Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
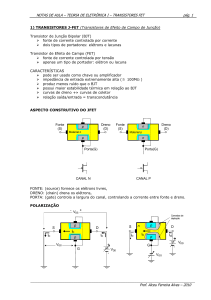
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
PUBVET, Publicações em Medicina Veterinária e Zootecnia.
Disponível em: <http://www.pubvet.com.br/texto.php?id=244>.
O manejo de pastagens e as relações fonte/dreno na planta forrageira
Lilian Elgalise Techio Pereira1 e Adenilson José Paiva1
1
Zootecnista Aluna de mestrado do Programa de Pós-Graduação em Ciência
Animal e Pastagens ESALQ/USP
Resumo
O crescimento de plantas, entendido como o acúmulo de matéria seca,
depende quase que inteiramente da fotossíntese, processo caracterizado pela
utilização da luz solar para fixar o gás carbônico da atmosfera e utilização na
síntese de uma série de moléculas orgânicas complexas que atuam nos
processos fisiológicos responsáveis pela manutenção de material vegetal.
Existe uma hierarquia na partição de assimilados produzidos pela fotossíntese
na fabricação dos diferentes compartimentos que compõem a biomassa
vegetal, que prioriza a expansão e manutenção de órgãos essenciais. O padrão
de translocação de assimilados a partir da fontes e os órgãos “alvo” ou drenos
dependem do estágio fisiológico da planta ou condições de ambiente e manejo
a qual está submetida. A importância destes processos reside no fato de que é
possível maximizar o crescimento vegetal, em relação aos órgãos de interesse,
tais como produção de sementes ou produção folhas no caso de pastagens,
quando
se
conhecem
os
princípios
que
regem
o
carregamento
e
descarregamento dos produtos da fotossíntese através do floema, pois
algumas práticas de manejo de certas culturas são uma interferência direta
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
nas relações fonte/dreno. Em pastagens sob condição de pastejo a desfolhação
é o fator de manejo que mais interfere na partição de assimilados, cujo efeito
no crescimento da planta durante o período de rebrotação é gradativo e
dependente da intensidade de remoção da área foliar e da decapitação dos
perfilhos. Essa interferência pode ser positiva ou negativa, conforme a
habilidade da planta em restabelecer uma nova partição de assimilados das
novas fontes para os novos drenos. Portanto conhecer a habilidade de planta
na partição e alocação de assimilados é de fundamental importância para se
adotar manejos em que se tenha o maior rendimento produtivo (exemplo kg
de
carne
produzidos
por
área),
sem
comprometer
a
persistência
e
produtividade da planta.
The pasture management and relations source/sink in forage crops
Abstract – Plant growth, taken as dry matter accumulation, depends almost
entirely on photosynthesis, process characterized by the use of the sunlight to
sequestrate carbon gas from the atmosphere and use it in the synthesis of a
series of complex organic molecules that act in the physiologic processes
responsible for the maintenance of vegetal material. There is a hierarchy in
partition of assimilates produced from photosynthesis in the production of the
different compartments that compose the vegetal biomass, which prioritizes
the expansion and maintenance of essential organs. The translocation pattern
of assimilates from sources and target organs or sink depend on the
physiologic stage of the plant or atmosphere conditions and management
which it is submitted to. The importance of these processes lies in the fact that
it is possible to maximize the vegetal growth, regarding organs of interest,
such as seeds production or leaves production in pastures, when it is known
the principles that govern the shipment and unloading of photosynthesis
products through the phloem, because some crops management practices are
a direct interference in the relationships source/sink.
In pastures under
grazing condition, the defoliation is the management factor that interferes the
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
most in the partition of assimilates, whose effect in the plant growth process
during the regrowth period is gradual and dependent on the intensity of
removal of the foliar area and of the decapitation of the tiller. This interference
can be positive or negative, according to the ability of the plant to reestablish a
new partition of assimilates of new sources to new sinks. Therefore, knowing
the plant ability in the partition and allocation of assimilates is of fundamental
importance to adopt management strategies to obtain optimal productivity
(example meat kg produced in the area), without affecting the persistence and
productivity of the plant.
1. Introdução
A mais importante característica das plantas é sua habilidade em
utilizar a luz solar para fixar o gás carbônico da atmosfera, sintetizando uma
série de moléculas orgânicas complexas. Esse processo fisiológico, que ocorre
nos tecidos clorofilados, é o principal responsável pela entrada de energia livre
na biosfera (Floss, 2006).
O crescimento das culturas, entendido como o acúmulo de matéria
seca, depende quase que inteiramente da fotossíntese. Segundo Taiz & Zieger
(2004), cerca de 90% da massa seca vegetal são originadas diretamente da
assimilação fotossintética de carbono através da fotossíntese.
A utilização dos compostos produzidos na fotossíntese segue uma
dinâmica que prioriza a expansão e manutenção de órgãos essenciais, sendo
que o padrão de translocação de assimilados e os órgãos “alvo” dependem do
estágio fisiológico da planta ou condições de ambiente e manejo a qual está
submetida. Segundo Moraes et al. (1995) existe uma hierarquia na partição de
assimilados para a fabricação dos diferentes compartimentos que compõem a
biomassa vegetal (folhas, hastes, ramificações e raízes). Na ausência de
limitações dos recursos temperatura, água, N e luz, a prioridade de alocação
dos assimilados é para atender a demanda de folhas, hastes, ramificações e
finalmente raízes. Na ocorrência de limitações de algum dos fatores que
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
determinam diminuição na oferta de carbono (C), a intensidade de demanda
de cada um dos compartimentos é alterada, e a prioridade de alocação de
assimilados passa a ser para raízes e reservas.
A translocação dos produtos da fotossíntese da fonte (local de produção
ou reserva de assimilados) para os diversos drenos se faz pelo floema. Embora
o modelo de transporte pelo floema represente simplesmente o movimento da
fonte para o dreno, o caminho específico entre eles é altamente complexo, pois
nem todos os drenos são igualmente supridos por todas as folhas da planta,
uma vez que cada fonte supre drenos específicos, sendo que o modelo de
distribuição dos assimilados é, em grande parte, função da proximidade entre
os órgãos fontes e os drenos (Taiz & Zeiger, 2004).
O conhecimento das modificações no consumo de C e de sua partição
entre os principais componentes do rendimento (folhas, hastes, órgãos
reprodutivos e raíz) provocadas por limitações ambientais é essencial não
apenas para realizar diagnósticos, mas, sobretudo, para construir modelos
preditivos. Identificar e quantificar os fatores ambientais que limitam a
expressão do potencial das espécies forrageiras possibilita estabelecer a
melhor forma de manejar esses fatores, com a finalidade de otimizar a
produção animal (Silva et al., 2001).
2. Produção de assimilados através da fotossíntese
A fotossíntese é o principal processo realizado pelas plantas, em que
transforma a energia luminosa em energia química processando o dióxido de
carbono (CO2), água (H2O) e elementos minerais em compostos orgânicos,
com formação de matéria orgânica (96% da planta) e liberação de oxigênio
(O2). Em plantas superiores, o mesófilo é o tecido mais ativo na assimilação de
CO2, em razão da riqueza de suas células em cloroplastos (Taiz & Zeiger,
1998).
Cerca de 90% da massa seca vegetal são originadas diretamente da
assimilação
fotossintética
de
carbono
através
da
fotossíntese.
A
taxa
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
fotossintética da folha determina a quantidade total de carbono fixada
disponível para a folha (Taiz & Zeiger, 1998) e é função de fatores intrínsecos,
como o nível de inserção e a idade, e de fatores extrínsecos às folhas, como a
irradiância, a água no solo, a temperatura e os nutrientes. Como resultado
desse conjunto de fatores, as folhas de mais alto nível de inserção no perfilho,
mais
jovens
e
melhor
iluminadas,
fotossintetizam
mais
e
geralmente
contribuem mais para o pool de fotoassimilados da planta que as folhas mais
velhas, de mais baixos níveis de inserção. A biomassa vegetal pode ser
limitada tanto pela produção como pelo uso dos fotoassimilados por parte dos
meristemas foliares (Lemaire & Agnusdei, 1999).
O carbono fixado pelas folhas pode ser alocado para três caminhos
metabólicos: (i) para utilização no metabolismo celular, fornecendo energia e
esqueletos de carbono para a síntese de outros compostos, (ii) para a síntese
de compostos de transporte, exportados para os diversos drenos da planta e
(iii) para síntese de compostos armazenados, para utilização durante a noite
(Taiz & Zeiger, 1998).
2.1 Destino dos Fotoassimilados
A partir dos produtos intermediários do Ciclo de Kalvin, são sintetizados
pelas plantas os diferentes compostos orgânicos, cuja composição depende da
espécie. As monocotiledôneas caracterizam-se pela alta produção de amido
armazenado nos grãos (Trigo, milho, arroz, aveia, cevada, centeio, triticale,
painço, sorgo, alpiste) ou de sacarose armazenada nos colmos (cana e sorgo
sacarino). Nas dicotiledôneas predominam as reservas de proteínas e óleos
como produtos de reserva. Os principais produtos resultantes da fotossíntese
são:
-
Monossacarídeos: Os principais monossacarídeos, glicose e frutose, são
sintetizados através do Ciclo de Kalvin (C3) ou da PEP carboxilase (C4).
-
Dissacarídeos: A sacarose é um importante dissacarídeo nas plantas
cultivadas (frutose+glicose) envolvida no transporte de glicídeos. A maltose é
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
outro dissacarídeo, obtido pela degradação do amido de reserva do grão de
cevada, utilizando como substrato para Sacharomyces cerevisae.
-
Polissacarídeos: Consistem em cadeias de monossacarídeos sendo os
mais importantes em células vegetais o amido e a celulose.
-
Proteínas: Parte das trioses, pentoses e hexoses formadas durante a
fotossíntese é convertida em α-cetoácidos e estes, combinando-se com amônia
ou enxofre formam os aminoácidos. Esses aminoácidos são convertidos em
proteínas vegetais, com funções estruturais, funcionais (enzimas) ou de
reserva (grãos de aleurona). As sementes contém grande quantidade de
proteínas de reserva, como albuminas, globulinas e prolaminas.
-
Lipídios: Fosfolipídeos (constituinte de membranas), esfingolipídeos,
cerídeos, cutina e suberina são alguns lipídeos produzidos nas plantas.
-
Compostos
fotoassimilados
secundários:
também
são
além
do
convertidos
metabolismo
em
outros
primário,
compostos
os
pelo
metabolismo secundário. A natureza desses compostos depende do fenótipo e
das condições de solo e clima do local de cultivo. Também são diversas as
funções
desses
compostos,
como
estruturais,
pigmentos,
hormônios
e
compostos que protegem as plantas de predadores e patógenos, com base na
sua toxicidade e na capacidade de repelir herbívoros e microorganismos. De
maneira geral, os compostos secundários são divididos em três grandes
grupos: (i) terpenos ou terpenóides; (ii) fenóis e (iii) compostos secundários
nitrogenados.
3. Os processos e rotas da partição de assimilados
3.1 Transporte no floema
O acúmulo de matéria seca no órgão de interesse econômico é
controlado pela troca de metabólitos entre os tecidos fornecedores de
fotoassimilados (a fonte) e o órgão colhido (o dreno colhido). Portanto, são
considerados fontes
os órgãos
ou tecidos que exportam
assimilados,
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
sintetizando-os (tecidos verdes), ou estocando-os anteriormente (como dreno
reversível) e cedendo-os posteriormente. Já os drenos são os órgãos ou
tecidos que utilizam (drenos em crescimento) ou armazenam fotoassimilados
(drenos de reserva) (Pimentel, 1998; Marenco & Lopes, 2005). Esses drenos
podem ser irreversíveis ou permanentes, como os meristemas vegetativos de
parte aérea ou raíz; e reversíveis, como o meristema intercalar de folhas, de
flores e de inflorescências, os entrenós de cana-de-açúcar, o tronco de
árvores, os tecidos de reserva de frutos, de sementes, de tubérculos, de
bulbos e de bulbilhos (Pimentel, 1998).
Vários fatores influenciam a alocação e a partição de assimilados nos
distintos órgãos da planta, sendo os principais a atividade fotossintética da
fonte, a força do dreno e os fatores que afetam o descarregamento no floema.
Em espécies que crescem em associação, a partição de assimilados também é
influenciada pela competição interespecífica (Marenco & Lopes, 2005). A
alocação de assimilados refere-se ao destino ou uso do carbono fixado
(sacarose ou amido para uso na respiração), enquanto que a partição refere-se
a distribuição diferencial dos fotoassimilados nos diferentes órgãos da planta.
O floema é o tecido que transloca os produtos da fotossíntese
(fotoassimilados) das fontes para os drenos, incluindo raízes, folhas novas e
frutos em desenvolvimento.
Em uma folha fotossinteticamente ativa, através do Ciclo de Kalvin
ocorre a conversão da triose fosfato em glicose, por enzimas reguladoras. No
citosol, as enzimas de síntese de sacarose combinam uma molécula de glicose
a uma de frutose (um intermediário da síntese de glicose). A sacarose é um
importante composto na partição e transporte de assimilados. Várias plantas,
inclusive a maioria dos cereais tem capacidade para armazenar sacarose no
vacúolo das células foliares.
Sob condições onde a concentração de sacarose na folha é alta,
pastagens temperadas e espécies de cereais podem também armazenar
frutanas solúveis (polímeros curtos de frutose) no vacúolo das células do
mesófilo. Contudo, dentro do cloroplasto a frutose é convertida em glicose, a
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
qual é polimerizada para dar amido insolúvel. Estes componentes do sistema
mantém o nível de sacarose no citosol das células foliares, pela troca de
sacarose e ou frutanas no vacúolo, e por mobilização e deposição de amido no
cloroplasto.
Durante períodos de rápida fotossíntese, as reservas de sacarose e
amido podem ser preenchidas. Contudo, no escuro ou intensa demanda de
assimilados (para o local de uso ou transporte para reservatórios distantes) a
sacarose vacuolar pode ser mobilizada rapidamente. A subseqüente hidrólise
do amido e o transporte do fosforilato intermediário resultante atravessa a
membrana do cloroplasto e pode prover a demanda por períodos de escuro
prolongados.
Os níveis de sacarose no citosol são mantidos baixos por hidrólise da
glicose e frutose, e pela incorporação dos açucares resultantes, normalmente
amido, armazenados nos plastídeos. Os diferentes resultados da concentração
de sacarose geram uma pressão de turgor no lúmen dos elementos do floema,
promovendo uma força que dirige, por fluxo de massa, a sacarose de órgãos
fonte para os drenos. Os assimilados são também transportados e depositados
em outros sítios de reserva na planta, estes incluem pequenas reservas em
todos os meristemas ativos e grandes reservas nos colmos e também na
bainha das folhas, raízes e estruturas reprodutivas, como nas paredes das
vagens
em legumes. Nestes
sítios
de
curto-prazo, as
reservas estão
normalmente na forma solúvel: monossacarídeos, sacarose e frutanas. Por
exemplo, as frutanas predominam nas células dos colmos em pastagens
temperadas e cereais, e sacarose é estocada em espécies C4 (milho e cana-deacúcar).
Ao contrário do movimento da água e sais minerais absorvidos pelas
raízes, cuja direção no xilema é essencialmente no sentido ascendente,
seguindo o gradiente de potencial hídrico, os fotoassimilados movimentam-se
pelo floema em qualquer sentido (folhas e raízes, por exemplo). A translocação
de assimilados das raízes para a parte aérea é um fenômeno comum em
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
plantas que acumulam reservas em estruturas subterrâneas da fase vegetativa
para a época de frutificação.
O transporte de assimilados nos tubos do floema não depende de gasto
energético, pois trata-se de um movimento passivo, que acompanha um
gradiente de pressão (∆P) no interior dos elementos condutores, seguindo o
modelo de fluxo-pressão. Este modelo associa o movimento de materiais pelo
floema ao gradiente de pressão, ou seja, há diferenças de pressão de
turgescência
entre
o
local
de
carregamento
(P-fonte)
e
o
local
de
descarregamento no floema do dreno (P-dreno). Este assunto será discutido
em capítulo posterior.
O transporte de elementos do tubo crivado com fotoassimilados realizase geralmente nos terminais do floema associado à células do mesófilo foliar
ou células-fonte. Entretanto, transporte lateral de solutos do floema para
tecidos adjacentes também ocorre, sendo de grande importância o acúmulo de
assimilados nos caules, constituindo um depósito de reserva temporário capaz
de suprir a planta em períodos de escassez de fotoassimilados. Nesse caso, os
açucares de reserva são remobilizados e redistribuídos via floema. O caule
normalmente funciona como órgão de reserva temporária, remobilizando e
redistribuindo assimilados durante a noite.
Os solutos mais importantes transportados pelo floema são produtos da
fotossíntese. Levando-se em conta que somente algumas partes da planta
fazem fotossíntese, o transporte no floema é essencial para que todos os
órgãos do vegetal sejam supridos. No caso do transporte de solutos
inorgânicos, o floema também é de importância fundamental. Como a força
motriz para a absorção de solutos inorgânicos normalmente é a transpiração,
os nutrientes provenientes do solo tendem a se acumular nos órgãos que
transpiram mais, como as folhas maduras, em detrimento dos que transpiram
menos, como os brotos novos e frutos. Para corrigir isso, os vegetais
redistribuem os nutrientes de um órgão para outro através do floema. Uma
conseqüência disso é que aqueles nutrientes que não são transportados pelo
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
floema (Ca+2 , SO4-2e Fe) ficam em baixas concentrações em órgãos que
transpiram pouco.
O movimento de sacarose do floema para os sítios de reserva envolve:
efluxo dos elementos crivados, difusão em gradientes de baixa concentração
na semente por apoplasto, absorção nos grãos em desenvolvimento por células
especializadas e transporte simplasto para os sítios de deposição de amido. Por
sua vez, a taxa de movimento das moléculas de sacarose das reservas
(cloroplasto das folhas ou reservas a curto prazo) para os drenos pode ser
limitado por: eventos bioquímicos nas folhas, transporte do floema ou sítios de
reservas para os elementos crivados, transporte dos elementos crivados para
os sítios de reserva no dreno e ou eventos bioquímicos no dreno.
3.2 O modelo de Fluxo-Pressão
O
modelo
de
fluxo-pressão
fundamenta-se
no
fato
de
que
o
carregamento do floema com assimilados da fonte baixa seu potencial
osmótico (PS) para valores de –2 a –3 Mpa, muito menor do que os
encontrados nas células do mesófilo (-1,3 a –1,8 Mpa). O baixo PS do floema
na fonte (carregamento) provoca absorção de água e, conseqüentemente,
aumento no potencial de pressão (PP) para valores próximos a 1 Mpa.
Estima-se que uma diferença de 0,12 a 0,46 Mpa entre o sítio de
carregamento na fonte e o dreno seja suficiente para impulsionar o movimento
em massa de assimilados pelo floema. Na maioria das espécies a sacarose, o
K+ e seus ânions acompanhantes são os solutos que mais contribuem com a
manutenção da pressão de turgescência do floema. O ∆P entre o local de
carregamento (fonte) e o de descarregamento (dreno) é um dos fatores que
determinam a velocidade de transporte no floema.
Até o começo da década de 80 o movimento a longa distância no floema
era o principal componente estudado, e para tal, muitas teorias foram
levantadas. Nesse contexto, o estudo dos processos de carregamento e
descarregamento ficou relativamente negligenciado. Embora hoje tenhamos
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
claro que esses dois processos são na verdade os mais relevantes para, por
exemplo, manipular um dreno de interesse agronômico, o conhecimento do
transporte a longa distância não pode, por sua vez, ser negligenciado. Sobre
esse tópico, sabe-se que para uma distância de um metro, um determinado
soluto demorará em média uma hora para ser translocado. Embora a
velocidade média no floema seja, portanto de 1 m/h, ela pode variar de 0,3 a
1,5 m/h. Essa velocidade é rápida e evidencia que o transporte não é por
difusão.
A título de comparação, a taxa de difusão é de 1 m a cada 32 anos.
Outra característica do transporte no floema é que ele, ao contrário do xilema,
é feito por pressão e não por tensão. Uma prova disso é o fato dos afídios não
necessitarem sugar o conteúdo floemático, sendo que esse continua fluindo
pelos estiletes desses insetos, mesmo quando o resto do corpo é removido.
Atualmente a teoria mais aceita para explicar o transporte à longa
distância no floema é a do “Fluxo de Pressão”. Essa teoria foi proposta por
Münch em 1930 e perfaz os requisitos expostos acima. O fluxo de pressão é
gerado pelo gradiente de potencial de pressão (yP) entre a fonte e o dreno. O
gradiente de pressão, por sua vez, gerado pelas alterações no potencial hídrico
(yH) devido aos processos de carregamento descarregamento. Na fonte o
carregamento de solutos (fotoassimilados) abaixa o potencial osmótico (yS) e
conseqüentemente o yH do tubo crivado. O abaixamento do yH provoca a
entrada de água vinda do xilema, aumentando o potencial de pressão (yP) do
tubo crivado nessa região. No dreno, o descarregamento de solutos aumenta o
yH do TC, fazendo com que a água saia para as células adjacentes. Há uma
diminuição do yP e o gradiente de yP entre a fonte e dreno é a pressão de
turgescência necessária ao movimento no floema. Em última análise, pode se
dizer que o gradiente de pressão é estabelecido como conseqüência do
carregamento do floema na fonte e do descarregamento no dreno (Figura 1).
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
Figura 1. Teoria do fluxo de pressão para o transporte no floema.
É interessante notar que na teoria de Münch, a água move-se no floema contra
um gradiente de potencial hídrico (yH) da fonte para o dreno. Esse movimento
de água não vai contra as leis da termodinâmica, pois ela está se movendo por
fluxo de massa e não por osmose. O movimento de água é, portanto conduzido
por um gradiente de pressão e não por um gradiente de yH. Embora o fluxo de
pressão seja passivo, ele é gerado por processos ativos de absorção de solutos
na fonte e no dreno. Por fim, é importante ressaltar que se não existissem as
placas crivadas nos tubos crivados, os gradientes de yS, yH e yP não seriam
possíveis, pois os sistemas tenderiam rapidamente a um equilíbrio.
Matematicamente,
o
volume
do
fluxo
de
seiva
(Jv)
pode
ser
representado pela expressão:
Na expressão acima, Lp representa a condutividade hidráulica e yP1 e
yP2 são os potenciais de pressão da fonte e do dreno, respectivamente. Essa
equação indica que uma diminuição artificial do potencial de pressão no dreno
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
(yP2) aumentaria o volume do fluxo de seiva em um dreno. Uma maneira de
se fazer isso seria diminuir o potencial osmótico (através da adição de solutos)
de uma solução externa ao dreno, o que provocaria a saída de água e
conseqüente diminuição do potencial de pressão das células do dreno.
Consistente com essa observação, o tratamento de raízes de plântulas
de ervilha com manitol aumentou a importação de [14 C]-sacarose (Schultz,
1994), possivelmente porque houve diminuição do potencial de parede nas
células do dreno. No referido experimento, a adição de 350 nM de manitol
aumentou a incorporação de sacarose em mais de 300% (Schultz, 1994).
Embora tenhamos enfatizado até aqui que os processos de carregamento e,
sobretudo de descarregamento sejam os alvos primários para manipulações
biotecnológicas visando aumentar a produtividade agrícola, os experimentos de
Schutz (1994) evidenciam que o controle do transporte a longa distância
também pode favorecer o descarregamento em drenos de importância
agronômica.
3.3 Carregamento de fotoassimilados
O carregamento do floema na região da fonte envolve o movimento dos
produtos dos cloroplastos nas células do mesófilo para as células do tubo
crivado (TC). Esse processo ocorre nas nervuras terminais das folhas.
Analisando o processo de carregamento sob o ponto de vista anatômico e
bioquímico, vemos que a triose fosfato produzida no estroma dos cloroplastos
deve ir para o citoplasma e se converter em sacarose. A sacarose deve migrar
das células do mesófilo para a vizinhança dos TC nas nervuras terminais das
folhas. Finalmente, a sacarose deve entrar no complexo células-companheiras
(CC)- tubos crivados (TC). Existem dois tipos principais de carregamento: o
simplástico (através dos plasmodesmas) e o apoplástico (Fig. 2). Nesse
último caso, os açúcares presentes no espaço intercelular e na parede celular
(apoplasto)
devem
ser
transportados
ativamente
para
membrana citoplasmática e entrarem no complexo CC-TC.
atravessarem
a
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
Figura 2. Carregamento simplástico (setas azuis) e apoplástico (setas
vermelhas), (Taiz e Zeiger, 1998)
Há uma distinção entre carregamento do floema e carregamento do
complexo CC-TC. O carregamento do floema refere-se ao caminho como um
todo que os fotoassimilados tomam a partir do mesófilo até o complexo CC-TC.
Por outro lado, o carregamento do complexo CC-TC restringe-se à descrição da
entrada do fotoassimilado neste. O carregamento do complexo CC-TC é
simplástico quando há uma continuidade simplástica entre este e as células
adjacentes e é apoplástico quando envolve a passagem dos solutos pela
membrana. O carregamento do floema é simplástico quando todo o caminho é
simplástico. De modo diferente, o carregamento do floema será apoplástico se
os plasmodesmas estão ausentes em algum ponto do caminho, independente
do local onde há a descontinuidade simplástica. Desse modo, uma espécie
pode ter carregamento apoplástico do floema, mesmo que o carregamento do
complexo CC-TC seja simplástico.
A via apoplástica de carregamento ocorre em plantas que possuem CC
comum ou células de transferência nas nervuras terminais. Essas espécies
transportam
quase
que
exclusivamente
a
sacarose
(exemplo,
família
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
Solanaceae e Fabaceae). Espécies com CC intermediárias fazem carregamento
via simplasto. Essas espécies transportam 20 a 80% de seus açúcares na
forma de rafinose e/ou estaquiose, além da sacarose. Um requisito importante
para o carregamento apoplástico é a ausência de transpiração na região onde
ele ocorre. Caso haja transpiração, a tensão gerada pelas forças de adesão e
coesão no apoplasto pode carrear a sacarose, impedindo seu carregamento no
complexo CC-TC.
Nas folhas fontes o acúmulo de sacarose no complexo CC-TC faz com
que o potencial osmótico (ys) seja mais negativo nesse local do que no
mesófilo. Em plantas com carregamento apoplástico, a sacarose do apoplasto
entra no complexo CC-TC através de um carregador localizado na membrana
plasmática que faz simporte entre sacarose e prótons (H+) (Fig.3). A presença
do carreador de sacarose é importante também para garantir um fluxo
contínuo desse açúcar vindo do mesófilo para o apoplasto, pois ele mantém o
potencial químico dessa substância sempre baixo no apoplasto.
Figura
3.
Transportadores
de
sacarose
necessário
no
carregamento
apoplástico, Taiz e Zeiger (1998)
No carregamento apoplástico, antes da sacarose ser carregada no TC da
fonte, deve haver um efluxo desse açúcar do mesófilo para o apoplasto. É
possível que existam células especializadas no mesófilo para fazer tal processo.
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
Sabe-se que o potássio estimula essa etapa de saída de fotoassimilados do
simplasto para o apoplasto. Portanto, uma boa nutrição com potássio tende a
aumentar a sacarose no apoplasto, e isso induz seu carregamento no TC e, em
última análise, o crescimento do dreno. Por outro lado, uma diminuição do
conteúdo de sacarose no apoplasto da fonte pode reduzir drasticamente o
carregamento de plantas que tem esse processo via apoplasto.
No carregamento por simplasto, obviamente, não há a necessidade de
um carreador de sacarose para que ela atravesse a membrana plasmática.
Contudo, o acúmulo de açúcares no complexo CC-TC impede que esse
processo seja feito por simples difusão, pois com o passar do tempo o fluxo se
reverteria. Uma hipótese para esse tipo de transporte seria o modelo de
armadilha de polímero proposto por Turgeon (1991). Nesse modelo, a sacarose
move-se do mesófilo para a célula intermediária onde ela se converte em
rafinose mantendo o gradiente de difusão. Rafinose e estaquiose não
retornariam ao mesófilo, pois são muito grandes (os plasmodesmas que ligam
o mesófilo à CC devem ser menores que os que ligam o CC ao TC).
Uma questão interessante é o porquê da existência de dois tipos de
carregamento (simplástico e apoplástico). Sabe-se que o carregamento
simplástico é mais comum em árvores e arbustos da região tropical úmida. Por
outro lado, o carregamento apoplástico predomina em plantas herbáceas de
regiões temperadas e zonas áridas. Uma das hipóteses para explicar a
existência de tipos diferentes de carregamento seria uma adaptação à
temperatura e à seca. Pode ser que o carregamento apoplástico seja uma
adaptação à inibição do carregamento simplástico em baixas temperaturas e
estresse hídrico. Em baixas temperaturas há um aumento da viscosidade da
seiva e conseqüente diminuição do fluxo de pressão. Pelo menos em regiões
temperadas, as espécies com carregamento simplástico geram menor fluxo de
pressão, possivelmente devido ao transporte de oligossacarídeos, os quais, em
baixas temperaturas tendem a aumentar a viscosidade. Além disso, a
capacidade das células intermediárias de produzirem oligossacarídeos pode
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
diminuir
muito
em
baixas
temperaturas.
De
modo
geral,
a
taxa
de
carregamento simplástico tende a ser menor que o apoplástico.
3.4 Descarregamento
De todas as etapas do transporte no floema, o descarregamento é o que
parece estar mais diretamente ligado com as diferenças na capacidade de
armazenamento entre os diferentes tipos de drenos. Com as perspectiva de se
aumentar o descarregamento em drenos de interesse econômico, tais como
frutos, sementes, tubérculos e raízes tuberosas, esse processo vem sendo
objeto de intensos estudos. Assim como o carregamento do floema que ocorre
na fonte, o descarregamento que ocorre nos drenos envolve uma série de
etapas, tais como: 1) Descarregamento do soluto vindo do TC. 2) Transporte a
curta distância depois do descarregamento até as células do dreno 3)
Armazenamento e metabolismo do açúcar no dreno. Essa terceira etapa
(armazenamento e metabolismo do fotoassimilado) é o que denominamos
alocação de assimilados e será discutida posteriormente. A duas primeiras
etapas constituem o descarregamento propriamente dito, o qual pode ser do
tipo simplástico ou apoplástico (Fig. 4). O descarregamento apoplástico admite
duas possibilidades, sendo que o descarregamento do TC pode ser apoplástico
(tipo1, Fig. 4) ou pode ser simplástico, possuindo uma etapa apoplástica
posterior (tipos 2A e 2B, Fig. 4).
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
Figura 4. Tipos de descarregamento. No descarregamento apoplástico, a saída
do tudo crivado pode ser apoplástica (tipo 1) ou simplástica com uma
etapa apoplástica posterior (tipos 2A e 2B). C = célula companheira;
T = tubo crivado; R = célula receptora, Taiz e Zeiger (1998).
A ocorrência de descarregamento simplástico ou apoplástico varia de
acordo com a espécie vegetal, o tipo de tecido ou órgão e a fase de
desenvolvimento (Fig. 5). Um bom exemplo é o fruto de tomate, o qual possui
um descarregamento simplástico e uma acumulação de amido no começo do
desenvolvimento e posteriormente um descarregamento apoplástico e um
acúmulo de açúcares solúveis. O tipo de descarregamento com maior
capacidade de transporte é o simplástico e por isso o apoplástico é reservado
para situações especiais.
O descarregamento do floema tende a ser simplástico (Fig. 5A e B) em
drenos nos quais o carbono é estocado em polímeros como amido e proteínas,
ao invés de sacarose. Regiões meristemáticas caulinares e radiculares
normalmente também possuem descarregamento do tipo simplástico. Nos
ápices radiculares, os fotoassimilados chegam à região subapical desses órgãos
através dos elementos do protofloema. Existem plasmodesmas ligando os TCs
desses elementos com as células meristemáticas nos ápices radiculares. O
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
descarregamento nos ápices caulinares é parecido com o que ocorre no ápice
radicular. Um exemplo de ápice caulinar que se transforma em um órgão de
reserva é o tubérculo de batata.
Figura 5. Exemplos de descarregamento simplástico (a, b) e apoplástico (c,
d), Patrik (1997)
Desse modo, o descarregamento desse dreno de estoque de polímeros é
simplástico (Fig. 5A), assim como o meristema caulinar que o originou. Apesar
da cana-de-açúcar acumular açúcares solúveis e não polímeros, o processo de
descarregamento nos colmos que acumulam sacarose é necessariamente
simplástico, pois há uma barreira apoplástica na parede celular das células da
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
bainha do feixe vascular (Fig. 5B). O descarregamento apoplástico (Fig. 5C e
D) é comum em monocotiledôneas (ex. folhas jovens de milho), órgãos de
reserva que acumulam mono e dissacarídeos ao invés de polímeros (ex. raízes
de beterraba e cenoura), frutos (tomate, uva, laranja) e em sementes em
desenvolvimento. Nas
sementes
em desenvolvimento
é
necessário um
descarregamento apoplástico, pois não há conexão simplástica entre o tecido
materno e os tecidos do embrião (Fig. 5D). O descarregamento apoplástico em
raízes que acumulam carboidratos como, a beterraba e a cenoura, é feito em
três etapas: 1) efluxo do açúcar para o apoplasto, 2) passagem do açúcar para
as células parenquimáticas do floema e 3) passagem simplástica para as
células parenquimáticas de estoque (Fig. 5C).
Durante o processo de descarregamento, é necessário que se mantenha
o potencial de sacarose sempre baixo na célula receptora, para que não haja
refluxo desse açúcar. Em órgãos com descarregamento simplástico, o baixo
potencial químico de sacarose é mantido pela respiração ou pela conversão dos
açúcares transportados em compostos necessários para o crescimento ou em
polímeros de estoque. Já em órgãos com descarregamento apoplástico, as
invertases desempenham um papel central na manutenção de um baixo
potencial de sacarose para que haja uma chegada contínua desse composto
nas células receptoras.
No descarregamento apoplástico pode haver necessidade de gasto de
energia para que os assimilados atravessem membranas. Nesse tipo de
descarregamento, o açúcar deve atravessar duas membranas: a do TC e a da
célula receptora. Às vezes há necessidade de atravessar a membrana do
tonoplasto também, se a sacarose for armazenada no vacúolo. Embora não
haja necessidade de atravessar membranas no descarregamento simplástico,
há necessidade de energia para o metabolismo e biossíntese de polímeros de
reserva. Em leguminosas, tanto a saída do TC quanto a entrada na célula
receptora são processos ativos. Em trigo, a saída é passiva (a favor de
gradiente) e a entrada é ativa.
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
4. Características morfofisiológicas da fonte e do dreno
4.1 Fonte
As folhas de maior importância para a produção dos drenos colhidos são
aquelas próximos a estes. Em uma frutífera como a macieira, são necessárias
cerca de 15 folhas próximas ao fruto para o enchimento deste. No milho, a
principal fonte de fornecimento de fotoassimilados para a espiga é a folha onde
essa espiga se apóia. Nessa planta, as partes verdes da espiga também
contribuem para o seu enchimento (WARDLAW, 1990). Para a vagem de uma
leguminosa, que também contribui para o seu enchimento, o trifolíolo que se
desenvolve junto a ela terá a maior participação para o seu enchimento
(DAVIES, 1995). Em trigo e arroz, a chamada folha bandeira tem uma
importância enorme para o fornecimento de fotoassimilados para a panícula
(FAGERIA, 1992). No trigo de barba, a arista desenvolvida faz também
fotossíntese. Essas características da fonte que afetam a produtividade da
cultura, levaram HSIAO (1990) a ressaltar as seguintes características
importantes para a fonte: Uma grande área foliar para uma planta de clima
tropical, além de permitir alta interceptação de energia luminosa, vai significar
também uma grande superfície de transpiração, o que é indesejável. O
melhoramento vegetal tradicional sempre selecionou plantas com alta área
foliar, pois em clima temperado a luminosidade é um fator limitante. Isto
dificilmente ocorrerá em clima tropical, principalmente para plantas C3, que se
saturam a baixas intensidades luminosas.
Energia luminosa não é fator limitante para a agricultura tropical,
portanto, segundo SINCLAIR & HORIE (1989), devemos selecionar plantas com
maior área específica de folha, isto é, plantas com maior peso de folha por
unidade
de
área.
A
atividade
fotossintética
é
função
do
número
de
cloroplastos, sejam eles dispostos horizontalmente (maior área foliar), sejam
dispostos verticalmente (maior espessura e área específica). Uma folha mais
espessa e menos larga vai manter alta atividade fotossintética por unidade de
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
área (intensidade da fonte), com menor superfície de transpiração, o que é
desejável em clima tropical.
A área foliar fotossintéticamente efetiva é extremamente importante.
Folhas que não sejam iluminadas adequadamente, por serem sombreadas
pelas folhas superiores, serão mais um dreno ao invés de fonte. Em
gramíneas, como o arroz, a seleção de plantas com folhas eretas (seleção para
lígula e aurícula bem desenvolvidas), permitindo maior penetração de luz no
dossel, trouxe aumento de produtividade, o que ainda não se conseguiu com o
trigo, cevada e o milho, que têm baixo índice de área foliar (WARDLAW,1990).
A folha, em seus estádios iniciais de desenvolvimento, é considerada um
tecido heterotrófico. Na própria folha, a zona de crescimento, em geral na base
dessa, funciona como dreno para as outras partes já desenvolvidas e somente
o excedente é exportado para outros órgãos (WARDLAW, 1990). Em folhas de
dicotiledôneas, somente ao atingir de 30 a 60% de sua área foliar máxima, ela
passa a ser autotrófica e exportadora de fotoassimilados. Em gramíneas, como
a cana-de-açúcar, a importação de carboidratos pelo floema se mantém até a
folha atingir 90% de sua área foliar máxima.
A transição na folha, de dreno para fonte, em plantas cujo principal
carboidrato de transporte é a sacarose, está associada a um aumento da
atividade da sacarose-P sintase, enzima para a síntese de sacarose, e
decréscimo da atividade da invertase ácida e sacarose sintase, enzimas de
hidrólise da sacarose que chega pelo floema (MARSCHNER, 1995).
Na folha, após atingir o tamanho máximo, inicia-se o processo de
senescência, com diminuição da fotossíntese e exportação de compostos
principalmente nitrogenados. Enzimas hidrolíticas, seqüestradas no vacúolo,
são liberadas para o citoplasma, iniciando a hidrólise protéica, começando
então, o processo de perda da funcionalidade membranar e degeneração da
integridade celular, em maior ou menor intensidade, dependendo do vegetal
(VIERSTRA, 1996). O declínio do teor de clorofila, causando clorose, é um
sintoma visível da senescência. A composição do floema muda, com diminuição
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
de açúcares e aumento de compostos nitrogenados de baixo peso molecular e
de nutrientes minerais móveis.
A duração do período autotrófico da folha é primordial para obtenção de
máxima produtividade, pois se no momento do desenvolvimento dos órgãos de
interesse a fonte estiver em processo de senescência, o enchimento desses
órgãos
ficará
comprometido.
A
habilidade
da
folha
em
exportar
fotoassimilados, no entanto, é regida pelo balanço entre a produção e o
consumo de carbono nesta e nos drenos existentes. Se o consumo de carbono
for baixo, há uma diminuição da atividade fotossintética na fonte. A
acumulação de amido no grão e conseqüente produtividade em trigo são
também dependentes do número de células e de plastídeos na folha e no
endosperma do grão, assim como em milho e soja.
Normalmente o potencial fotossintético de uma folha não é totalmente
utilizado, e a remoção de algumas folhas-fonte aumenta a fotossíntese nas
restantes. O declínio, entretanto, da taxa fotossintética no período da tarde, é
a expressão de uma limitação do dreno devido à acumulação de açúcares,
assim como do fechamento estomático. Em condições de alta luminosidade
pode ocorrer fotoinibição, a menos que hajam drenos temporários, como as
bainhas e ramos de cereais. Quando a demanda por fotoassimilados é alta, há
uma diminuição de ácido abscísico nas folhas, levando à máxima abertura
estomática (como será visto adiante). Com a grande abertura estomática,
aumenta a transpiração e a síntese de citocininas nas raízes, que é
transportada para as folhas, impedindo a senescência foliar (também será
visto adiante).
Logo, os fitormônios também estão envolvidos no controle da senescência
(PELL & DANN, 1990).
4.2 Características do Dreno
Quando se inicia o desenvolvimento do fruto e de sementes, há uma
dominância do crescimento destes órgãos sobre os órgãos vegetativos (parte
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
aérea e raíz), enquanto as flores ainda são pobres competidores. Alguns
tecidos de reserva subterrâneos também são dominantes. Logo, em plantas
perenes de produção anual, como é o caso do café e de frutíferas, uma
frutificação pesada em um ano vai reduzir o crescimento vegetativo (novas
folhas) para o ano seguinte, e resultar em baixa frutificação (WARDLAW,
1990).
Algumas práticas de manejo de certas culturas são uma interferência
direta nas relações fonte/dreno. Por exemplo, com frutíferas, o “raleio” dos
primeiros frutos vai permitir ainda um desenvolvimento vegetativo, e os frutos
produzidos, depois, em menor número, serão de maior tamanho, característica
desejável para a comercialização. A “capação” no tomateiro, que é a
eliminação da gema vegetativa terminal, vai diminuir o tamanho da planta e o
número de frutos, que serão maiores (WARDLAW, 1990).
Para as características do dreno, HSIAO (1990) sugere as seguintes
características: Um dreno a ser priorizado em clima tropical é o sistema
radicular, pois água e nutrientes são fatores limitantes e podem afetar
drasticamente o comportamento da fonte e do dreno a ser colhido, fato menos
evidente em clima temperado, pois a evapotranspiração em geral é menor. Por
isto, o melhoramento vegetal tradicional, durante a revolução verde nos anos
50 e 60, não buscou plantas com o sistema radicular muito desenvolvido, pois
nem água nem nutrientes eram, na maioria dos cultivos, limitantes. Hoje em
dia tal pensamento já se modificou mesmo em relação ao clima temperado,
buscando-se plantas mais eficientes no uso de nutrientes.
Em clima tropical, no entanto, grande parte da agricultura para a
produção de alimentos é feita com baixa tecnologia, em regiões de alta
evapotranspiração, sem irrigação e, visto o alto custo da aplicação de
fertilizantes, conduzida com baixa disponibilidade de nutrientes. Portanto, a
seleção de plantas com sistema radicular bem desenvolvido, para profundidade
e área radicular, apesar de não ser um órgão colhido para a maioria das
culturas, permitirá aumentos de produtividade (PUGNAIRE et al., 1994).
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
HENDRIX (1994) propõe que 44% do carbono fixado pela fotossíntese
vão para a raíz, com 1/4 desse valor utilizado no crescimento, e o restante na
respiração de manutenção. O mesmo autor afirma que para plantas em
simbiose com o Rhizobium, pelo menos 12% dos fotossintatos produzidos pela
planta são gastos na respiração e crescimento dos nódulos, assim como em
plantas micorrizadas, 5 a 10% destes fotossintatos são usados pelo fungo.
Estes gastos com a raiz, assim como na simbiose, são vantajosos para a planta
de clima tropical devido à carência de nitrogênio, que será assimilado pelo
sistema radicular mais eficiente e/ou pela simbiose com o Rhizobium. Da
mesma forma, a associação com fungos micorrízicos trará benefícios ao
aumento do volume radicular e conseqüente absorção de água e fósforo,
ambos limitantes em solos tropicais. Portanto, este gasto de energia em um
órgão não colhido trará vantagens para a eficiência no uso de água, nitrogênio
e fósforo, que são limitantes na agricultura tropical.
Tendo em vista a maior probabilidade de ocorrência de temperaturas
altas e falta de água em clima tropical, um vegetal que possua uma floração e
frutificação mais longas, indeterminadas, terá mais chances de produzir um
maior número de drenos reprodutivos, como a prolificidade do milho, com mais
de uma espiga aproveitável por colmo, ou a soja, que comparada ao feijão,
apresenta um período de floração e produção de vagens mais longo. Havendo
um estresse ambiental durante uma parte do período de floração e de
frutificação, ainda existem chances de que esse estresse termine antes do final
da produção dos drenos (WARDLAW, 1990). Além dessa vantagem, o milho
prolífero é mais eficiente no uso de N do que o milho não prolífero.
O transporte de carbono é dependente da idade fisiológica dos tubos
crivados. Com o envelhecimento destes, os poros das discutíveis placas
porosas começam a ser bloqueados por P-proteínas e calose. Para a maioria
das plantas, à exceção de monocotiledôneas que não sintetizam P-proteínas,
esses dois compostos é que regem a permeabilidade das placas porosas.
Em clima tropical, os estresses ambientais, como altas temperaturas e
falta de água são bastantes comuns, e por isso quanto maior for o período de
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
enchimento do dreno, maiores as possibilidades de manter o crescimento de
pelo menos alguns drenos, e conseqüentemente, a produção.
A duração do dreno é também função da duração da fonte, pois não
adianta ter um dreno a ser enchido sem uma fonte de fotoassimilados. Quase
todo o carbono que vai para o dreno é proveniente da atividade fotossintética
durante o período de enchimento deste dreno, sendo que pouco carbono é
retranslocado, ao contrário do nitrogênio (NORMAN et al, 1995).
4.2.1 A força dos drenos
A força de um dreno é determinada pela intensa atividade metabólica
nessas áreas. Altas taxas de divisão e expansão celular, respiração e
biossíntese, conseqüentemente, precisam de uma grande quantidade de
assimilados.
Nessas regiões e em determinadas fases de desenvolvimento existem
reguladores do crescimento (hormônios) que afetam a força dos drenos e,
certamente, um dreno forte é uma região que possui altos níveis de certos
reguladores que são conhecidos por estimular a divisão e expansão celular.
Além disso, outro fator pode ter considerável importância na força do dreno: a
capacidade de mobilizar solutos e transportá-los para o local.
A partição de nutrientes feita através do floema segue um critério
relativamente simples: ela é feita sempre no sentido da fonte para o dreno,
sendo que os drenos mais “fortes” recebem mais nutrientes.
Segundo Floss (2006), quanto aos fatores que definem a força do
dreno, três são os principais:
1) Proximidade. Normalmente as fontes translocam nutrientes para os
drenos que estão mais próximos delas. Uma conseqüência prática disso é que
folhas que sombreiam outras folhas mais próximas dos drenos de interesse
devem ser eliminadas. Isso ocorre em videira, onde as folhas próximas aos
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
cachos são as responsáveis pela qualidade dos frutos. Como critério geral, as
folhas da porção superir da planta costumam translocar nutrientes para as
folhas novas e caules em crescimento e as folhas da porção basal tendem a
exportar para o sistema radicular.
2) Desenvolvimento. Durante a fase vegetativa os maiores drenos são
raízes
e
ápices
caulinares. Na
fase
reprodutiva
os frutos
se
tornam
dominantes.
3)
Conexão
Fontes
vascular.
translocam
assimilados
preferencialmente para drenos com os quais elas têm conexão vascular direta.
Nos drenos, os fotoassimilados podem ser usados para o crescimento,
como ocorre nos meristemas e nas folhas em desenvolvimento. Também
podem ser destinados ao armazenamento, na forma de amido, ou de sacarose
ou outros açucares que se acumulam no vacúolo. O tamanho e a atividade
metabólica do dreno determinam a sua força e são influenciados por fatores
hormonais.
As condições de luminosidade durante o crescimento também afetam a
partição
de
assimilados.
Assim,
em
plantas
que
crescem
sob
baixa
luminosidade, os órgãos da parte aérea são drenos preferenciais. O contrário
ocorre em plantas cultivadas sob alta irradiância, nas quais as raízes são
depósitos preferidos. A remoção dos drenos reprodutivos pode aumentar o
peso específico da folha, reduzindo concomitantemente a taxa de fotossíntese.
4.3 Ações de Fitormônios na fonte e no dreno
O metabolismo de carbono na fonte e no dreno, assim como o transporte
entre esses tecidos, é regulado por um controle de “feedback” pelos níveis dos
metabólitos intermediários e finais. O controle por “feedback” é feito
diretamente pelo metabólito a nível de transcrição de enzimas específicas do
seu metabolismo, ou mediado por fitormônios, os quais controlam essas
enzimas. Não havendo uso desses metabólitos ocorre uma diminuição de sua
formação (Pimentel, 1998).
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
O movimento de fotoassimilados do local de síntese, na fonte, para o
local de uso ou acumulação, no dreno, pode ser regulado em diversos pontos.
O gradiente de concentração entre a fonte e o dreno é geralmente aceito como
principal determinante da taxa de transporte e partição entre órgãos. Tais
processos também são controlados, em parte, por fitormônios que atuam
como mensageiros entre células, tecidos e órgãos, promovendo a ativação de
genes específicos ou processos metabólicos.
As auxinas, citocininas e giberelinas modulam o crescimento dos órgãos
da planta e, portanto, afetam a força de dreno de suas diferentes partes. Em
particular, altas concentrações de citocininas são encontradas em órgãos e
tecidos em crescimento ativo, como sementes e frutos em desenvolvimento e
ápices radiculares.
A citocinina é um hormônio vegetal que aumenta a força do dreno,
retardando a senscência. Em geral, os assimilados da folha fonte são
direcionados para os drenos fortes mais próximos, o que significa que as folhas
do terço superior direcionam os assimilados para os ápices e folhas novas em
desenvolvimento, enquanto que folhas de terço inferior direcionam seus
produtos de exportação para as raízes. Em culturas anuais, os drenos fortes,
como a semente em desenvolvimento, têm prioridade em relação aos drenos
vegetativos e, em conseqüência, o crescimento vegetativo cessa ou é
sensivelmente reduzido (Pimentel, 1998).
Os grãos têm uma concentração de fitormônios maior que outras partes
da planta. Os níveis de auxinas, citocininas, giberelinas e ácido abscísico (ABA)
variam distintamente durante o desenvolvimento do fruto e do grão. À
excessão de citocininas, a máxima concentração dos outros fitormônios
geralmente ocorre durante o período de máxima acumulação de matéria seca
no dreno. Por exemplo, o ABA é encontrado em altas concentrações no eixo
embrionário de soja, ou no embrião do milho, isto é, ABA é encontrado em
máximas concentrações no local de descarregamento do floema no dreno,
durante o período de rápido enchimento do grão. A ocorrência de auxinas em
sementes em desenvolvimento é similar ao ABA, pois a máxima concentração
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
somente é observada alguns dias antes do máximo de ABA. Da mesma forma,
giberelinas em altas concentrações são encontradas no endosperma de ervilha,
no momento de máxima elongação da vagem; assim como as citocininas,
atingem os maiores
teores no endosperma
de
grãos
de
cereais
em
desenvolvimento. Em trigo, há um aumento da concentração de citocininas 4 a
5 dias pós-antese (Pimentel, 1998).
Os fitormônios regulam as relações fonte/dreno, provavelmente em uma
ação conjunta, onde um fitormônio funciona como promotor de um evento e
outro fitormônio como inibidor deste evento. Assim, o vegetal tem como
controlar passo a passo todo o processo de assimilação, transporte e utilização
de carboidratos, fazendo uma sintonia fina, em função de variações ambientais
instantâneas, como teores de CO2 ou de H2O, para maximizar o processo de
produção vegetal. Provavelmente, nenhum desses processos é controlado por
um único fitormônio, mas sim pelo balanço destes. O estímulo floral, por
exemplo, acredita-se ser simultaneamente produzido em folhas diferentes, via
fitocromos, enviado e amplificado em reações de cascata em dois níveis, com a
cooperação entre microfunções intracelulares e macrofunções intercelulares.
Tal resposta é dependente também das mudanças ocorridas nas estruturas
existentes, e da atividade enzimática que manteve o crescimento vegetativo
(Pimentel, 1998).
O vegetal controla o movimento de fitormônios entre diferentes tecidos,
os quais regulam a absorção de água e nutrientes e o trânsito de assimilados
entre a fonte e o dreno, atuando ao nível da plasmalema e causando
modificações químico-estruturais em substâncias celulares. Apesar de já
termos conhecimento da ação hormonal sobre a fonte e o dreno, devemos
acautelar-nos na afirmação de que as relações fonte/dreno são controladas
exclusivamente pelos fitormônios. Estes estão envolvidos no controle dos
processos responsáveis pelo crescimento e desenvolvimento vegetal, mas
provavelmente agindo em conjunto com outros metabólitos, com a expressão
gênica de enzimas e estímulos exógenos e endógenos, que em conjunto,
produzirão a resposta do vegetal.
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
Tabela 1. Relação de hormônios, local de síntese e principais efeitos na planta
A) Auxinas
Provavelmente também as auxinas ou a toxina fúngica (fusicosina)
estimulam o carregamento e descarregamento do floema pelo controle da
atividade
da
ATPase
membranar, funcionando
como
um
regulador da
concentração de K+ no floema (JAEGHER & BOYER, 1990). As auxinas se
movem do grão de soja para as folhas próximas, em maiores concentrações do
meio para o fim do período de enchimento do grão. A remoção dos frutos
reduz o exudado de auxinas no pecíolo, indicando que o fruto é a maior fonte
dessas. Na fase vegetativa, as auxinas são exportadas do apex de caule,
inibindo o crescimento de gemas laterais, demonstrando dominância apical. Na
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
fase de frutificação de feijão, o crescimento de gemas vegetativas é inibido e,
provavelmente, a fonte dessa inibição sejam os grãos dos frutos dominantes.
Quando as sementes de frutos mais velhos são retiradas, as gemas axilares
retomam o crescimento. Frutos maiores exportam mais auxinas do que frutos
menores, mostrando sua maior habilidade para exercer a dominância sobre
outros órgãos.
B) Citocininas
Talvez os órgãos dominantes reprimam o crescimento de gemas laterais,
agindo como um dreno, ativado pelas auxinas, para acumular a citocinina
oriunda de raízes, impedindo o transporte dessas citocininas para as estruturas
subordinadas. Estudos mostram que o nível e a distribuição de citocininas são
controlados pelo apex vegetativo, que controla, inclusive, a síntese da forma
ativa das citocininas (Pimentel, 1998). Citocininas produzidas nas raízes são
transportadas via xilema para a parte aérea, retardando a senescência,
parecendo induzir o início do desenvolvimento reprodutivo, redirecionando o
fluxo de assimilados do apex para as gemas reprodutivas. Sob deficiência de
nitrogênio, menos citocininas são sintetizadas no apex radicular, e o que é
produzido, pouco exportado para a parte aérea, havendo então indução do
crescimento radicular em detrimento da parte aérea (Pimentel, 1998). A
aplicação de citocininas em folhas promove a atividade fotossintética, pelo
aumento do conteúdo foliar de clorofilas, acelerando o desenvolvimento de
cloroplastos ou modulando a atividade de enzimas fotossintéticas, retardando
assim a senescência. No dreno, as citocininas estimulam a formação de
amiloplastos para acúmulo de amido, reduzindo o aborto de grãos.
C) Giberelinas
A retirada de frutos de videira causa diminuição dos teores de giberelinas
nas folhas, sugerindo que frutos em desenvolvimento também exportam
giberelinas para as folhas próximas. A remoção de tais frutos diminui a
atividade fotossintética da folha fonte, provavelmente pela diminuição do
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
conteúdo de giberelinas nessa folha. Giberelinas aumentam a atividade da
sacarose-fosfato sintetase, estimulando a exportação de sacarose da folha,
sendo consideradas indutoras de desenvolvimento reprodutivo (Pimentel,
1998). Em ervilha, a giberelina estimula o efeito inibidor de auxinas quando as
duas são aplicadas juntas no apex vegetativo, provavelmente estimulando a
exportação da auxina no ápice dominante. Da mesma forma, há uma interação
entre giberelinas e auxinas estimulando a atividade da invertase ácida de
drenos em desenvolvimento, e a expansão dos entrenós de cana-de-açúcar. As
giberelinas
também
estão
envolvidas
na
dormência
e
iniciação
do
desenvolvimento de tubérculos de batata, associado aos níveis de inibidores,
de carboidratos disponíveis e dos fatores ambientais como luz, temperatura e
fotoperíodo.
D) Ácido abscísico
O movimento do ABA entre fonte e dreno parece ser dinâmico. ABA
move-se rapidamente de raízes e folhas maduras para todos os outros órgãos
vegetativos, cumulando-se, geralmente, em tecidos de dreno. Quando aplicado
em folhas fonte de plantas em frutificação, o ABA é exportado para os grãos
em
desenvolvimento,
enquanto
ao
ser
aplicado
sobre
o
grão
em
desenvolvimento, fica imóvel. Um dos efeitos mais conhecido do ABA é sobre o
controle estomático em folhas. Os drenos, principalmente os grãos, podem
regular a atividade da fonte, retirando este ABA, que se move para o dreno,
estimulando o descarregamento do floema e aumentando a atividade das
invertases. ABA move-se das folhas para as vagens em desenvolvimento (na
soja) e é também transportado, via floema, para as raízes, sendo reciclado de
volta para a parte aérea, via xilema (Pimentel, 1998).
5. Partição de fotoassimilados nas fases de desenvolvimento vegetal
Uma vez produzidos, os fotoassimilados são transportados para muitas
áreas da planta, podendo ser transformados em muitos compostos, alguns
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
deles estruturais, tais como celulose e hemicelulose, que fortalecem a
estrutura física das plantas e usualmente permanecem onde são sintetizados.
Compostos de armazenamento são extremamente importantes, pois mantêm o
crescimento e a constância de desenvolvimento apesar das flutuações
fotossintéticas,
o
movimento
desses
compostos
do
local
onde
foram
armazenados para locais onde poderão ser reutilizados é chamado de
remobilização (Floss, 2006).
Durante certos estádios de desenvolvimento, as plantas podem assimilar
mais compostos do que a real utilização, sendo o excesso distribuído para os
locais de armazenamento e em um dado momento esses compostos
armazenados são utilizados para suprir a necessidade de algum órgão,
exemplo no momento da frutificação, folhas em senescência os açúcares,
aminoácidos, fósforo, enxofre, nitrogênio e outros elementos móveis são
remobilizados e translocados para os drenos ativos (Floss, 2006).
A partição é definida como a distribuição diferencial de fotoassimilados
pelos diferentes órgãos da planta e, portanto, determina o modelo de
crescimento da planta, que pode ser balanceado entre parte aérea e sistema
radicular (Alexandrino et al., 2005).
5.1 Partição de fotoassimilados durante a fase vegetativa
Algumas plantas cultivadas, tais como a maioria das gramíneas, não tem
essencialmente crescimento caulinar durante o desenvolvimento vegetativo,
havendo uma partição favorável para folhas e raízes. Nas plantas anuais, o
principio operativo consiste em usar, primeiramente, a maior proporção
possível dos fotoassimilados para formação de folhas. Estas, então, participam
na produção e aumentam a absorção da planta. Enquanto esses órgãos
fotossinteticamente ativos vão se desenvolvendo preferencialmente, a massa
das partes que apenas respira permanece pequena. Após a expanção total e
sob condições ambientais favoráveis de fotossíntese, as folhas podem
exportar, em média, 60% de seus fotoassimilados para outras áreas da planta,
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
mas com o envelhecimento das folhas e a posterior senescência, esse processo
diminui
até
cessar,
nessa
condição
a
folha
também
não
importada
fotoasssimilados, a respiração celular é grandemente reduzida ocorrendo
apenas para que a folha sobreviva (Floss, 2006).
5.2 Partição de fotoassimilados durante a fase reprodutiva
Órgãos como flores, frutos e sementes são considerados em algumas
culturas o rendimento econômico, tendo sido selecionados muitas vezes para
aumentar a partição entre o total de MS e as partes reprodutivas. Após o
florescimento o dreno reprodutivo torna-se extremamente forte, limitando a
partição de fotoassimilados entre a folha adicional, o caule e o crescimento de
raízes, sendo que na maioria dos casos cessa o crescimento, mas há casos de
crescimento indeterminado em que pode se ter o crescimento vegetativo e
reprodutivo ao mesmo tempo (Floss, 2006).
Na fase de floração de plantas anuais, o sistema de distribuição passa a
favorecer os órgãos reprodutores, que recebem uma tão grande que todas as
outras partes da planta mudam consideravelmente. A maior mudança ocorre
na fração da massa total (folhas), que passa, em média de 45% durante a fase
de elongação, para 15% na época em que os frutos estão maduros.
5.3 Partição de fotoassimilados durante o enchimento de grãos.
Os fotoassimilados depositados no grão podem ser oriundos de três
fontes principais: da fotossíntese das folhas ativas, da fotossíntese das demais
partes verdes e
da remobilização de fotoassimilados depositados em outros
órgãos da planta.
Objetivando estudar o efeito de diferentes épocas de semeadura na taxa
e duração do período de enchimento de grãos e de que modo esses fatores
determinam o peso final dos grãos de híbridos do milho (Zea mays L.), Didonet
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
et al. (2001) citado por Floss (2006) concluíram que a taxa efetiva do
enchimento dos grãos foi diretamente dependente da temperatura média do
ar, tendo sido maior nas semeaduras mais precoces e resultando em grãos
mais pesados do que os obtidos em semeaduras mais tardias em relação á
semeadura mais tardia. Concluído que o principal fator limitante do rendimento
de grãos em semeadura tardia é a fonte (produção de fotoassimilados), não a
capacidade dos grãos em acumulá-los.
6. A partição de fotoassimilados e o manejo de pastagens
Alexandrino at al., (2005), estudando a distribuição dos fotoassimilados
em plantas de Panicum maximum cv. Mombaça e a dinâmica da relação fonte
dreno conforme a inserção da folha completamente expandida, de diferentes
tipos de perfilhos de plantas intactas ou submetidas a desfolhação, através da
utilização de carbono marcado.
O nível de inserção da folha foi classificado em folha de topo que era
aquela recém expandida (mais jovem) do perfilho, e a da base, a que estava
no segundo nível de inserção abaixo da folha do topo, analisando no perfilho
principal, 1º e 3º perfilhos primários (Tabela 2).
Tabela 2. Tratamento experimentais
A desfolhação, quando realizada, foi praticada pela remoção das lâminas
foliares da planta no nível da lígula, minutos antes da exposição ao
14
CO2, em
todas as folhas da planta, inclusive a folha emergente, com exceção da lamina
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
a ser exposta ao carbono marcado. No momento em que aplicou o
14
C a
gramínea encontrava-se em pleno processo de perfilhamento, cujo perfilho
principal estava com 48 dias. As folhas eram expostas por 30 minutos ao
carbono marcado, após esse tempo as plantas ficaram na casa de vegetação
por 24 horas e decorrido esse tempo elas foram individualmente colhidas e
fracionadas nos seguintes componentes: Perfilho principal e sua raiz; Primeiro
perfilho primário e sua raiz; Segundo perfilho primário e sua raiz; Terceiro
perfilho primário e sua raiz; Quarto perfilho primário; e Perfilhos secundários
(originados na gema axilar da folha de perfilhos primários). Além disso, o
perfilho cuja lâmina foliar foi exposta foi desmembrado em: Meristema
terminal (meristema apical + folhas em expansão e a folha emergente); Folha
adulta exposta ao
14
CO2; Demais folhas adultas, completamente expandidas
verdes (bainha + lâmina ou apenas bainha das plantas desfolhadas); e Base
do perfilho (segmento entre a raiz e o meristema apical).
Os tratamentos experimentais, resultantes da combinação dos níveis de
inserção da folha, da hierarquia dos perfilhos e da desfolhação, não tiveram
efeito (P>0,05) sobre o modelo de distribuição percentual dos assimilados
recuperados entre a parte aérea e o sistema radicular, seja do perfilho da folha
exposta seja dos outros perfilhos da planta, após 24 horas de exposição, a
partição de
14
C pela planta inteira alcançou valores médios de 82,5% para a
parte aérea e 17,5% para o sistema radicular (Figura 6). O autor relatou que
os valores de partição de fotoassimilados para a raiz podem ser bem mais
superiores aos encontrados nessa pesquisa, mas devido ao estágio de pleno
perfilhamento do capim-mombaça onde os novos perfilhos constituem-se no
principal dreno, os resultados são condizentes com tal fase.
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
Figura 6. Distribuição dos assimilados entre o perfilho portador da folha
exposta e os demais perfilhos da planta, em função dos tratamentos
experimentais, conforme Tabela 2. Valores acima e abaixo da abscissa
correspondem, respectivamente, à parte aérea e ao sistema radicular
dos perfilhos.
Ressalta-se a tendência de mais alto percentual de exportação de
assimilados para o sistema radicular pelas folhas dos perfilhos primários,
sobretudo de suas folhas da base. Estas observações são consistentes com
relatos
de
que
preferencialmente
folhas
para
o
ontogenicamente
sistema
radicular.
mais
Maior
velhas
quantidade
exportam
14
C
foi
recuperada no sistema radicular de perfilhos mais jovens: perfilho principal
15,3%; 1º perfilho primário 17,7%; e 3º perfilho primário 21,6%, o que
provavelmente esteja relacionado à prioridade para o desenvolvimento do
sistema radicular daqueles perfilhos. Tendência de substancial translocação de
assimilados da folha do topo do 3º perfilho primário para o sistema radicular
foi observada sob condições de desfolha da planta, demonstrando que o
sistema radicular é um forte dreno em perfilhos novos.
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
Em plantas que não perfilham (milho) ou perfilham pouco (sorgo), a
translocação de assimilados da folha da base se faz preferencialmente para as
raízes, enquanto, em plantas com intenso perfilhamento (trigo e azevém
perene), a prioridade cabe aos perfilhos (Ryle & Powell (1974) citado por
Alexandrino et al. (2005)). Neste estudo, a prioridade de translocação das
folhas da base para os perfilhos foi condizente com os relatos da literatura
quanto ao padrão observado em trigo e azevém. Isso decorre possivelmente
da fase de desenvolvimento vegetativo do capim-mombaça, que, apresentando
curto pseudo-colmo, propiciou igual proximidade dos diferentes drenos em
relação à fonte de assimilados. Entretanto, maior percentual de radioatividade
na parte aérea correspondeu sempre ao perfilho portador da folha exposta,
relativamente àquela encontrada na parte aérea de qualquer um dos demais
perfilhos, independentemente de sua hierarquia (Figura 7). Entretanto, houve
tendência de perfilhos mais jovens exportarem maior quantidade de
sistema radicular.
14
C para o
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
Figura 7. Distribuição dos assimilados pelos diferentes perfilhos da planta, em
função dos tratamentos experimentais, conforme Tabela 2. Valores
acima e abaixo da abscissa correspondem, respectivamente, à parte
aérea e ao sistema radicular dos perfilhos.
A exportação de assimilados para os diferentes órgãos da parte aérea do
perfilho portador da folha exposta não variou (P>0,05) com os tratamentos
experimentais, mas algumas tendências foram observadas. Assim, as folhas do
topo tenderam a translocar mais assimilados para o meristema terminal que as
folhas da base, principalmente para condição do perfilho principal, com valores
de 15 e 10,8%, respectivamente.
Independentemente do tratamento, a translocação de assimilados para a
base (base da planta + pseudocolmo) atingiu valores de 4,27 a 7,80%, sempre
inferior àquela translocada para outros componentes do perfilho da folha
exposta. Avaliando-se individualmente todos os componentes da planta,
conclui-se que o meristema terminal comportou-se como o dreno de maior
força da planta de capim-mombaça quando a folha exposta foi a do topo,
independentemente do tipo de perfilho ou da presença de desfolhação.
A translocação de assimilados em plantas de capim-mombaça em fase de
intenso perfilhamento foi mais acentuada para a parte aérea que para o
sistema radicular. Independentemente da hierarquia do perfilho, do nível de
inserção da folha e da desfolhação, o perfilho portador da folha exposta ao
14
CO2 reteve aproximadamente 50% dos assimilados encontrados na parte
aérea da planta.
Lupinacci (2002), quantificando os teores e estoques de compostos de
reserva (carboidratos não estruturais) em pastos da Brachiaria brizantha cv.
Marandu, submetida a quatro intensidades de alturas 10, 20, 30 e 40 cm por
pastejo de bovinos de corte, no período de agosto de 2001 a fevereiro de
2002. Observou que os teores de carboidratos não estruturais (CNE) possuíam
os menores teores na menor altura de pastejo a 10 cm e o maior valor nos
pastos mantidos a 40 cm, com 40,6 e 118,4 g kg-1, respectivamente durante a
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
fase vegetativa. Considerando-se que a altura de pasto de 10 cm apresentou
valores de taxa de acumulo próximos aos observados para os demais
tratamentos durante esse período, pode-se inferir que os menores teores de
CNE desses pastos em relação às alturas de 20, 30 e 40 cm indicam que a
quantidade de assimilados alocados
para os tecidos de reserva foi menor,
provavelmente como conseqüência da priorização de produção de tecidos
fotossintetizantes.
Já em janeiro e fevereiro, meses em que a planta encontrava-se em
estádio reprodutivo, notou-se uma redução nos teores de CNE em todos os
tratamentos, tal fato pode ser compreendido se for considerado que a
inflorescência deve ter atuado como maior dreno metabólico durante a fase
reprodutiva e, portanto, a produção de fotoassimilados não deve ter sido
suficientemente para assegurar os processos envolvidos com a formação de
inflorescência e manutenção dos processos respiratórios e de síntese de
tecidos, principalmente nos pastos mantidos nas menores alturas. Como
conseqüência, as plantas precisaram lançar mão dos compostos armazenados
nos órgãos de reservas, de modo que quanto mais assimilados produzido pela
planta menor deve ter sido a quantidade de reserva mobilizada. Pastos
mantidos a 10 cm corresponderam a uma condição de maior estresse para a
planta forrageira, condição essa justificada pelos menores teores apresentados
durante a fase de desenvolvimento.
Com relação ao órgão de armazenamento dos CNE, foi observado que
durante o estádio vegetativo que as raízes apresentaram os maiores teores
desses compostos. Á medida que a planta passa para o estádio reprodutivo,
houve uma inversão na ordem de grandeza dos teores em relação aos órgãos
de armazenamento e modo que os maiores teores de CNE foram observados
na parte aérea.
O padrão de comportamento dos estoques de CNE ao longo do período
experimental foi semelhante ao descrito para os teores desses compostos onde
os maiores estoques foram observados nos pastos mantidos mais altos e
durante a fase vegetativa e que com a chegada do estádio reprodutivo houve
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
uma redução do estoque de CNE em todos os tratamentos. Adicionalmente a
redução dos estoques de CNE foi superior na parte aérea em relação ao
sistema radicular, chegando a atingir 49,38 e 63,70% em dezembro e
fevereiro, respectivamente, em relação aos maiores estoques observados
durante o período experimental. Esse comportamento pode ser justificado pela
maior proximidade da parte aérea aos principais drenos da planta.
Com relação ao órgão de armazenamento dos CNE, foi notado que a
parte
aérea
apresentou
consistentemente
os
maiores
estoques
desses
compostos em relação ao sistema radicular ao longo do período experimental,
o que foi devido fundamentalmente á maior massa da parte aérea em
comparação ao sistema radicular.
Teixeira
et
al.
(2005),
avaliando
o
destino
dos
fotoassimilados
sintetizados em folhas de diferentes níveis de inserção no perfilho principal de
Panicum maximum cv. Mombaça, em dois estádios de desenvolvimento, em
diferentes tempos de colheita, após a exposição da folha ao
14
CO2. Os
tratamentos resultaram de um arranjo fatorial 2x2x3 - dois níveis de inserção
de folha no perfilho principal (topo e base), dois estádios de crescimento e três
momentos de colheita (três, oito, e 24 h) após a exposição da folha ao
14
CO2.
Os estádios de crescimento corresponderam: (i) ao momento em que o
perfilho principal alcançou valores próximos a nove folhas completamente
expandidas e quatro a cinco perfilhos; (ii) dez folhas completamente
expandidas e oito a nove perfilhos, respectivamente. Depois de serem
colhidas, cada planta teve seu perfilho principal separado da raiz e dos demais
perfilhos.
Posteriormente,
o
perfilho
principal
foi
dissecado
em:
folha
completamente expandida exposta (bainha mais lâmina), folhas adultas não
expostas (bainha mais lâmina), meristema terminal (folha emergente mais
folha em expansão mais meristema apical), base e raiz. Os demais perfilhos
foram desmembrados em perfilhos primários e secundários surgidos no
primeiro e segundo estádios.
As maiores biomassas da planta do capim-Mombaça corresponderam aos
perfilhos primários e secundários, ao sistema radicular e às folhas adultas do
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
perfilho principal. Estas frações constituíram seus principais drenos, onde se
acumularam as maiores quantidades exportadas de fotoassimilados-14C. Dessa
forma, o tamanho do órgão foi fator determinante da sua força dreno,
provavelmente, porque as plantas se apresentavam ainda muito jovens, nos
dois estádios estudados, e estavam com seus órgãos em pleno crescimento e
grande atividade metabólica. Carvalho (2002), trabalhando com os cultivares
Mombaça e Tanzânia (Panicum maximum), observou que a translocação de
assimilados do perfilho principal para o perfilho primário mais jovem foi menor
(6,5%) que do perfilho primário para o perfilho principal (14%) em razão da
maior massa do perfilho principal. A distância e a maior força dreno do perfilho
principal (o maior perfilho) teriam sido os principais fatores a determinar a
maior quantidade de fotoassimilados-14C exportada para o mesmo, em relação
às demais categorias de perfilhos. A folha do topo, por apresentar maior
massa, reteve mais fotoassimilados que a da base (Tabela 3). No entanto,
essas folhas expostas ao
14
CO2 diferiam não apenas quanto à massa e ao nível
de inserção no perfilho, mas também à idade e ao ambiente em que se
encontravam. As folhas da base, mais velhas e desenvolvidas sob condições de
baixa luminosidade, têm menor capacidade fotossintética.
As folhas do topo e da base diferem em idade e proximidade dos drenos
da planta. A folha do topo, mais jovem, encontra-se mais próxima do
meristema apical, ao passo que a da base, mais velha, localiza-se mais
próxima da raiz e dos perfilhos, o que lhes confere diferente padrão de
distribuição de assimilados, que este é resultado da força e da distância dos
drenos à fonte. Menor percentagem de radioatividade foi encontrada nas folhas
adultas não expostas e nos perfilhos primários e secundários do primeiro
estádio, quando a folha do topo foi exposta. A quantidade de fotoassimilados
exportada para a raiz, segundo maior dreno da planta, tendeu a ser maior
quando da exposição da folha da base, porém, sem significância (P>0,05) para
a diferença observada. A quantidade de fotoassimilados presente no perfilho
principal alcançou percentual mais alto (56,1%), quando da exposição da folha
do topo, que a observada quando da exposição da folha da base, que foi de
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
53,2% (P<0,05%). Esse fato reflete a menor retenção de fotoassimilados na
folha exposta da base e sua maior exportação de fotoassimilados para os
perfilhos surgidos na primeira fase, maiores drenos da planta, ficando menos
fotoassimilados no perfilho principal.
Tabela 3. Percentagem de fotoassimilados
14
C em cada órgão do perfilho
principal e em outros perfilhos, em função do nível de inserção da
folha exposta ao
14
CO2
A quantidade de fotoassimilados-14C exportada do perfilho principal para
os perfilhos primários e secundários também foi elevada, provavelmente em
razão da condição de baixa luminosidade que antecedeu a exposição ao
14
CO2
nos dois estádios de desenvolvimento, o que pode ter elevado ainda mais a
dependência de assimilados por parte do maior dreno da planta (os perfilhos).
7. Considerações Finais
A dinâmica da partição de fotoassimilados e da relação fonte dreno é
influenciada
por uma série
de
fatores
ambientais, de
manejo
e das
características da planta. Sob condição de pastejo a desfolhação é o fator de
manejo que mais
interfere na partição de assimilados, cujo efeito no
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
crescimento da planta durante o período de rebrotação é gradativo e
dependente da intensidade de remoção da área foliar e da decapitação dos
perfilhos. Essa interferência pode ser positiva ou negativa, conforme a
habilidade da planta em restabelecer uma nova partição de assimilados das
novas fontes para os novos drenos.
Portanto conhecer a habilidade de planta na partição e alocação de
assimilados é de fundamental importância para se adotar manejos em que se
tenha o maior rendimento produtivo (exemplo kg de carne produzidos por
área), sem comprometer a persistência e produtividade da planta.
8. Referêcias Bibliográficas
ALEXANDRINO, E. et al. Distribuição de fotoassimilados em plantas de Panicum maximum cv.
Mombaça. Revista Brasileira de Zootecnia. v.34, n. 5, p. 1449-1458, 2005.
CARVALHO, D.D. Leaf morphogenesis and tillering behaviour in single plants and
simulated swards of guinea grass (Panicum maximum jacq) cultivars. New
Zealand: University Palmerston 2002. 186p. Tese (PhD Plant Sience) - University
Palmerston, 2002.
DAVIES, W. J., MANSFIELD, T. A. & HETHERING, A. M. 1990. Sensing of soil water status and
the regulation of plant growth and development. Plant Cell and Environment, v.13,
p.709-719.
FAGERIA, N. K. 1992. Maximizing crop yields. Marcel Dekker, Inc. New York.
FLOSS, E. L. Fisiologia das palntas cultivadas. 3 ed. Passo Fundo – RS, Editora: UPF, 2006.
751p.
HENDRIX, J. E. 1994. Assimilate transport and partitioning. In: Handbook of plant and crop
physiology. PESSARAKLI, M. (ed.) Marcel Dekker. New York. pp. 357-385.
HSIAO, T. C. 1990. Measurements of plant water status. In: Irrigation of agricultural crops.
STEWART, B. A. & NIELSEN, D. R. (eds.) American Society of Agronomy. Madison. pp.
244-280
JAEGHER, G. de & BOYER, N. 1990. On the role of membranes and calcium in signal perception
and transduction in thigmomorphogenesis of Bryonia dioica. In: Intra-and intercellular
communications in plants. MILLET, B. & GREPPIN, H. (eds.). INRA publ. Paris. pp. 2939.
LEMAIRE, G.; AGNUSDEI, M. Leaf tissue turnover and efficiency of herbage utilization. In:
GRASSLAND AND ECOPHYSIOLOGY AND ECOLOGY, 1999, Curitiba. Anais…Curitiba:
Universidade Federal do Paraná, 1999. p.165-186.
LUPINACCI, A. V. Reservas orgânicas, índice de área foliar e produção de forragem em
Brachiaria brizantha cv. Marandu submetida a intensidades de pastejo por bovinos de
corte. Piracicaba, 2002. 160p. Dissertação (Mestrado). Escola Superior de Agricultura
“Luiz de Queiroz”, Universidade de São Paulo.
MARENCO, R. A.; LOPES, N. F. Fisiologia Vegetal. Fotossíntese, respiração, relações hídricas
e nutrição mineral. Viçosa, ed.3. Editora: UFV., 2005. 432p.
MARSCHNER, H. 1995. Mineral nutrition of higher plants. Academic Press. San Diego.
Pereira, L.E.T. e Paiva, A.J. O manejo de pastagens e as relações fonte/dreno na planta
forrageira. PUBVET, V.2, N.22, Art#244, Jun1, 2008.
MORAES, A.; MARASCHIN, G. E.; NABINGER, C. Pastagens nos ecossistemas de clima subtropical: pesquisas para o desenvolvimento sustentável. In: SIMPÓSIO SOBRE
PASTAGENS NOS ECOSSISTEMAS BRASILEIROS, 32., 1995, Brasília. Anais... Brasília:
Sociedade Brasileira de Zootecnia, 1995. p.147-200.
NORMAN, M. J. T., PEARSON, C. J. & SEARLE, P. G. E. 1995. The ecology of tropical food
crops. Cambridge University Press. Cambridge PATRICK, J. W. Phloem unloading: sieve
element unloading and post-sieve element transport. Annual Review of Plant
Physiology and Plant Molecular Biology. v.48, p. 191-222, 1997.
PELL, E. J. & DANN, M. S. 1990. Multiple stress-induced foliar senescence and implications for
whole-plant longevity. In: Environmental injury to plants. KATTERMAN, F. (ed.).
Academic Press. San Diego. pp. 189-205.
PIMENTEL, C. Metabolismo do carbono na agricultura tropical. UFRRJ. Seropédica: Edur,
1998. 150p.
PUGNAIRE, F. I., ENDOLZ L. S. & PARDOS J. 1994. Constrains by water stress on plant growth.
In: Handbook of plant and crop stress. PESSARAKLI, M. (ed.) Marcel Dekker Inc. New
York. pp. 247-260.
SCHULTZ, A. Phloem transport and differential unloading in pea seedlings after source and sink
manipulations. Planta, v.192, p.239-248, 1994.
SINCLAIR, T. R. & HORIE, T. 1989. Crop physiology & metabolism. Leaf nitrogen,
photosynthesis, and crop radiation use efficiency: A Review. Crop Science, 29: 90-98.
TAIZ, L. & ZEIGER, E. Plant physiology. California, The Benjamin/Cummigs Publishing Co.,
1998, 559 p.
TAIZ, L.; ZIEGER, E. Fisiologia Vegetal. Editora: Artmed S.A., ed.3, Porto Alegre, 2004.
719p.
TEIXEIRA, A. C. B. et al. Distribuição de fotoassimilados de folhas do topo e da base do capimMombaça (Panicum maximum cv. Mombaça) em dois estádio de desenvolvimento.
Revista Brasileira de Zootecnia. v. 34, n. 2 p. 479-488, 2005.
TURGEON, R. Symplastic phloem loading and the sink-source transition in leaves: a model. In
BONNEMAIN, J. L.; DELROT, S.; LUCAS, W. J.; DAINTY, J. (Eds). Recent advances in
phloem transport and assimilate compartmentation. Nantes, Ouest Editions.
1991. p. 18-22.
VIERSTRA, R. D. 1996. Proteolysis in plants: mechanisms and functions. Plant Molecular
Biology, 32: 275-302.
WARDLAW, I. F. 1990. The control of carbon partioning in plants. The New Phytologist, 116 :
341-381.
http://www.ufmt.br/bionet/conteudos/15.01.05/fotossintese.htm. Fotossíntese. Capturado
em 11 de Maio de 2007.