E CINETINA (KIN) NO")
UNIVERSIDADE DO EXTREMO SUL CATARINENSE – UNESC
CIÊNCIAS BIOLÓGICAS – BACHARELADO
DEISE BACKES
EFEITOS DO ÁCIDO INDOLILACÉTICO (AIA) E CINETINA (KIN) NO
ENRAIZAMENTO DE ESTACAS DE Maytenus ilicifolia Mart. ex
Reiss. e Casearia sylvestris (Sw.)
CRICIÚMA, NOVEMBRO DE 2010
1
DEISE BACKES
EFEITOS DO ÁCIDO INDOLILACÉTICO (AIA) E CINETINA (KIN) NO
ENRAIZAMENTO DE ESTACAS DE Maytenus ilicifolia Mart. ex
Reiss. e Casearia sylvestris (Sw.)
Trabalho
de
conclusão
de
curso,
apresentado para obtenção do grau de
Biólogo Bacharel no curso de Ciências
Biológicas da Universidade do Extremo Sul
Catarinense, UNESC.
Orientador: Prof. MSc. Roberto Recart dos
Santos.
CRICIÚMA, NOVEMBRO DE 2010
2
DEISE BACKES
EFEITOS DO ÁCIDO INDOLILACÉTICO (AIA) E CINETINA (KIN) NO
ENRAIZAMENTO DE ESTACAS DE Maytenus ilicifolia Mart. ex
Reiss. e Casearia sylvestris (Sw.)
Trabalho de Conclusão de Curso aprovado
pela Banca Examinadora para obtenção do
Grau de Biólogo Bacharel no Curso de
Ciências
Biológicas
da
Universidade
do
Extremo Sul Catarinense, UNESC, com Linha
de Pesquisa em Manejo de Recursos Naturais.
Criciúma, 26 de novembro de 2010.
BANCA EXAMINADORA
____________________________________________________
Prof. MSc. Roberto Recart dos Santos – Eng. Agrônomo - Orientador - (UNESC)
__________________________________
Prof. MSc. Marcos Back – Eng. Agrônomo - (UNESC)
___________________________________
Prof. Dr. Jairo José Zocche - Biólogo - (UNESC)
3
AGRADECIMENTOS
Em primeiro lugar agradeço aos meus pais, Günter Backes e Clara de
Godoi Backes por sempre acreditaram em mim e me forneceram suporte e conforto
ao longo dessa minha caminhada. Agradeço também aos meus colegas e amigos,
que se fizeram sempre presentes, assim como de modo geral aos meus professores.
Agradeço ao meu orientador Prof. Msc. Roberto Recart dos Santos, pela
sua disponibilidade e auxílio, bem como a equipe do Horto Florestal – UNESC em
especial Alencar Semler.
Agradeço também a Marcos Back e Jairo Zocche que aceitaram fazer
parte da banca examinadora.
E por último, mas não menos importante, agradeço a Deus, que sempre
esteve presente fornecendo-me paciência, força e compreensão nos momentos
mais difíceis desta minha jornada de 4 anos como estudante de Ciências Biológicas
– Bacharelado, UNESC.
4
“O que ocorre com a terra recairá sobre os
filhos da terra. O homem não teceu o tecido
da vida: ele é simplesmente um de seus fios.
Tudo o que fizer ao tecido, fará a si mesmo”.
Carta
do
Chefe
indígena
Seatle
ao
Presidente dos Estados Unidos da América
(1854).
5
RESUMO
Maytenus ilicifolia Mart. ex Reiss. e Casearia sylvestris (Sw.) são espécies
medicinais com ampla utilização no mercado de fitoterápicos e uso popular, porém o
extrativismo tem comprometido o estoque natural das espécies assim como a baixa
taxa de germinação têm prejudicado a regeneração das mesmas. A utilização de
técnicas de propagação vegetativa, como a estaquia, possibilita a produção de um
maior número de mudas em um curto período de tempo. O objetivo deste estudo foi
avaliar a eficiência dos hormônios vegetais Ácido indolilacético (AIA) e Cinetina
(KIN), no enraizamento de estacas de Maytenus ilicifolia Mart. ex Reiss. e Casearia
sylvestris (Sw.). Foram confeccionadas estacas herbáceas e lenhosas, com 8,0 cm
de comprimento, com base cortada em bisel e com lesões longitudinais na base,
com imersão de 15 segundos nos tratamentos correspondentes: AIA (0; 125; 250;
500; 1.000; 2.000 ppm). Na sequência, um segundo grupo de estacas foi submetido
às mesmas concentrações de AIA mais 25 ppm de KIN. Após a aplicação foram
colocadas para enraizar em meio contendo cinza de casca de arroz e substrato
comercial, ambos autoclavados. O delineamento foi totalmente casualizado com seis
tratamentos distintos, para cada 12 estacas. Foram avaliadas estacas vivas, com
calos e com brotação após 90 dias de tratamento. Os resultados foram submetidos à
análise pelo programa estatístico Qui-Quadrado, para comparar se houve diferença
significativa entre as diferentes variáveis. Verificou-se que a aplicação de KIN pode
favorecer o enraizamento das estacas assim como menores concentrações de AIA
mostram-se mais eficientes do que maiores dosagens, mesmo não sendo
estatisticamente significativos. Outros estudos devem ser conduzidos com estacas
herbáceas e lenhosas de ambas espécies com tempo maior de acondicionamento
no substrato.
Palavras-chave: Propagação vegetativa. Hormônios vegetais. Maytenus ilicifolia
Mart. ex Reiss.. Casearia sylvestris (Sw.).
6
LISTA DE ILUSTRAÇÕES
Figura 1: Estrutura química do AIA.
Figura 2: Estrutura química do KIN.
Figura 3: Imagem satélite do Fragmento de floresta Mata Atlântica, Horto – UNESC.
Figura 4: Fragmento de Floresta Ombrófila Mista Montana, São Joaquim – Serra
Catarinense, 2010.
Figura 5: Fragmento de Mata Ciliar, Bom Jardim - Serra Catarinense, 2010.
Figura 6: Maytenus ilicifolia Mart. ex Reiss., aspecto geral da planta em São Joaquim
– Serra Catarinense, 2010.
Figura 7: Detalhe do ramo, folhas e frutos do tipo cápsula de Maytenus ilicifolia Mart.
ex Reiss..
Figura 8: Aspecto geral de Casearia sylvestris (Sw.).
Figura 9: Ramos de C. sylvestris (Sw.) em florescimento.
Figura 10: Estaca herbácea de Maytenus ilicifolia Mart. ex Reiss.
Figura 11: Estacas emersas em água destilada.
Figura 12: Imagem satélite EPAGRI, Urussanga –SC.
Figura 13: Acondicionamento em tubetes de Maytenus ilicifolia Mart. ex Reiss..
Figura 14: Estacas de M. ilicifolia Mart ex Reiss. A) Estaca enraizada do primeiro
experimento (AIA) na concentração de 250 ppm. B) Estaca com formação de calos
na concentração de 25 – 1000 ppm.
Figura 15: Estacas de M. ilicifolia Mart ex Reiss. A) Estaca morta do primeiro
experimento (AIA) na concentração de 2.000 pmm. B) Estaca enraizada na
concentração de 25 – 250 ppm.
Figura 16: Estacas de Casearia sylvestris (Sw.) enraizadas nas concentrações 125
(AIA) e 25 – 0 (KIN/AIA) respectivamente.
7
LISTA DE TABELAS
Tabela 1 - Classificação Taxonômica Segundo Engler e Cronquist.
Tabela 2: Comparação estatística entre as variáveis usando Qui-Quadrado, do
primeiro experimento (AIA) com estacas herbáceas de inverno de Maytenus ilicifolia
Mart. ex Reiss..
Tabela 3: Comparação estatística entre as variáveis usando Qui-Quadrado, do
segundo experimento (KIN/AIA) com estacas herbáceas de inverno de Maytenus
ilicifolia Mart. ex Reiss..
Tabela 4: Comparação estatística entre as variáveis usando Qui-Quadrado, do
primeiro experimento (AIA) de Maytenus ilicifolia Mart. ex Reiss..
Tabela 5: Comparação estatística entre as variáveis usando Qui-Quadrado, do
segundo experimento (KIN/AIA) de Maytenus ilicifolia Mart. ex Reiss..
Tabela 6: Comparação estatística entre as variáveis usando Qui-Quadrado, do
primeiro experimento (AIA) de Casearia sylvestris (Sw.).
Tabela 7: Comparação estatística entre as variáveis usando Qui-Quadrado, do
segundo experimento (KIN/AIA) de Casearia sylvestris (Sw.).
Tabela 8: Comparação estatística entre as variáveis usando Qui-Quadrado, do
primeiro experimento (AIA) de Maytenus ilicifolia Mart. ex Reiss..
Tabela 9: Comparação estatística entre as variáveis usando Qui-Quadrado, do
segundo experimento (KIN/AIA) de Maytenus ilicifolia Mart. ex Reiss..
Tabela 10: Comparação estatística entre as variáveis usando Qui-Quadrado, do
primeiro experimento (AIA) de Casearia sylvestris (Sw.).
Tabela 11: Comparação estatística entre as variáveis usando Qui-Quadrado, do
segundo experimento (KIN/AIA) de Casearia sylvestris (Sw.).
8
SUMÁRIO
1 INTRODUÇÃO ......................................................................................................09
2 OBJETIVOS ..........................................................................................................12
2.1 Objetivo geral ...................................................................................................12
2.2 Objetivos específicos .......................................................................................12
3 REFERENCIAL TEÓRICO ....................................................................................12
3.1 Plantas medicinais ...........................................................................................12
3.2 Propagação vegetativa ....................................................................................14
3.3 Fatores que afetam o enraizamento na estaquia ...........................................15
3.4 Hormônios vegetais e suas aplicabilidades ..................................................16
3.4.1 Ácido indolilacético (AIA) .............................................................................18
3.4.2 Cinetina (KIN) .................................................................................................19
4 MATERIAIS E MÉTODOS ....................................................................................20
4.1 Descrição da área de estudo ...........................................................................20
4.2 Maytenus ilicifolia Mart. Ex Reiss. ..................................................................23
4.3 Casearia sylvestris (Sw.) ..................................................................................27
4.4 Metodologia ......................................................................................................30
4.4.1 Coleta das estacas ........................................................................................30
4.4.2 Preparação das estacas em laboratório ......................................................30
4.4.3 Acondicionamento das estacas do primeiro experimento - EPAGRI ........32
4.4.4 Acondicionamento das estacas do segundo experimento – Horto Florestal - UNESC .............................................................................................................34
4.4.5 Variáveis analisadas .....................................................................................34
4.3.6 Análise estatística .........................................................................................34
5 RESULTADOS E DISCUSSÃO ............................................................................35
5.1 Estacas herbáceas de verão ...........................................................................35
5.1.1 Estacas herbáceas de inverno .....................................................................37
5.2 Estacas lenhosas .............................................................................................41
6 CONCLUSÃO .......................................................................................................46
REFERÊNCIAS.........................................................................................................47
9
1 INTRODUÇÃO
O uso de plantas medicinais pela população mundial tem sido muito
significativo nos últimos tempos. Dados da Organização Mundial de Saúde (OMS)
mostram que cerca de 80% da população mundial fez uso de algum tipo de erva na
busca de alívio de alguma sintomatologia dolorosa ou desagradável. Desse total,
pelo menos 30% deu-se por indicação médica. São muitos os fatores que vêm
colaborando no desenvolvimento de práticas de saúde que incluam plantas
medicinais, principalmente econômicos e sociais (MARTINS, 2000), proporcionando
atualmente ao mercado mundial de fitoterápicos um montante de 20 a 40 milhões de
dólares por ano (MARTINS et al., 1995).
Segundo a ONU/OMS, existem cerca de 30% de preparações
farmacêuticas que são obtidas direta ou indiretamente de plantas, sendo que no
Brasil passa a ser 40% de medicamentos farmacêuticos que têm princípios ativos
retirados desses vegetais (FRANCO, 2001).
A utilização das plantas medicinais pelo povo faz parte da cultura, como
resultado das experiências de gerações passadas, que foram transmitidas por meio
de aprendizagens. “O resultado empírico das virtudes curativas dos vegetais
remonta a épocas antigas, quando o homem primitivo escolhia nos prados e selvas,
guiado pelo instinto, a erva para acalmar a dor e curar o próprio mal” (PANIZZA,
1997).
O Brasil, apesar de não possuir uma medicina tradicional propriamente
dita, isto é, caracterizada pela transmissão escrita dos conhecimentos, desenvolveu
uma Medicina Popular, calcada nas informações dos índios, europeus e negros,
principais formadores da etnia brasileira, e na influência de outras populações
imigrantes. Tal conhecimento vem sendo perdido ao longo do tempo, após o grande
desenvolvimento da indústria farmacêutica que, em contrapartida, demanda
pesquisas e tecnologia avançada, de alto custo, tornando seus produtos, os
quimioterápicos, muito caros, inacessíveis às populações de baixa renda, além de
proporcionarem a ocorrência de doenças causadas por efeitos colaterais e pelo seu
uso excessivo ou inadequado.
A recente expansão do mercado de plantas medicinais, baseado quase
que exclusivamente no extrativismo, vem contribuindo para a erosão das espécies.
10
O extrativismo de plantas medicinais tem sido feito ao longo do tempo, sem
nenhuma orientação a respeito do manejo e dos limites de coleta. Com o aumento
na demanda, têm crescido a pressão sobre determinadas espécies, o que pode
levar à eliminação de indivíduos e populações. Cascas, sementes e raízes têm sido
extraídas, muitas vezes sem a preocupação com a manutenção ou reposição dos
estoques naturais (MING et al., 1996).
Sendo a procura por plantas medicinal bastante crescente, existe a
necessidade de maiores estudos sobre processos de propagação que possam
garantir a produção de mudas em quantidade e qualidade necessárias para suprir
este mercado crescente.
A propagação vegetativa ou assexuada pode ser considerada como um
método economicamente viável de baixo custo na estratégia de preservação de
espécies nativas ameaçadas de extinção, além de contribuir para a formação de
bancos de germoplasma (PAIVA, 1993; SANTOS, 1994).
Segundo Rodrigues (1990), a falta de técnica na produção de mudas para
espécies nativas, e em alguns casos, a falta de viabilidade de sementes, indica a
propagação
assexuada
como
alternativa
à
multiplicação,
possibilitando
a
manutenção de boas características das plantas matrizes e a redução do período
juvenil, o que leva à antecipação do mecanismo reprodutivo.
Entre as técnicas de propagação vegetativa existente, a estaquia se
destaca por ser um método com resultados em um curto período de tempo e sendo
economicamente viável.
A propagação vegetativa pode ser considerada como a produção de
plantas utilizando caules, raízes, folhas ou outros órgãos no lugar das sementes.
Esse processo só é possível devido à capacidade que certos órgãos vegetais
possuem de se recomporem. Na verdade, algumas células desses órgãos contêm
informações genéticas necessárias para produzir uma planta toda quando em
condições
favoráveis,
propriedade
essa
chamada
de
totipotência
(ONO;
RODRIGUES, 1996).
Um dos fitorreguladores que apresentam maior efeito na formação de
raízes em estacas são as auxinas (FACHINELLO et al., 1995). As auxinas são os
reguladores vegetais com maior efetividade na promoção do enraizamento, cujo
principal efeito está ligado à sua ação sobre a iniciação dos primórdios radiciais
(HARTMANN et al., 1997).
11
Outro elemento promotor de crescimento é a cinetina, uma substância
sintética. Quando aplicada em certas células vegetais, como as de determinados
calos ou raízes crescidos em cultura de tecidos, essa substância atua junto com a
auxina e produz um enorme aumento das divisões celulares. A auxina sozinha
produz unicamente tumefação e aumento das células existentes, enquanto que a
cinetina estimula a divisão celular (mitose) e a formação de gema (LORENZI, 1998).
Maytenus ilicifolia Mart. ex Reiss., pertencente à família Celastraceae,
nativa do Brasil, secundária tardia, recebeu o nome popular de Espinheira - Santa
devido às folhas possuírem bordas com espinhos e propriedades medicinais
(MAGALHÃES, 2002). Um sub-arbusto ou árvore. Esta espécie tem sido
intensamente explorada nas populações naturais (CARLINE, 1988). A crescente
demanda tem levado a um extrativismo predatório, acarretando em erosão genética.
Casearia Sylvestris (Sw.), conhecida popularmente por guaçatonga,
pertencente à família Flacourtiaceae, pioneira é bastante promissora como opção
agrícola (MONTANARI JÚNIOR, 2002) e foi listada como uma das espécies
arbóreas prioritárias para conservação e manejo sustentável durante a primeira
Reunião Técnica sobre Recursos Genéticos de Plantas Medicinais e Aromáticas
(VIEIRA et al. 2002). Além do uso farmacológico, esta espécie apresenta grande
importância para o repovoamento de áreas degradas (LORENZI, 2002). Como a
maioria das espécies nativas com propriedades fitoterápicas, a C. sylvestris não é
cultivada, havendo extrativismo em áreas naturais o que promove a degradação da
flora nativa e distúrbios em ecossistemas (SILVA JUNIOR, 2005).
Trabalhos de domesticação de plantas medicinais são escassos ou
inexistentes para a maioria das espécies, sendo necessário o desenvolvimento de
estudos relacionados à adaptação destas plantas às condições de cultivo,
principalmente em virtude do aumento da demanda por parte das indústrias
farmacêutica e cosmética.
Partindo deste pressuposto, há a necessidade de
maiores informações a respeito do enraizamento de tais estacas para suprir a
demanda do crescente mercado de uso de plantas medicinais bem como, a
conservação de ambas as espécies.
12
2 OBJETIVOS
2.1 Objetivo geral
Avaliar a eficiência dos hormônios vegetais: ácido 3 indolilacético (AIA)
e cinetina (KIN), no enraizamento de estacas de Maytenus ilicifolia Mart. ex
Reiss. e Casearia sylvestris (Sw.).
2.2 Objetivos específicos
Testar diferentes concentrações de ácido indolilacético (AIA) e cinetina
(KIN) como indutores de formação de raízes em estacas lenhosas e herbáceas de
Maytenus ilicifolia Mart ex Reiss. e Casearia sylvestris (Sw.).
Analisar
as
respostas
das
espécies
em
relação
as
diferentes
concentrações de AIA e KIN aplicados.
Verificar a influência da consistência das estacas em relação aos
tratamentos de AIA e KIN.
3 REFERENCIAL TEÓRICO
3.1 Plantas medicinais
No Brasil, o conhecimento acerca das plantas medicinais, é uma rica
mistura de conhecimento dos indígenas, europeus e africanos, baseados em
espécies tropicais desde a colonização. A inter-relação do homem com as plantas e
13
seu ambiente é de grande utilidade para a medicina moderna ocidental (MING,
2003).
Atualmente, o comércio de fitoterápicos cresce 17% ao ano, enquanto
que o de medicamentos alopáticos cresce 10% (SIMÕES et al., 2003). O grande
desafio do último século tem sido desvendar e validar cientificamente espécies
vegetais com potencial terapêutico (BUTLER, 2005).
Segundo
Ferreira
(1998),
em
seu
projeto
de
pesquisa
sobre
“Medicamentos a partir de plantas medicinais do Brasil” ele enfatiza que dos
medicamentos vendidos em farmácias de todo o Brasil, 5,5% das vendas eram de
produtos contendo exclusivamente princípio ativo de origem vegetal.
A exploração de recursos genéticos de plantas medicinais no Brasil está
relacionada, em grande parte, à coleta extensiva e extrativa do material silvestre.
Apesar do volume considerável da exportação de várias espécies medicinais na
forma bruta ou de seus subprodutos, pouquíssimas espécies chegaram ao nível de
ser cultivadas, mesmo em pequena escala. O fato torna-se mais marcante quando
consideramos as espécies nativas, cujas pesquisas básicas ainda são incipientes
(VIEIRA, 1994).
As plantas medicinais representam a principal matéria médica utilizada
pelas chamadas medicinas tradicionais, ou não ocidentais, em suas práticas
terapêuticas, sendo a medicina popular a que utiliza o maior número de espécies
diferentes (HAMILTON, 2003).
As plantas são utilizadas por muitas pessoas na forma in natura como
produtos
saudáveis
ou
isentos
de
contra-indicação.
A
maioria
considera
medicamento natural com ausência de compostos secundários maléficos. O
conhecimento tradicional é rico em informações benéficas, porém, a falta de
conhecimento tóxico-farmacológico, deixa as pessoas mal informadas acerca das
plantas como as principais fornecedoras dos grandes venenos da história da
humanidade. As plantas não são uma fábrica de remédios prontos para doenças
específicas. Elas são um complexo de inúmeras substâncias, em sua maioria
desconhecidas, muitas delas capazes de exercer ação tóxica sobre o organismo
(RITTER et al., 2002).
Vários fatores poderão alterar a quantidade e a qualidade do princípio
ativo produzido pelas plantas como metabólitos secundários. Esses fatores podem
ser de ordem genética, técnica e fitotécnicas como: a forma de plantio, adubação,
14
tratos culturais e época de colheita. Os fatores fitossanitários e as condições
ambientais também influenciam na qualidade da planta como: o fotoperíodo,
umidade do solo, temperatura, disponibilidade de nutrientes, época do ano, vento e
poluição atmosférica, latitude e altitude também são fundamentais para um bom
cultivo com obtenção de substâncias químicas de qualidade (STEFANINI et al.,
2002; ROSAL, 2008).
O valor comercial de plantas medicinais e aromáticas tem refletido
qualitativa e quantitativamente na produção destas espécies. O uso de reguladores
de crescimento vegetal está sendo muito usado na agricultura devido a suas
influências positivas na quantidade e na qualidade de produção. Porém, pouco se
conhece sobre os efeitos destas substâncias em plantas medicinais e aromáticas.
Alguns estudos têm mostrado que essas substâncias têm sido muito utilizadas em
plantas medicinais visando à promoção do crescimento e ao aumento na
produtividade (SHUKLA FAROOQI, 1990).
Assim o estudo em relação à propagação vegetativa e a aplicabilidade de
hormônios vegetais de espécies utilizadas na medicina popular têm sido
intensificado nos últimos anos, devido ao crescente investimento em pesquisas para
a descoberta de novos fármacos e da utilização da fitoterapia como um meio
alternativo.
3.2 Propagação vegetativa
A propagação vegetativa é uma importante ferramenta no melhoramento
de espécies lenhosas e herbáceas e vem sendo amplamente utilizada, visando a
melhorar e manter variedades de importância econômica e medicinal (EHLERT et
al., 2004). É o método de propagação mais utilizado na produção comercial de
diversas culturas ornamentais e frutíferas tendo como vantagens a reprodução de
todas as características da planta matriz, uniformidade nas populações e produção
de produtos padronizados de alta qualidade (HARTMANN et al., 2002).
A propagação vegetativa pode ser também uma boa alternativa para a
reprodução de plantas que produzem poucas sementes e/ou para aquelas cujas
sementes germinam com dificuldade, assim como para a perpetuação de clones
15
onde as árvores são altamente heterozigotas e perdem suas características
desejáveis com a propagação sexuada (LORENZI, 1998).
As estacas podem ser produzidas a partir de porções vegetativas de
caules, rizomas, tubérculos e bulbos, folhas e raízes (BORTOLINI, 2006).
Na reprodução por estaquia há quatro fases que se pode distinguir,
iniciando-se com a produção de brotos, seguida da preparação da estaca e do meio
de crescimento, o enraizamento e por fim a aclimatação das mudas. As fases mais
importantes são o enraizamento e a produção de brotos, porque limitam a
possibilidade ou não e a quantidade de mudas a produzir (FLORIANO, 2004).
A estaca caulinar, objeto deste trabalho é constituída por segmento de
ramo com gemas apicais ou laterais, podendo ser lenhosa ou herbácea. As estacas
coletadas em um período de crescimento vegetativo intenso, primavera e verão,
apresentam-se com poucas reservas de carboidratos e com consistência mais
herbácea. Já estacas coletadas no inverno possuem um maior grau de lignificação e
teores mais elevados de carboidratos (FACHINELLO et al., 1994).
A propagação in vitro de plantas medicinais tem sido realizada por vários
autores (PINTO et al., 1994; LAMEIRA, 1994) e incrementada nos últimos anos
devido a vários fatores, entre eles, dificuldades na reprodução, baixa taxa de
germinação ou exploração irracional, resultando na quase extinção de algumas
espécies e modificação do meio ambiente, dificultando a coleta de plantas saudáveis
(KAJIKI, 1996).
3.3 Fatores que afetam o enraizamento na estaquia
A capacidade da estaca emitir raízes é uma função da interação de
fatores endógenos e das condições ambientais proporcionadas ao enraizamento.
Tem sido observado que a formação de raízes adventícias deve-se à interação de
fatores existentes nos tecidos e da translocação de substâncias sintetizadas nas
folhas e gemas em desenvolvimento. Entre estes fatores, os níveis de carboidratos,
água, fitohormônios e nutrientes minerais são de fundamental importância (WANG e
ANDERSEN, 1989; SMALLEY et al., 1991; MAYNARD e BASSUK, 1992; HENRY et
al., 1992; RIEGER, 1992; FACHINELLO et al.,1994).
16
Muitos fatores afetam o enraizamento de estacas. Práticas baseadas
nestes fatores têm sido desenvolvidas para promover o enraizamento em espécies
com dificuldade para o enraizamento. Estes fatores podem ser divididos em: a)
fatores químicos (endógeno ou exógeno) que promovam o enraizamento; b) fatores
da planta que afetam o enraizamento: a juvenilidade dos brotos, a posição do broto
do qual as estacas são retiradas, diâmetro das estacas, a presença de gemas e/ou
folhas, efeito do período de coleta das estacas, influência das espécies, efeito do
período de dormência e, influência do estado nutricional; c) efeitos ambientais no
enraizamento: controle da umidade; luminosidade; aquecimento do substrato;
fotoperíodo e; tratamento e/ou acondicionamento dos brotos e estacas antes da
estaquia; d) outros fatores que afetam a resposta ao enraizamento: composição
química e física do substrato, alguns estresses ambientais e efeito do ferimento
(COUVILLON, 1988).
Para cada espécie, diversos fatores estão relacionados com a época ideal
de colheita, tais como o grau de lenhificação, o estado fenológico, o balanço
hormonal e o estado nutricional das plantas-mãe (TOFANELLI et al., 2001).
Considerando que o objetivo principal na produção de plantas medicinais
é a obtenção de matéria-prima em quantidade e qualidade desejadas, deve-se tentar
diminuir a interferência dos fatores ambientais, técnicos e da variabilidade química
natural das espécies (MARTINS et al., 1995).
3.4 Hormônios vegetais e suas aplicabilidades:
As auxinas podem ser utilizadas combinadas ou isoladas, com a
concentração variando de acordo com a espécie e o genótipo (SOUZA & PEREIRA,
2007). Compõem o grupo de fitorreguladores de crescimento que apresenta o maior
efeito na formação de raízes em estacas, pois possuem ação na formação de raízes
adventícias, na ativação de células do câmbio e na promoção do crescimento das
plantas. Além de influenciarem a inibição das gemas laterais e a abscisão de folhas
e frutos (FACHINELLO et al., 1995).
17
A concentração adequada dos indutores de enraizamento, entretanto, é
um dos quesitos a ser considerado uma vez que pode fazer com que atuem de
forma sincrônica promovendo o enraizamento, ou inibindo-o (BATISH et al., 2008).
O controle do desenvolvimento de raízes adventícias é influenciado por
substâncias reguladoras de crescimento. As auxinas são os únicos reguladores de
crescimento que aumentam a formação de primórdios radiculares (TAIZ & ZEIGER,
1991). A ação positiva das auxinas sobre o enraizamento das estacas, deve estar
relacionada com a divisão das células que darão origem à raízes (HAISSIG, 1972).
Além disso, estes hormônios levam ainda à síntese de RNA, o qual intervém na
iniciação do primórdio radicular (HESS, 1969), favorecendo a atividade metabólica,
necessária para o desenvolvimento de novos tecidos de raiz, estimulando seu
crescimento (BREEN & MURAOKA, 1973).
Sampaio (1998) relata que o transporte das auxinas é polar. Uma toxidez
de auxina durante o enraizamento pode manifestar-se apenas na fase de
alongamento das raízes. Por esta razão é, às vezes, recomendada a utilização de
dois meios de cultura para a rizogênese. Primeiramente, as partes aéreas
permanecem em meio com auxina, favorecendo a indução e posteriormente
passaria para meio sem auxina, estimulando assim a rizogênese e o crescimento
das raízes (FETT NETO et al., 1992).
Já as citocininas, que foram descobertas em cultivos de callus, provocam
divisão celular nas plantas e são reguladoras do crescimento, atuando desta forma
na morfogênese. As citocininas estimulam o crescimento pela expansão mais do que
pelo alongamento. São sintetizados nos meristemas da raiz e em frutos em
desenvolvimento; ocorrem como parte do RNAt solúvel (GEORGE, 1996; SAMPAIO,
1998). O transporte de citocininas, sob a forma de ribosídeo da zeatina, ocorre
principalmente pelo xilema, mas se dá também pelo floema. O tipo e a concentração
de citocinina influenciam o processo de multiplicação in vitro, (JORDAN &
ITURRIAGA, 1980).
Para o sucesso do cultivo in vitro é necessária a utilização de reguladores
de crescimento em associação, como auxinas e citocininas, que, segundo
Murashige e Skoog (1962), influenciam na forma e textura do calo, extensão da
formação do órgão, tamanho e número de órgãos formados, vigor e longevidade das
culturas. Para Noggle e Fritz (1976), estas substâncias iniciam as reações químicas,
mudam a composição química dentro da planta e quando associadas aos fatores
18
ambientais como luz, temperatura etc, interagem com os processos bioquímicos
durante os processos de crescimento e diferenciação.
Meredith et al. (1970), em trabalho com ácido indolacético (AIA) e cinetina
(KIN), constataram que o AIA pode melhorar o enraizamento de estacas, por
estimulação, alongamento e desenvolvimento de raízes iniciais pré-formadas; e a
cinetina pode ter estimulado a formação de raízes.
Além de estimular a divisão celular, variações nas proporções de auxinas
e cinetina colocadas em uma cultura de tecidos podem influenciar fortemente o tipo
de diferenciação celular. Quando a proporção do AIA é superior à de cinetina, certas
regiões dos tecidos em cultura formam raízes. Proporções maiores de cinetina
resultam no desenvolvimento de caules. Estes fatos demonstram que a divisão e a
diferenciação celular exigem a ação conjunta e harmônica de dois reguladores
vegetais: auxinas e cinetina (NOGGLE & FRITZ 1976).
3.4.1 Ácido indolilacético (AIA)
Os estudos sobre metabolismo das auxinas têm-se concentrado quase
que exclusivamente no AIA, devido as provas convincentes de que é a principal
auxina nas plantas, sendo ativa em concentrações extremamente baixas. O AIA não
é apenas sintetizado pelas plantas, mas também inativado durante os processos de
crescimento e diferenciação. A oxidação do AIA pode ocorrer por um processo de
foto-oxidação e através de reações de oxidação catalisadas por enzimas (AIAoxidase). Além dos processos de oxidação, o AIA pode ligar-se a outras moléculas
na planta, produzindo conjugados que podem reter ou perder a atividade auxínica
(SALISBURY, 1991).
O AIA é sintetizado a partir do aminoácido triptofano em rotas trp
dependentes (rota do ácido indol-pirúvico, rota do indol-3-acetaldeído e rota da
triptamina). As plantas podem obter AIA também pela β oxidação do ácido 3indolbutírico (AIB) ou por hidrólise de conjugados nos quais o AIA pode
eventualmente
estar
ligado
como
aminoácidos,
açúcares
ou
peptídeos
(WOODWARD & BARTEL, 2005). De um modo geral, a auxina natural mais
abundante é o AIA, entretanto, dependendo da espécie, da idade da planta e das
19
condições sob as quais a planta se desenvolve, outras auxinas naturais podem ser
encontradas: ácido 4-cloroindol-3-acético (4-cloroAIA) e ácido fenilacético. Dentre as
auxinas sintéticas encontram-se o ácido naftalenoacético (ANA), ácido 2,4diclorofenoxiacético (2,4-D), ácido 2,4,5-triclorofenoxiacético (2,4,5-T), dentre outras
(KRIKORIAN, 1991).
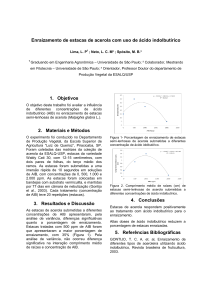
O ácido indolilacético (Figura 1), provoca a indução do sistema radical em
conjunto
com
carboidratos
e
compostos
nitrogenados.
Este
hormônio
é
principalmente sintetizado no meristema apical das folhas jovens, onde é
transportado em sentido polarizado, a partir do meristema apical até as
extremidades das raízes (TAIZ; ZEIGER, 2004).
Figura 1: Estrutura química do AIA (C10H9NO2).
Fonte: Taiz e Zeiger, 2004.
3.4.2 Cinetina (KIN)
Foi descoberta como um fator adicional à auxina, necessário em
quantidades muito pequenas para a divisão celular em certos tipos de tecidos
vegetais em cultura. A cinetina é um derivado da base purina, adenina, que é
importante em ácidos nucléicos. A cinetina é a citocinina sintética que é utilizada em
pesquisa vegetal. Não existe em plantas, mas um certo número de compostos
presentes nas plantas, conhecidos como citocininas, apresentam atividades
similares a cinetina (SAMPAIO, 1998).
20
Figura 2: Estrutura química do KIN
Fonte: Sampaio, 1998.
A formação de raízes adventícias é geralmente favorecida por uma
relação auxina/cinetina elevada, enquanto uma relação baixa favorece a formação
de gomos adventícios. Verifica-se, assim, que as estacas de espécies que
apresentam naturalmente um nível elevado de cinetinas têm maior dificuldade em
enraizar do que aquelas com níveis baixos de cinetinas. A influência das cinetinas
na iniciação da rizogênese pode depender da fase particular da iniciação e da
concentração hormonal. Alguns autores sugerem que os poucos casos de sucesso
com a aplicação exógena de cinetinas indicam que estas podem ter um papel
indireto que pode manifestar-se através dos efeitos no rejuvenescimento e na
acumulação de carboidratos na base das estacas (HARTMANN et al., 2002).
4 MATERIAIS E MÉTODOS
4.1 Descrição da área de estudo
21
A coleta do material vegetal foi realizada em um fragmento de floresta
Mata Atlântica (Figura 3) do
Horto Florestal da Universidade do Extremo Sul
Catarinense (UNESC), situada na região sul de Santa Catarina no município de
Criciúma, localizado nas coordenadas geográficas, latitude 28º 40’ 39”S e longitude
49º 22’ 11”W e a uma altitude média de 46 metros. Assim como em São Joaquim Serra Catarinense, em um fragmento de Mata Ombrófila Mista Montana (Figura 5),
localizada nas coordenadas geográficas, latitude 28º 15’ 31.5”S e longitude 49 53’
50.7”W, com altitudes média de 1.720 metros e Bom Jardim da Serra, em um
fragmento de mata ciliar floresta Ombrófila Mista Montana (Figura 6), localizada nas
coordenadas geográficas, latitude 28º 20’ 13”S, 49º 27’ 13”W, e altitude média de
1.245 metros. Compreendendo os meses de Maio, Julho e Outubro de 2010.
Figura 3: Vista satélite do Fragmento de floresta Mata Atlântica, Horto – UNESC.
Fonte: Google Earth, 2010.
22
Figura 4: Fragmento de Floresta Ombrófila Mista Montana, São Joaquim – Serra Catarinense,
2010.
Fonte: Do autor.
Figura 5: Fragmento de Mata Ciliar, Bom Jardim - Serra Catarinense, 2010.
Fonte: Do autor.
4.2 Maytenus ilicifolia Mart. ex Reiss.
23
Maytenus Ilicifolia Mart. ex Reiss. (Celastraceae), pertence ao gênero
Maytenus que conta com 200 espécies tropicais, sendo que a América do Sul dispõe
do maior número delas. Aproximadamente 76 espécies e 14 variedades ocorrem no
Brasil, sendo encontrada principalmente na região sul e nos países vizinhos,
Paraguai, Uruguai e Leste da Argentina, com ocorrência no interior de mata nativa e
mata ciliar, onde o solo é rico em matéria orgânica, em solo argiloso e bem drenado
(CORREA JUNIOR et al., 1994). Ocorre, principalmente, na Floresta Ombrófila
Mista, na Floresta Estacional Semidecidual, nas porções mais elevadas da Floresta
Ombrófila Densa, em sub-bosques em fase de sucessão secundária tardia ou
climácicas (KLEIN, 1968). A tabela a seguir descreve a classificação taxonômica da
espinheira-santa verdadeira segundo Engler e Cronquist:
TABELA 1 - CLASSIFICAÇÃO TAXONÔMICA SEGUNDO ENGLER E CRONQUIST
Táxon
Engler (Engler, 1964)
Divisão
Angiospermae
Magnoliophyta
Classe
Dicotyledoneae
Magnoliopsida
Subclasse
Archichlamydeae
Rosidae
Ordem
Celastrales
Celastrales
Família
Celastraceae
Celastraceae
Gênero
Maytenus
Maytenus
Espécie
Maytenus ilicifolia
Maytenus ilicifolia
Cronquist (Concrist, 1988)
A espinheira-santa apresenta diversos usos, são descritos ações
anticonceptiva, abortiva, anti-séptica, digestiva, antiespasmódica, tratamento de
feridas, de úlceras estomacais, diurética, laxativa, antiasmática, antitumoral,
problemas hepáticos e disfunção erétil porém a principal indicação das folhas de
espinheira-santa é o tratamento de gastrites, úlceras gástricas e duodenais
(HNATYSZYN et al., 2003; FERREIRA et al., 2004; REIS e SILVA, 2004).
De acordo com Carvalho-Okano (1992), M. ilicifolia é um subarbusto ou
árvore (Figura 6), ramificado desde a base, medindo até cerca de cinco metros de
24
altura. Apresenta ramos glabros; estípulas inconspícuas; folhas congestas,
coriáceas, glabras, com nervuras proeminentes na face abaxial, forma elíptica ou
estreitamente elíptica, base aguda a obtusa, ápice agudo a obtuso, mucronado ou
aristado. A margem das folhas pode se apresentar inteira ou com espinhos em
número de um a vários, distribuídos regular ou irregularmente no bordo, geralmente
concentrados na metade apical de um ou de ambos os semi-limbos. Apresenta
inflorescências em fascículos multifloros e frutos do tipo cápsula, bivalvar, orbicular.
As raízes são fortes e numerosas, avermelhadas externamente e amarelas no seu
interior (CARVALHO - OKANO, 1992; MARTINS, 2000).
Figura 6: Maytenus ilicifolia Mart. ex Reiss., aspecto geral da planta em São Joaquim – Serra
Catarinense, 2010.
Fonte: Do autor.
25
Figura 7: Detalhe do ramo, folhas e frutos do tipo cápsula.
Fonte: Reis e Silva, 2004.
As sementes de espinheira-santa são envoltas pelo arilo, excrescência
carnosa que se forma no funículo ou no hilo (VIDAL & VIDAL, 2000) que, para esta
espécie, é de coloração branca e fica exposta quando o fruto (cápsula) maduro abrese (CARVALHO-OKANO, 1992). Apesar de apresentarem boa germinação, as
plantas oriundas de sementes, apresentam a desvantagem da grande variabilidade
quanto à morfologia e ao teor de metabólitos, não havendo garantia quanto às
características do material propagado (PEREIRA, 1998).
Quanto à forma de dispersão natural das mesmas, M. ilicifolia apresenta
síndrome de dispersão zoocórica, especialmente realizada por aves (TABARELLI et
al.,1993).
O florescimento de M. ilicifolia começa no início da primavera e
permanece durante o verão (SCHEFFER, 2001). A frutificação ocorre nos meses de
novembro, dezembro e janeiro (CARVALHO-OKANO, 1992; SCHEFFER E
ARAUJO, 1998). Scheffer e Araujo (1998), observaram a ocorrência de frutos
somente em árvores que recebem insolação direta durante pelo menos parte do dia.
Segundo estes autores, mesmo recebendo luz direta, nem todos os indivíduos da
população frutificam no mesmo ano.
O estudo fitoquímico revelou presença de triterpenos e compostos
polifenólicos, como flavonóides e taninos nas folhas (YARIWAKE et al., 2005). Os
26
compostos fenólicos são encontrados principalmente como polifenóis: flavonóides e
taninos, sendo estes facilmente oxidáveis por enzimas, metais, luz e calor. Os
taninos possuem alto peso molecular e apresentam a propriedade de formar
complexos insolúveis em água, com alcalóides e com proteínas e, são responsáveis
pela adstringência em vegetais (SIMÕES et al., 2003).
MARTINS et al. (2003)
encontraram nas folhas de Maytenus ilicifolia, 8,72; 5,61 e 3,11 g% de polifenóis
totais, polifenóis não-tanantes e taninos totais.
Estima-se que cerca de 40% das plantas comercializadas como M.
ilicifolia sejam na verdade de outras espécies (SCHEFFER, CORRÊA JUNIOR e
GRAÇA, 2004), mas a maior demanda hoje é por folhas de Maytenus ilicifolia,
considerada a verdadeira (REIS e SILVA, 2004).
O comércio de espinheira-santa atualmente é de 160 ton/ano. No
Programa “Paraná 12 meses”, da EMATER-PR e UFPR foram recolhidas amostras
para avaliar a qualidade das plantas medicinais produzidas no estado. Os laudos
apontaram que 80% das amostras estavam abaixo do padrão exigido (REIS e
SILVA, 2004).
A propagação de espinheira-santa pode ocorrer por via sexuada, por
sementes, ou assexuadamente, por rebentos nascidos das raízes, alporquia e
mergulhia (SILVA JÚNIOR e OSAIDA, 2006), estacas caulinares herbáceas (SILVA,
1999) e também por meio da micropropagação (PEREIRA, 1998). A estaquia
normalmente é método eficiente para a propagação de espinheira-santa e obtenção
de material homogêneo, com caracteres genéticos desejáveis, produzidos a partir de
plantas matrizes selecionadas (SILVA, 1999).
4.3 Casearia sylvestris (Sw.)
Casearia sylvestris, é tradicionalmente considerado como pertencente à
família Flacourtiaceae (JOLY, 1998). No entanto, Flacourtiaceae foi recentemente
considerada um grupo não monofilético, e o gênero Casearia passou a ser
considerado como pertencente à família Salicaceae (SOUZA e LORENZI, 2005).
O gênero Casearia Jacq. é pan-tropical e conta com cerca de 180
espécies (CRONQUIST, 1981).
A América tropical e subtropical abriga a maior
27
diversidade do gênero, com cerca de 75 espécies (TORRES e YAMAMOTO, 1986).
No Brasil, o gênero Casearia é representado por aproximadamente 70 espécies
(MAISTRO et al., 2004). Casearia sylvestris Sw. é uma espécie de ampla
distribuição geográfica, ocorrendo do México até a Argentina em diversos ambientes
(SLEUMER, 1980, TORRES & RAMOS, 2007). Cresce preferencialmente na floresta
pluvial da encosta Atlântica, em áreas úmidas, aluvião, várzeas, encostas suaves e
até pedregosas. É seletiva higrófila, heliófita ou esciófita. São encontradas em todo o
território brasileiro, em praticamente todas as formações florestais (LORENZI, 1992).
Trata-se de uma espécie de alta densidade e ampla distribuição em
matas ciliares (DURIGAN et al., 2000) e em diversas outras formações C. sylvestris
apresenta grande variação com relação ao tamanho, forma e textura das folhas, à
pilosidade dos ramos mais jovens e da inflorescência, ao número de flores por
inflorescência e ao comprimento dos pedicelos (TORRES & YAMAMOTO, 1986).
28
Figura 8: Aspecto geral de Casearia sylvestris (Sw.)
Fonte : TORRES e RAMOS, 2007.
Casearia sylvestris (Sw.) apresenta inflorescência em fascículos, com
cerca de 20 a 40 flores hermafroditas por inflorescência (TORRES & YAMAMOTO,
1986), podendo chegar a 70 flores por inflorescência, (TORRES & RAMOS, 2007).
Do ponto de vista da interação planta-animal, as espécies são dispersas por aves
que consomem suas sementes (SANCHOTENE, 1989). Isto provavelmente, explica
sua grande dispersão pelo neotrópico. Suas flores apresentam néctar aromático o
que atrai uma grande quantidade de visitantes (insetos). Machado e Oliveira (2000),
relatam que os polinizadores mais importantes de Casearia grandiflora, uma espécie
próxima, são moscas da família Syrphidae, tendo sido observados também outros
insetos.
29
Figura 9: Ramo de C. sylvestris em florescimento.
Fonte: LORENZI, 1998.
A propagação é por sementes e estacas de ramos. A emergência das
sementes, cujo poder germinativo é baixo, ocorre entre 20 e 30 dias (LORENZI,
1992).
De acordo com Santos (2008), Casearia sylvestris, em particular, é
vastamente utilizada na medicina popular da América Latina. As indicações como
antiofídico, antitérmico, cicatrizante e no tratamento de úlceras e gastrites são as
mais citadas na literatura, sendo as folhas as partes mais utilizadas, seguidas por
cascas do caule e raízes. Muitas das aplicações populares desta espécie vêm sendo
corroboradas por estudos científicos (FERREIRA, 2006).
O grande interesse em C. sylvestris por parte da comunidade científica, e,
sobretudo, por parte da indústria farmacêutica, surgiu após a descoberta de seus
diterpenos clerodânicos, e de sua possível ação no tratamento do câncer (MORITA
et al., 1991).
A presença de óleos essenciais foi detectada na droga seca em um teor
de 0,12%, com uma grande porcentagem de compostos sesquiterpênicos
(POSSOLO
E
FERREIRA,
1949).
Silva
e
Bauer
(1970),
obtiveram
por
hidrodestilação das folhas frescas, o equivalente a 0,4% de óleo essencial, com
elevada porcentagem de terpenos (77,78%). Na desterpenação deste óleo
encontrouse o ácido hexanóico (0,58%), ácido este usado como matéria-prima na
preparação de ésteres que servem de adjuvantes em composições odoríferas.
30
Scavone (1970) detectou a presença de flavononas, saponinas e óleo essencial 2,1% em relação à matéria seca, líquido amarelado, com odor semelhante ao do
cedro e sabor não característico. Yamamoto (1995) e Luz et al. (1998), em
abordagem fitoquímica, evidenciaram a presença de taninos, flavonóides, saponinas
e traços de alcalóides.
4.4 Metodologia
4.4.1 Coleta das estacas
As estacas herbáceas de verão de Maytenus ilicifolia Mart. ex Reiss. do
primeiro experimento foram coletadas de matrizes presentes em São Joaquim, Serra
Catarinense. As estacas do segundo experimento, do tipo lenhosas e herbáceas de
inverno, coletadas em Bom Jardim. Já as estacas de Casearia sylvestris (Sw.), tanto
herbáceas quanto lenhosas foram coletadas de matrizes presentes no Horto
Florestal, UNESC. Ambas as espécies coletadas em épocas diferentes observando
o estado fisiológico em que a planta matriz encontrava-se, sendo realizado nos
meses de maio e julho de 2010.
Para confecção das estacas do primeiro experimento, foram selecionados
ramos herbáceos, coletados no mês de maio. Já as estacas coletadas para a
realização do segundo experimento, no mês de julho, foram de ramos lenhosos e
herbáceos de inverno. As espécies apresentavam-se em início de frutificação. Após
a coleta, as estacas foram deixadas em baldes previamente desinfetados com
hipoclorito 2%. Posteriormente ficaram de molho na água por aproximadamente 24
horas, para melhor hidratação das mesmas.
4.4.2 Preparação das estacas em laboratório
31
No dia seguinte da hidratação as estacas foram preparadas com
aproximadamente 8 cm de comprimento e 3-5 mm de diâmetro como (Figura 9),
fissuras foram feitas por estilete na base da estaca para aumentar a área do câmbio,
corte reto no ápice e em forma de bisel na base, com a finalidade de aumentar a
área de exposição do câmbio, sendo mantidas duas folhas superiores, com a
finalidade de aumentar a síntese de açúcares pela fotossíntese.
Figura 10: Estaca herbácea de Maytenus ilicifolia Mart. ex Reiss.
Fonte: Do autor.
Após serem desinfetadas em solução de hipoclorito de sódio 2% por 10
segundos, com imersão de cerca de 30% da base da estaca, as mesmas foram
banhadas com água destilada por cerca de 10 segundos para tirar o excesso de
hormônios.
32
Figura 11: Estacas emersas em água destilada.
Fonte: Do autor
As bases das estacas foram submetidas a diferentes tratamentos. Em um
primeiro grupo foram emersas somente no Ácido indolilacético (AIA) nas
concentrações de: 2.000 ppm, 1.000 ppm, 500 ppm, 250 ppm, 125 ppm.
Um
segundo grupo de estacas submetidas às mesmas concentrações de AIA mais 25
ppm de KIN.
O fitorregulador ácido indolilacético (AIA) C10H9NO2, foi diluído em
hidróxido de sódio (NaOH) 2 molar e completado com água destilada até 1L. A
partir disso diluído até as concentrações desejadas.
O tempo de imersão nas soluções com AIA e KIN foi de 15 segundos,
com 30% da base da estaca, posteriormente foram acondicionas em um béquer
contendo água destilada para retirada do excesso dos hormônios.
4.4.3 Acondicionamento das estacas do primeiro experimento - EPAGRI
Na Empresa de Pesquisa Agropecuária e Extensão Rural - EPAGRI de
Urussanga (Figura 12), as estacas de consistência lenhosa foram distribuídas em
tubetes de 10 centímetros de comprimento e 2 cm de diâmetro (Figura 13), contendo
50% de cinza e 50% de substrato comercial CAROLINA ® com a composição: turfa
de sthagno, vermiculita espandida, calcário dolomítico, gesso agrícola, fertilizante
NPK. Os substratos foram autoclavados e acondicionados na estufa de plástico UBV
associado a tela sombrite 50 %, com irrigação intermitente por nebulização.
33
Figura 12: Imagem satélite EPAGRI, Urussanga –SC.
Fonte: Google Earth, 2010.
Figura 13: Acondicionamento em tubetes de Maytenus ilicifolia Mart. ex Reiss..
Fonte: Do autor.
34
4.4.4 Acondicionamento das estacas do segundo experimento – Horto
Florestal – UNESC
Foram acondicionadas estacas com consistência herbácea de inverno
em canteiro contendo 50% de substrato comercial, associados a 50% de cinza de
casca de arroz, com sombrite 50%, em estufa tipo túnel com plástico UBV.
Com delineamento totalmente casualizado para as espécies de Maytenus
ilicifolia Mart ex Reiss. e Casearia sylvestris (Sw.), com 12 estacas para cada
tratamento, totalizando 144 estacas para cada espécie. Permanecendo em leito de
enraizamento de 90 dias para posterior análise.
A quantidade de estacas confeccionadas para os experimentos foi
delimitada pela quantidade de material vegetal disponível assim como pelo espaço
de armazenamento.
4.4.5 Variáveis analisadas
Para o processo de enraizamento as variáveis a seguir apresentam
influência direta:
•
Formação de raízes;
•
Número de brotações;
•
Formação de calos;
•
Estacas vivas
4.4.6 Análise estatística
O teste estatístico utilizado é o de Qui-Quadrado, com nível de
significância α= 5% com 1 grau de liberdade, para comparar as hipóteses levantadas
35
sobre diferenças entre os tratamentos de AIA e KIN em relação as variáveis
analisadas.
5 RESULTADOS E DISCUSSÂO
5.1 Estacas herbáceas de verão
Estacas herbáceas de verão da Casearia sylvestris (Sw.) foram
descartadas pelo fato da irrigação intermitente por nebulização presente na EPAGRI
não ser devidamente utilizada, ocasionando assim a morte de todo experimento.
Como o espaço é limitado, outros experimentos desenvolvidos no local
apresentaram contaminação por fungos justamente pela quantidade excessiva de
água, necessitando o desligamento da irrigação.
Pelo fato de Maytenus ilicifolia Mart. ex Reiss. apresentar uma
consistência mais coreácea e resistente, pode ter contribuído para a sobrevivência
das estacas, mesmo sofrendo estresse hídrico. Porém, devemos salientar que a não
hidratação contínua das estacas pode ter influenciado também no resultado das
estacas herbáceas de verão da espécie.
Para estacas de M. ilicifolia do primeiro experimento com AIA, observouse melhores resultados nas concentrações de 250 ppm, 500 ppm, 1000 ppm
(Tabela 2). Embora o resultado não seja estatisticamente significante observou-se a
formação de raízes, formação de calos e um maior número de estacas vivas
comparado com os demais tratamentos.
36
Tabela 2: Comparação estatística entre as variáveis usando Qui-Quadrado, do primeiro experimento
(AIA) com estacas herbáceas de inverno de Maytenus ilicifolia Mart. ex Reiss..
F.
Ñ. F.
Est.
Est.
Diluição AIA
Raízes
Raízes
Vivas
Mortas
F. Brot.
Brot.
2.000
2
10
1
11
6
6
0
12
1.000
2
10
2
10
9
3
0
12
500
2
10
1
11
7
5
0
12
250
3
9
4
8
7
5
0
12
125
0
12
1
11
5
7
0
12
0
1
11
0
12
5
7
0
12
q²
3.133
0.505
6.167
0.881
1.731
2.045
0
0
F. Calos Ñ. F. Calos
Ñ F.
N. F. = Não Formação/ F. = Formação.
Para as estacas do segundo experimento (KIN/AIA), os melhores
resultados ficaram entre 25 – 250 ppm, 25 - 500 ppm e 25 - 1000 ppm (Tabela 3).
Além de formação de raízes e maiores números de estacas vivas, a formação de
calos foi abundante, tendo diferença estatística significativa no tratamento 25 – 1000
ppm, onde a formação de calos ocorreu em todas as estacas do tratamento. Os
tratamentos 25 - 0 e 25 - 125 demonstraram ser menos eficientes quando
comparado com os demais tratamentos. Sendo que para 25 - 0, a variável não
formação de calos teve diferença significativa. Antunes et al. (2000), observaram
que em estacas de amora-preta obteve-se padrão similar entre enraizamento e
formação de calos, e mencionaram que estes dois processos se completam.
Tabela 3: Comparação estatística entre as variáveis usando Qui-Quadrado, do segundo experimento
(KIN/AIA) com estacas herbáceas de inverno de Maytenus ilicifolia Mart. ex Reiss..
Diluição
N. F.
Est
KIN/AIA
F.Raizes
Raíz
F. Calos N. F. Calos
25 - 2000
0
12
11
25 - 1000
2
10
25 - 500
3
25 - 250
Ñ. F.
Est Vivas
Mortas
F. Brot
Brot
1
12
0
0
12
12ª
0
12
0
0
12
9
10
2
11
1
0
12
4
8
6
6
12
0
0
12
25 - 125
1
11
2
10
7
5ª
0
12
25 - 0
0
12
1
11a
7
5ª
0
12
q²
6,333
1,022
13,714
19,200
2,702
14,995
0
0
Números seguidos da letra “a” na vertical não diferem entre si para alfa = 5% a 1 grau de liberdade.
37
Figura 14: Estacas de M. ilicifolia Mart ex Reiss. A) Estaca enraizada do primeiro experimento (AIA)
na concentração de 250 ppm. B) Estaca com formação de calos na concentração de 25 – 1000 ppm.
Botelho et al. (2005), indica que as estacas herbáceas retiradas durante o
verão apresentaram maiores concentrações de auxinas do que as semilenhosas e
lenhosas que foram colhidas no outono e inverno respectivamente, pelo fato das
plantas-mãe das primeiras se encontrarem em pleno crescimento.
5.1.1 Estacas herbáceas de inverno
Nas estacas herbáceas de inverno, assim como as herbáceas de verão
de Maytenus ilicifolia Mart ex Reiss., observou-se melhor resposta de enraizamento
nos tratamentos utilizados, comprovando a afirmação de Bastos et al. (2005), onde a
utilização de estacas herbáceas também permitiu a obtenção de melhores respostas
ao enraizamento do que com estacas lenhosas ou semilenhosas para o caquizeiro.
No
primeiro
experimento
(AIA)
(Tabela
4),
embora
não
tenha
demonstrado diferença significativa entre as variáveis, observou-se que o tratamento
com 250 µgg¯¹ teve melhor resposta quando comparado com os demais
tratamentos. Com maior número de formação de raízes, assim como maior número
de calos formados e um número considerável de estacas vivas.
38
A formação de calos sugere que se as estacas ficassem acondicionadas
por um tempo maior no substrato, há a possibilidade de surgirem novas raízes.
Hartmann (2002), afirma que o calo é uma massa irregular de células
parênquimatosas em diversos estados de lenhificação que geralmente se
desenvolvem na base das estacas quando colocadas em condições ambientais
favoráveis ao enraizamento, havendo relatos que as primeiras raízes aparecem com
frequência através do calo, conduzindo à suposição de que a formação do calo é
essencial para o enraizamento. Hill (1996), diz que a formação de calos seria uma
regeneração dos tecidos, resultante de um acúmulo de células na extremidade da
estaca cortada, precedendo ao enraizamento.
Tabela 4: Comparação estatística entre as variáveis usando Qui-Quadrado, do primeiro experimento
(AIA) de Maytenus ilicifolia Mart. ex Reiss..
Diluição
F.
Ñ. F.
Est.
Est.
AIA
Raízes
Raízes
Vivas
Mortas
F. Brot.
Brot.
2.000
2
10
1
11
6
6
0
12
1.000
2
10
2
10
9
3
0
12
500
2
10
1
11
7
5
0
12
250
3
9
4
8
7
5
0
12
125
0
12
1
11
5
7
0
12
0
1
11
0
12
5
7
0
12
q²
3.133
0.505
6.167
0.881
1.731
2.045
0
0
F. Calos Ñ. F. Calos
Ñ F.
Para M. ilicifolia Mart ex Reiss., os tratamentos de 25 - 125 e 25 - 250
(Tabela 5) obtiveram melhores resultados quando comparados com os demais
tratamentos. Apresentando maior número de formação de raízes, formação de calos
e estacas vivas. Para 25 - 125 houve ainda o surgimento de brotações. A aplicação
do AIA em associação ao KIN, mostrou-se bastante eficiente para estacas
herbáceas de inverno de M. ilicifolia. Para Hartmann (2002), a formação de raízes
adventícias é geralmente favorecida por uma relação auxina/cinetina.
39
Tabela 5: Comparação estatística entre as variáveis usando Qui-Quadrado, do segundo experimento
(KIN/AIA) de Maytenus ilicifolia Mart. ex Reiss..
Diluição
F.
Ñ. F.
Est.
Est.
KIN/AIA
raízes
Raízes
Vivas
Mortas
F. Brot.
Brot.
25 - 2.000
1
11
3
9
5
7
1
11
25 - 1.000
1
11
4
8
5
7
0
12
25 - 500
2
10
3
9
6
6
2
10
25 - 250
4
8
5
7
10
2
0
12
25 - 125
4
8
4
8
10
2
2
10
25 - 0
3
9
8
4
10
2
3
9
q²
2.900
0.763
3.389
2.033
3.420
6.051
5.417
0.677
F. Calos Ñ. F. Calos
Ñ F.
Figura 15: Estacas de M. ilicifolia Mart ex Reiss. A) Estaca morta do primeiro experimento (AIA) na
concentração de 2.000 µgg¯¹ . B) Estaca enraizada na concentração de 25 – 250 µgg¯¹.
Warmund et. al. (1986), afirmam que alguns inconvenientes estão
associados à propagação de estacas herbáceas, tais como: a necessidade de
manter elevada umidade durante o enraizamento, o que implica um custo adicional
com um sistema de nebulização na estrutura da estufa; o trabalho de propagação
40
ser mais intensivo; e a imposição de tratar rapidamente as estacas para prevenir
eventuais défices internos de humidade durante a propagação.
Para as estacas herbáceas de C. sylvestris (Sw.), não houve indício de
raízes no experimento contendo somente AIA (Tabela 6), porém houve diferença
estatística nos tratamentos 250, 125 ppm para a variável estaca viva.
Tabela 6: Comparação estatística entre as variáveis usando Qui-Quadrado, do primeiro experimento
(AIA) de Casearia sylvestris (Sw.).
Diluição
F.
Ñ. F.
Est.
Est.
AIA
Raízes
Raízes
Vivas
Mortas
F. Brot.
Brot.
2.000
0
12
0
12
1
11
0
12
1.000
0
12
0
12
1
11
0
12
500
0
12
1
11
2
10
0
12
250
0
12
0
12
8ª
4
0
12
125
0
12
2
10
8ª
4
0
12
0
0
12
0
12
0
12
0
12
q²
0
0
6.500
0.283
18.567
7.141
0
0
F. Calos Ñ. F. Calos
Ñ F.
Para o segundo experimento com KIN 25 ppm sem adição de AIA (Tabela
7), houve uma melhor resposta quando comparado com os demais tratamentos e
observando as variáveis estacas vivas e enraizadas. As dosagens de ácido
indolilacético 2000 ppm, 1000 ppm, 500 ppm, para as C. sylvestris podem ter agido
como agente herbicida, corroborando com a colocação de Fachinello (1995), onde
relata a ação herbicida quando usado AIA em altas concentrações, pois o teor
adequado de AIA para o estímulo do enraizamento depende da espécie e da
concentração de auxina existente no tecido.
41
Tabela 7: Comparação estatística entre as variáveis usando Qui-Quadrado, do segundo experimento
(KIN/AIA) de Casearia sylvestris (Sw.).
Diluição
F.
Ñ. F.
Est.
Est.
KIN/AIA
Raízes
Raízes
Vivas
Mortas
F. Brot.
Brot.
25 - 2.000
0
12
0
12
0
12
0
12
25 - 1.000
0
12
0
12
0
12
0
12
25 - 500
0
12
0
12
0
12
0
12
25 - 250
0
12
0
12
0
12
0
12
25 - 125
0
12
0
12
0
12
0
12
25 - 0
1
11
0
12
4
8
0
12
q²
4.833
0.068
0
0
19.333
1.137
0
0
F. Calos Ñ. F. Calos
Ñ F.
5.2 Estacas lenhosas
Nos tratamentos feitos com AIA para M. ilicifolia, destacaram-se as
concentrações de 250 ppm e 125 ppm (Tabela 8), onde houve diferença significativa
na variável estaca viva. As estacas tratadas nas concentrações de 2.000 ppm, 1.000
ppm e 500 ppm estão de acordo com a colocação de Fachinello (1995), onde o
mesmo relata a ação herbicida quando aplicado AIA em altas concentrações, pois o
teor adequado de AIA para o estímulo do enraizamento depende da espécie e da
concentração de auxina existente no tecido.
Outro fator que pode estar associado com o não aparecimento de raízes e
morte das estacas deve estar relacionado com o estudo de Sharma e Aier (1989) ao
trabalhar com estacas lenhosas observaram que à baixa capacidade de formar
raízes adventícias em estacas lenhosas, pode se dar pela presença do anel
esclerenquimatoso e ao aumento de substâncias inibidoras durante a dormência.
42
Tabela 8: Comparação estatística entre as variáveis usando Qui-Quadrado, do primeiro experimento
(AIA) de Maytenus ilicifolia Mart. ex Reiss..
Diluição
F.
Ñ. F.
AIA
Raízes
Raízes
F. Calos Ñ. F. Calos
Est.
Est.
Vivas
Mortas
a
Ñ F.
F. Brot.
Brot.
0
12
2.000
0
12
0
12
0
12
1.000
0
12
0
12
0
12
0
12
500
0
12
1
11
8
4
0
12
250
0
12
0
12
11
1
0
12
125
0
12
1
11
11
1
0
12
0
0
12
0
0a
7
5
0
12
q²
0
0
3.667
10.909
14.401
15.224
0
0
a
a
a
Para o segundo experimento contendo KIN/AIA (Tabela 9), observou-se a
morte de todas as estacas dos tratamentos, havendo diferença significativa para a
variável estaca morta na concentração de 25 - 500 ppm. Para a concentração de 25
- 250 ppm, embora não tenha ocorrido o enraizamento de nenhuma das estacas do
tratamento, assegurou a integridade de todas as estacas, mantendo-as vivas.
Hill (1996) relata que estacas lenhosas devem ser coletas no final do
outono até final de inverno, pois é onde a planta apresenta maior taxa de
regeneração, alto teor de lignificação e encontra-se em dormência, sendo condições
ideais para o enraizamento de estacas lenhosas.
Tabela 9: Comparação estatística entre as variáveis usando Qui-Quadrado, do segundo experimento
(KIN/AIA) de Maytenus ilicifolia Mart. ex Reiss.
Ñ. F.
Est.
Est.
F. Calos
Calos
Vivas
Mortas
F. Brot.
Brot.
12
0
12
1
11
0
12
0
12
0
12
4
8
0
12
25 - 500
0
12
0
12
0
12ª
0
12
25 - 250
0
12
0
12
11
1
0
12
25 - 125
0
12
0
12
9
3
0
12
25 - 0
0
12
0
12
5
7
0
12
q²
0
0
0
0
6.783
11.143
0
0
Diluição
F.
Ñ. F.
KIN/AIA
raízes
Raízes
25 - 2.000
0
25 - 1.000
Ñ F.
43
Para ambos os experimentos aplicados às estacas de C. sylvestris,
evidenciaram um completo insucesso no enraizamento das estacas. Jordanov (apud
STANKOVA; PANETSOS, 1997) sugere que o não enraizamento das estacas pode
estar relacionado ao curto período de leito de enraizamento, sugerindo um período
maior, pois alguns autores como destaca que as espécies apresentam períodos
variáveis para a indução de enraizamento, como é caso das estacas de Cupressus
sempervirens que foram acompanhadas por 220 dias. Ribas (1993), verificou que o
tempo necessário para se estabelecer o sistema radicial em estacas de Macadamia
integrifólia foi de 120 dias após a instalação do experimento.
No primeiro experimento (AIA) (Tabela 10), observou-se melhor resposta
na concentração de 125 ppm, houve formação de raiz, assim como manteve o maior
número de estacas vivas comparadas com as demais dosagens.
Tabela 10: Comparação estatística entre as variáveis usando Qui-Quadrado, do primeiro experimento
(AIA) de Casearia sylvestris (Sw.).
Diluição
F.
Ñ. F.
AIA
Raízes
Raízes
2.000
0
1.000
Ñ. F.
Est.
Est.
Ñ F.
F. Calos
Calos
Vivas
Mortas
F. Brot.
Brot.
12
0
12
2
10
0
12
0
12
0
12
3
9
0
12
500
0
12
0
12
3
9
0
12
250
0
12
0
12
7
5
0
12
125
1
11
2
10
10
2
0
12
0
0
12
0
12
0
12
0
12
q²
4.833
0.068
9.667
0.276
14.913
7.933
0
0
Para o tratamento feito com KIN/AIA (Tabela 11), a melhor resposta
ocorreu no tratamento contendo 25 ppm de KIN, com surgimento de raiz assim como
para a variável estaca viva foi estatisticamente significante quando comparado com
os demais tratamentos.
44
Tabela 11: Comparação estatística entre as variáveis usando Qui-Quadrado, do segundo experimento
(KIN/AIA) de Casearia sylvestris (Sw.).
Diluição
F.
Ñ. F.
KIN/AIA
Raízes
Raízes
25 - 2.000
0
12
Ñ. F.
Est.
Est.
F. Calos
Calos
Vivas
Mortas
0
12
0
12
a
a
Ñ F.
F. Brot.
Brot.
0
12
0
12
25 - 1.000
0
12
0
12
0
12
25 - 500
0
12
0
12
0
12
0
12
25 - 250
0
12
0
12
7
5
0
12
25 - 125
0
12
0
12
8
4
0
12
a
a
25 - 0
1
11
0
12
10
2
0
12
q²
4.833
0.068
0
0
21.953
11.677
0
0
Figura 16: Estacas de Casearia sylvestris (Sw.) enraizadas nas concentrações 125 (AIA) e 25 – 0
(KIN/AIA) respectivamente.
45
Estacas herbáceas em sua grande maioria independente da espécie
estudada tende a ter um número maior de estacas enraizadas quando comparadas
com estacas lenhosas, como observado por Browse (1979), a estaca caulinar
herbácea demanda maior controle ambiental no viveiro, entretanto possui maior
potencial de enraizamento. Porém no presente estudo, C. sylvestris não demonstrou
diferença significativa entre a consistência da estaca.
46
6 CONCLUSÃO
Para M. ilicifolia na consistência estacas herbáceas de inverno e verão o
melhor resultado ficou em torno de 25 – 250 ppm, em função do maior número de
estacas enraizadas e mantendo a integridade da grande maioria das estacas.
Para C. sylvestris a aplicação dos hormônios exógenos não tiveram
resultados significativos, acarretando a morte da grande maioria das estacas dos
tratamentos. Para o segundo experimento (KIN/AIA), o tratamento com 25 – 0 ppm
mesmo não sendo estatisticamente
significativa, representou melhor resposta
quando comparado com os demais tratamentos pois assegurou maior número de
estacas vivas assim como surgimento de raiz em uma das estacas.
Para as estacas lenhosas de M. ilicifolia a aplicação dos hormônios
exógenos também não demonstrou-se significativa para o tempo de 90 dias, porém
as concentrações de 125 ppm e 25 – 250 ppm, foram consideradas melhores
respostas pois mesmo não formando raízes, mantiveram um maior número de
estacas vivas assim como formação de calo em 125 ppm.
Para C. sylvestris, os tratamentos com 125 ppm e 25 - 0 ppm mesmo não
sendo estatisticamente significativo, obtiveram melhores resultados, aparecendo raiz
assim como maior número de estacas vivas.
Mesmo os resultados não sendo em sua grande maioria estatisticamente
significativos, as concentrações mais baixas de hormônios para ambas as espécies
demonstraram-se mais eficientes.
Em relação à consistência das estacas, constatou-se que estacas
herbáceas de M. ilicifolia obtiveram melhores resultados quando comparadas com
lenhosas, porém para C. sylvestris, a consistência das estacas não mostrou-se
significativa para este trabalho. A associação AIA/KIN foi positiva, levando ao
entendimento que KIN age como potencializador na presença de AIA.
Com o baixo índice de estacas enraizadas de Maytenus ilicifolia Mart ex
Reiss. e Casearia sylvestris (Sw.) sugere-se então, um novo estudo mantendo
estacas herbáceas e lenhosas nas mesmas concentrações, porém em leito maior de
enraizamento no substrato e um controle mais rígido das variáveis ambientais,
principalmente relacionados à irrigação.
47
REFERÊNCIAS
ANTUNES, L.E.C. Amora -preta: nova opção de cultivo no Brasil. Ciência Rural.
v.32. n.1. Santa Maria. p.151-158. Jan./fev. 2002.
BASTOS, D. C.; PIO, R.; SCARPARE FILHO, J. A.; LIBARDI, M. N.; ALMEIDA, L. F.
P. de; ENTELMANN, F. A. Enraizamento de estacas lenhosas e herbáceas de
cultivares de caquizeiro com diferentes concentrações de ácido indolbutírico.
Revista Brasileira de Fruticultura, Jaboticabal, v. 27, n. 1, p. 182-184,mês 2005.
BATISH, D. R. et al. Caffeic acid affects early growth, and morphogenetic response
of hypocotyl cuttings of mung bea (Phaseolus aureus) Plant Physiology, Ed. 165 p.
297- 305, (2008).
BORTOLINI, M. F. Uso de ácido indol butírico na estaquia de Tibouchina
sellowiana (Cham.) Cogn. 2006. 86 f. Dissertação. (Mestrado em Ciências
Agrárias). Área de Concentração em Produção Vegatal – Universidade Federal do
Paraná, Curitiba.
BOTELHO, R.V., Maia, A.J., Pires, E.J.P., Terra, M.M., Schuck, E. (2005). Efeitos de
reguladores vegetais na propagação vegetativa do porta-enxerto de videira "43-43"
(Vitis vinifera x Vitis rotundifolia). Revista Brasileira de Fruticultura, 27: 6-8.
BREEN, P. J. & MURAOKA, T. Effect of indolebutyric acid on distribution of (14)
Cphotosinthate in softwood cutting of Marianna 2624 Plum. Journal of American
Society for Horticultural Science, Alexandria, 98: 436-9, 1973.
BROWSE, P. M. A propagação das plantas. Lisboa: Europa-América, 1979. 229 p.
BUTLER, M.S. Natural products to drugs: natural product derived compounds
in clinical trials. Nat. Prod. Rep, n. 22, p. 162-95, 2005.
CARVALHO-OKANO, R. M. Estudos taxonômicos do gênero Maytenus Mol
emend. Mol. (CELASTRACEAE) do Brasil extra-amazônico. Campinas:
UNICAMP, 1992. 253p. Tese (Doutorado).
CARLINI, E.A. (coord.). Estudo da ação antiúlcera gástrica de plantas
brasileiras: Maytenus ilicifolia (espinheira-santa) e outras. Brasília : CEME/AFIP,
1988. 87p.
CORRÊA-JUNIOR C, Ming LC, Scheffer MC 1994. Cultivo de plantas medicinais,
condimentares e aromáticas. 2. ed. Jaboticabal: FUNEP.
COUVILLON, G.A. Rooting responses to different treatments. Acta horticulturae,
n.227, p.187-196, 1988.
CRONQUIST A. An integrated system of classification of flowering plants. New
York: Columbia University Press, 1981. 1261p.
48
CRONQUIST, A. The evolution and classification of flowering plants. 2.ed. New
York: New York Botanical Garden, 1988.
DURIGAN, G.; RODRIGUES, R.R.; SCHIAVINI, I. A heterogeneidade ambiental
definindo a metodologia de amostragem da floresta ciliar. In: RODRIGUES,
R.R.;LEITÃO FILHO, H.F.(eds.). Matas ciliares: conservação e recuperação.
EDUSP, São Paulo. 2000. p. 159-167.
EHLERT, P.A.D. et al. Propagação vegetativa da alfavacacravo utilizando diferentes
tipos de estacas e substratos. Horticultura Brasileira, Brasília, v.22, n.1, p.10-13,
2004.
ENGLER, H.G.A. Syllabus der Pflanzenfamilien. Berlin: Gebrüder Borntraeger,
1964. 666p.
FACHINELLO, J. C.; HOFFMAN, A; NACHITIGAL, J. C.; KERSTEN, E.; FORTES, G.
R. de L. Propagação de plantas frutíferas de clima temperado. 2 ed. Pelotas:
UFPEL, 1995. 178 p.
FACHINELLO, J.C., HOFFMAN, A., NACHTIGAL, J.C., et al. Propagação de
plantas frutíferas de clima temperado. Pelotas: UFPEL, 1994. 179 p.
FERREIRA, P.M.P. Determinação do potencial antitumoral de diterpenos
isolados das folhas de Casearia sylvestris Swarts. Fortaleza, 2006. 116 f.
Dissertação (Mestrado em Farmacologia) – Departamento de Fisiologia e
Farmacologia, Universidade Federal do Ceará, 2006.
FERREIRA, P.M. et al. A lyophilized aqueous extract of Maytenus ilicifolia
leaves inhibitis histamine-mediated acid secretion in isolated frog gastric
mucosa. Planta, v.219, p.319-324, 2004.
FERREIRA, S.H., et al., Medicamentos a partir de plantas medicinais no Brasil.
Ferreira, S.H., et al., Medicamentos a partir de plantas medicinais no Brasil.
Academia Brasileira de Ciências, São Paulo, 1998.133p.
FETT NETO,A.G.; TEIXEIRA, S.L.; SILVA, E.A.M.; SANTNNA, R. Biochemical and
morphological changes during in vitro rhizogenesis in cuttings of Sequoia
sempervirens (D. Don) Endl. Journal of Plant Physiology, Stuttgart, v. 140, p. 720728, 1992.
FLORIANO, E. P. Produção de mudas clonais por via assexuada. Série
Cadernos Didáticos. Santa Rosa: nº. 3, 1º ed., 2004. 37 p.
FRANCO, L. L. As sensacionais 50 plantas medicinais campeãs de poder
curativo. 5. ed. Curitiba: Editora Lobo Franco Ltda, 2001. 238 p
GEORGE, E. F. Plant propagation by tissue culture: the technology. 2. ed.
Edington: Exegetics, 1996. 1574 p.
49
HAISSIG, B.E. Meristematic activity during adventitious root primordium development
influences of endogenous auxin and applied gibberellic acid. Plant Physiology,
ethesda, 49: 886-92, 1972.
HARTMANN, H. T. et al. Plant Propagation: principles and practices. 6.ed. New
Jersey: Prentice-Hall, 1997. 770 p.
HARTMANN, H.T., Kester, D.E., Davis Jr, F.T., Geneve. R.L. Plant Propagation:
principles and practices. 7th ed. Prentice-Hall, New Jersey, USA, 2002, 880 p.
HAMILTON, A. Medicinal plants and conservation: issues and approaches.
International Plants Conservation Unit, WWF-UK, 2003.
HENRY, P.A., BLAZICH, F.A., HINESLEY, L.E. Influence of stock fertility on
adventicious rooting of stem cuttings. Journal of American Society Horticulture
Science, Alexandria, v. 117, p. 568-570, 1992.
HESS, C.E. Internal and external factors regulating root initiation; root growth.
2 ed. London, Buttersworth, 1969. p. 42-53.
HILL, L. Segredos da propagação de plantas: cultive suas próprias flores,
legumes, frutas, sementes, arbustos, árvores e plantas de interior. São Paulo: Nobel,
1996. 245 p.
HNATYSZYN, O. et al. Argentinian plant extracts with relaxant effect on the smooth
muscle of the corpus cavernosum of Guinea pig. Phytomedicine, v.10, p.669-674,
2003.
JOLY, A.B. Botânica – Introdução à taxonomia vegetal. 12ª ed. São Paulo,
Companhia Editora Nacional, 1998. 777p
JORDAN, M.; ITURRIAGA, L. Formación de raices en entrenudos de higuera (Ficus
carica L. cv. Adriatic) cultivados in vitro. Ciencia Investiones Agricolas, Buenos
Aires, v. 7, n. 2, p. 149-151, 1980.
KAJIKI, F.O. Estudo das condições para indução de calos e produção de
compostos secundários em Pfaffia glomerata (Spreng) Pedersen. Piracicaba:
ESALQ, 1996. 96 p. (Dissertação mestrado).
KRIKORIAN, A.D Propagación clonal in vitro. In: ROCA, W.M. E MROGINSKI. L.A.,
(Ed.). Cultivo de tejidos en al agricultura: fundamentos y aplicaciones. Cali,
CIATp.
95-25, (1991).
LAMEIRA, O.A.; COSTA, M.P.; PINTO, J.E.B.P. The efficiency of shoot and
plantlet formation of Cephaelis ipecacuanha after three subcultures in vitro.
Ciência Rural, Santa Maria, v. 24, n. 3, p. 523-526, 1994.
50
LORENZI, H..Árvores brasileiras: manual de identificação e cultivo de plantas do
Brasil. 4.ed. Nova Odessa: Plantarum, 2002. 368p.
LORENZI, H. Árvores brasileiras: manual de identificação e cultivo de plantas
arbóreas nativas do Brasil. Nova Odessa, SP: Plantarum, 1998. v. 2
LORENZI, H. Árvores brasileiras. Nova Odessa: Plantarum, 1992. p. 115.
LUZ S.F.B.; SATO M.E.O.; DUARTE M.R.; SANTOS C.A.M. Parâmetros para
controle de qualidade de folhas de C. sylvestris SW. –guaçatunga, 1998. Rev Bras
Farmacogn 7: 1-11.
MACHADO, A.O.; OLIVEIRA, P.E.A.M., Biologia floral e reprodutiva de Casearia
grandiflora Camb. (Flacourtiaceae). Revista Brasileira de Botânica v.23, p.283290, 2000.
MAGALHÃES, P. M. Agrotecnologia para o cultivo da Espinheira Santa.
Campinas, SP: UNICAMP, 2002.
MAISTRO, E.L.; CARVALHO, J.C.; MANTOVANI; M. S. Evaluation of the
genotoxic potential of the Casearia sylvestris extract on HTC and V79 cells by
the comet assay. Toxicology In Vitro, v.18, p. 337-342, 2004.
MARTINS, E. R. Plantas medicinais. Viçosa: UFV, 2000. 220 p.
MARTINS E.R.; CASTRO D.M.; CASTELLANI D.C.; DIAS J.E. 2003. Plantas
Medicinais. 5. ed. Ver. Viçosa: UFV, p. 15-19.
MARTINS, E. R.; CASTRO, D. M.; CASTELLANI, D. C.; DIAS, J. E. 1994. Plantas
medicinais. UFV-Viçosa. 220p.
MAYNARD, B.K., BASSUK, N.L. Stock plant etiolation, shading, and banding effects
on cutting propagation of Carpinus betulus. Journal of American Society
Horticulture Science, Alexandria, v. 117, n. 5, p. 740-744, 1992.
MEREDITH, W.C; JOINER, J.N.; BIGGS, R.H. Influences of indole-3-acetic acid and
kinetin on rooting and Índole metabolism of Feijoa sellowiana. Journal of the
American Society Horticultural Science, Alexandria, v.95, n.1, p.49-52, 1970.
MING, L. C.; SILVA, S. M. P.; SILVA, M. A. S.; HIDALGO, A. F.; MARCHESE, J. A.;
REIS, M.S. dos. Manejo sustentado de plantas medicinais em ecossistemas
tropicais. In: DI STASI, L. C. (organizador) 1996. Plantas Medicinais: arte e ciência.
Um guia de estudo multidisciplinar. 1a. ed. São Paulo, Ed. Unesp. p. 199-215.
MING, L.C.; SILVA, S.M.P.; SILVA, M.A.S.; HIDALGO, A.F.; MARCHESE, J.A.;
CHAVES, F.C.M. Manejo e cultivo de plantas medicinais: algumas reflexões sobre
perspectivas e necessidades no Brasil In: Diversos olhares em etnobiologia,
etnoecologia e plantas medicinais. Cuiabá: Unicen, 2003. p.149-156.
51
MONTANARI JÚNIOR, I. Aspectos da produção comercial de plantas medicinais
nativas. Centro Pluridisciplinar de Pesquisas Químicas Biológicas e Agrícolas UNICAMP. 2002.
MORITA, H.; NAKAYAMA, M.; KOJIMA, H.; TAKEYA, K.; ITOKAWA, H.;
SCHENKEL, E.P.; MOTIDOME, M. Structures and cytotoxic activity
relationship of casearins, new clerodane diterpenes from Casearia sylvestris
Sw. Chem. Pharm. Bull. v.39, p. 693-97. 1991.
MURASHIGE, T. 81 SKOOG, F. A Revísed Medium for Rapid Growth and Bio
Assays With Tobacco Tissues Cultures. Physiol.
Plant.,15:47;3-97,1962.
NICOLOSO, F. T.; LAZZARI, M.; FORTUNATO, R. P. Propagação vegetativa de
Platanus acerifolia Ait.: (I) Efeito de tipos fisiológicos das estacas e épocas de coleta
no enraizamento de estacas. Ciência Rural, Santa Maria, v.29, n.3, p 479-485,
1999.
NOGGLE, G. R. 81 FRITZ, G. J. Introductory Plant Physiology. New Jersey,
Prentice-Hall, 1976, 688 p.
ONO, E.; RODRIGUES, J. D. Aspectos da fisiologia do enraizamento de estacas
caulinares. Jaboticabal: FUNEP, 1996. 83 p.
PAIVA, H. N.; GOMES, J. M. Propagação vegetativa de espécies florestais.
Viçosa: UFV, 1993. 40 p.
PANIZZA, S. Plantas que curam: cheiro de mato. IBRASA, Instituição Brasileira de
Difusão Cultural LTDA, São Paulo, SP, 25ª ed, 280p, 1997.
PEREIRA, A.M.S. Micropropagação de Maytenus aquifolium Mart. e Maytenus
ilicifolia Mart. (espinheira-santa). In: MING., L.C. (Coord.); SCHEFFER, M.C.;
CORRÊA JÚNIOR, C.; BARROS, I.B.I.; MATTOS, J.K.A. Plantas medicinais,
aromáticas e condimentares: avanços na pesquisa agronômica. v. 2. Botucatu:
UNESP, 1998. p. 19-32.
PINTO, J.E.B.P.; ARELLO, E.F.; PINTO, C.A.B.P.; BARBOSA, M.H.P. Uso de
diferentes explantes e concentrações de benzilaminopurina na multiplicação in
vitro de brotos de Kielmeyera coriacea. Pesquisa Agropecuária Brasileira,
Brasília, v. 29, n. 6, p. 867-873, 1994.
POSSOLO, H; Ferreira C. Saponinas e outros compostos interessantes na família
das Flacourtiaceae. An Fac Farm Odont Univ São Paulo, 1949. 7: 377-385.
REIS, M.S.; SILVA, S.R. Conservação e uso sustentável de plantas medicinais e
aromáticas: Maytenus spp., espinheira-santa. Brasília: Ibama, 2004.
RIBAS, K.C. Efeito de auxinas, ácido bórico e sua interações no enraizamento
de estacas de Macadamia integrifolia Maiden & Betche. Botucatu, 1993.
Dissertação de Mestrado - UNESP, 128p.
52
RIEGER, M. Growth, gas exchange, water uptake, and drought response of
seedling- and cutting- propagate peach and citrus rootstock. Journal of American
Society Horticulture Science, Alexandria, v. 117, n. 5, p. 834-840, 1992.
RITTER, M.R.; SOBIERAJSKI, G.R.; SCHENKEL, E.P.; MENTZ, L.A. Plantas
usadas como medicinais no município de Ipê, RS, Brasil, Revista Brasileira de
Farmacognosia, (online), v.12, n.2, p. 51-62, 2002.
RODRIGUES, V. A. Propagação vegetativa de aroeira Schinus terebinthifolius
Raddi, canela sassafrás Ocotea pretiosa Benth e Hook e Cedrela fissilis
Vellozo através de estacas radiciais e caulinares. Curitiba, 1990, 90 f.
ROSAL, L.F. Produção de biomassa, óleo essencial e características
fisiológicas e anatômicas foliares de Plectranthus neochilus schlechter em
função da adubação orgânica, malhas coloridas e idade das plantas. 2008.
123p. Tese (Doutorado em Fitotecnia) Departamento de Ciências Agrárias,
Universidade Federal de Lavras, Lavras.
SALISBURY, F. B.; ROOS, C.W. Plant Physiology. Wadswoth, California: 1991.
cap.17, p.357-378.
SAMPAIO, E. Fisiologia Vegetal: Teoria e Experimentos. Ponta Grossa, PR: UEPG,
1998, 190 p. ALTMAN, A. The role of auxin in root initiation in cutting. Proceedings
of international Plant Propagation Society, London, 22: 280, 1972.
SANCHOTENE, M. C. C. Frutíferas nativas úteis à fauna na arborização urbana.
2 ed. Sagra, Porto Alegre, 1989. 306p.
SANTOS, A.G. Identificação dos princípios ativos antiulcerogênicos das folhas
de Casearia sylvestris: contribuição para o desenvolvimento de um fitoterápico.
Araraquara, 2008. 174 f. Tese (Doutorado em Química) – Instituto de Química,
Universidade Estadual Paulista.
SANTOS, M. C, dos. O uso da clonagem na silvicultura intensa. Revista
Silvicultura. São Paulo, v.15, p. 28-30,1994.
SCAVONE O.; GRECCHI R.; PANNIZA S.; SILVA RAP S. Guaçatonga (C. sylvestris
Swartz) Aspectos botânicos da planta, ensaios fitoquímicos e propriedades
cicatrizantes da folha. An Farm Quim São Paulo, 1970. 19: 73-81.
SCHEFFER, M.C. Sistema de cruzamento e variação genética entre populações
e progênies de espinheira-santa. 2001. 104f. Tese (Doutorado em Engenharia
Florestal) – Curso de Pós-graduação em Engenharia Florestal, Universidade Federal
do Paraná.
SCHEFFER, M.C.; CORRÊA JUNIOR, C.; GRAÇA, L.R. Aspectos da Cadeia
Produtiva da Espinheira-Santa. In: _____. Complexo Agroindustrial das plantas
medicinais, aromáticas, e condimentares no Estado do Paraná: diagnósticos e
53
perspectivas. Curitiba: Sociedade Paranaense de Plantas Medicinais: EMATER
Paraná: Embrapa Florestas, 2004. p. 253-271.
SHARMA, S.D.; AIER, N.B. Seasonal rooting behaviour of cuttings of plum cultivars
as influenced by IBA treatments. Scientia Horticulturae, Amsterdam, v. 40, n. 4, p.
297-303, Nov. 1989.
SHUKLA, A.; FAROOQI, A.H.A.E. Review: utilization of plant growth regulators in
aromatic plant production. Current Research Medicinal & Aromatic Plants,
Amsterdam, v.12, p.152-177, 1990
SILVA, C. de P. Efeitos do ANA, ácido bórico, paclobutrazol e da época de
coleta, no enraizamento de estacas caulinares de espinheira-santa (Maytenus
aquifolia Mart.). 1999. 99 f. Tese (Doutorado em Ciências Biológicas, Botânica) Universidade Estadual Paulista, Botucatu, 1999.
SILVA GAA, Bauer L 1970. Análise do óleo essencial de C. sylvestris Sw-I. Rev
Bras Farm 6: 328-331.
SILVA JÚNIOR, A.A.; OSAIDA, C.C. Espinheira-santa (Maytenus ilicifolia) – da flora
ao medicamento. Agropecuária Catarinense, v. 19, n. 3, p. 36-40, 2006.
SILVA JÚNIOR, M. C. DA. 100 árvores do Cerrado: guia de campo. Brasília, DF:
Rede de Sementes do Cerrado, 2005. p. 278.
SILVA, M. L. V.; ALVES, A. G. C.; ALMEIDA, A. V. 2004. A zooterapia do Recife
(Pernambuco) uma articulação entre as práticas e a história. Biotemas, 17 (1): 95116.
SIMÕES, C. M. O.;MENTZ, L. A.; SCHENKEL, E. P.; IRGANG, B. E.;
STEHMANN, J. R. Plantas da medicina popular do Rio Grande do Sul. 5.ed.
Porto Alegre: Ed. Universidade/UFRGS, 1998.
SIMÕES, C. M. O. et al. Plantas da Medicina Popular no Rio Grande do Sul.
Porto Alegre: Univ. Fed. Rio Grande do Sul, 2003. 173p.
SLEUMER, H.O. Flora Neotropica Monograph n.22 (FLACOURTIACEAE).
New York: The New York Botanical Garden, 1980. 499 p.
SMALLEY, T.J, DIR, M.A., ARMITAGE, A.M., et al. Photosynthesis and leaf water
carbohydrate and hormone status during rooting of stem cuttings of Acer rubrum.
Journal of American Society Horticulture Science, Alexandria, v. 116, p. 10521057, 1991.
SOUZA, A.V.; PEREIRA, A.M.S. Enraizamento de plantas cultivadas in vitro. Revista
Brasileira de Plantas Medicinais. Botucatu, v.9, n.4, p.103-117, (2007).
SOUZA, V.C.; LORENZI, H. Botânica sistemática – Guia ilustrado para
identificação das famílias de Angiospermas da flora brasileira, baseado em
APG II. Nova Odessa: Instituto Plantarum, 2005. 640p.
54
STANKOVA, T.; PANETSOS, K. Vegetative propagation of Cupressus sempervirens
L. of cretan origin by softwood stem cuttins. Silvae Genética, Frankfurt, v.46, p.2-3,
1997.
STEFANINI, M.B.; RODRIGUES, S.D.; MING, L.C. Ação de fitorreguladores no
crescimento da erva-cidreira-brasileira, Horticultura Brasileira, Brasília, v. 20, n. 1,
p. 18-23, março 2002.
TABARELLI, M.; VILLANI, J. P. & MANTOVANI, W. 1993. Aspectos da sucessão
secundárias em floresta atlântica no Parque Estadual da Serra do Mar, SP. Revista
do Instituto Florestal 5(1): 99-112.
TAIZ, L.; ZEIGER, E. Plant physiology, Redwood City California: 1991. cap.16,
p.398-424.
TOFANELLI, M.B.D., Chalfun, N.N.J., Hoffmann, A., Chalfun Jr, A. Capacidade de
enraizamento de estacas lenhosas e semilenhosas de cultivares de pessegueiro.
Ciência e Agrotecnologia, 2001, p. 847.
TORRES, R. B; YAMAMOTO, K. Taxonomia das espécies de Casearia Jacq.
(Flacourtiaceae) do estado de São Paulo. Revista Brasileira de Botânica, v.9,
p.239-258, 1986.
TORRES, R.B.; RAMOS, R. Flacourtiaceae. In: WANDERLEY MGL, SHEPHERD
GJ, MELHEM TSA & GIULIETTI AM (coord.). Flora Fanerogâmica do
Estado de São Paulo. São Paulo: Instituto de Botânica. 2007. p. 201-223.
VIDAL, W.N.; VIDAL, M.R.R. Botânica: organografia; quadros sinóticos
ilustrados de Fanerógamos. 4.ed. Viçosa : Universidade Federal de Viçosa, 2000.
124p.
VIEIRA, R.E., SILVA, S.R., ALVES, R.B.N., SILVA, D.B., WETZEL, M.M.V.S., DIAS,
T.A.B., UDRY, M.C. ; MARTINS, R.C. Estratégias para conservação e manejo de
recursos genéticos de plantas medicinais e aromáticas: resultados da primeira
reunião técnica. Brasília, DF: Embrapa Recursos Genéticos e Biotecnologia: Ibama:
CNPq, 2002. 184p.
VIEIRA, R.F. Coleta e conservação de recursos genéticos de plantas
medicinais. In: CONGRESSO BRASILEIRO DE MEDICINA E TERAPIAS
NATURAIS, 1., 1994, Brasília. Trabalhos...São Paulo: Instituto Médico Seraphis,
1994. p.44-49.
WANG, Q., ANDERSEN, A.S. Propagation of Hibiscus rosa-sinensis: relations
between stock plant cultivar, age, environment and growth regulator treatments. Acta
Horticulturae, Wageningen, v. 245, p. 289-309, 1989.
WARMUND, M.R., STARBUCK, C.J., LOCKSHIN, L. GROWTH. Cold hardiness and
carbohydrate content of Vidal blanc grapevines propagated by hardwood vs.
softwood cuttings. American Journal of Enology and Viticulture, 1986. p. 219.
55
WERLE, Almeci L. B; ZANETTI, Gilberto D.; CERON, Carla S.; MANFRON, Melânia
P. Avaliação da atividade de Casearia sylvestris Swartz (Flacourtiaceae) sobre
os níveis séricos de triglicerídeos em ratos. Rev. bras. farmacogn. vol.19 no.
2a João Pessoa. Apr/June 2009.
WOODWARD, A.W; BARTEL, B. Auxin: Regulation, Action and Interaction. Annals
of Botany. v. 95, p. 707-735, 2005.
YAMAMOTO, M. M. C. sylvestris Sw.: Aspectos botânicos, fitoquímicos e
farmacológicos. Curitiba, 1995. 54p. Monografia -Curso de Especialização em
Ciências Farmacêuticas -Produtos Naturais. Universidade Federal de Paraná.
YARIWAKE JH, LANÇAS FM, CAPPELARO EA, VASCONCELOS EC, TIBERTI LA,
PEREIRA AMS, Franca SC 2005. Variabilidade sazonal de constituintes químicos
(triterpenos, flavonóides e polifenóis) das folhas de Maytenus aquifolium Mart
(Celastraceae). Rev Bras Farmacogn 15: 162-168.
 E CINETINA (KIN) NO")