelas próprias
comunidades indígenas. Este declínio é explicado por um conjunto de fatores
relacionados ao aumento da população indígena no parque, juntamente com o
crescimento das áreas de desmatamento e, pela introdução de novas técnicas e
ferramentas de captura introduzidas pelo “homem branco”, como o uso de barco a
motor, lanterna e espinhel. Esses fatores têm aumentado significativamente a
pressão de captura sobre os animais, que antes eram caçados com caminhadas
noturnas apenas nas proximidades das aldeias e com uso de flechas e arpões
(Projeto Tarekaja’a Xingu/EMBRAPA).
PEZZUTI et al. (2010) apontaram a importância dos quelônios continentais como
recurso alimentar e incremento protéico à dieta de populações indígenas e
ribeirinhas da região amazônica. A utilização deste recurso, bem como seus ovos e
seu óleo, envolve uma série de fatores que incluem preferências alimentares, tabus,
usos medicinais e até valores de mercado, sendo estes variáveis entre as regiões.
PEZZUTI et al. (2010) fazem referência sobre a preferência pela ingestão de fêmeas
por determinadas culturas, uma vez que os machos possuem sabor menos
agradável e existe a crença de que sua ingestão pode causar injúrias e não devem
ser consumidos em certas épocas (gravidez, lactação, período menstrual ou
acometido por doenças), o que pode aumentar ainda mais a pressão de captura
sobre as fêmeas.
A dificuldade de acesso a um recurso alimentar tão importante também é
preocupante no que diz respeito aos valores nutricionais que P. unifilis acrescenta à
dieta das comunidades, uma vez que praticamente todo incremento protéico da dieta
indígena do alto Xingu é proveniente da pesca e as tartarugas fornecem um alto
aditivo calórico à dieta. A preocupação em manter P. unifilis como recurso
13
sustentável surgiu da própria comunidade indígena, particularmente dos membros
da aldeia Kamayurá-Morená, que iniciaram um programa de proteção de ninhos e
filhotes, com intuito de promover o aumento da população a através da diminuição
da predação de ninhos e da mortalidade dos filhotes e animais adultos
(Comunicação pessoal).
O rio Xingu apresenta uma grande heterogeneidade ambiental em suas praias, com
sedimento em diversas frações granulométricas, disponibilizando uma área de
grande diversidade para a postura dos ovos. Observa-se em uma mesma praia
zonas com graus distintos de sombreamento e vegetação de entorno, altura
diferenciada em relação ao leito do rio, diferentes frações granulométricas
semelhante ao que foi descrito para outros ambientes fluviais do rio Crixás-Açu
(FERREIRA JÚNIOR & CASTRO 2006), Araguaia (FERREIRA JÚNIOR & CASTRO 2005) e no
rio Javaés (FERREIRA JÚNIOR et al. 2003). Todos esses fatores interferem diretamente
no desenvolvimento dos embriões (FERREIRA JÚNIOR & CASTRO 2010), tornando as
praias do rio Xingu um local apropriado para o desenvolvimento de estudos sobre a
ecologia reprodutiva de P. unifilis.
FERREIRA JÚNIOR & CASTRO (2003, 2006) e FERREIRA JÚNIOR et al. (2007)
demonstraram a preferência de P. unifilis em desovar nas áreas mais baixas,
próximas ao nível do rio, tornando os ninhos mais susceptíveis ao alagamento e
morte dos embriões. O alagamento é a principal causa de perdas de ninhos para as
espécies de quelônios continentais devido à instabilidade climática, que causa a
subida repentina dos rios e afogamento dos embriões de ninhos das áreas mais
baixas. Para P. unifilis e P. expansa o alagamento é uma das principais causas de
perda de ninhos e filhotes nas mais diferentes áreas de ocorrência destas espécies
(ALHO & PÁDUA 1982, IBAMA 1989, MITCHELL & QUIÑONES 1994, SOINI 1995,
14
FERREIRA JÚNIOR & CASTRO, 2005, 2006). Para evitar perda dos ovos o RAN/ICMBio
recomenda a transferência dos ninhos para locais mais altos ou mais protegidos de
predação por animais. Essa estratégia é utilizada há algum tempo por projetos de
conservação de quelônios, como o Projeto TAMAR/ICMBio e o Projeto Quelônios da
Amazônia/RAN/ICMBio (IBAMA 1989). No entanto quando se utiliza a transferência
de ninhos deve-se atentar não apenas para altura da praia em relação ao ninho ou à
proteção contra predadores, mas também se o local escolhido para receber os ovos
apresenta características semelhantes ao local da desova. Essa preocupação se
deve ao fato de que alterações no tamanho e na composição mineralógica dos
sedimentos podem alterar substancialmente o perfil de transmissão de calor no
interior o ninho (MILTON et al. 1999, FERREIRA JÚNIOR et al. 2007, FERREIRA JÚNIOR &
CASTRO 2006), interferindo no desenvolvimento e alterando a determinação do sexo
dos embriões. O ideal, nesse caso, é que o ambiente que irá receber os ovos
proporcione condições de desenvolvimento que assegure uma razão sexual próxima
daquela encontrada no local de origem do ninho. A razão sexual de equilíbrio para
espécies com determinação sexual dependente da temperatura de incubação é
desconhecida e de difícil determinação, uma vez que as interações entre fatores
ambientais e biológicos são muito diversas (VALENZUELA & LANCE 2004). Assim,
ressaltamos a importância da execução de experimentos laboratoriais conjuntos a
trabalhos de campo, a fim de se estabelecer a razão sexual e as dinâmicas
populacionais das espécies, para que os resultados sejam aplicados em projetos de
manejo e conservação.
A obtenção de dados, como a razão sexual dos filhotes, a temperatura pivotal, as
faixas de temperatura responsáveis pela produção de cada sexo e a influência do
local da desova no desenvolvimento embrionário são fundamentais para estabelecer
as melhores estratégias de manejo para a população em questão.
15
REFERENCIAS BIBLIOGRÁFICAS 1
ALHO, C.J.R. & L.F.M. PÁDUA. 1982. Sincronia entre o regime de vazante do rio e o
comportamento de nidificação da tartaruga da Amazônia Podocnemis expansa
(Testudinata: Pelomedusidae). Acta Amazônica 12(2): 323-326.
ALHO, C.J.R.; T.M.S. DANNI & L.F.M. PÁDUA. 1984. Influência da temperatura de
incubação na determinação do sexo da tartaruga-da-amazônia (Podocnemis
expansa, Testudinata: Pelomedusidae). Revista Brasileira de Biologia 44(3): 305311.
BAPTISTOTTE, C.; J.T. SCALFONI & N. MROSOVSKY. 1999. Male-producing thermal
ecology of a southern loggerhead turtle nesting beach in Brazil: implications for
conservation. Animal Conservation 2: 9-13.
BRODERICK, A.C.; B.J. GODLEY; S. REECE & J.R DOWNIE. 2000. Incubation periods and
sex ratios of green turtles: highly female biased hatchling production in the eastern
Mediterranean. Marine Ecology Progress Series 202: 273-281.
BULL, J.J. 1980. Sex determination in reptiles. Quarterly Review of Biology 53(1):
3-20.
ERNST, C.H.; BARBOUR, R.W. 1989. Turtles of the World. Washington, Smithsonian
Institution Press, 313 p.
FERREIRA JÚNIOR, P.D. 2009a. Aspectos ecológicos da determinação sexual em
tartarugas. Acta Amazonica 39(1): 139-154.
FERREIRA JÚNIOR, P.D. 2009b. Efeitos de fatores ambientais na reprodução de
tartarugas. Acta Amazonica 39(2): 319-334.
16
FERREIRA JÚNIOR, P.D. & P.T.A CASTRO. 2003. Geological control of Podocnemis
expansa and Podocnemis unifilis nesting areas in Rio Javaés, Bananal Island, Brazil.
Acta Amazonica 33(3): 445-468.
FERREIRA JÚNIOR, P.D. & P.T.A. CASTRO. 2005. Nest placement of the giant Amazon
river turtle, Podocnemis expansa, in the Araguaia River, Goiás State, Brazil. Ambio
34(3): 212-217.
FERREIRA JÚNIOR, P.D.; O.S. GUIMARÃES & A. MALVASIO. 2003. The influence of
geological factors on reproductive aspects of Podocnemis unifilis (Testudines,
Pelomedusidae), on the Javaés river, Araguaia National Park, Brazil. Chelonian
Conservation and Biolology 4(3): 626-634.
FERREIRA JÚNIOR, P.D.; A.Z. CASTRO & P.T.A CASTRO. 2007. The importande of
nidification environment in the Podocnemis expansa and Podocnemis unifilis
phenotypes
(Testudines:
Podocnemididae).
South
American
Journal
of
Herpetology 2(1): 39-46.
FERREIRA JÚNIOR, P.D.; M.F. ROSA; M. LORENZO; M.F. MONTEIRO & R.R. AZEVEDO
JUNIOR. 2008. Influência dos sedimentos na reprodução de Caretta caretta na praia
da Guanabara, Anchieta, Espírito Santo, Brasil. Iheringia. Série Zoologia 98(4):
447-453.
FERREIRA JÚNIOR, P.D. & P.T.A. CASTRO. 2010. Nesting ecology of Podocnemis
expansa (Schweigger, 1812) and Podocnemis unifilis (Troschel, 1848) (Testudines,
Podocnemididae) in the Javaés River, Brazil. Brazilian Journal of Biology 70: 8594.
GEORGES, A. 1989. Female turtle from hot nests: Is it duration of incubation or
proportion of development that matters? Oecologia 81: 323-328.
17
GEORGES, A. 1992. Thermal characteristics and sex determination in field nests of the
pig-nosed turtle, Carattochelys insculpta (Chelonia: Carettochelydidae), from
Australia. Australian Journal of Zoology 40: 511-521.
GEORGES, A.; C. LIMPUS & R. STOUTJESDIJK. 1994. Hatchling sex in the marine turtle
Caretta caretta is determined by proportion of development at a temperature, not
daily duration of exposure. Journal of Experimental Zoology 270 (3): 432-444.
GEORGES, A.; S. DOODY; K. BEGGS & J. YOUNG. 2004. Thermal models of TDS under
laboratory and field conditions. In: Valenzuela, N.; Lance, V.A. TemperatureDependent Sex Determination in Vertebrates (Eds). Smithsonian Books,
Washington. p. 79-89.
GIRONDOT, M; P.R. DELMAS; F. COURCHAMP; A.C. PRÉVOT-JULLIARD & M.H GODFREY.
2004. Implications of temperature-dependent sex determination for population
dynamics.
In:
Valenzuela,
N.;
Lance,
V.A.
Temperature-Dependent
Sex
Determination in Vertebrates (Eds). Smithsonian Books, Washington. p. 148-155.
GODFREY, M.H.; A.F; D’AMATO; M.Â. MARCOVALDI & N. MROSOVSKY. 1999. Pivotal
temperature and predicted sex ratios for hatchling hawksbill turtles from Brazil.
Canadian Journal of Zoology 74(9): 1465-1473.
GODLEY, B.J.; A.C. BRODERICK & N. MROSOVSKY. 2001. Estimating hatchling sex ratios
of loggerhead turtles in Cyprus from incubation durations. Marine Ecology Progress
Series 210:195-201.
Gomes, A.S. & Ferreira Júnior, P.D.F. 2011. Management and relocation of nests of
Podocnemis expansa (Scheweigger, 1812) (Testudines, Podocnemididade) on the
Crixás-Açu River, Brazil. Brazilian Journal of Biology 71 (4): 975-982.
18
HAYS, G.C; A.C. BRODERICK; F. GLEN & GODLEY, B.J.2003. Climate change and sea
turtles: a 150-year reconstruction of incubation temperatures at a major marine turtle
rookery. Global Change Biology 9: 642-646.
IBAMA – INSTITUTO BRASILEIRO
DO
MEIO AMBIENTE
E DOS
RECURSOS NATURAIS
RENOVÁVEIS. 1989. Projeto Quelônios da Amazônia, 10 anos. Brasília, IBAMA,
119p.
IUCN 2011. IUCN Red List of Threatened Species. Version 2011.2. Available online
at: http://www.iucnredlist.org. [Accessed: 01de dezembro de 2011].
KEMENES, A. & J.C.B. PEZZUTI. 2007. Estimate of trade traffic of Podocnemis
(Testudines, Pedocnemididae) from the middle Purus River, Amazonas, Brazil.
Chelonian Conservation and Biology 6 (2): 259-262.
LANCE, V.A., N. VALENZUELA & H.
VON
HILDEBRAN. 1992. A hormonal method to
determine the sex of hatchling giant river turtle, Podocnemis expansa. Application to
endangered species research. American Zoologist 32:16A.
Legler, J. M. 1993. Morphology and Physiology of the Chelonia. In: GLASBY, C.J. et
al. (Eds.). Fauna of Austrália 2A, cap.16: 108-119.
LUBIANA, A. & P.D. FERREIRA JÚNIOR. 2009. Pivotal temperature and sexual
dimorphism of Podocnemis expansa hatchlings (Testudines: Podocnemididae) from
Bananal Island, Brazil. Zoologia 26 (3): 527-533.
MALVASIO, A. 2001. Aspectos do mecanismo alimentar e da biologia reprodutiva
em Podocnemis expansa (Schweigger, 1812), P. unifilis (Troschel, 1848) e P.
sextuberculata
(Cornalia,
1849)
(Testudines,
Emydidae).
São
Paulo,
19
Universidade de São Paulo. 199p. (Tese de Doutorado em Zoologia, Instituto de
Biociências da USP)
MALVASIO, A.; A.M. SOUZA; P.D. FERREIRA JÚNIOR; E.S. REIS & F.A.A. SAMPAIO. 2002.
Temperatura de incubação dos ovos e granulometria dos sedimentos das covas
relacionadas à determinação sexual em Podocnemis expansa (Schweigger, 1812) e
P. unifilis (Troschel, 1848) (Testudines, Pelomedusidae). Publicações Avulsas do
Instituto Pau Brasil de História Natural 5(1): 11-25.
MARCOVALDI, M.Â.; M.H. GODFREY & N. MROSOVSKY. 1997. Estimating sex ratios of
loggerhead turtles in Brazil from pivotal incubation durations. Canadian Journal of
Zoology 75:755-770.
MARCOVALDI, M.A. & LAURENT, A. 1996. A six season study of marine turtle nesting at
Praia do Forte, Bahia, Brazil, with implications for conservation and management.
Chelonian Conservation Biology 2:55–59.
MILTON, S.L.; A.A. SCHULMAN & P.L. LUTZ. 1997. The effect of beach nourishment with
aragonite versus silicate sand on beach temperature and loggerhead sea turtle
nesting success. Journal of Coastal Research 13(3): 904-915.
MITCHELL, C. & L QUIÑONES. 1994. Manejo y conservacion de la taricaya (Podocnemis
unifilis) en la Reserva de Biofera del Manu, Madre de Dios. Boletin de Lima 16: 425436.
MORREALE, S.J; G.J. RUIZ; J.R. SPOTILA & E.A. STANDORA. 1982. Temperaturedependent sex determination: current pratices threaten conservation of sea turtles.
Science 216(11): 1245-1247.
20
MROSOVSKY, N. 1994. Sex ratios of sea turtles. The Journal of Experimental
Zoology 270(1): 16-27.
MROSOVSKY, N. & C.L. YNTEMA. 1980. Temperature dependence of sexual
differentiation in sea turtles: implications for conservation practices. Biological
Conservation 18: 271-280.
MROSOVSKY, N. & J. PROVANCHA. 1992. Sex ratio of hatchling loggerhead sea turtles:
data and estimates from a 5-year study. Canadian Journal of Zoology 70: 530-538.
MROSOVSKY, N., C. BAPTISTOTTE & M.H. GODFREY. 1999. Validation of incubation
duration as an index of the sex ratio of hatchling sea turtles. Canadian Journal of
Zoology 77(5): 831-835.
NARO-MACIEL, E.; N. MROSOVSKY & M.Â. MARCOVALDI. 1999. Thermal profiles of sea
turtle hatcheries and nesting areas at Praia do Forte, Brazil. Chelonian
Conservation and Biology 3(3): 407-413.
PACKARD, G.C.; M.J. PACKARD & L. BENIGAN. 1991. Sexual differentiation, growth, and
hatching success by embryonic painted turtles incubated in wet and dry
environments at fluctuating temperatures. Herpetologica 47(1): 125-132.
PEZZUTI, J.C.B. & R.C. VOGT. 1999. Nesting ecology of Podocnemis sextuberculata
(Testudines, Pelomedusidae) in the Japurá River, Amazonas, Brazil. Chelonian
Conservation and Biology 3(3): 419-424
PEZZUTI, J.C.B.; G.H. REBELO; G. MOREIRA & L. LUGLI,. 2005. Pesca artesanal de
quelônios no Parque Nacional do Jaú. Boletim do Museu Paraense Emilio Goeldi
1(1): 109-125.
21
PEZZUTI, J.C.B; J.P.LIMA; D.F. SILVA & A. BEGOSSI. 2010. Uses and taboos of turtles
and tortoises along Rio Negro, Amazon Basin. Journal of Ethnobiology 30(1): 153–
168.
PIEAU, C. 1982. Modalities of the action of temperature on sexual differentiation in
field developing embryos of the European pond turtle Emys orbicularis (Emydidae).
Journal of Experimental Zoology 220: 353-360.
PIEAU, C. & M. DORIZZI. 1981. Determination of temperature sensitive stage for sexual
differentiation of the gonads in embryos of the turtle, Emys orbicularis. Journal of
Morphology 170: 373-382.
PRITCHARD, P.C.H. & P. TREBBAU. 1984. The turtles of Venezuela. Oxford, Society
for the Study of Amphibians and Reptiles, 403p.
RUEDA-ALMONACID, J.V. ; J.L. CARR; R.A. MITTERMEIER; J.V. RODRÍGUEZ-MAHECHA;
R.B. MAST; R.C. VOGT; A.G.J. RHODIN; J. OSSA-VELÁSQUEZ; J.N. RUEDA & C.G.
MITTERMEIER. 2007. Las tortugas y los cocodrilianos de los países andinos del
Trópico. Bogotá, Colombia, Conservación Internacional, 537p.
SOINI, P. 1995. Investigaciones en la Estación Biológica Cahuana. Reporte PacayaSamiria, Universidad Nacional Agraria La Molina, 435p.
SOUZA, R.R. & R.C. VOGT 1994. Incubation temperature influences sex and hatchling
size in the neotropical turtle Podocnemis unifilis. Journal of Herpetology 28(4): 453464.
Spotila, J.R. & E.A. Standora. 1986. Sex determination in the desert tortoise: a
conservative management strategy is needed. Herpetologica 42(1):67-72
22
VALENZUELA, N.; R. BOTERO; & E. MARTÍNEZ. 1997. Field study of sex determination in
Podocnemis expansa from Colombian Amazonia. Herpetologica 53(3):390-395.
VALENZUELA, N. 2001.
Constant, shift, and natural temperatures effects on sex
determination in Podocnemis expansa turtles. Ecology 82: 30 10-3024.
VALENZUELA N. & V.A. LANCE. 2004. Temperature dependent sex determination in
vertebrates. Washington D.C, Smithsonian Books.
W YNEKEN, J.; T.J. BURKE; M. SALMON & D.K. PEDERSEN. 1988. Egg failure in natural
and relocated sea turtle nests. Ecology 79(6): 1884-1892.
23
3. CAPÍTULOS
Os capítulos a seguir foram redigidos seguindo as normas de citações e
referências da revista Zoologia (ISSN 1984-4670), no entanto não se encontram formatados
para submissão, uma vez que alterações serão consideradas, de acordo com correções ou
sugestões propostas pela banca examinadora. As normas encontram-se disponíveis em:
http://www.scielo.br/revistas/zool/iinstruc.htm
3.1
CAPÍTULO I
RAZÃO SEXUAL DE PODOCNEMIS UNIFILIS (TROSCHEL 1848) (TESTUDINES:
PODOCNEMIDIDAE) NO PARQUE INDÍGENA DO XINGU, MATO GROSSO, BRASIL.
Aline Silva Gomes1,3, José Roberto Moreira², Fábio de Oliveira Freitas² &
Paulo Dias Ferreira Júnior¹
1. Programa de Pós-graduação em Ecologia de Ecossistemas, Universidade Vila
Velha, Rua Comissário José Dantas de Melo, 21, Boa Vista, 29102-770, Vila
Velha, ES, Brasil.
2. Embrapa Recursos Genéticos e Biotecnologia, Parque Estação Biológica, Av.
W5 Norte, Caixa Postal 02372, 70770-917, Brasília, DF, Brasil.
3. [email protected], autor correspondente.
RESUMO
A razão sexual dos filhotes é fundamental para o conhecimento da dinâmica
populacional e história de vida das espécies. Neste trabalho estimamos a razão
sexual de ninhos naturais de Podocnemis unifilis no Parque Indígena do Xingu, no
estado do Mato Grosso. Para isto, estabelecemos a temperatura pivotal desta
população a partir da incubação em laboratório sob temperaturas constantes. A
temperatura pivotal de P. unifilis no Parque Indígena do Xingu é de 31,8 °C. Para o
cálculo da razão sexual foi monitorada a temperatura de 120 ninhos in situ durante
toda a incubação. A razão sexual foi estimada com base na temperatura média no
segundo terço de incubação e no CTE (Equivalente de Temperatura Constante),
estabelecido a partir das variações diárias de temperatura nos ninhos. Para a
temporada reprodutiva de 2010, 47% dos filhotes de P. unifilis eram fêmeas e a
24
duração de incubação média foi de 92 dias. Foi possível estabelecer uma correlação
entre o CTE e a duração de incubação, indicando que este pode ser utilizado na
estimativa da razão sexual.
Palavras-chave: Incubação, Temperatura pivotal, Tracajá, manejo, CTE.
INTRODUÇÃO
A determinação do sexo de Podocnemis unifilis (TROSCHEL 1848) depende da
temperatura de incubação dos ovos (SOUZA & VOGT 1994), sendo classificada como
tipo TSD Ia (EWERT et al. 1994). Neste modelo, machos são gerados em
temperaturas baixas e fêmeas em temperaturas elevadas. A temperatura
responsável pela produção de machos e fêmeas em proporções iguais (1:1) é
denominada temperatura pivotal (PIEAU 1976, BULL 1980, YNTEMA & MROSOVSKY
1982). Em um regime de temperaturas variáveis, em que as oscilações diárias são
marcantes, são as variações acima e abaixo da temperatura pivotal que
determinarão o sexo dos embriões (GEORGES et al. 2004).
A determinação da temperatura pivotal é importante, pois ela pode variar entre as
espécies e entre populações de acordo com a região geográfica (EWERT & NELSON
1991, EWERT et al. 1994). Uma espécie com ampla distribuição pode apresentar
variações em torno de sua temperatura pivotal ao longo de sua área de ocorrência,
como no caso de Chelydra serpentina e Trachemys scripta, (EWERT et al. 1994,
2004). Este mecanismo visa compensar as variações climáticas (EWERT & NELSON
1991, EWERT et al. 1994), uma vez que para a manutenção de uma população viável
é necessário que a temperatura pivotal seja condizente com as características
ambientais, para que ambos os sexos sejam gerados (BULL et al. 1982).
25
O conhecimento da temperatura pivotal para cada população pode auxiliar na
escolha da técnica de manejo adequada as necessidades de cada uma delas (VOGT
1994). Devido à interação de fatores ambientais e biológicos, torna-se difícil
determinar a uma razão sexual de equilíbrio para uma população com a
determinação sexual dependente da temperatura de incubação (VALENZUELA &
LANCE 2004). Desta forma, experimentos em laboratório e trabalhos de campo se
tornam necessários para estabelecer a razão sexual de diferentes populações,
principalmente em projetos de manejo e conservação que envolvem o manuseio dos
ninhos (LUBIANA & FERREIRA JÚNIOR 2009).
Na incubação em laboratório sob temperatura constante ou em ninhos naturais com
pequena variação diária da temperatura, como por exemplo, os ninhos de tartaruga
marinha cuja câmara de ovos situa-se a mais 50 cm de profundidade (NARO-MACIEL
et al. 1999), o uso da temperatura média no segundo terço da incubação apresenta
uma alta correlação com a razão sexual, permitindo inclusive que a duração da
incubação seja utilizada na predição da proporção de filhotes machos e fêmeas
(MROSOVSKY et al. 1997, Marcovaldi et al. 1999, GODFREY et al. 1999).
Em estudos realizados em campo, onde vários fatores exercem influência na
temperatura, é importante que além da temperatura pivotal, se conheça a dinâmica
das variações diárias da temperatura (GEORGES et al. 2004). As oscilações em torno
da temperatura pivotal terão grande influência no sexo dos embriões, uma vez que a
diferenciação gonadal é influenciada pelo tempo acima ou abaixo da temperatura
pivotal, no qual o desenvolvimento embrionário ocorre (BULL et al. 1985, MROSOVSKY
et al. 1984, GEORGES 1989). O simples conhecimento da temperatura pivotal e da
temperatura média de incubação, portanto, não são suficientes para estabelecer a
razão sexual de ninhos naturais (GEORGES et al. 2004). Para contornar este
26
problema, GEORGES et al. (1994) sugeriram a utilização do “Equivalente de
Temperatura Constante” (CTE), que consiste em um modelo matemático que
sintetiza as variações diárias de temperatura em um único valor, estabelecendo uma
relação direta com a taxa de desenvolvimento embrionário (GEORGES et al. 2004).
No rio Xingu os dados sobre a ecologia reprodutiva de quelônios, especialmente de
P. unifilis, espécie mais abundante na região, são escassos, não havendo
informações relevantes sobre a estrutura e dinâmica populacional. O conhecimento
da temperatura pivotal e do CTE será fundamental para a conservação e o manejo
da espécie, evitando que as atividades de manejo que atualmente são executados
nesta área possam afetar a razão sexual que normalmente seria gerada nos ninhos
naturais.
Realizamos o presente trabalho sob a hipótese de que o desenvolvimento de macho
e fêmeas é dependente das variações diárias da temperatura de incubação. O
objetivo deste trabalho foi determinar a razão sexual dos ninhos in situ de P. unifilis
da porção norte do rio Xingu.
MATERIAIS E MÉTODOS
O estudo foi conduzido no Parque Indígena do Xingu (PIX), no estado do Mato
Grosso, junto à área de influência da aldeia Kamayurá-Morená (53°33’W 11°56S),
onde é executado o Projeto Tarekaja’a Xingu, que em conjunto com a comunidade
indígena monitora, maneja e protege as desovas de P. unifilis. O PIX possui, cerca
de 30.000 km², abrigando uma população de aproximadamente 6.000 índios (IBGE,
2010). A aldeia Kamayurá-Morená está localizada na região norte do PIX, próxima à
confluência dos rios Batovi, Kuluene e Ronuro (MONDINI et al. 2007) (Figura 1).
27
Figura 1. Localização da área de estudo com as praias utilizadas na nidificação por P. unifilis
(FERREIRA JÚNIOR et al. 2011)
O território da aldeia possui nove praias fluviais arenosas, normalmente monitoradas
desde 2007 pelos agentes do Projeto Tarekaja’a Xingu, a partir do mês de junho,
quando as fêmeas de P. unifilis iniciam a desova.
28
Para a determinação da temperatura pivotal foram coletados 150 ovos oriundos de
10 ninhos das praias do entorno da aldeia. Todos os ovos foram coletados na
manhã seguinte à desova, foram identificados e armazenados em caixa de isopor
preenchida com areia e vermiculita umedecida. Esse procedimento protegeu os ovos
contra rotações e choques mecânicos durante o transporte e garantiu a manutenção
de uma temperatura compatível com o desenvolvimento dos embriões. Os ovos
foram levados para o Laboratório de Ecologia Aquática e Terrestre, no Complexo de
Atividades Biopráticas, Centro Universitário Vila Velha, onde foram distribuídos
uniformemente entre dez incubadoras, com temperaturas variando entre 24 ºC e 35
ºC (variação aproximada de 1 ºC entre cada incubadora). Cada caixa foi preenchida
com vermiculita umedecida. Juntamente com os ovos foi inserido um termógrafo
digital do tipo i-button (Maxim DSG1921G), programado para medir e registrar a
temperatura a cada quinze minutos, durante toda a incubação.
Após a eclosão, os filhotes permaneceram na incubadora por aproximadamente
cinco dias para completar a absorção do vitelo e o endurecimento da carapaça. Os
filhotes foram marcados e encaminhados ao Biotério, também localizado no
Complexo de Atividades Biopráticas, onde permaneceram por aproximadamente três
meses. Os animais foram eutanasiados com uma injeção intracardíaca de 0,5 mL de
tiopental sódico diluído a 25%. As gônadas foram extraídas para preparação de
lâminas histológicas, coradas com HE, e analisadas segundo os critérios descritos
por MALVASIO et al. (2002) e classificadas em ovários ou testículos.
Após a sexagem dos filhotes e do estabelecimento da razão sexual de cada
incubadora foi utilizado software TSD V.3.2.2 (GIRONDOT 1999, GODFREY & DELMAS
2003), para o cálculo da temperatura pivotal. Este programa permite escolher entre
cinco descrições matemáticas, a equação que melhor se ajusta ao estabelecimento
29
da correlação entre a razão sexual e a temperatura, com base no AIC (Akaike
Information Criterion), além de apresentar o erro padrão, permitindo a comparação
com outros estudos (GIRONDOT 1999).
Simultaneamente à incubação no laboratório, foi monitorada a temperatura de 120
ninhos nas praias do rio Xingu com termógrafos i-button (Maxim DSG1921G),
inseridos no interior da câmara de ovos. Estes termógrafos registraram a
temperatura a cada uma hora durante toda a incubação.
A razão sexual destes ninhos foi estimada de duas maneiras, a partir da temperatura
média no segundo terço de incubação (Temp ⅔) e do Equivalente de Temperatura
Constante (CTE). O CTE se baseia no princípio de grau-dia e é indicado para
estimar a razão sexual em um regime de temperatura variável (GEORGES et al. 1994,
2004). Para o estabelecimento do CTE é necessário que se conheça a taxa de
desenvolvimento embrionário. Neste trabalho utilizou-se a relação obtida na
incubação sob temperaturas constantes em laboratório (GEORGES 1989, DEMUTH
2001, CHU et al. 2002, BOOTH & FREEMAN 2006) durante o segundo terço da
incubação, período termo sensível da determinação sexual, (YNTEMA & MROSOVSKY
1982). A relação encontrada neste trabalho, para P. unifilis, é expressa por:
Duração de Incubação (dias) = 182 – (3, 727 x temperatura de incubação °C)
Desta forma a fração do desenvolvimento embrionário que ocorre durante
determinado período do dia é dado por:
1/ (-3,827+ 185,63 T)
Onde T é a temperatura do intervalo de tempo. A soma de todas as frações do
desenvolvimento embrionário durante o segundo terço de incubação foi multiplicada
por 0,34 (a fração do desenvolvimento no qual o sexo é definido), permitindo o
30
estabelecimento do CTE para um dado ninho (BOOTH & FREEMAN 2006, CHU et al.
2008).
Análise Estatística
Para o cálculo da razão sexual dos ninhos foi utilizado o programa TSD V.3.2.2
(GODFREY et al.1994), que fornece os resultados a partir da temperatura. Para cada
ninho foram calculadas duas razões sexuais, a primeira a partir da temperatura
média, e a segunda do CTE, ambas utilizando o segundo terço da incubação. Para
comparar a razão sexual nestes dois tratamentos foi aplicado o teste não
paramétrico de Kruskal- Wallis. Para testar a relação do CTE e da temperatura
média no segundo terço com a duração de incubação e com o sucesso de eclosão
foi utilizada regressão linear e as equações de cada reta foram estabelecidas. Estas
análises foram realizadas com o auxílio do software Sigma Stat 3.5, assumindo
significância mínima de 95% (p < 0,05).
RESULTADOS
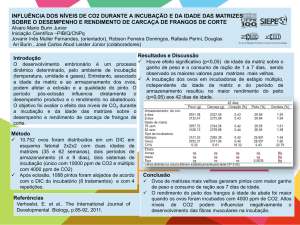
A temperatura pivotal de Podocnemis unifilis do rio Xingu foi de 31,77° C (Figura 2).
Os valores de temperatura e razão sexual utilizados para o cálculo da razão sexual
são apresentados na tabela 1.
Tabela 1. Razão sexual de P. unifilis incubados em temperatura constante.
Temperatura Média
n
Razão Sexual (%M)
Duração de Incubação (dias)
28,4 ± 0,04
10
90
84
30,3 ± 2,11
8
75
63
30,9 ± 0,16
8
62,5
57
32,1 ± 0,19
15
46,6
65
31
Figura 2. Relação entre a freqüência de fêmeas em diferentes temperaturas de incubação
constantes. A intercessão das linhas tracejadas representa o ponto onde às proporções de
machos e fêmeas são equivalentes (31,77 °C).
Para o calculo do CTE foi utilizada a correlação existente entre a temperatura de
incubação e a duração de incubação (Figura 3). O CTE apresentou maior relação
com as variáveis avaliadas (duração de incubação, razão sexual e sucesso de
eclosão) que a temperatura média no segundo terço da incubação (Temp. ⅔)
(Tabela 2).
Existem diferenças significativas entre o CTE e a Temp. ⅔ (H= 54, 601; gl= 1; p< 0,
001) e a razão sexual gerada por elas, de modo que a razão sexual para o CTE foi
de 47, 0 ± 0,28% de fêmeas, enquanto que para a Temp. ⅔ foi de 17,2 ± 0,07% de
fêmeas. A análise de variância demonstrou que existe diferença altamente
significativa entre as temperaturas (H= 55, 365; gl= 1; p< 0,001) (Figura 4).
32
Figura 3. Relação entre a temperatura e a duração de incubação em laboratório. Essa
relação é utilizada para o calculo da taxa de desenvolvimento.
Figura 4. Diferença entre temperatura média (Temp. ⅔) e o Equivalente da Temperatura
Constante (CTE) do segundo terço da incubação.
33
Tabela 2. Relação existente entre a temperatura média do segundo terço da incubação
(Temp. ⅔) e o Equivalente da Temperatura Constante (CTE) com a duração de incubação
(DI), razão sexual (RS) e sucesso de eclosão (SE). Note que o R² é sempre maior para o
CTE.
Temp. ⅔
CTE
Equação
R²
p
Equação
R²
p
DI
CTE= 6, 465 + (0,273 DI)
0,582
< 0,001
Temp. ⅔= 31, 210 - (0,0211DI)
0,017
0,23
RS
CTE = 27,501 +(8,687RS)
0,94
< 0,001
Temp. ⅔= 27, 330 +(11,35RS)
0,596
< 0,001
SE
CTE= 34, 257 - (3,280SE)
0,11
< 0,001
Temp. ⅔= 30, 036 - (0,918SE)
0,04
0,04
DISCUSSÃO
A temperatura pivotal de Podocnemis unifilis no rio Xingu foi de 31,8 °C, próxima dos
32 °C estabelecidos por SOUZA & VOGT (1994) para o rio Guaporé (RO). A
semelhança das temperaturas pivotais destas duas populações é explicada pela
localização dos rios Xingu (latidude 11,57°) e Guaporé (latitude 12,30° S). Dessa
forma, as populações de P. unifilis que habitam estas regiões experimentam
temperaturas de incubação muito semelhantes, o que leva ao estabelecimento de
uma temperatura pivotal similares (PRITCHARD & TREBBAU 1984).
A temperatura pivotal de P. unifilis no rio Xingu é próxima às temperaturas pivotais
descritas para outros Podocnemidídeos, como P. expansa (33,5 °C) (LUBIANA &
FERREIRA JÚNIOR 2009), P. sextuberculata (31,5 °C) (PEZZUTTI et al. 1998) e P.
lewyana (33,4 °C) (PAÉZ et al. 2009). P. expansa, apresenta distribuição geográfica
mais próxima à de P. unifilis. No entanto fatores intrínsecos à biologia das espécies
levaram ao estabelecimento de uma temperatura pivotal mais elevada, como por
exemplo, o comportamento reprodutivo de P. expansa, que desova no final do
período de seca e apresenta um ninho mais profundo, o que os torna mais
34
susceptíveis ao alagamento. Desta forma uma temperatura mais elevada propicia
um desenvolvimento embrionário acelerado encurtando a duração de incubação e
permitindo a eclosão dos ovos (FERREIRA JÚNIOR & CASTRO 2006).
Se comparadas com espécies de tartarugas marinhas, que também desovam em
áreas abertas e ensolaradas, e possuem temperatura pivotal em torno de 29 °C
(MROSOVSKY
1988,
MARCOVALDI
et
al. 1997),
a
temperatura
pivotal
dos
Podocnemidídeos é mais elevada (VALENZUELA & LANCE 2004). Essa característica é
explicada pela ocorrência tropical dos Podocnemidídeos, (PRITCHARD & TREBBAU
1984), assim a incubação comumente ocorre em temperaturas elevadas, o que
promove o aumento da temperatura pivotal como resultado da seleção natural, uma
vez que a temperatura pivotal mais alta, nesses casos, garantirá o nascimento de
ambos os sexos (BULL et al. 1982). Quelônios continentais sofrem com a
instabilidade climática, que promove a subida repentina ou inconstante dos rios.
Desta forma uma duração de incubação mais curta é essencial para as espécies que
desovam em praias fluviais tropicais, o que não ocorre com os ninhos de tartarugas
marinhas, que não estão sujeitos ao alagamento e apresentam ninhos relativamente
mais frios, um desenvolvimento mais lento, e uma temperatura pivotal mais baixa
(BULL & VOGT 1981, PIEAU & DORIZZI 1981, MROSOVSKY et al. 1984, Marcovaldi et al.
1997).
Mesmo sem a validação do CTE para o presente estudo, recomenda-se sua
utilização para a estimativa da razão sexual dos filhotes. A utilização do CTE para o
cálculo da razão sexual (GEORGES et al. 1994, 2004) produziu resultados mais
consistentes que o emprego da temperatura média de incubação. A razão sexual
encontrada com a utilização da temperatura média de incubação (17% de fêmeas)
poderia inviabilizar a manutenção da população na região do rio Xingu (CONGDON et
35
al. 2000). Já a proporção de fêmeas obtida utilizando o CTE (47%) é mais
consistente com a manutenção de uma população viável e ecologicamente
equilibrada.
Nossos dados sugerem que em um regime de temperaturas diárias variáveis a
temperatura média de incubação não é um bom preditivo da razão sexual (GEORGES
et al. 1994, VALENZUELA et al. 1997, VALENZUELA 2001). A taxa de desenvolvimento
embrionário é diretamente relacionada à temperatura da incubação (BULL & VOGT
1981, MROSOVSKY et al. 1984, MARCOVALDI et al. 1997), mas as oscilações diárias de
temperatura, são melhor quantificadas com o uso do CTE, pois neste caso é o
tempo desprendido acima e abaixo da temperatura pivotal que definirá o sexo do
embrião (GEORGES et al. 1994).
Para Peltocephalus dumerilianus a razão sexual de adultos é desviada para fêmeas
(LA OSSA & VOGT 2010, PEZZUTI et al. 2008). Já P. unifilis, no rio Guaporé (RO),
mostrou uma razão sexual desviada para machos (TÉRAN & MÜLHEN 2003). Para as
tartarugas marinhas do litoral brasileiro é freqüente a razão sexual desviada para
fêmeas (MARCOVALDI et al. 1997; MROSOVSKY et al. 1999; GODFREY et al. 1999;
GODLEY et al. 2001) e apenas localmente tem-se áreas com uma produção maior de
machos (BAPTISTOTTE et al. 1999). Estes fatos demonstram que pode não haver um
padrão pré-estabelecido para a razão sexual em quelônios, ou indicam problemas
relacionados à
padronização
da
amostragem.
Além disso,
diferenças no
comportamento das espécies podem levar a uma interpretação errônea dos dados.
GIBBONS (1970) afirma que a determinação da razão sexual funcional deve ser
baseada nos indivíduos que atingiram a maturidade sexual e portanto, a proporção
de machos e fêmeas na eclosão deve ser utilizada com cautela nos programas de
manejo e conservação, uma vez que essas espécies apresentam alta taxa de
36
mortalidade dos filhotes (FERREIRA JÚNIOR 2009) e a razão sexual no nascimento
pode ser alterada ao longo dos anos.
Para a utilização da duração de incubação como ferramenta preditiva da razão
sexual, de espécies de ciclo de vida longo como P. unifilis, deve-se avaliar por
longos períodos (MARCOVALDI et al. 1997), já que mudanças sazonais podem causar
diferenças nas características de cada temporada reprodutiva (MROSOVSKY 1984).
Este fato deve ser considerado, principalmente em ninhos rasos como P. unifilis,
onde a taxa de desenvolvimento embrionário é muito sensível às variações
ambientais (GEORGES et al. 1989). Os dados obtidos no presente trabalho
demonstram que existe uma relação positiva entre o CTE, a duração de incubação e
a razão sexual, indicando que tanto a temperatura quanto a duração de incubação
possuem potencial preditivo em relação à porcentagem de machos e fêmeas que
nascem no rio Xingu, bem como para as demais espécies de quelônios continentais.
O CTE apresenta maior correlação (R²) que a temperatura média do segundo terço
de incubação, sugerindo que sua utilização é mais adequada para o cálculo da
razão sexual. Nossa hipótese de que as variações diárias de temperatura é que são
responsáveis pela produção de machos e fêmeas, como proposto por GEORGES et
al. (2004), foi aceita, assim recomendamos a utilização do CTE em estudos futuros.
É importante ressaltar que os dados obtidos no presente trabalho são referentes à
temporada reprodutiva de 2010, e apenas um ano não é suficiente para descrever a
reprodução de uma espécie de vida longa, sendo necessário o monitoramento
contínuo na região. Somente assim será possível estabelecer a dinâmica
populacional de P. unifilis no rio Xingu e a influência da razão sexual dos filhotes na
estrutura populacional. Nossos dados devem ser utilizados como ponto de partida
para estudos sobre a ecologia e biologia reprodutiva da espécie, a fim de determinar
37
as melhores técnicas de manejo e conservação. Além disso, é importante que se
amplie os esforços de pesquisa e se inicie um programa de captura e recaptura de
juvenis e adultos (PEZZUTI et al. 1998, TERÁN & MÜLHEN 2003, LA OSSA & VOGT 2010)
para caracterização efetiva da população protegida e manejada pelo Projeto
Tarekaja’a Xingu.
AGRADECIMENTOS
Ao CNPq, pela concessão da bolsa de mestrado. A Universidade Vila Velha, pela
estrutura disponibilizada. A Petrobras Ambiental e a EMBRAPA/CENARGEN, pelo
apoio logístico e estrutural. Ao RAN/ICMBio e ao Projeto Tarekaja’a Xingu e todos os
seus agentes pelo auxílio na coleta de dados. Agradecemos à comunidade indígena
Kamayurá da aldeia Morená, pelo apoio no desenvolvimento das atividades de
campo. Ao Laboratório de Histopatologia da UVV. A toda equipe de pesquisa do
Laboratório de Ecologia Terrestre e Aquática da UVV, pelo apoio e colaboração.
REFERÊNCIAS
BAPTISTOTTE, C.; J.T. SCALFONI & N. MROSOVSKY. 1999. Male-producing thermal
ecology of a southern loggerhead turtle nesting beach in Brazil: implications for
conservation. Animal Conservation 2: 9-13.
BOOTH, D. T. & C. FREEMAN. 2006. Sand and nest temperatures and an estimate of
hatchling sex ratio from the Heron Island green turtle (Chelonia mydas) rookery,
Southern Great Barrier Reef. Coral Reefs 25 629–633.
BULL, J.J. 1980. Sex determination in reptiles. Quarterly Review of Biology 53(1):
3-20.
38
BULL, J.J. & R.C. VOGT. 1981. Temperature-sensitive periods of sex determination in
Emydid turtles. Journal of Experimental Zoology 218:435–440.
BULL, J.J.; R.C. VOGT, R.C. & C.J. MCCOY. 1982. Sex determining temperatures in
turtles: a geographic comparison. Evolution 36(2): 326-332.
BULL, J.J.; J.M. LEGLER & R.C VOGT. 1985. Non-temperature dependent sex
determination in two suborders of turtles. Copeia 3: 784-786.
CONGDON, J.D.; R.D. NAGLE; O.M. KINNEY; M. OSENTOSKI; H.W. AVERY; C.R.S.
VAN
LOBEN, & D.W. TINCKLE. 2000. Nesting ecology and embryo mortality: implications for
hatchling success and demography of Blanding’s turtles (Emydoidea Blandingii).
Chelonian Conservation and Biology 3 (4): 569-579.
EWERT, M.A. & C.E NELSON. 1991. Sex determination in turtles: diverse patterns and
some possible adaptive values. Copeia 1: 50-68.
EWERT, M.A.; D.R. JACKSON & C.E. NELSON. 1994. Patterns of temperature-dependent
sex determination in turtles. Journal of Experimental Zoology 270(1): 3-15.
EWERT, M.A.; C.R. ETCHEBERGER & C.E. NELSON. 2004. Turtle sex-determination
modes and TSD patterns, and some TSD patterns correlates, p. 21-32. In: N.
Valenzuela & V.A. Lance (Eds). Temperature-dependent sex determination in
vertebrates. Washington, Smithsonian Books, 194p.
FADINI, L. S.; A.G. SILVA & FERREIRA JÚNIOR, P. D. 2011 . Sedimentary
characteristics and their effects on hatching success and incubation duration of
Caretta caretta (Testudines: Cheloniidae) in Espirito Santo, Brazil. Zoologia 28: 312320.
39
FERREIRA JÚNIOR, P.D. 2009. Efeitos de fatores ambientais na reprodução de
tartarugas. Acta Amazonica 39(2): 319-334.
Ferreira Júnior, P.D.; R.A.M. Balestra; J.R. Moreira; F.O. Freitas; A. P. G. Lustosa;
R.F. Jorge; A.J. M.Rosa; A.A. Sampaio & A.S. Gomes. 2011. Nesting of Phrynops
geoffroanus (Testudines: Chelidae) on sandy beaches along the Upper Xingu River,
Brazil. Zoologia 28: 571-576.
GEORGES, A. 1989. Female turtle from hot nests: Is it duration of incubation or
proportion of development that matters? Oecologia 81: 323-328.
GEORGES, A.; C. LIMPUS & R. STOUTJESDIJK. 1994. Hatchling sex in the marine turtle
Caretta caretta is determined by proportion of development at a temperature, not
daily duration of exposure. Journal of Experimental Zoology 270 (3): 432-444.
GEORGES, A.; S. DOODY; K. BEGGS & J. YOUNG. 2004. Thermal models of TDS under
laboratory and field conditions. In: Valenzuela, N.; Lance, V.A. TemperatureDependent Sex Determination in Vertebrates (Eds). Smithsonian Books,
Washington. p. 79-89.
GODFREY, M.H.; A.F; D’AMATO; M.Â. MARCOVALDI & N. MROSOVSKY. 1999. Pivotal
temperature and predicted sex ratios for hatchling hawksbill turtles from Brazil.
Canadian Journal of Zoology 74(9): 1465-1473.
GODLEY, B.J.; A.C. BRODERICK & N. MROSOVSKY. 2001. Estimating hatchling sex ratios
of loggerhead turtles in Cyprus from incubation durations. Marine Ecology Progress
Series 210:195-201.
40
IBAMA – INSTITUTO BRASILEIRO
DO
MEIO AMBIENTE
E DOS
RECURSOS NATURAIS
RENOVÁVEIS. 1989. Projeto Quelônios da Amazônia, 10 anos. Brasília, IBAMA,
119p.
LUBIANA, A. & P.D. FERREIRA JÚNIOR. 2009. Pivotal temperature and sexual
dimorphism of Podocnemis expansa hatchlings (Testudines: Podocnemididae) from
Bananal Island, Brazil. Zoologia 26 (3): 527-533.
MARCOVALDI, M.A. & LAURENT, A. 1996. A six season study of marine turtle nesting at
Praia do Forte, Bahia, Brazil, with implications for conservation and management.
Chelonian Conservation Biology 2:55–59.
NARO-MACIEL, E.; N. MROSOVSKY & M.Â. MARCOVALDI. 1999. Thermal profiles of sea
turtle hatcheries and nesting areas at Praia do Forte, Brazil. Chelonian
Conservation and Biology 3(3): 407-413.
PIEAU, C. & M. DORIZZI. 1981. Determination of temperature sensitive stage for sexual
differentiation of the gonads in embryos of the turtle, Emys orbicularis. Journal of
Morphology 170: 373-382.
PRITCHARD, P.C.H. & P. TREBBAU. 1984. The turtles of Venezuela. Oxford, Society
for the Study of Amphibians and Reptiles, 403p.
SOUZA, R.R. & R.C. VOGT 1994. Incubation temperature influences sex and hatchling
size in the neotropical turtle Podocnemis unifilis. Journal of Herpetology 28(4): 453464.
VALENZUELA, N.; R. BOTERO; & E. MARTÍNEZ. 1997. Field study of sex determination in
Podocnemis expansa from Colombian Amazonia. Herpetologica 53(3):390-395.
41
VALENZUELA, N. 2001.
Constant, shift, and natural temperatures effects on sex
determination in Podocnemis expansa turtles. Ecology 82: 30 10-3024.
VALENZUELA N. & V.A. LANCE. 2004. Temperature dependent sex determination in
vertebrates. Washington D.C, Smithsonian Books.
VOGT, R.C. 2008. Tartarugas da Amazônia. Lima, Peru, Gráfica Biblos, 104p.
42
3.2 CAPÍTULO II
INFLUÊNCIA DO LOCAL DA DESOVA
NO DESENVOLVIMENTO EMBRIONÁRIO DE PODOCNEMIS
UNIFILIS (T ROSCHEL 1848) (TESTUDINES: PODOCNEMIDIDAE) NO PARQUE INDÍGENA DO
XINGU, MATO GROSSO, BRASIL.
Aline Silva Gomes*¹, José Roberto Moreira², Fábio de Oliveira Freitas², Rafael
Antônio Machado Balestra³ & Paulo Dias Ferreira Júnior¹.
1 Programa de Pós-graduação em Ecologia de Ecossistemas, Universidade Vila
Velha, Rua Comissário José Dantas de Melo, 21, Boa Vista, 29102-770, Vila
Velha, ES, Brasil.
2. Embrapa Recursos Genéticos e Biotecnologia, Parque Estação Biológica, Av.
W5 Norte, Caixa Postal 02372, 70770-917, Brasília, DF, Brasil.
3. Centro Nacional de Pesquisa e Conservação de Répteis e Anfíbios, Rua 229,
95, Setor Leste Universitário, 74.605-090, Goiânia, GO, Brasil.
4. [email protected], autor correspondente
RESUMO
As características do meio físico são determinantes no desenvolvimento embrionário
de Podocnemis unifilis, determinando o sexo dos embriões e influenciando o
momento da eclosão. O presente trabalho avalia como o local da desova interfere no
desenvolvimento embrionário de P. unifilis no rio Xingu e avalia o manejo dos ninhos
realizado no local. Foi monitorada a temperatura de 120 ninhos durante todo o
período de incubação e amostras de sedimento foram coletadas para análise
granulométrica. Foram comparados o sucesso de eclosão, a razão sexual e a
duração de incubação de ninhos oriundos de áreas arenosas, com alto teor de lama
e no berçário com ninhos transferidos. O local da desova não influenciou a
temperatura ou o desenvolvimento embrionário de P. unifilis, demonstrando que o
manejo realizado é adequado, não havendo diferença significativa na razão sexual
ou no sucesso de eclosão dos ninhos naturais e transferidos.
Palavras-chave: determinação sexual, incubação, manejo, tracajá, transferência,
razão sexual.
43
INTRODUÇÃO
As características do ambiente de
nidificação são fundamentais para
o
desenvolvimento e determinação sexual dos filhotes de Podocnemis unifilis
(TROSCHEL 1848) (MALVASIO et al. 2002). A determinação sexual desta espécie se dá
de acordo com a temperatura a qual os ovos são submetidos durante a
embriogênese, sendo o conjunto das características ambientais o responsável pelo
estabelecimento desta temperatura (SOUZA & VOGT 1994, MALVASIO et al. 2002). Para
P. unifilis, temperaturas acima de 32 °C tendem a gerar prioritariamente fêmeas,
enquanto temperaturas mais baixas são responsáveis pela produção de machos
(SOUZA & VOGT 1994). Dessa forma, o momento da desova, as características do
sedimento (tamanho do grão, albedo e composição mineralógica) e as condições de
exposição e sombreamento são fatores determinantes da temperatura no interior
dos ninhos e assim, exercem forte influência no sucesso de eclosão e no sexo dos
filhotes (FERREIRA JÚNIOR 2009, FERREIRA JÚNIOR & CASTRO 2010, FADINI et al. 2011).
A escolha do ambiente de nidificação é um fator crucial para o desenvolvimento
embrionário, sendo a fêmea responsável por selecionar locais que sejam adequadas
à postura. Estes locais devem ser altos o suficiente para evitar o alagamento dos
ninhos e propiciar uma temperatura adequada ao desenvolvimento dos embriões,
garantindo a manutenção de uma razão sexual compatível com a população local
(MORREALE et al. 1982, VALENZUELA et al. 1997).
No desenvolvimento embrionário de diversas espécies a temperatura pivotal é a
responsável pela produção de machos e fêmeas em proporções iguais (50%), e é
considerada um fator chave para a ecologia reprodutiva desses animais (PIEAU 1976,
BULL 1980, YNTEMA & MROSOVSKY 1982). Machos e fêmeas desenvolvem-se em
velocidades e temperaturas diferentes, de modo que temperaturas mais elevadas
44
promovem um desenvolvimento mais rápido e estimulam a produção de fêmeas.
Assim fêmeas normalmente apresentam uma duração de incubação mais curta do
que os machos (PIEAU 1976, YNTEMA & MROSOVSKY 1982). GEORGES et al. (2004)
propuseram que, em regime variável de temperatura, fêmeas serão geradas quando
mais da metade do desenvolvimento embrionário ocorrer acima da temperatura
pivotal e machos serão produzidos quando mais da metade do desenvolvimento
embrionário ocorrer abaixo da temperatura pivotal.
Dessa forma pode-se relacionar a temperatura de incubação à taxa de
desenvolvimento embrionário e, conseqüentemente, à duração de incubação dos
ovos (MARCOVALDI et al. 1997). Para tartarugas marinhas a duração de incubação é
utilizada na predição da razão sexual, e baseia-se no conhecimento da temperatura
de incubação dos ovos e das faixas responsáveis pela produção de cada sexo
(MARCOVALDI et al. 1997; MROSOVSKY et al. 1999; GODFREY et al. 1999; BRODERICK et
al. 2000; GODLEY et al. 2001). Entretanto não existem estudos que confirmem a
viabilidade da utilização da duração de incubação na estimativa da razão sexual
para espécies de quelônios continentais.
GOMES
E
FERREIRA JUNIOR (2011) demonstraram que a transferência de ninhos
sujeitos ao alagamento, ou à predação pode ser uma alternativa viável para o
manejo de espécies que se encontram em situação vulnerável, podendo esta ação
representar um aumento significativo no sucesso de eclosão com reflexos no
recrutamento de juvenis na população. Porém é necessário que se observe as
condições de transferência, pois alterações no ambiente de nidificação podem
repercutir na duração da incubação e na determinação sexual dos filhotes (FERREIRA
JÚNIOR & CASTRO 2003). Assim, é imprescindível que se analise os parâmetros
45
fisiográficos do sítio de nidificação, bem como a influência que estes parâmetros
exercem no desenvolvimento embrionário das tartarugas.
A temperatura pivotal é fundamental na ecologia destes animais por se tratar de uma
característica que pode definir a existência de uma determinada espécie em uma
região (BULL et al. 1982). Espécies com grande distribuição geográfica tendem a
possuir variações em torno de sua temperatura pivotal a fim de compensar a
variações climáticas e viabilizar a ocupação da região (EWERT & NELSON 1991,
EWERT et al.1994).
Conhecer a maneira como o ambiente interfere no desenvolvimento dos embriões é
fundamental para a definição de áreas apropriadas para a conservação e a
execução do manejo sustentável da espécie (GOMES & FERREIRA JÚNIOR 2011). O
presente trabalho
tem como
objetivos identificar
como
as características
sedimentológicas do local da desova afetam a temperatura no interior dos ninhos, e
as conseqüências no sucesso de eclosão e na razão sexual de P. unifilis no rio
Xingu, analisando a eficiência das técnicas de manejo executadas atualmente na
região. Para avaliar este último objetivo será analisada a influência da transferência
de ninhos no sucesso de eclosão e na razão sexual dos filhotes gerados.
MATERIAIS E MÉTODOS
Durante a temporada reprodutiva de 2010, foram monitoradas as temperaturas de
incubação de 120 ninhos distribuídos em duas das nove praias localizadas na região
norte do Parque Indígena do Xingu (Figura 1). As praias estão situadas no território
da aldeia Kamayurá-Morená e são monitoradas pelo Projeto Tarekaja’a Xingu, que
46
busca a conservação e executa o manejo de P. unifilis na região. Esta iniciativa tem
o apoio da comunidade indígena para a realização do trabalho.
Figura 1. Localização do Parque Indígena do Xingu e da área de estudo. O território da
aldeia Kamayurá-Morená engloba as nove praias estudadas no rio Xingu (Ferreira Júnior et
al 2011). Foi monitorada a temperatura de incubação dos ninhos das praias 1 (Morená) e 3
(Korone)
Para avaliar a distribuição espacial dos ninhos foi realizado o mapeamento das
praias com o auxílio de bússola e trena na escala de 1:1000 (FERREIRA JÚNIOR &
47
CASTRO 2003). Nesta escala foi possível plotar os ninhos e as principais feições
geomorfológicas das praias.
A temperatura no interior dos ninhos foi monitorada através de termógrafos digitais
do tipo i-button (Maxim DSG1921G) inseridos no interior da câmara de ovos. Os
termógrafos mediram e registraram a temperatura do ninho a cada hora durante toda
a incubação. Foram coletadas amostras de sedimento do interior dos ninhos,
destinados à análise granulométrica segundo o protocolo de FOLK (1974).
Foram coletadas as informações sobre o sucesso de eclosão (razão entre filhotes
vivos e ovos viáveis), a duração de incubação (número de dias transcorridos desde
a desova até a eclosão) e a razão sexual (tomada como a porcentagem de fêmeas
que cada ninho produziu). O nível do rio foi monitorado durante todo o período de
incubação, caso houvesse necessidade de transferência por risco de alagamento,
considerou-se o sucesso de eclosão igual a zero. Os ninhos alagados foram
quantificados e analisados separadamente.
As transferências de ninhos, no rio Xingu, são realizadas como alternativa para
aumentar o número de eclosões por temporada e são motivadas, principalmente
pela alta predação dos ninhos por canídeos, principalmente Cerdocyon thous
(FERREIRA JÚNIOR et al. 2011). A predação é a principal causa de perda de ovos e
filhotes na região, por esse motivo, todos os ninhos das praias 8 (Tepotsiy) e 9
(Janypawu) foram transferidos logo após a desova para um berçário localizado na
praia 3 (Korone). Os ovos foram transportados em caixas de isopor preenchidas com
areia oriunda do interior do próprio ninho, a fim de manter a temperatura e evitar o
deslocamento dos ovos e, conseqüentemente, dos embriões (MARCELLINI & DAVIS
1982, MALVASIO et al. 2002). Todos os ninhos, transferidos ou não, foram protegidos
com tela de metal e identificados com lacre numerado. Nas praias com maior
48
ocorrência de canídeos as telas foram reforçadas com ganchos de ferro enterrados
na areia.
Para a razão sexual dos ninhos foi utilizada a temperatura pivotal de 31,8 °C,
calculada por GOMES & FERREIRA JÚNIOR (ver capítulo 1) que realizaram a incubação
dos ovos desta população em temperatura constante. No cálculo da razão sexual foi
empregado o CTE do segundo terço de incubação, que é mais indicado que a
temperatura média em regimes de temperatura variável, como no caso de P. unifilis.
O CTE é indicado nos trabalhos que envolvem um regime variável de temperatura
(GEORGES 1989) em que as variações diárias são elevadas, como é o caso dos
ninhos de P. unifilis (GEORGES et al. 1994, 2004). A taxa de desenvolvimento
embrionário, necessária ao estabelecimento do CTE, foi calculada a partir da relação
entre a duração de incubação em temperaturas constantes (GOMES & FERREIRA
JÚNIOR capítulo 1) e expressa por:
Duração da incubação (dias) = 182 – (3,727 Temperatura de incubação)
A fração diária do desenvolvimento embrionário, portanto, é dada por:
1/ (-3,827+185,63 T)
A soma de todas as frações durante o segundo terço da incubação foi multiplicada
por 0,34 (correspondente à fração do desenvolvimento embrionário no qual o sexo é
determinado irreversivelmente) obtendo-se assim o CTE do ninho.
A análise granulométrica foi realizada por peneiramento e a porcentagem de cada
classe de tamanho foi transformada para o seu arcoseno buscando a normalidade
dos dados. O sucesso de eclosão e a razão sexual também foram transformados
para seu arcoseno. Foi aplicado o teste de Shapiro-Will para avaliar a normalidade
da distribuição dos dados.
49
Foram comparados ninhos transferidos e ninhos in situ, divididos em ninhos
localizados em sedimentos francamente arenosos e sedimentos silto-arenosos,
através de Análise de Variância para comparação entre o sucesso de eclosão, a
duração de incubação, o CTE e a razão sexual (calculada com base no CTE). Para
avaliar se a duração da incubação, o sucesso da eclosão e o tamanho dos
sedimentos variaram entre os ninhos foi aplicado o teste não paramétrico de
Kruskall-Wallis (quando dados não apresentaram distribuição normal mesmo após
transformação), com significância de 5%. A influência do tamanho do sedimento na
temperatura de incubação (CTE e temperatura média no segundo terço) e no
sucesso da eclosão foi testada através da regressão linear múltipla. Devido à alta
colinearidade algumas frações granulométricas foram removidas dos cálculos para
que não inflacionassem os resultados. Os cálculos foram realizados com o auxílio do
programa Systat 3.5, e a classificação dos sedimentos foi realizada com o programa
Gradistat 4.0. Todas as análises estão de acordo com ZAR (2010).
RESULTADOS
Na temporada reprodutiva de 2010 ocorreram 1458 desovas de P. unifilis nas nove
praias monitoradas. Foram transferidos 60 ninhos das praias 8 e 9 para o berçário
da praia 3. No total, 524 ninhos foram encontrados predados e não puderam ser
protegidos. Dos ninhos protegidos, 227 sofreram predação, principalmente por
Cerdocyon thous (Linnaeus, 1706), e secundariamente por gaviões (Polyborus
plancus, Miller 1777) urubus (Coragyps atratus, Bechstein 1793) e jacarés (Caiman
crocodilos, Linnaeus, 1758). Vinte e nove ninhos foram alagados ou necessitaram
ser transferidos por risco de afogamento dos embriões. Também foram registrados
31 ninhos de Phrynops geoffroanus, que compartilham com P. unifilis as áreas de
50
desova (FERREIRA JÚNIOR et al. 2011). Os ninhos se distribuem por toda a praia, mas
pode-se perceber uma maior concentração nas áreas planas e baixas nas margens
do canal e nas proximidades com a vegetação na porção externa do banco arenoso
(Figura 2).
A análise granulométrica indicou diferenças no tamanho dos sedimentos,
caracterizando um ambiente heterogêneo ao longo das praias. Os sedimentos
podem ser classificados primordialmente como arenosos, areno-lamosos e arenocascalhosos (Figura 3).
Não houve diferença significativa para nenhuma das variáveis analisadas, ou seja, a
localização e a transferência dos ninhos das praias 8 e 9 para a praia 3 não
interferem no CTE (F= 1,011; p= 0,368), no sucesso de eclosão (H= 2,393; gl= 2; p=
0,302), na razão sexual (F= 0,663; p= 0,518) ou na duração de incubação (F= 2,330;
p= 0,103) dos ninhos.
Apenas a fração lama exerceu influencia na temperatura média no segundo terço de
incubação (p= 0,006), indicando que as variáveis ambientais influenciam a
temperatura média no segundo terço de incubação (F= 2,941; p= 0,034). O sucesso
de eclosão também não foi afetado pelas variáveis ambientais (r²= 0,25; F= 1,641;
p= 0,186).
A data da desova e a altura dos ninhos são elementos que podem auxiliar na
predição do alagamento das praias e afogamento dos embriões. Todos os ninhos
alagados foram desovados no período entre 29 de julho até o final de agosto. A
altura destes ninhos em relação ao rio no momento da desova para variou entre 22
cm e 100 cm nos ninhos alagados.
51
Figura 2. Distribuição dos ninhos de Podocnemis unifilis e Phrynops geoffroanus na Praia 1
(Morená) no Parque Indígena do Xingu na temporada reprodutiva de 2010.
52
Figura 2. Classificação do sedimento de acordo com Folk (1974).
DISCUSSÃO
Apesar das variações no tamanho do sedimento, na posição e altura dos ninhos em
relação ao nível do rio, excluindo-se os ninhos sujeitos ao alagamento, o local da
desova não apresentou grande importância para a reprodução de Podocnemis
unifilis no rio Xingu e as características do meio físico não interferiram no
desenvolvimento embrionário dos filhotes. As diferenças no tamanho do sedimento
não foram suficientes para alterar o ambiente termal dos ninhos e influenciar o
desenvolvimento embrionário. A pequena influência exercida pela fração “lama” não
foi suficiente para afetar a temperatura. Desta forma, a razão sexual e o sucesso de
eclosão não variaram entre os ninhos localizados em sedimentos arenosos, arenolamosos e areno-cascalhosos.
53
Esta relativa homogeneidade termal permitiu que a transferência dos ninhos não
afetasse a razão sexual e o sucesso de eclosão. Em projetos de manejo que
envolvem a transferência dos ninhos, estes são os principais objetivos a serem
alcançados (MARCOVALDI & MARCOVALDI 1997, IBAMA 1989). Garantir que os locais
de origem e de incubação dos ovos sejam semelhantes e não alterem a viabilidade
de desenvolvimento, ou a razão sexual dos filhotes gerados é essencial para a
manutenção das características populacionais da espécie na região (NARO-MACIEL et
al. 1999, MILTON et al. 1999, GOMES & FERREIRA JÚNIOR 2011). Os resultados obtidos
no presente trabalho demonstram que os locais de estudo não variam e não
influenciam a reprodução de P. unifilis, o que amplia as possibilidades de manejo na
região. As transferências de ninhos demonstram-se como alternativa viável de
manejo dos ovos e garantem o aumento do número de eclosões por temporada
reprodutiva (MARCOVALDI & MARCOVALDI 1997, IBAMA 1989), no entanto devem-se
realizar as transferências do modo seguro (MARCELLINI & DAVIS 1982, MALVASIO et al.
2002) para a promoção do manejo sustentável (GOMES & FERREIRA JÚNIOR 2011).
A transferência se mostrou viável e o sucesso de eclosão semelhante aos ninhos
naturais indica que ela é realizada de forma adequada. A proteção, entretanto não
se mostrou muito eficiente, pois cerca de 25 % dos ninhos protegidos com tela ainda
foram predados, demonstrando que a proteção dos ninhos é um fator que necessita
ser melhorado para ampliação das eclosões e da conservação de P. unifilis no
Xingu.
A distribuição dos ninhos de Podocnemis unifilis no PIX segue o padrão observado
em outras áreas como nas praias arenosas do rio Javaés, no estado do Tocantins
(FERREIRA JÚNIOR & CASTRO 2006, 2010). Apesar de não ocorrerem desovas
regulares de P. expansa nesta parte do rio Xingu (MOREIRA et al. no prelo), P. unifilis
54
seleciona principalmente áreas baixas, nas proximidades do canal e as partes
próximas à vegetação. A desova nas áreas mais baixas foi apontada como
estratégia para evitar a sobreposição com P. expansa (LIMA et al. 2009), que desova
mais tardiamente e poderia destruir os ninhos rasos de P. unifilis. No PIX esta
possível destruição dos ninhos não é um problema e a manutenção da desova em
áreas mais susceptíveis ao alagamento pode representar outra estratégia associada
a uma menor exposição das fêmeas adultas à predação durante a desova (e.g.:
predação por onça) (SALERA JÚNIOR et al. 2009), ou necessidades específicas
voltadas às características dos ovos (PACKARD et al. 1981).
O fato é que a escolha das áreas próximas ao leito do rio aumenta a predisposição
ao alagamento e afogamento de filhotes e embriões durante à subida do rio
(FERREIRA JÚNIOR & CASTRO 2003). Por isso o manejo de P. unifilis é indispensável
para conservação da espécie nas áreas de ocorrência.
Com a adesão da comunidade da aldeia Kamayurá-Morená ao projeto, a coleta de
ovos e de fêmeas durante a desova cessam por completo e para P. unifilis, no PIX, a
predação dos ninhos dos ninhos por canídeos e o alagamento são as principais
causas de perda de ninhos.
Em 2010 a utilização das telas se mostrou eficiente contra a predação. Para avaliar,
entretanto, a influência da altura do ninho em relação e o risco d alagamento é
necessário o monitoramento de médio prazo para que se possa avaliar a dinâmica
dos períodos de cheia e vazante. O momento em que o rio inicia o alagamento das
praias varia ao longo dos anos com reflexos distintos no sucesso da eclosão. Em
uma mesma praia o percentual de alagamento dos ninhos varia ao longo dos anos e
das espécies (FERREIRA JÚNIOR & CASTRO 2010) indicando a importância do
55
conhecimento da dinâmica fluvial no estabelecimento do manejo (GOMES & FERREIRA
JÚNIOR 2011).
No Parque Indígena do Xingu ninhos acima de 100 cm, independente do momento
da desova completaram a incubação. Para os ninhos situados abaixo desta altura o
momento da desova influenciou o risco de alagamento.
Estabelecer, portanto, o momento de ascensão do nível do rio, a partir de uma série
histórica, é essencial para indicar, no momento da desova, os ninhos mais
susceptíveis ao alagamento e que, por isso, devem ser transferidos.
É necessário quantificar a porcentagem de ninhos perdidos em outras temporadas
reprodutivas e verificar quais as praias e trechos de praia mais susceptíveis ao
alagamento. Assim será possível realizar o manejo específico, de acordo com a
data e local da desova. Estudos aplicados possibilitariam a aplicação de estratégias
de manejo sustentável junto à comunidade indígena que utiliza os ovos e filhotes
como recurso alimentar em sua dieta (Pezzutti et al. 2010).
Apesar de alcançarmos os objetivos propostos, mais estudos na região são
necessários para efetivar os dados obtidos. Por se tratar de uma espécie de longa
vida, dados de uma única temporada são insuficientes para avaliar dinâmica
reprodutiva (MARCOVALDI et al. 1997, CONGDON et al. 2000, FERREIRA JÚNIOR 2009).
São necessários maiores esforços de amostragem para analisar a utilização das
praias ao longo os anos e, em especial, a influência da dinâmica fluvial no
alagamento dos ninhos. Assim será possível definir com exatidão o status de
conservação da população de P. unifilis na região do Alto Xingu, a eficiência do
manejo e o possível uso sustentável pela comunidade Kamayurá-Morená.
56
AGRADECIMENTOS
Ao CNPq, pela concessão da bolsa de mestrado. A Universidade Vila Velha, pela
estrutura disponibilizada. A Petrobras Ambiental e a EMBRAPA/CENARGEN, pelo
apoio logístico e estrutural. Ao RAN/ICMBio e ao Projeto Tarekaja’a Xingu e todos os
seus agentes pelo auxílio na coleta de dados. Agradecemos à comunidade indígena
Kamayurá da aldeia Morená, pelo apoio no desenvolvimento das atividades de
campo. Ao Laboratório de Histopatologia da UVV. A toda equipe de pesquisa do
LETA, pelo apoio e colaboração.
REFERÊNCIAS
BRODERICK, A.C.; B.J. GODLEY; S. REECE & J.R DOWNIE. 2000. Incubation periods and
sex ratios of green turtles: highly female biased hatchling production in the eastern
Mediterranean. Marine Ecology Progress Series 202: 273-281.
BULL, J.J. 1980. Sex determination in reptiles. Quarterly Review of Biology 53(1):
3-20.
BULL, J.J.; R.C. VOGT, R.C. & C.J. MCCOY. 1982. Sex determining temperatures in
turtles: a geographic comparison. Evolution 36(2): 326-332.
CONGDON, J.D.; R.D. NAGLE; O.M. KINNEY; M. OSENTOSKI; H.W. AVERY; C.R.S.
VAN
LOBEN, & D.W. TINCKLE. 2000. Nesting ecology and embryo mortality: implications for
hatchling success and demography of Blanding’s turtles (Emydoidea Blandingii).
Chelonian Conservation and Biology 3 (4): 569-579.
EWERT, M.A. & C.E NELSON. 1991. Sex determination in turtles: diverse patterns and
some possible adaptive values. Copeia 1: 50-68.
57
EWERT, M.A.; C.R. ETCHEBERGER & C.E. NELSON. 2004. Turtle sex-determination
modes and TSD patterns, and some TSD patterns correlates, p. 21-32. In: N.
Valenzuela & V.A. Lance (Eds). Temperature-dependent sex determination in
vertebrates. Washington, Smithsonian Books, 194p.
FADINI, L.S.; A.G. SILVA & FERREIRA JÚNIOR, P.D. 2011. Sedimentary characteristics
and their effects on hatching success and incubation duration of Caretta caretta
(Testudines: Cheloniidae) in Espirito Santo, Brazil. Zoologia 28: 312-320.
FERREIRA JÚNIOR, P.D. 2009. Efeitos de fatores ambientais na reprodução de
tartarugas. Acta Amazonica 39(2): 319-334.
FERREIRA JÚNIOR, P.D. & P.T.A CASTRO. 2003. Geological control of Podocnemis
expansa and Podocnemis unifilis nesting areas in Rio Javaés, Bananal Island, Brazil.
Acta Amazonica 33(3): 445-468.
FERREIRA JÚNIOR, P.D. & P.T.A. CASTRO. 2010. Nesting ecology of Podocnemis
expansa (Schweigger, 1812) and Podocnemis unifilis (Troschel, 1848) (Testudines,
Podocnemididae) in the Javaés River, Brazil. Brazilian Journal of Biology 70: 8594.
Ferreira Júnior, P.D.; R.A.M. Balestra; J.R. Moreira; F.O. Freitas; A. P. G. Lustosa;
R.F. Jorge; A.J. M.Rosa; A.A. Sampaio & A.S. Gomes. 2011. Nesting of Phrynops
geoffroanus (Testudines: Chelidae) on sandy beaches along the Upper Xingu River,
Brazil. Zoologia 28: 571-576.
FOLK, R.L. 1974. Petrology of Sedimentary Rocks. Austin, Hemphill Publication
Company, 182p.
58
GEORGES, A. 1989. Female turtle from hot nests: Is it duration of incubation or
proportion of development that matters? Oecologia 81: 323-328.
GEORGES, A.; C. LIMPUS & R. STOUTJESDIJK. 1994. Hatchling sex in the marine turtle
Caretta caretta is determined by proportion of development at a temperature, not
daily duration of exposure. Journal of Experimental Zoology 270 (3): 432-444.
GEORGES, A.; S. DOODY; K. BEGGS & J. YOUNG. 2004. Thermal models of TDS under
laboratory and field conditions. In: Valenzuela, N.; Lance, V.A. TemperatureDependent Sex Determination in Vertebrates (Eds). Smithsonian Books,
Washington. p. 79-89.
GODFREY, M.H.; A.F; D’AMATO; M.Â. MARCOVALDI & N. MROSOVSKY. 1999. Pivotal
temperature and predicted sex ratios for hatchling hawksbill turtles from Brazil.
Canadian Journal of Zoology 74(9): 1465-1473.
GODLEY, B.J.; A.C. BRODERICK & N. MROSOVSKY. 2001. Estimating hatchling sex ratios
of loggerhead turtles in Cyprus from incubation durations. Marine Ecology Progress
Series 210:195-201.
Gomes, A.S. & Ferreira Júnior, P.D.F. 2011. Management and relocation of nests of
Podocnemis expansa (Scheweigger, 1812) (Testudines, Podocnemididade) on the
Crixás-Açu River, Brazil. Brazilian Journal of Biology 71 (4): 975-982.
IBAMA – INSTITUTO BRASILEIRO
DO
MEIO AMBIENTE
E DOS
RECURSOS NATURAIS
RENOVÁVEIS. 1989. Projeto Quelônios da Amazônia, 10 anos. Brasília, IBAMA,
119p.
LIMA, J. P.; J.C.B. PEZZUTI; L.A.S. MONJELO; A.S. TEIXEIRA; A. KEMENES; D.F. SILVA &
G.H. REBELO. 2009. Seleção de locais de desova e sobrevivência de ninhos de
59
quelônios Podocnemis no baixo Rio Purus, Amazonas, Brasil. RECIA - Revista
Colombiana de Ciência Animal 1: 37-59.
MALVASIO, A.; A.M. SOUZA; P.D. FERREIRA JÚNIOR; E.S. REIS & F.A.A. SAMPAIO. 2002.
Temperatura de incubação dos ovos e granulometria dos sedimentos das covas
relacionadas à determinação sexual em Podocnemis expansa (Schweigger, 1812) e
P. unifilis (Troschel, 1848) (Testudines, Pelomedusidae). Publicações Avulsas do
Instituto Pau Brasil de História Natural 5(1): 11-25.
MARCOVALDI, M.Â.; M.H. GODFREY & N. MROSOVSKY. 1997. Estimating sex ratios of
loggerhead turtles in Brazil from pivotal incubation durations. Canadian Journal of
Zoology 75:755-770.
MILTON, S.L.; A.A. SCHULMAN & P.L. LUTZ. 1997. The effect of beach nourishment with
aragonite versus silicate sand on beach temperature and loggerhead sea turtle
nesting success. Journal of Coastal Research 13(3): 904-915.
MOREIRA, J. R; R.A.M. BALESTRA; F.O. FREITAS; A.P.G. LUSTOSA; A.J.M. ROSA; A.A.
SAMPAIO & P.D. FERREIRA JÚNIOR. 2011. Podocnemis expansa (Giant South American
River Turtle). First record of nesting in the Upper Xingu River. Herpetological
Review 42: 595-596.
MORREALE, S.J; G.J. RUIZ; J.R. SPOTILA & E.A. STANDORA. 1982. Temperaturedependent sex determination: current pratices threaten conservation of sea turtles.
Science 216(11):1245-1247.
MROSOVSKY, N., C. BAPTISTOTTE & M.H. GODFREY. 1999. Validation of incubation
duration as an index of the sex ratio of hatchling sea turtles. Canadian Journal of
Zoology 77(5): 831-835.
60
NARO-MACIEL, E.; N. MROSOVSKY & M.Â. MARCOVALDI. 1999. Thermal profiles of sea
turtle hatcheries and nesting areas at Praia do Forte, Brazil. Chelonian
Conservation and Biology 3(3): 407-413.
PACKARD, G.C.; M.J. PACKARD & T.J. BOARDMAN. 1981. Patterns and possible
significance of water exchange by flexible-shelled eggs of painted turtles (Chrysemys
picta). Physiological Zoology 54(1): 165-178.
PEZZUTI, J.C.B; J.P.LIMA; D.F. SILVA & A. BEGOSSI. 2010. Uses and taboos of turtles
and tortoises along Rio Negro, Amazon Basin. Journal of Ethnobiology 30(1): 153–
168.
SALERA JUNIOR, G.; T. C. G. PORTELINHA; & A. MALVASIO. 2009. Predação de fêmeas
adultas de Podocnemis expansa Schweigger (Testudines, Podocnemididae) por
Panthera onca Linnaeus (Carnivora, Felidae), no Estado do Tocantins. Biota
Neotropica 9: 01.
VALENZUELA, N.; R. BOTERO; & E. MARTÍNEZ. 1997. Field study of sex determination in
Podocnemis expansa from Colombian Amazonia. Herpetologica 53(3):390-395.
VOGT, R.C. 2008. Tartarugas da Amazônia. Lima, Peru, Gráfica Biblos, 104p.
61
4. CONSIDERAÇÕES FINAIS
O Parque Indígena do Xingu apresenta-se como uma área com grande potencial
para a realização de estudos de ecologia de quelônio, tanto para Podocnemis unifilis
quanto para Phrynops geophranus. Observamos uma grande densidade de desovas
e interações de P. unifilis com outros fatores ambientais, tornando este um local
capaz de fornecer informações para analises de dinâmica e estrutura populacionais,
ecologia reprodutiva, densidade e diversidade de espécies, além das interações
ecológicas, como predação e coexistência de espécies e sobreposição de habitats.
Por isso, encorajamos a realização de estudos avançados na região, a fim de
incrementar os conhecimentos entorno da fauna do parque e, principalmente das
espécies de tartarugas que ali vivem.
Observamos também que o manejo realizado pelo Projeto Tarekaja’a Xingu é
adequado e não gera alterações na população de P. unifilis, no entanto outras
possibilidades de manejo são possíveis. Percebemos a necessidade de ampliação
da observação dos ninhos sob risco de alagamento, e aumento da proteção dos
ninhos, e praias, sob risco de predação. A perda desses ninhos ainda representa um
grande percentual do total de filhotes que deveriam nascer em uma temporada,
assim para que a proteção seja efetiva, estes pontos devem ser analisados com
maior atenção e outras estratégias devem ser utilizadas, além das técnicas
utilizadas atualmente.
Sugerimos a ampliação do Projeto Tarekaja’a Xingu, iniciando trabalhos com
indivíduos juvenis e adultos, além da caracterização genética e morfológica dos
indivíduos. É importante, para um projeto de conservação, saber se existe fluxo
gênico com outras populações próximas, assim é possível determinar o status e a
viabilidade da população.
62
Por fim, indicamos a continuidade dos estudos realizados até o momento. Os dados
obtidos no presente trabalho demonstram que estudar e caracterizar a população de
P. unifilis no alto Xingu é possível, no entanto é indispensável a validação do CTE
como índice da razão sexual dos filhotes, estudos de médio e longo prazo devem
ser realizados para a confirmação dos dados obtidos.
Todas as sugestões realizadas nesta seção foram feitas levando em conta o bem
estar e a manutenção dos costumes e tradições da comunidade indígena KamauyráMorená, que centraliza o Projeto Tarekaja’a Xingu no Parque Indígena do Xingu.