Bioquímica – Sistema Imune
Profa. Elizabeth Chaves
Parte 1 – Carlos Eduardo
Então, vamos falar um pouco sobre sistema imune, e falar sobre a composição
bioquímica de moléculas que atuam nesse sistema.
Por que o Sistema Imune existe? É um sistema que só é encontrado em vertebrados,
dado que os invertebrados têm mecanismos mais simples de defesa, por exemplo, enzimas
que quebram DNA estranho.
Essas doenças infecciosas que vamos estudar no Sistema Imune são responsáveis por
1/3 das mortes no mundo. Nós temos vários tipos de agentes infectantes, como bactérias,
vírus, parasitas, príons. Tudo isso é elemento estranho que, ao infectar e se instalar no
organismo hospedeiro, vai desenvolver uma resposta por parte desse hospedeiro, que nós
denominamos Resposta Imune.
A tuberculose, por exemplo, é uma coisa que todos vocês ouviram falar, e está
novamente em definição, porque a tuberculose era mais ou menos controlada com
medicamentos. Em países desenvolvidos, cujas condições sanitárias são melhores, isso estava
praticamente extinto. Só que essa tuberculose hoje em dia está adquirindo resistência às
medicações existentes, uma série de mutações estão surgindo, e ela está voltando a ser uma
das causas de mortes em todos os países.
A malária é outra doença, que embora restrita ao continente africano, está se
globalizando, porque há uma série de pessoas que visitam a África. O continente está abrindo
mais suas portas do comércio. O que é preciso para difundir a malária? Que ele encontre um
vetor que torne possível essa propagação. É preciso um mosquito que propague a malária.
AIDS é uma doença mundial.
Úlcera gástrica, antes todos pensavam ser um problema de estresse, hoje se sabe que
é uma inflamação produzida por uma bactéria, a H. pylori, que é capaz de iniciar um processo
inflamatório, e daí surgir a úlcera. Hoje se pede sempre uma endoscopia, exame para saber se
há ou não a bactéria.
O que leva a que a gente tenha esses casos de infecções aumentando cada vez mais
no mundo? O aumento da população leva a um aumento dessas doenças infecciosas. Mas,
principalmente as baixas condições sanitárias que nós temos, além de educação. É muito mais
fácil você encontrar populações pobres infectadas por diferentes agentes, poliinfectadas, de
que você encontrar nas populações mais desenvolvidas.
E sistema público de saúde é outro problema sério, porque muitas de nossas doenças
seriam muito menos nocivas para a população se houvesse um bom sistema de esgoto,
fossas, etc. Mas, não tem, por aí a fora todo mundo pisa na água e está se contaminando.
Quais são os mecanismos de resistência que a gente tem pra essas infecções por
esses agentes, e essa exposição constante que temos às doenças? Existem 2 tipos de
respostas encontradas: INATA e ADAPTATIVA.
Resposta INATA é imediata, não-específica, não depende de contato prévio com o
patógeno (agente infectante). Por exemplo, você sofre um corte, e o agente infectante penetra.
Imediatamente, há uma série de recursos que os organismos usam para tentar combater
aquela infecção logo que ela está tentando entrar no organismo.
Se isso falha, então a Resposta ADAPTATIVA continua o processo para combater o
material estranho. Nesse tipo de resposta, o Sistema Imune é estimulado por um agente, e
desenvolve-se uma prevenção futura. Por isso, sarampo, catapora... são ditas doenças “de
criança”, porque após um primeiro contato, você fica prevenido contra ela por uma série de
fatores que ao longo da aula nós vamos falar.
A reposta inata está presente nos mais diferentes tecidos e atua como primeira forma
de defesa. Já a resposta adaptativa é específica, estimulada pelo próprio antígeno, e constitui
uma linha de defesa que atua somente quando o organismo invasor escapa do Sistema Inato.
A imunidade inata responde à exposição ao agente infeccioso de imediato, e esse
agente pode ter ser adquirido através do contato da pele, pela ingestão ou inalação, ocorre nas
primeiras horas ou dias da infecção; enquanto a Resposta Adaptativa só se expressa após 7
dias, nós vamos ver isso. Além disso, a inata é não específica, não se voltando contra um
único agente infectante.
Aqui vocês têm um desenho. Vamos supor que você tem um tecido qualquer, em que
houve um dano e um rompimento. Então, através dele, algumas bactérias estão tentando
penetrar no tecido. Com essa penetração, algumas células do corpo se põem no Sistema inato,
e fazem a liberação de alguns agentes vasoativos, e também de fatores quimiotáticos. O que
esses agentes fazem? Os agentes quimiotáticos provocam a atração para aquele local, onde
ocorre a penetração, e vários outros tipos de células e fatores que vão combater essa infecção
localizada. E o vasoativo vai aumentar a permeabilidade dos vasos, pra haver um influxo pro
tecido, saindo dos capilares sangüíneos, não só de células que extravasam, mas também de
líquido, exsudato do próprio sangue. Esse material vai fagocitar, por exemplo, a bactéria. E, ao
mesmo tempo que eles fagocitam, eles provocam o surgimento naquela área de uma série de
fatores que vão aumentar ainda o poder fagocítico das células.
Queremos dizer que, durante a penetração de qualquer agente num ferimento
qualquer, esse agente deve ser de imediato um estimulador da presença de substâncias que
vão aumentar a permeabilidade vascular e vão atrair substâncias, porque o agente quimiotático
vai puxar uma série de proteínas que vem desse exsudato, proteína complemento, Anticorpos,
proteínas reativas. E elas vão, com esse efeito, causar a morte desse agente infectante.
Quando isso não é feito muito bem pelo Sistema Imune inato é que essa bactéria vai
alcançar a corrente circulatória, vai se distribuir, dar alguma infecção mais tardia, e se prolonga
por mais tempo. A duração vai depender muito da resposta que o sistema dê.
PERGUNTA INAUDÍVEL. RESPOSTA: Complemento é uma série de proteínas que se
transformam durante o processo, e geram algumas moléculas de natureza protéica ou
peptídica, que vão surgir como agentes quimiotáticos.
PERGUNTA INAUDÍVEL. RESPOSTA: A própria célula que está sofrendo o ataque
forma os fatores vasoativos e quimiotáticos, com isso permitem o extravasamento. Então o
material que vem do sangue vai ajudar mais ainda.
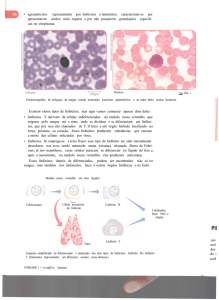
Essa fotografia eu achei muito interessante. Em verde são bactérias que são
marcadas; em roxo, macrófago tentando pegar essas bactérias; neutrófilos em azul que foram
atraídos por algumas substâncias, chamadas citocinas, que são proteínas liberadas pelo
macrófago à medida que ele fagocita as bactérias; e, em vermelho, uma hemácea. E, aqui uma
descrição daquilo, ou seja, a bactéria torna-se ligada à membrana por invaginação. Depois a
bactéria é ingerida pelas células, nesse caso, uma célula dendrítica. Então, ela entra, estimula
e é quebrada por enzimas lisossomais, e esse material é exposto para a superfície.
Aqui está sendo mostrado o material liberado. Na superfície dessas células, chamadas
Apresentadores de Antígenos, a gente vai ter receptores que vão mostrar esses fragmentos
que foram fagocitados e degradados. Então, com isso, você estimula todo o Sistema imune.
ACABOOOOOOOOOOOOOOOOU!!! Ninguém merece transcrever aula
Enzimabeth... Meus dedos tão doendo até umas horas!!! Mas, ta aí, galera, abraços...
de
Parte 2 – Carolina Reis
Bem, esse esqueminha aqui é somente para mostrar para vocês que eu lembro que eu
disse que vários tipos celulares, macrófagos, células dendríticas e outros vão ser atraídos para
o local. Se por acaso essas bactérias não forem controladas logo naquele local, elas
extravasam e num efeito mais tardio vocês terão a estimulação da resposta adaptativa. Dessa
resposta adaptativa vão entrar linfócitos B, e isso aqui representa receptores na superfície dos
linfócitos B, na realidade, são imunoglobulinas, principalmente o tipo de imunoglobulina M fica
mais presente aqui, e esses receptores são capazes, ali em cima você já vê através desses
pontinhos, como se fosse um Y, essas extremidades são capazes de interagir com o material
estranho, claro, dependendo da especificidade de cada um. Então, estimula, elas vão se
transformar em plasmócitos produtores de anticorpos etc. e dependendo também vocês têm a
estimulação dos linfócitos T. Então, quase sempre numa resposta imune vocês vão encontrar
uma resposta associada, tanto você tem a resposta imune humoral, que é a líquida, presente
nos líquidos, no sangue, na linfa e nos fluidos extra-celulares, como você tem a resposta
celular, que é aquela desenvolvida pelos linfócitos T e interagem célula-célula, mas quase
sempre a gente vai ver que sempre que a gente tem um o outro está paralelamente atuando
também. Bem, o que é um patógeno? São agentes causadores de doenças infecciosas. Como
é que eles fazem para penetrar na célula? Eles exploram atributos bioquímicos para penetrar.
Por exemplo, como é que a gente se banha em rios contaminados e a cercaria, que causa o
schistossoma, como entra? Ela entra porque ela é capaz de produzir uma enzima, uma
mucopolissacaridase, uma enzima chamada hialuronidase que rompe o ác. hialurônico que
está presente no tecido conjuntivo. Então, aquela malha que tem o ác. hialurônico prendendo
vários sulfatos de condroitina, como se fosse uma escova de cabelo. Então, ele rompe aquele
ác. hialurônico central e aí destrói e permite fazer como de fossem furos no tecido e aí
penetram até a corrente sangüínea. Têm outros que atuam por outros tipos de mecanismos
que interagem na superfície de determinados tipos celulares porque quimicamente se a gente
tem uma composição naquela célula que favorece ela se ligar, por que é que a gente é
infectado pelos mais diferentes vírus? Porque esses vírus têm mecanismos capazes de
interagir e eles são seletivos, nem toda célula é capaz de internalizar todo vírus, porque para
internalizar é preciso que tenha a determinada coisa química que ele possa realmente
reconhecer e aproveitar. Então, às vezes eles vivem numa perfeita simbiose com o meio que
eles possam se propagar. Então, se a gente pensar direitinho, o reino todo animal e vegetal, a
gente sempre tá destruindo alguma coisa para o outro viver. Então, eles exploram atributos
biológicos das células e o conhecimento do mecanismo de ação axila muito a combatê-los,
porque no momento que você sabe, por exemplo, o schistossoma sobrevive no sangue depois
que ele penetra porque na superfície dele ele tem receptores que interagem com a LDL. Por
que ele faz isso? Por duas razões: uma, porque quando ele se reveste todo com aquela
proteína do hospedeiro, pode ser albumina, uma série de proteínas nossas, ele ta camuflando
o reconhecimento pelo sistema imune. É difícil ser reconhecido porque o que ele externaliza
são proteínas nossas; segundo, eles pegam a LDL, entre outras proteínas, porque eles não
são capazes de fazer ác. graxos, que a membrana celular deles, como a qualquer membrana
celular tem fosfolipídios. Então, ele adquire os ác. graxos capturando isso das lipoproteínas do
próprio hospedeiro. Eles camuflam e ao mesmo tempo adquirem do hospedeiro e vivem muito
bem. E tem uma coisa interessante também, ele baixa o colesterol, o pessoal ta tentando
descobrir porque a schistossomose baixa o colesterol. Então, se você sabe esse mecanismo,
voltando à questão do mecanismo para combater, você pode desenhar e sintetizar drogas que
possam bloquear a interação, por exemplo, da lipoproteína com ela. No momento que você
bloqueia o sítio de interação, aquele bichinho vai morrer porque ele precisa dos ác. graxos do
hospedeiro para a sobrevida dele. Então, têm inúmeras coisas desse tipo, todas as drogas
desenhadas para qualquer tipo de doença são sempre baseadas em algum mecanismo
químico que é capaz de ser ativado ou bloqueado facilitando então a cura da doença.
Bem, essas superfícies epiteliais que a gente fala na imunidade inata
constituem a primeira barreira de defesa, a própria composição dela. Então, a gente tem como
exemplo a pele, o epitélio pulmonar e intestinal. Estas superfícies epiteliais internas, para
serem o primeiro elemento de defesa, contêm substâncias de natureza mucóide, um muco com
a composição que é uma garantia de que nem todo material que chegar no organismo da gente
vai penetrar. Porque o muco tem uma composição fixa que proteínas, principalmente mucinas
que dá aquela viscosidade, e vários tipos de glicoproteínas (é favorecida a síntese de
glicoproteínas pela vitamina A). A função desse muco é proteger. Essa substância de natureza
mucóide impede justamente esse acesso de microorganismo. Esses microorganismo interagem
com as superfície para penetrar de alguma maneira, ou pra furar e penetrar, ou pra interagir e
ser internalizada. Então, esse muco impede justamente essa adesão, na hora que impede a
adesão fica muito mais difícil penetrar. Então, a função é proteger a adesão dos patógenos e
também contem peptídeos antimicrobianos, denominados de defensinas, e essas defensinas
são realmente peptídeos de proteínas, mas de baixo peso molecular (12 a 50 aminoácidos) e
são positivamente carregados e esses peptídeos vão, de certa forma, estar presentes no muco
e vão ajudar a matar ou inibir o crescimento desses microorganismos. Apresentam domínios
hidrofóbicos que favorecem interações e constituem agentes terapêuticos úteis no combate a
infecções. Mas isso que eu estou me referindo é a defensina, que tem esse papel. Então o
muco ajuda a evitar essa adesão e por sua vez contem proteínas que favorecem o combate a
essas infecções.
Ainda no sistema inato, eu já falei antes, ele reconhece classes genéricas de moléculas
produzidas por vários agentes causadores de doenças. Esse sistema imune inato também usa
receptores chamdos Toll-like (TLRs), que são proteínas transmembrânicas que se encontram
na maioria das células, particulamente naquelas células com capacidade fagocítica, como o
macrófago. Então, esse tipo de receptor pode ser vantajoso ou não, vai depender. Se eles são
fabricados e apresentarem alta atividade, eles podem causar doenças crônicas. Se eles
atuarem demais, eles vão produzir tantas citocinas, estimular tanto o sistema imune, que
podem vir a produzir artrite e lúpus, exemplos de doenças auto-imunes. E seria bom definir
também. E quando eles estão pouco ativos, eles, ao contrário, em vez de estimular eles não
estão estimulando, e ai eles começam a deixar passar, digamos, muitas infecções se instalam
lá, porque não têm combate, então os receptores Toll-like são importantes também. E
estimulam a resposta inflamatória que, as vezes vocês vêem, aqui a gente dá um
ferimentozinho ai, as vezes, fica vermelho, incha aquele local um pouquinho, ai dependendo
da infecção pode dá febre, etc. Então o sistema imune estimula a resposta inflamatória que é
caracterizada por essas características e também a fagocitose. É interessante dizer que é
muito bom você está com febre porque quer dizer que o organismo está reagindo contra, que
está havendo uma reação.
Quais são essas moléculas que eu falei para vocês: reconhecem classes genéticas de
moléculas que são imunoestimulantes patógeno-associados. Que classe de moléculas são
essas? Como ele não é específico é preciso que tenha algumas características químicas que
ele possa reconhecer. Por exemplo, é muito comum os peptídeos que são formados, as
proteínas de uma maneira geral, nossas proteínas, nós iniciamos síntese com metionina. Então
vejam, quais essas moléculas que são imunoestimulantes, mas estão no agente infectante?
Lembrem que elas estimulam nosso sistema imune. Então uma delas é peptídeos contendo
formiometionina. O que é que esse peptídeo faz? Nossas proteínas quando são sintetizadas,
o primeiro aminoácido que é colocado nela, mesmo que depois ela tenha transformações e
modificações, mas durante a síntese protéica a nível dos ribossomos, primeiro aminoácido que
nós colocamos na proteína é a metionina, mas em bactérias não é a metionina é a
formiometionina, então vejam tem um padrão diferente do nosso, então qualquer proteína que
entre no organismo que seja formiometilada na extremidade amina, que é o início da proteína,
imediatamente estimula a ação do sistema imune inato, qualquer que seja ela, não interessa
de quem é, mas proteínas fabricadas por bactéria tem forniometionina. Qual é o outro padrão?
Parede celular de peptídeoglicano. Nós falamos isso semestre passado, quando a gente
mostrou a constituição de algumas bactérias, que tinha d-alanina, lembrem que os nossos
aminoácidos são L, mas na parede das bactérias tem uma constituição de peptídeos
associados com polissacarídeo, que nós chamamos de peptídeoglicanos. Esses peptídeos
contêm, entre outras coisas, d-alanina, que é um aminoácido diferente. Mas o sistema imune é
capaz de diferenciar o isômero D de um L? É. Ele tem uma especificidade tão fina que é capaz
de distinguir sim. Então bactérias que tenham essa característica, esse revestimento, são
identificadas. Flagelos de bactérias. O flagelo das bactérias, aquela tirinha que fica, aquele
flagelo tem uma composição protéica diferente da nossa. Então, como são proteínas
anormalmente presentes no nosso organismo elas são (moléculas imunoestimuladoras). Vocês
poderiam até me perguntar: e a gente num tem a flora intestinal? Dentro da flora intestinal as
bactérias têm funções. Agora, por exemplo, quando vocês tiverem uma cirurgia qualquer
abdominal, ou por algum rompimento qualquer desse intestino, uma pessoa com ascite, o que
é esse problema de ascite? É uma infecção ao nível do peritônio, porque as bactérias que são
contidas dentro da luz do intestino migraram para um tecido em que não é normal a presença
delas. Então, esse flagelo de bactérias, lipopolissacarídeos (LPS) das bactérias gramnegativas, ácidos tecóicos de bactérias gram-positivas, a parede celular de alguns
fungos que contêm uma série de polissacarídeos tipo zimosan, glucanos e quitina e
glicosilfosfatidilinositol em parasitas, é muito comum você vê proteínas que têm esse
resíduo, o glicosilfosfatidilinositol, que é um lipídio que fica na extremidade. No parasita, tem
uma proteína que está associada com esse resíduo, então esse resíduo é muito usado para
aderir essas moléculas, que eu falei que se mascara, às moléculas do hospedeiro. Então, ele
se liga através desses resíduos e se reveste com a molécula do hospedeiro. E também DNA
de bactérias, DNA estranho também estimula. Esses são apenas alguns exemplos para você
ver que quimicamente tem muita coisa, às vezes, em comum entre algumas bactérias, entre
alguns sistemas, entre alguns polissacarídeos, então não é especificamente aquele agente
infectante, mas é o grupo de agente que pode apresentar determinada característica diferentes
da nossa. E tem que ser diferente porque senão instalam-se doenças auto-imunes, que são
doenças quando o sistema imune nosso começa a não diferenciar o que é próprio do que não
é. Então, quando ele perde essa capacidade de diferenciar o que é nosso do que não é nosso
se instala a doença auto-imune.
Parte 3 – Carolina Asevêdo
Nós falamos nas estruturas químicas imunoestimulantes, e agora que receptores
são capazes de reconhecer esse material.Então o reconhecimento desses padrões de
substâncias que são imunoestimulantes vcs têm um principalmente um complemento.O que é
um complemento?Um complemento são proteínas solúveis que estão presentes em torno de
30 proteínas.São solúveis no sangue e na linfa também e que são capazes de atuar, mas são
lançadas no sangue mesmo,são formadas pelo e fígado.E esses receptores(?), que são
ligados às membranas das células hospedeiras que ajudam a iniciar a fagocitose e também
estimulam a expressão gênica das ditas citocinas,que são proteínas produzidas em resposta
ao sistema imune,quando ele é estimulado.Então existem vários tipos de proteínas que atuam
no sistema imune,que são produzidas por diversas células e que no geral a gente chama de
citocina.Então ele estimula a expressão gênica dessas citocinas que vão atuar na resposta
imune.Bem então eu vou mostrar quem é o complemento e depois quem é só o toll-like.
Vamos falar um pouco sobre o complemento.Esse complemento,cm eu disse,é
formado por várias proteínas existentes no sangue e que servem pra amplificar a resposta por
meio de uma cascata enzimática.O que isso quer dizer?Vcs jah viram a parte de sangue e
lembram da coagulação sanguínea.A gente tem, na coagulação sanguínea, vários fatores, e
um vai estimulando o seguinte, e assim por diante até a formação do coágulo.Então,aí é mais
ou menos isso, a gente começa com uma primeira proteína que estimula a segunda que
estimula a terceira e assim por diante, até nós chegarmos à destruição da bactéria,por
exemplo.Então, essa amplificação da resposta é feita por meio de uma cascata
enzimática,utilização de diferentes vias de ativação, a gente tem 3 vias, nós vamos
mostrar.Limitação do início da cascata a locais onde possa ser útil, vc não distribui para o
corpo todo, mas ele vai atuar quando se trata de resposta inata,principalmente, naquele
local.Ele vai junto quando existe um extravasamento que eu falei no início da aula.Aumenta a
permeabilidade vascular então extravasa material do sangue para o espaço do tecido onde
está havendo a inflamação.Então nesse caso limita àquela área ali.Regulação da resposta ao
invasor por proteínas solúveis e associadas à membrana celular.Então eu vou mostrar a vcs
que nessa cascata de reações que nós temos, algumas proteínas vão ficar aderidas à
superfície da molécula até pro fim, uma proteína que vcs já ouviram muito falar, que é a
perfurina, que é a última da casacata de reação.Ela fura a membrana da bactéria e
desequilibra o meio em que a bactéria está vivendo.Ela tanto extravasa material, quanto
internaliza líquido e outros materiais.E termina pro romper aquela estrutura e é aí que as
células fagocíticas atuam. Então o próprio complemento já tem substâncias que são
quimiotáticas e vão atrair essas células fagocíticas que vão destruir o material que está
infectado.Então isso aí é uma visão geral da função desempenhada pelo sistema
complemento.
Pergunta...Resposta:Quando a gente chama hospedeiro, é o organismo que está
sendo infectado.Então, quando eu falar várias células do hospedeiro,eu tou dizendo q as
células, em geral, são capazes de gerar em sua superfície uma família de proteínas que no
geral são chamadas de receptores toll-like.Então essa família de proteínas é que vai, no início,
dar início à ação do sistema imune.Ele realmente é muito bom, cm célula certo?Pq a célula em
si é que vai atuar, lá não, são proteínas que estão presente no líquido.Então eu vou explicar o
complemento e depois passo pra isso certo?
Bem, então o complemento, cm eu disse, a função do complemento, as vias de
ativação do complemento.Na verdade, o complemento tem 3 vias de atuação já bem distintas,
conhecidas e estudadas.Todas essas vias, a via clássica, a via da MBL e a via alternativa
todas elas convergem pra um ponto comum.O início delas é diferente, mas todas vão convergir
pra um ponto onde existe uma proteína chamada C3, essa proteína é o controle de toda
resposta imune. Os pacientes,por exemplo, que têm deficiência na produção de C3, que é uma
das proteínas do complemento,eles têm uma resposta imune baixíssima, eles podem morrer de
infecção a qualquer hora, por quê?Porque eles não respondem , principalmente, às
inflamações que estão em líquidos,que é a resposta humoral.De humos que é líquido.Então a
resposta humoral fica muito enfraquecida quando vc não tem a produção de C3, ou um C3
deficiente.Então, todas estão convergindo.Cm a acetil-coA, não é uma substância que
converge o processo degradativo de proteínas,carboidratos e lipídios pra pra daqui por diante
seguir?Então é isso, todas as 3 vias convergem pra C3,e daqui de C3 vc segue até haver a
morte da bactéria.Então veja, quem é a via clássica?A via clássica é uma via bem mais
demorada de que as outras duas, ela é bem mais demorada pq ela é mais elaborada de
início.Ela precisa, por exemplo, que haja a formação do complexo antígeno-anticorpo.Veja, pra
eu formar o complexo antígeno-anticorpo,quando é que eu vou formar ele?Eu vou formar o
complexo antígeno-anticorpo depois de estimular os linfócitos B.Então pra isso leva dias,por
quê?Os linfócitos B estão na circulação,acredita-se que, quando nós nascemos,nós temos uma
população de linfócitos.E vc não chega a diferenciar mt bem quando não há estimulação quem
é linfócito B ou T.Tamanho, etc, são mt semelhantes.Então o que é que acontece com essa
população?No momento em que,digamos, penetrou uma bactéria no meu sangue...Vamos
imaginar que nessa bactéria, vc tem várias substâncias químicas expostas aqui na superfície
diferentes.Vamos dizer que aqui vc tenha uma determinada seqüência de 5 aa, e aqui uma
outra seqüência diferente.Então claro, que quando vc estimula pela bactéria, o seu sistema
imune, um determinado linfócito vai conhecer essa estrutura.No momento que ele reconhece
essa estrutura aqui, que é diferente dessas outras,o segundo linfócito ta reconhecendo essa
daqui e o terceiro tá cá.Então vc está estimulando uma resposta de mais de um linfócito.Isso a
gente chama de uma resposta policlonal,porque o que é o clone?Vc pega uma célula, ela é
dividida forma 2, as filhas são exatamente iguais à mãe.As 2 filhas vão se tornar mães e vão
gerar 4 rebentinhos iguais a elas vc propaga células iguais a elas,isso é um clone.mas quando
vc ta propagando um linfócito que reconheceu essa seqüência aqui, um outro linfócito
propagou-se também aqui, então outro clone, e um outro linfócito propagou cá, então vc tem
uma resposta de vários clones, uma resposta policlonal.Enão toda resposta que nós
desenvolvemos contra qualquer agente infectante ela é policlonal.O que é mt bom, pq às vezes
um linfócito produz anticorpos com maior afinidade de que outros.A combinação anticorposantígeno é mt semelhante à enzima-substrato.Tem enzimas,cm a hexocinase,ela atua em
frutose, glicose, um bocado de coisas,mas é mais ativa com determinado
monossacarídeo.Então é a mesma coisa, quando vc estimula,alguns daqueles anticorpos a
serem formados podem ter mais ou menos afinedade pelo material.Então de qualquer forma a
resposta existe.Então vc dá uma penerada pra que a resposta seja efetiva.Então o que eu
queria dizer a vcs é o seguinte:pra vc formar o anticorpo pra ele atuar é preciso que primeiro,os
linfócitos que nunca foram estimulados fiquem na circulação procurando né, fiscalizando a
coisa.Reconheceu, foi lá ligou.Quando ele se liga,através de receptores da superfície,ele
estimula diferenciação celular, divisão e tal, e a formação dos plasmócitos que serão os
produtores
de
anticorpos.Então
desse
mecanismo
aqui,
de
identificação,reconhecimento,estimulação e produção de anticorpos, leva em torno, de um
primeiro contato, de 7 dias,a resposta.Então,por isso que vc não tem na via clássica, uma
resposta imediata, pq depende da formação antígeno-anticorpo.E somente 2 tipos de
imunoglobulinas estimulam, nós temos, na realidade, 5 classes principais. A imunoglobulina G,
a M, a A, a E e a T.A G é a que a gente tem em maior quantidade no organismo, então a G tem
subclasses.E a M, que é a primeira a ser produzida em qualquer infecção.Somente elas 2 têm
capacidade de estimular a via clássica.Mas claro que é pro elas mesmo,pq cm a IgM é a
primeira a ser produzida em uma infecção,só ela pode desenvolver,realmente, a resposta.E em
segundo lugar vem a IgG, que é a que existe em maior quantidade no organismo e que
substitui a IgM, pq a IgM é pentamérica e é uma proteína que requer mt dispêndio de energia
pra sintetizar ela.Já a via lectina MBL, ela forma aglomerados com 6 unidades globulares em
torno de uma proteína central.E o q é que essa reunião faz.Ele liga-se a manose e fucose
presente na superfície das bactérias.Então a via manose é mt interessante pra infecção por
bactérias.Ela não precisa da estimulação, ela se liga diretamente à bactéria, é mt mais rápida.E
a via alternativa idem.Ela não precisa de antígeno-anticorpo.Daí elas atuarem mt mais no
sistema inato do que no adaptativo.Então ela atua mt mais, a via alternativa, quando vc tem
polissacarídeos na parede das bactérias e leveduras tb, pq elas têm uma composição q atua
na via alternativa.
Pergunta...Resposta:Essa MBL, ela ta livre na circulação, então ela vai se ligar a
manose ou fucose que são tipos de monossacarídeos que vc vai encontrar nas paredes de
certas bactérias, elas vão interagir lá.Bem na hora que ele interage, é uma cascata de reação q
vai ocorrer.é cm se fosse uma marcação, o q vai se desenvolver daí por diante, eu vou mostrar,
q é uma cascata de reação.É uma marcação, cm a via alternativa tb, existem determinados
tipos de proteínas q vão se ligar à superfície de determinados tipos de bactérias e leveduras.Aí
isso feito, que é oq a gente chama de primeira etapa q seria o reconhecimento, aí viria uma
ativação e depois uma de morte mesmo da bactéria.
Pergunta...Resposta: Qdo a gente falou de proteína a gente falou sobre estrutura
quaternária, q é formada por mais de uma cadeia.Então é exatamente isso, é uma estrutura
espiralada e forma uma estrutura cm se fosse uma haste, e junto a ela tem repetições de uma
outra cadeia peptídica globular q fica ligada a essa primeira proteína.
Parte 4 – Daniela Pernambuco
O sistema complemento é composto por 30 proteínas distintas. Então, quando eu for
falar da via clássica, da via alternativa e da via MBL está dentro dessas 30 proteínas que eu
estou dizendo que existem dentro do sangue.
Pergunta: Ela depende da formação do anticorpo?
Professora: Ela depende sim, ela interage com ele. Eu vou mostrar como é.
Então, as vias vão mostrar como é que o complemento responde, ele pode ser através
da via clássica, da via alternativa e o da MBL. Agora eu vou mostrar a vocês onde é que se
junta isso. Vejam: aqui você tem a via clássica e a via alternativa, a MBL parece muito com
essas, mas eu vou mostrar só as duas já que foi esse esquema que eu achei.
A via clássica é formada por pelo menos 11 proteínas distintas. E o que essas
proteínas fazem? A proteína C1 se une ao complexo antígeno-anticorpo que está lá em cima
no slide (slide____). A imunoglobulina, aquela estrutura em forma de Y, está ligada ao antígeno
mas o pé desse Y está livre para atrair C1.
Quando vocês forem estudar imunoglobulinas vocês vão ver que funcionalmente a
cabeçinha do Y serve para identificação do agente infectante, do patógeno, e o pé dele serve
para estimular, no caso do IgG e IgM, o sistema complemento através da via alternativa.
Quando essa proteína vem e interage com isso aqui, ela vai ser estimulada. Essa proteína já
está no sangue.
Então C1 ela é formada por três cadeias polipeptídicas unidas entre si, é como se você
tivesse um buquê de flores, o talo e as flores em volta. Através dessa parte de cima é que ela
interage com o pé do Y, quando há essa interação ela estimula as três cadeias que formam
essa proteína única chamada C1 a se aderir mais fortemente entre si. Sendo essa três cadeias
chamadas de C1s, C1r e C1q. Uma vez que tudo isso está estimulado a C1s passa a constituir
uma enzima proteolítica ativa.
Então, repetindo, eu tenho o complexo antígeno e anticorpo, no pé do Y eu ligo C1 que
é formado de C1q, C1r e C1s e elas aderem fortemente a proteína pra ficar tudo juntinho lá. Só
que nessa adesão, estimulada também pela presença de cálcio que circula no sangue, o C1s
perde uma parte de sua estrutura que vai ser quebrada. Nós falamos sobre isso quando
estudamos proteínas onde eu disse que você pode ter uma proteína grande que para ser
ativada perde uma parte de sua estrutura, como acontece com os zimogênios. Nesse caso do
sistema imune, essas proteínas são produzidas no estado inativo exatamente para impedir que
elas fiquem atuando sem necessidade, então, no momento que é estimulado o sistema a C1s
perde uma parte da sua estrutura e passa a constituir uma enzima ativa.
Então, agora o conjunto tem uma enzima ativa, portanto C1 agora está ativado com
propriedades de enzimas proteolítica. C1 ativado vai ligar C4 e C2, não pensem que vai pela
ordem dos números pois esses números foram dados a medida que foram sendo descobertas
as proteínas. C2 e C4 catalisados pela C1s ativa vão ser estimulados e vão se dividir em duas
partes. Então, isso aqui é C4 e isso aqui é C2 e eu tiro um pedaçinho menor de C4 e C2 e
libero. Esse pedacinho menor que é liberado é chamado de fragmento A, formando assim C4A
e C2A. Esse pedacinho A tem propriedades quimiotáteis. Os fragmentos maiores que ficaram
da C4 e C2 são chamado de C4B e C2B e se ligam as bactérias e essa ligação do peptídeo a
membrana do invasor é um fenômeno chamado de opcinização (galera não sei escrever
isso). Eles vão recobrir a membrana dessa bactéria. Esse C4B e C2B juntos e aderidos a
bactéria vão constituir o agente ativador do C3 que é a proteína central pra qualquer dessas
vias. C4B e C2B podem ser chamados de C3 convertase já que vão quebrar C3 em C3B e C3A
que novamente o C3A é um agente quiomiotátil.
Pergunta: ???
Resposta: Porque eles têm propriedades químicas que os aderem a membrana. É
tudo químico.
Pergunta: ??
Resposta: Imagine assim, me parta ao meio, um dos meus lados será C2 e o outro
será C4. Meus braços serão uma das proteínas. Eu tiro a minha mão que será o fragmento
menor(A) e o resto do meu braço é o fragmento maior(B). Os dois fragmentos A que eu cortei
vão sair e atrair fagócitos para aquela região. Os outros pedaços maiores, C2B e C4B, que
ficaram estão agora juntos na superfície daquele negócio. Então, eles dois juntos constituem
agora uma nova enzima (C3 convertase) que vai ativar C3. Isso tudo é via clássica. Essa C3
convertase quebra C3 em C3A e C3B. Sendo o segmento A o que irá atrair os fagócitos, certo?
E o B fica novamente exposto na superfície.
Bem, esse C3B se ligando com C4B e C2B ele já vai ativar C5. Então, C5 já vai ser
quebrada também em C5A e C5B, lembrando que sempre o fragmento B é o que fica. Então,
C3B agora uniu com C5 inativa que será ativada após a atuação de C4B, C2B e C3B. Todos
esses juntos já estão disposto lá na superfície e vão atuar na conversão de C5 para C5A e
C5B. Daí por diante, embora já se conheçam as proteínas C6, C7, C8 e C9, não se sabe
exatamente como é que acontece. Admite-se que continue essa cascata proteolítica onde a
enzima anterior estimula a seguinte e assim por diante.
Então no final o que é que nós temos? O C9 que é a ultima dessas proteínas aqui vai
promover a quebra da membrana (ataque a membrana) propriamente dita. O C9 é uma
proteína relativamente pequena mas que tem propriedades hidrofóbicas então ele penetra na
membrana formando algumas estruturas que estruturarão um poro na membrana. Através
desse poro é que ocorre o extravasamento de material da bactéria pro meio. Com isso,
moléculas estranhas (material bacteriano) vão estimular o sistema imune e os fagócitos estarão
lá pra pegar tudo, os fragmentos celulares da bactéria e as substâncias internas dela que estão
sendo liberadas.
Então qual a diferença quando a gente tem a via alternativa ou MBL? Nós não temos
C1, C4 e C2. Aqui você tem um pouquinho de C3 que fique da via alternativa (eu acho que
depois ela corrige isso e diz que é da via clássica, foi até Bruno que perguntou... mas n tenho
certeza) já é suficiente pra que você reaja ele com um fator que é chamado D já que isso é
uma reação enzimática e poucas quantidades já são suficientes. Tudo isso é proteína. Então, o
C3B junto com esse outro fator protéico chamado B, no caso da via P, a via alternativa, ele se
une e forma C3B que já está lá. O B vai se dividir em dois fragmentos então B(bezão) e
b(bezinho) isso daqui substitui, no caso, essa da via clássica que é o C4B e C2B, então será
ela a C3 convertase da via alternativa. E daí por diante a coisa continua exatamente igual.
Porque isso é feito?! Porque nesse caso nós não estamos precisando de estimulação pelo
antígeno-anticorpo então, a presença de C3 estimulante existindo um elemento infectante
carboidrato onde ele se ligue, imediatamente ele vai se ligar a superfície e o resto corresponde.
Por isso que eu disse que a falha imunológica de um paciente que tenha deficiência de
produção, ou produza um C3 diferente, vai causar uma falha muito grande na resposta humoral
dele fica sendo feito tudo a nível de linfa, sangue e líquidos extracelulares. E a MBL em vez de
ter isso ela se liga diretamente com o agente infectante e daí ela já converte C3B em B e b e já
continua a via. Então, recordando tudinho pra vocês entenderem. O complemento é um
conjunto de proteínas que a gente tem no sangue, essas proteínas podem migrar para os
tecidos por extravasamento dos líquidos celulares no momento de algum ferimento ou entrada
de bactérias. Quando isso chega lá, o que é melhor? Bom, se for a resposta inata o ideal é que
a resposta seja dada ou pela via alternativa, reconhecendo a estrutura das bactérias ou pela
MBL que se liga à glicose e manose. Se essas duas vias falharem mas adiante a gente vai ter
essa daqui, mas só essa aqui só vai ocorrer quando tiver um estimulo para produção de
anticorpo e esse estimulo já é adaptativo.
Pergunta: Qual a função da via clássica?
Resposta: A via clássica é pra quando você tem um infecção no sangue.
Pergunta: Mas a função do complemento não é estimular a resposta?
Resposta: É.
Pergunta: Então, se ela estimulou... pra que a via clássica?
Resposta: Ah... aí é que está. É que o anticorpo é apenas marcador. É porque
normalmente a gente não sabe mas quando um anticorpo liga-se ao invasor é apenas para que
a resposta venha depois, ele não destrói material nenhum não. Por isso que eu digo que o
sistema imune é complexo porque ele é formado por muitas proteínas e muitas células. O
anticorpo é tipo um marcador e ele fazendo essa marcação ele estimula o processo todo.
Pergunta: ??
Resposta: O C3B normalmente você tem um pequena quantidade que está lá na
circulação, então, se você tiver um pouquinho você já vai formando mais C3B, é autocatalitica
a coisa.
Pergunta: ??
Resposta: Essas proteínas são produzidas pelo fígado, pelas células hepáticas. É por
isso que uma lesão do fígado é considerada grave pois muitas proteínas do sangue são
produzidas no fígado.
Pergunta: Então todas essas proteínas já estão preparadas para promover uma
resposta eficaz ou tem algum mecanismo que estimula? (acho que foi isso)
Resposta: Não, ela já deve está pronta para uma resposta eficaz, de imediato. Agora,
claro que você pode que com isso estimular um pouco mais a produção mas já deve existir
pois você não pode ficar esperando ela aparecer.
Pergunta: ??
Resposta: Não, eu disse que o anticorpo que a gente tava vendo aqui era marcador...
Quem destrói são os fagócitos, e principalmente no caso de bactérias, são essas proteínas
chamadas de complemento.
Pergunta: ??
Resposta: É a perfurina (acho que é a C9). Agora existe mais de um tipo de perfurina,
ela é que está furando e fazendo o furo. Então é isso que leva a morte da bactéria e não a
marcação do anticorpo. O anticorpo ele reconheceu um pontinho químico aqui diferente que eu
não tenho e que portanto é estranho a ele, e ele se liga.
Pergunta: ??
Resposta: Também, mas você não tem único tipo de proteína que é chamada de
perfurina.
Pergunta: A via clássica pode ser um resposta inata?
Resposta: Normalmente não, porque para ter uma resposta inata você teria que ter a
produção de anticorpo. É mais comum que você tenha somente a MBL e o outro na resposta
inata.
Pergunta: ??
Resposta: No caso da vacina, eu ou já chegar lá.
Pergunta: ??
Resposta: A C9 é a perfurina. Essas proteínas são um espécie de família que são
produzidas em algumas células. Então, no caso, essa perfurina particular aí é produzida no
fígado.
Pergunta: ??
Resposta: Normalmente, nós não temos manose e frutose (acho que é) expostas.
Porque lembra que eu falei pra vocês que as nossas proteínas tem acido cialico nas
extremidades exatamente para proteger. Então, na hora que você expõe resíduos de galactose
são normalmente... mas o que é que a gente tem mais?
Parte 5 – Edda Carvalho
(.....) não entendi
Vamos lá, e aqui tem (.....) manose, tem um proteoglic...pronto! receptores toll like, que
alguém tava aí curioso pra saber o que é. Olhe, vamos supor que isso aqui seja uma célula,
eles até (......) como se fosse uma célula dendrítica, né... então essa célula dendrítica é uma
célula que existe uma concentração muito baixa dela no sangue, mas ela tem uns tentáculos
soltos como se fosse um polvo, então essa célula dendrítica que ta sendo simplificada aqui
vocês tão vendo esses risquinhos aqui... na realidade esses receptores, essa família de
receptores que a gente chama de toll like, eles são encarregados de que? vejam, a bactéria,
(......) ali uma bactéria gram negativa que tem ali a liberação de LPS que é lipoproteína solúvel,
né... então essa LPS...desculpe, lipopolissacarídeo, então essa LPS é reconhecida, é como de
fosse... é uma interação química que a gente poderia comparar talvez com a interação que a
gente faz de antígeno/anticorpo, porque é químico/químico, quer dizer, ele simplesmente
reconhece. Quando ele se liga, imediatamente depois dessa ligação você estimula toda a
célula a produzir citocinas, então as citocinas são produzidas pela célula estimulada que
reconheceu o material antigênico e elas são liberadas. Então vários tipos de citocinas, aqui, por
exemplo, tem um grupo de citocinas que se chama interleucinas, acho que vocês já ouviram
falar, né... tem interleucina um, dois, três, quatro, cinco, seis... então aqui, interleucina seis, o
que é que ela faz? Promove a atividade de células B, ou seja, ela aumenta a atividade das
células B, consequentemente da produção de anticorpos. A interleucina doze promove a
atividade de células T, então aumenta a atividade... então ela faz aumentar o que eu falei lá no
início, ela faz aumentar a expressão gênica de determinadas proteínas que estarão envolvidas
na resposta imune. Então como a célula dendrítica é das primeiras a estar presente na
imunidade inata, ela faz isso muito bem... hoje em dia, inclusive, eu não sei se eu falei pra
vocês, houve uma época...ainda está se pesquisando isso... há uns dois anos atrás, foi
publicado um trabalho feito com um pessoal da França e lá do LIKA, e que eles usaram uma
vacina contra a AIDS com uma resposta muito boa, qual foi essa resposta? Teve uma paciente,
uma mulher que ela negativou completamente, hoje em dia ela não tem nem traços mais do
vírus, e os outros todos não chegaram a 100% porque também eu acho que foi uma dose ou
duas que deram, mas os outros tiveram uma resposta assim, negativaram em 80%, 85%, então
isso é uma das coisas que a gente quis muito estudar mas esses pacientes são um
pouquinho...se vocês chegarem lá espero que vocês não trabalhem com AIDS, e que já tenha
sido descoberta uma cura pra ela, mas eles são muito escorregadios, é difícil você contactar,
eles às vezes se acanham por conta do problema social, e tudo mais, então a gente não
conseguiu levar à frente... tá se voltando a trabalhar junto com o pessoal da biologia do Ageu
também... é esse processo... mas foi muito interessante, e o que é que eles faziam? Vejam, é
uma auto-vacina, se tirava do paciente o sangue dele, isolava as células dendríticas, que são
de primeira resposta, estimulava elas, porque elas já tinham sido pré-informadas do vírus,
então já tinha uma memória, então essas células são estimuladas, aumenta-se a população
delas e as reinjeta. Então como essas células dendríticas saem direto dos gânglios linfáticos
pra estimular toda a produção de anticorpos, então a resposta se torna assim...é um boom,
né...aumenta enormemente, então com isso a gente conseguiu praticamente... agora, sai caro
o tratamento? Sai, porque é individualizada a coisa, você tira da pessoa, prepara todo o
material e reinjeta, então não é um tratamento que você vai vender na farmácia, tem que usar
um laboratório, mas mesmo assim, se conseguir eu acho que é fantástico, né... o que a gente
conseguiu foram dezoito eu acho (......) e todos eles tiveram uma resposta fantástica com esse
tipo de célula... quem sabe se esses receptores toll like não são estimulados? eu não sei... Eu
contar a vocês 100% o mecanismo de estimulação eu não sei porque eu não trabalhei no meio
mas é muito interessante, então vejam...( alguém faz uma pergunta que não dá pra entender e
ela responde): talvez por isso mesmo, porque a população de células dendríticas nossas é
pequena em relação às outras células brancas do sangue, de uma maneira geral é
pequena...ela constitui mais ou menos 0,2% coisa desse tipo...(aluna interrompe novamente e
ela retoma): aí eles estimulam pra compensar... é... aumentar a quantidade. Bem, voltando
aqui, ( outra aluna faz pergunta e ela responde): é isso que tá se tentando, se estimulasse, mas
de qualquer maneira tem que se descobrir...porque o caminho é longo porque veja, a colega da
gente tá perguntando: não tem como estimular alguma coisa que estimule o aumento delas
sem que a gente faça essa auto-vacina? Pode ter mas pra a gente chegar lá, a gente tem que
descobrir, é ela que tá... é ela que é capaz... então, qual é o fator, o que eu é que ela tem que
pode fazer isso... como a gente pode aumentar... então são coisas que...(ela falou essas frases
soltas mesmo, aí ficou meio confuso) pesquisa é um negócio que você nunca pára porque
quando você descobre alguma coisa você vai em busca de outra... então não é fácil. Bem,
então voltando aqui, esse outro esquema mostra os mesmos receptores pra mostrar pra vocês
que a gente tem realmente uma família, TLR2, o outro que pode ser de um tipo, aliás quando
tem duas cores é bom lembrar pra vocês que pode ser o mesmo receptor toll ou em
combinação, quando aparece às vezes em dupla, então quando tem colorações diferentes é
que tem mais de um, então ali é o seis com dois, ali o quatro com quatro, aqui é o cinco
sozinho e assim por diante. Então o que é que já se estudou? Já se estudou que esses
receptores daqui eles são particulares...eu não vou ler um por um que não dá, mas por
exemplo, citar um deles, conectar ao RNA, esse daqui, o sete, ele conecta ao RNA viral de fita
simples como o do HIV, pode ser que precise estimular ele. Esse outro daqui, que adere ao
DNA viral de fita dupla como no caso do vírus do Nilo ocidental, esse outro liga-se ao RNA viral
de fita simples, então eles hoje em dia, já tá se estudando tanto essa família de receptores que
já chega a se identificar o papel funcional deles, então já tem muita coisa em imunologia
avançando, então esses receptores realmente têm sido muito estudados pelos imunologistas.
Bem, o que é que as células dendríticas fazem? Elas instruem o sistema imunológico a
reconhecer o que é próprio do que não é, então o papel funcional delas é importantíssimo, ela
é capaz de diferenciar as estruturas químicas nossas do que não é nosso. Ela inicia e controla
a resposta imunológica. São cruciais pra formação da memória imunológica... o que é memória
imunológica? É justamente quando eu...daqui a pouco a gente apresenta, mas aquilo que eu
disse há pouco pra vocês, que quando a gente estimula um linfócito, B por exemplo, ele se
divide, se divide, se divide, então a resposta amplia muito, então nessa divisão uma pequena
fração desses linfócitos estimulados permanece na circulação, elas não chegam a atingir o
estágio de plasmócito produtor, ela fica como linfócito, mas estimula, por exemplo, se você
pegar um linfócito for estimulado depois que crescer essa população, esse clone, você deixa
100 na circulação, então imaginem, é muito mais fácil que quando eu tiver uma reinfecção pelo
mesmo agente esses 100 reconheçam e combatam muito mais rapidamente do que quando a
gente tinha um só, que ao acaso, sete dias depois de circular ele achou... então é por isso que
quando a gente tem uma reinfecção, que eu tava citando no início da aula pelo sarampo, por
exemplo, a gente teve, constituiu a memória imunológica que pro resto da vida você vai
conservar, então aqueles linfócitos estimulados que já reconheceram, que tiveram o primeiro
contato com o antígeno e que vão ficar na circulação, esses linfócitos que constituem a
memória imunológica não vão dar trégua ao agente pela segunda vez e vão eliminar ele, então
não tem sete dias de espera, você com dois dias, três dias já tá desenvolvendo uma
quantidade de anticorpo enorme, que você não tem num primeiro contato, certo? Então é muito
importante a memória imunológica que a célula dendrítica favorece o aparecimento dele.
Constitui, como eu disse a vocês, bem pouquinho, é 0,2% dos glóbulos brancos, eles ingerem
patógenos e degradam, quando saem do tecido infectado amadurecem (......), nódulos
linfáticos e expõem antígenos na superfície celular ativando linfócitos T. Esses antígenos na
superfície celular é o que a gente chama de complexo principal de histocompatibilidade. (aluno
pergunta e ela responde): quando você tem a célula dendítica, vamos dizer que ela tá virgem,
não viu nada, não trabalhou ainda em nada, na hora em que ela... ela é uma célula fagocítica,
ela pegou o negócio, infectou, ela aí começa a degradar dentro dela esse material que ela
internalizou, como o macrófago faz, quebra... então esse material quebrado ela vai expor, por
isso que quando há pouco eu apresentei ali eu disse a vocês, tá mostrando esse material solto
mas ele não fica solto, ele chega na superfície dela, ele tem determinados...eu não quero fazer
a mesma coisa pra não parecer anticorpo... alguns receptores que vão externalizar esses
fragmentos quebrados no interior da célula, então quando a gente diz que ela amadurece é
porque no momento que o material estranho está sendo degradado pelas enzimas lisossomais
a célula tá estimulada a produzir os receptores que é o complexo principal de
histocompatibilidade que todo mundo tem em todas a células pra expor isso na superfície, aí
ela atrai o linfócito T porque a interação do linfócito T é célula/célula, entendeu? Por isso que
estimula, por isso que a gente diz que o amadurecimento dele é porque a estimulação interna
da presença do material antigênico fragmentado vai levar à formação dessas proteínas que
constituem receptores de superfície que vão externalizar e chamar atenção de que aquela
célula tem um material estranho e portanto o organismo tá em perigo. Bem, aqui é um resumo
daquilo que a gente já falou, vamos dizer, o corte, as bactérias, aqui não dá pra todo mundo
ver mas seria uma célula dendrítica, então ele tá aumentando ela trazendo pra cá, essa célula
dendrítica internalizou a bactéria, quebrou, externalizou fragmentos e vai para os nódulos
linfáticos onde estimula o amadurecimento dos linfócitos, certo? Então com essa estimulação
ele estimula não somente linfócitos B como também macrófagos, todo tipo de célula que atua
no sistema imune e aqui você tem exemplos, por exemplo o plasmócito já né... a célula B já
estimulada produzindo os plasmócitos e anticorpos que vão migrar pra aquele local, vai
estimular linfócitos T e assim por diante, certo? (aluna faz pergunta e ela responde): ela
estimula as próprias células imunes de uma maneira geral, elas quando estimuladas pela
presença do antígeno elas produzem uma série de citocinas que vão aumentar a resposta
imune, atraindo, ligando, fazendo uma série de papéis funcionais cada uma delas. Então nesse
caso, quando a célula dendrítica se sente estimulada e expõe esses fragmentos na superfície,
ela já está de certa forma permitindo que o linfócito T, qualquer linfócito T que tenha aqui e que
reconheça esse material antigênico se ligue a ele... no momento que ele liga é o mesmo tipo de
estimulação que tem pra célula B, só que o linfócito B ele reconhece nos líquidos ali o material
e o linfócito T tem outro tipo de receptor de superfície que reconhece também, então quando
ele se liga um ao outro esse linfócito T que reconheceu cria um clone dele, aí estimula.
Parte 6 – Edilane Guerra
Como havia dito, o sistema imune adaptativo é específico. Ele só existe quando é
estimulado pela presença do antígeno. O adaptativo tem duas respostas, uma resposta
humoral que é a resposta que ocorre dos fluídos de uma maneira geral, linfa, sangue e fluídos
extracelulares e nós temos a resposta que é dada pelos linfócitos B, na produção de anticorpos
no início. E tem um outro tipo de resposta que a gente chama de imune - celular, que é uma
resposta que é dada pelos linfócitos T, chamada de resposta celular, interação de linfócitos T
célula – à –célula.
A célula infectada interage com o linfócito T e no caso do linfócito B, não, é um
elemento estranho que se liga a ele, uma molécula ou uma macromolécula. Molécula pequena
dificilmente estimula o sistema imune, mas uma proteína, um DNA estranho, um carboidrato
estranho, polissacarídeos de uma maneira geral, todos eles estimulam. Moléculas muito
pequenininhas, se você injetar uma manose em um animal que não produza ou que não tenha,
passam despercebidas.
Então, quando se tem a presença de um antígeno, que é o organismo infectante, esse
antígeno vai ser reconhecido por alguma parte química. Essa parte química do antígeno que é
reconhecida e que se liga ao anti-corpo que está na superfície dos linfócitos, que a gente
chama receptores de linfócitos B, mas que na realidade esse pedacinho daqui (ela se refere a
perninha do Y da imunoglobulina) é hidrofóbico e se insere na membrana, mas isso aqui é o
anticorpo que depois será exportado, exatamente o mesmo.
Então, ele vai lá e reconhece; a pontinha química que ele reconhece e que ele se ligou
a gente chama de determinante antigênico. Tem uma seqüência de aminoácidos, de
carboidratos, é um trechinho químico daquele organismo que é estranho ao que nós temos,
então ele vai lá e se liga.
Quando ele se liga a esse determinante antigênico ou epitopo, é a mesma coisa, ele
estimula a... Nessa foto aqui ele está mostrando o linfócito B e o linfócito T. Eu tenho três tipos,
aqui de linfócitos B e T, será que todos são estimulados? Não! Por esse determinante
antigênico, só será estimulado somente o que reconhece ele, quimicamente isso aqui é
diferente, a gente vai ver jájá, é variável (se refere a parte superior e variável do Y) para se dá
uma variação grande de anticorpos. Então, o linfócito que reconheceu vai ser estimulado,
nesse caso ele está mostrando esse aqui mais triangular(receptor do Y), esse redondinho e
diferente desse que parece um V para mostrar que são diferentes receptores.
Então, quem reconheceu foi esse B2. Então, o B2 é estimulado a proliferar e produzir
plasmócitos para exportar os anticorpos, certas imunoglobulinas. Então, essas imunoglobulinas
são as mesmas daqui, só que quando ela é fabricada essa parte hidrofóbica da cauda daqui é
quem se liga para exportação, se não o plasmócito não faz. O plasmócito só faz a síntese da
cadeia polipeptídica daqui para aqui, então a parte que se liga a membrana não tem. Mas o
poder de reconhecimento que tem aqui é o mesmo, por isso que ele pode exportar e ele vai
direto atuar lá, deu para entender?
O linfócito T, ele não tem, veja que o desenho é diferente, ele não tem capinha, ele
está mostrando o tipo de receptores do linf. T diferentes, porque os receptores dos linfócitos T
jamais são exportados, a interação do linfócito T é diretamente com o material estranho.
Evidentemente ele não sai daqui. O que o linfócito T vai ter é a uma série de citocinas
que levam a apoptose das células e uma série de coisas, mas a interação continua sendo de
cel a cel. ( No desenho do slide não mostra a célula que o linf T está interagindo, mas ela disse
que estava errado, que na verdade era para ter.). Isso é a base do problema de rejeição,
porque a gente tem padrões de uma pessoa para outra. Então, o que causa rejeição é muito
isso. Então, aqui mais uma vez rapidamente, mostra que quando se forma um clone a partir da
célula eu formo cel produtoras de anticorpos (linfócito B) e formo células de memória. No caso
dos linfócitos T eu formo linfócitos ativos, produtores de citocinas e etc e as células de
memória.
Em ambos os casos, uma estimulação fará guardar uma memória imunológica, que
responderá mais facilmente em uma segunda infecção, certo? Esse é o mecanismo da primeira
infecção.
Aqui é as imunoglobulinas que chamamos de anticorpos que são proteínas formadas
por duas cadeias polipeptídicas, exatamente iguais, que são essa aí e 2 outras também iguais,
essas grandes chamamos de cadeias pesadas e essas são as cadeias leves e entre elas as
ligações são feitas por pontes de sulfetos. Então, essas pontes de sulfetos ligam essas
articulações das cadeias pesadas que chamamos de joelho o que permite um pouco de
mobilidade dessas estruturas para se ligar ao antígeno e todos os anticorpos tem pelo menos
dois sítos de interação, eu digo pelo menos porque há anticorpos que têm duplicações disso,
AGM, por exemplo, que veremos daqui a pouco.
Então, as mais simples são constituídas por essas quatro cadeias, duas pesadas e 2
leves, iguais entre si. Iguais entre si, se forem produzidas pelo mesmo linfócitos, se forem por
linfócitos diferentes elas diferem, mas a estrutura é sempre essa, 2 cadeias leves, 2 pesadas.
A estrutura da imunoglobulina G a gente pode visualizar agora melhor, na verdade isso
está sendo mostrado por esses bloquinhos para facilitar a visualização, mas as duas cadeias
pesadas e as duas leves a parte N, ou seja, o início da cadeia, que é a terminação N, está
voltado para cima, terminação amina, e a terminação carboxila para baixo.
Notem que somente as cadeias pesadas formam esse pezinho aqui, as cadeias leves
não formam. A gente também pode dividir isso em blocos porque? Porque a gente chama isso
aqui de domínios, domínio Ch1,Ch2,Ch3. (Domínio parte globular de uma cadeia
polipeptídica com funcionalidade própria). Então, nesse caso, essas cadeias pesadas da IGg
têm 4 domínios, sendo que 3 domínios aqui são iguais entre as globulinas Gs, mas aquela
parte lá de cima que eu chamo de Vh. Na cadeia leve eu tenho um domínio variável(Vl) e um
constante (Cl). Então, toda imunoglobulina tem no mínimo isso, então o que faz para diferir
uma da outra?(vejam os slides). Se nós tratássemos em especial a classe de IGg ela sempre
terá a parte constante da cadeia pesada que a gente chama gama, mas a parte variável dela é
diferente de um linfócito PARA outro, essa parte variável é que tem o poder de reconhecimento
do antígeno, é por isso que quando a gente mostra o desenho a gente mostra duas partes. A
cadeia leve é a mesma coisa, a parte C é constante e a parte V é variável, mas a leve difere,
porque enquanto na IGg os bloquinhos constantes são os mesmos: gama, na cadeia leve nós
temos 2 tipos de cadeias. O linfócito pode está produzindo IGg com cadeia leve lâmbida ou
kapa eu tenho 2 tipos de cadeia leve, lâmbida ou kapa , então essa parte cte pode ser lâmbida
ou pode ser kapa, deu para entender isso?
Eu tenho 5 CLASSES PRINCIPAIS:
IGg, IGm, IGb,IGe, IGa. No IGg a cadeia pesada cte é gama, não importa o linfócito,
sempre será gama, na imunoglobulina M por ser uma imunoglobulina que está muito na
superfície dos linfócitos ela tem um outro domínio a mais na cadeia pesada cte, essa cadeia
pesada cte da IGm é chamada de “mi”, também qualquer que seja o linfócito que produza
A cadeia será sempre a “mi”. A parte variável muda de um linfócito para outro, agora
ela e cte para o linfócito, se o linfócito produzir uma determinada parte variável “X” , o resto da
vida e os clones dele irão produzir sempre X. A PARTE VARIÁVEL É ENTRE OS
LINFÓCITOS, MAS ELA É CONSTANTE PARA O MESMO LINFÓCITO!
O mesmo raciocínio é usado para a cadeia kapa, por exemplo. A cadeia kapa é a
mesma coisa, por exemplo, você tem uma parte variável que muda de linfócito para linfócito e
uma parte cte, se um linfócito produzir cadeia kapa ele vai produzir para o resto da vida. Então,
o linfócito pode produzir IGg, gama e a parte cte da leve sendo kapa, um outro linfócito pode
produzir uma outra IGg com a cadeia lâmbida, entenderam? Cadeia leve só tem kapa e
lâmbida, que pode está presente em todas as classe de imunoglobulina citadas a cima. Mas se
um linfócito escolher produzir kapa é para o resto da vida, não tem volta.
Aqui é uma estrutura globular, justamente para vocês terem uma idéia, mas a formação
dela é mais ou menos a forma de um “y”, aqui é para vê quem melhor rege com o que e aí
tratou-se com uma série de proteases e as proteases que melhor funcionaram foram a pepsina
e a papaína, vocês estão vendo do lado esquerdo de vocês aí a papaína que quando ela
quebra, ela só consegue quebrar a região ali mais encima da???/ gerando 2 fragmentos
idênticos com a cadeia leve e pesada e o fragmento de baixo e fazendo-se experimentos se
verificou-se que esse pedaço(parte de cima do Y) tem ligação com o antígeno e esse(perna do
Y) aqui tem ligação como o complemento.
Então, dessas 5 classes principais de Ig, só têm 2 delas que tem mais de um
monômeros, uma delas é IGa que está presente no leite. Então, quando essa Iga está
atravessando membranas, ela está saindo da célula por exemplo, então, quando ela está
atravessando a membrana, ela está sobre essa forma de um dímero. Ela tem uma estrutura de
Y para cima e uma para baixo. Bom, esse componente aqui, chama-se de componente
secretor, e essa cadeia que faz a junção dos Y, cadeia J, não está presente somente nesse
dímero de Iga, mas também na IGm. Então, essas proteínas ajudam, favorecem a passagem
desse dímero através da membrana, depois do lado de fora para atuar, ela volta a forma
monomérica. Mas é um componente secretor que ajuda na transportação. E a outra que não é
monomérica é a IGm, vejam aqui, ela é formada por cinco cadeias, e o que eu disse a vocês?
É a primeira imunoglobulina em qualquer tipo de infecção a ser produzida.
Ufaaaaaaaaaaaaaaa....acabouuuuuuuuuuuu!!!!!!!!!!!!!!!!!!!
Parte 7 – Eduarda Tenório
Então, continuando aqui com as estruturas das imunoglobulinas, as imunoglobulinas G,
D e E são monoméricas, têm aquela estrutura de 4 cadeias polipeptidicas unidas entre si. A IgA
durante o processo de secreção se torna dimérica, em seguida ela volta a forma monomérica.
E essa daí que nós chamamos IgM, que é a primeira a ser produzida e é também essa
estutura, cada monômero desse, é a forma mais comum que você tem na superfície dos
linfócitos. Eu quero que vocês entendam, vamos dizer que nós temos aqui um linfócito, na
superfície deles nós temos as imunoglobulinas que a essa altura do campeonato nós
chamamos de receptores de linfócitos B, enquanto ela está ligada à superfície por uma massa
hidrofóbica, nós chamamos isso aqui de receptores de linfócitos B, quando esses receptores
reconhecem vamos dizer que a gente tem uma bactéria qualquer e temos uma estrutura aqui
que é reconhecida, uma estrutura qualquer aqui que indica que é reconhecida, então ela se liga
e ao se ligar à estrutura que nós chamamos determinante antigênico quando se liga
imediatamente isso faz com que haja uma estimulação do linfócito B e essa estimulação do
linfócito B leva à produção de mais linfócitos B e mais aquelas células grandes q são os
plasmócitos que vão produzir, nesses caso eles vão exportar os mesmos compostos, os
mesmos anticorpos com as mesmas capacidades de reconhecimento desse original só que a
essas horas essa parte hidrofóbica que está dentro da membrana aqui ela não é mais
sintetizada, pq? Porque agora esses anticorpos são feitos para ficarem solúveis, então eles
não vão mais se ligar à membrana, então isso é a primeira coisa que vocês devem lembrar.
Então quando eu falo da imunoglobulina E, é porque esses receptores dos linfócitos, na grande
maioria dos casos essa estrutura monomérica aqui é que vai constituir essa estrutura
pentamérica futura, porque a estrutura pentamérica vai sair a partir da estimulação do linfócito,
quando eu tiver um plasmócito já produtor ele já vai produzir uma estrutura pentamérica
idêntica a original.
O que é que acontece? Quando eu disse pra vocês q eu tenho isso aqui grande, um
parte variável e essas outras partes chamadas constantes, com isso tanto pra cadeia pesada
quanto pra leve, isso quer dizer que uma vez o linfócito estimulado o que acontece? Eu
comecei a produzir aquela molécula pentamérica e qual vantagem? A vantagem é que eu tô
produzindo aquela molécula e ela está rastreando, a parte variável delas é exatamente a
mesma em todas as estruturas monoméricas aqui. Por que se justifica um gasto tão grande pra
fazer uma estrutura pentamérica se o reconhecimento é feito naquelas extremidades? Bem é
de se supor que seja pra ter um melhor rastreamento, ou seja, ela enxerga toda a extensão,
que não é a mesma coisa q uma molécula única, então para um primeiro efeito é bom que seja
ela, mas não é uma questão de duração de dias, se uma infecção qualquer se prolonga por
mais tempo, imediatamente isso é mudado e aquele mesmo linfócito ele passa a mudar a
classe da imunoglobulina.
Então, o que anteriormente, no primeiro estagio, eu estaria fabricando um tipo de
cadeia mi que é a cadeia da estrutura da IgM, eu posso no futuro manter a mesma variável
naquele linfócito, mas só que o linfócito agora, numa segunda etapa da infecção, segundo
estágio mais ou menos, ela continua a ter a mesma parte variável continua idêntica pra cadeia
pesada e pra cadeia leve, mas essa parte aqui vai mudar, então essa parte constante agora
aqui vai ser da IgG, então é gama, então a constante vai ser gama certo!? Então eu posso
dizer o seguinte: um linfócito ele tem poder de reconhecimento apenas para um tipo de
receptor (acho que foi isso que ela disse), mas a medida que o estágio da doença muda, ou o
tipo de patógeno muda, ele vai mudar a classe da imunoglobulina, eu vou mostrar a vocês isso.
Mas eu quero que vocês entendam bem que um linfócito ele é capaz de mudar a classe, ele é
capaz de, por exemplo, reconhecer um antígeno que provoca alergia, mas a gente sabe que
nos estágios de alergia ou nas parasitoses a classe de imunoglobulina que se forma muito é
IgE, então, ai ele pode começar com IgM e passar pra IgE, então a classe da imunoglobulina
pode, o que não pode naquele linfócito mudar é o poder de reconhecimento dele, ou seja, o q a
gente chama de parte variável, para o linfócito, depois dele ser criado é constante, ela varia
entre os vários linfócitos, mas para ele, ela se torna constante.
Pergunta que não dá pra ouvir...
Resposta: Não, a estrutura em si individual e quando a gente chama isso de receptor,
certo? então essa estrutura aqui, essa cadeia pesada daqui, na grande maioria das vezes é a
mi, por isso que eu to dizendo que o monômero dela é IgM, agora pq isso, talvez até porque o
gene dela já fique estimulado a produzir em primeiro estágio a IgM aí por isso seja o fato de
que é sempre mais provável q você tenha a cadeia mi do que a gama ou a eta ou qualquer
outra.
Outra pergunta... Resposta: Ah, aí é que tá, olhe, é uma pergunta interessante, mas é
provável q seja dentro da célula e tenha vesículas exportadoras. Eu nunca vi detalhes de como
essa estrutura pentamerica é formada, mas no espaço extracelular é que não é, com certeza
ela já vem pronta e é exportada como qualquer proteína, agora ela é maior, deve vir em
vesículas, certo?
Essa cadeia J, é aquela mesma cadeia J que a gente vê na estrutura de dímeros da
IgA, também presente aqui. Aí você vê que essa cauda dela, talvez não dê pra visualizar mas é
um pouco maior, ela tem c1, c2, c3 e c4, tem uma estrutura globular a mais, um domínio a
mais.... e aí ela se junta através de pontes dissulfeto.., isso tudo são pontes dissulfeto. Essa
cadeia J é como se ela estabilizasse as estruturas pentamericas.
Bem, solúvel, igM sempre está na estrutura pentamérica, não adianta, ou outros são
monoméricos, mas ela é sempre pentamérica.
Pergunta... Resposta: estimulado pelos patógenos, pelos determinantes antigênicos,
quando ele se liga, ele vai ser estimulado a aumentar a divisão celular.
Pergunta... Resposta: olhe, normalmente o que pode acontecer é o seguinte: se você
tiver (a gente diz muito isso) reação cruzada, por exemplo, carboidrato a gente tem pouca
variabilidade, lembrem que as glicoproteinas sempre têm ácido sialico na extremidade, são
ricas em manose, então a variabilidade é pequena, resultado: a imunogenicidade do
carboidrato é muito alta, se você injetar um carboidrato você produz uma resposta, uma
produção de anticorpos muito alta, estimula muito, mas em compensação ele é pouco receptivo
porque ele pode cruzar reação, você pode ta estimulando pra uma determinada glicoproteina
que você quer X e estar produzindo também anticorpos que reconhecem Y, Z e assim por
diante. Então é por isso que eu posso dizer pra vocês o seguinte, ele reconhecendo o antígeno
que seja próprio do reconhecimento dele, ele pode vir a reconhecer antígenos semelhantes e
assim como as enzimas que eu tava falando a pouco tem uma exocinase da vida, ela
reconhece hexoses em geral, mas tem maior especificidade, ou maior afinidade por um
monossacarídeo do que outro, a mesma coisa pode acontecer aqui, ela poder ser estimulada
por um, mas ela pode vir a reconhecer alguma coisa semelhante.
Pergunta... Resposta: não, é isso que eu quero lhe dizer, talvez eu não tenha sido
clara, se ele vier a encontrar por acaso algum determinante antigênico idêntico ao que
estimulou ele, pode estimular. Só que a essas horas já tem... é por isso que eu disse a vocês
que a resposta é rápida da segunda vez que é visto aquilo, porque o segundo antígeno, que
seja outra, mas que tenha identidade química igual ao da primeira estimulação e que venha a
estimular ele, em vez de ele ter uma pra ser estimulado ele tem centenas de linfócitos na
memória, e ai ele responde mais, é porque a gente vai criando uma certa resistência a doença,
porque se não a gente morria com muita facilidade, então uma criança ela pega muito mais
fácil esse negócio dessas doencinhas de criança papeira, caxumba, depois q você tem, você
não tem mais e você fica mais resistente, vocês vão lidar muito em hospitais com gente doente
e nem por isso vocês vão adquirir a doença.
Pergunta... Resposta: entendi o q você quer, você quer dizer que esse aqui
permanecesse digamos na condição de virgem ainda, não, não, porque o que vai estimular ele
é a ligação com o antígeno e a ligação com o antígeno é provocada quando essa partezinha
daqui dita variável ficou de tal maneira que quimicamente ela interage com outra, certo? Então
este daqui que já foi estimulado... a população de memória, ele já foi estimulado, portanto ele
não pode se tornar virgem de novo, perdeu a virgindade, aí não tem mais. Então nesse caso
você pode voltar a produzir, o que ele quer dizer é mais ou menos por aí. Veja, você pode
voltar a estimular no bom sentido, se você novamente infectar pelo mesmo agente e ele tiver
nessa condição aqui cheio de receptorezinhos, certo? Mas é a sua população de guarda. E o
patógeno já desapareceu, já foi combatida a infecção, tudinho, aí você tem centenas deles
idênticos, se o patógeno volta, na hora que ele vai se ligar e vai rapidamente proliferar, então...
se expande, mas ele já foi de certa forma estimulado.
Pergunta... Resposta: é pq ele já é construído pra reconhecer só aquilo, entendeu? Só
um determinado tipo químico de coisa, então quando você modifica aquilo, já é outro que vai
ser estimulado. Pergunta... Resposta: Quando é mto parecido vc pode ter aquilo que eu disse,
uma menor afinidade, porque não é igual, mas pode estimular. E a resposta policlonal que eu
disse é exatamente isso. Vamos supor que você tenha aqui uma bactéria qualquer e um DNA e
então você tenha um determinado composto aqui, mas tenha uma outra estrutura química
diferente e tenha uma terceira estrutura química aqui que eu posso dizer que seja um triangulo
aí qualquer, certo? Então são três diferentes, se o linfócito de numero 1 for estimulado, então o
anticorpo dele que reconhece isso daqui reconhece isso, então isso daqui é proveniente do
linfócito 1, esse outro que é diferente quimicamente desse, então é um outro linfócito
estimulado, então o linfócito 2 vai produzir contra esse e novamente o linfócito 3 que foi
estimulado vai produzir contra este, por isso, embora cada um desses linfócitos vá fazer uma
população igual, que a gente diz o clone, a resposta é policlonal e claro que é de se esperar,
por isso a interação quimica que pode ser que a interação do anticorpo 1 produzido pelo
linfocito 1 seja mais especifica, mais forte quimicamente falando do que a do 2 ou a do 3, então
essa resposta policlonal é muito boa, porque digamos que ao longo do tempo por alguma
modificação conformacional e genética que você adquira, aquele linfócito não responda mais,
você tem populações de outros linfócitos que foram criados de memória também, entendeu? E
pode acontecer isso, você pode perder..
Alguém interrompeu com uma pergunta... Resposta: não, no sentido já foi estimulado,
no sentido de que pode ter uma estimulação, ele pode, porque toda vez que você ligar alguma
coisa q seja reconhecida por essa parte variável você tá estimulando o linfócito a se diferenciar,
certo? Mas o q a gente quer, que é a mesma coisa q a base da vacina que a gente tava
dizendo, é que essa população de linfócitos já fique predisposta a reagir de imediato quando
verificar algum determinante antigênico idêntico. Vamos supor que aqui seja uma seqüência
que depois um determinado vírus qualquer aqui tenha, então vamos supor q essa seqüência
aqui seja exatamente igual a esta, olha se ele já foi estimulado, se ele já tiver, você vai ao
longo da vida adquirindo cada vez mais resistência a infecções pq você vai criando no geral
uma população de linfócitos infectados e conseqüentemente uma população de células de
memória, entendeu?
Pergunta... Resposta: num mesmo linfócito a parte variável que é de reconhecimento,
pra ele é constante e foi isso que eu tava dizendo a pouco não foi? A diferença é isso, eu
chamo de variável entre os diferentes linfócitos, mas quando aquele linfócito tem aquela
variável, pra ele, pro resto da vida dele aquilo vai ser igual, então é constante pra ele, por isso
que eu to dizendo a vocês: pra ele voltar a ser estimulado é preciso que o determinante
antigênico seja igual.
Pergunta... Resposta: não, ele pode fabricar 2 classes diferentes, é diferente. Veja,
deixa eu tentar explicar isso. A parte variável aqui é de reconhecimento, a parte constante,
deixa eu desenhar aqui, daqui pra baixo é c1, c2 e c3, constante, certo? Então essa parte
constante, eu tenho várias constantes, se a imunoglobulina for G a constante é gama, se a
imunoglobulina for M a constante é mi, se a imunoglobulina for A é alfa, se for D é delta, se for
imunoglobulina E é eta. Então a parte constante para cada classe de imunoglobulina dessas é
diferente, então um mesmo linfócito não muda a parte variável, mas essa outra que é a parte
constante o linfócito pode mudar dependendo do estágio da doença, no inicio ele fabrica mi,
depois ele pode mudar pra gama, se for um parasita ele vai aumentar muito a produção de eta
(acho que foi isso que ela disse) e assim por diante, isso a gente chama mudança de classe.
Pergunta... resposta: é um gasto muito grande, mas por outro lado o poder de
reconhecimento de imediato é maior.
Parte 8 – Etiene Andrade
Pergunta ... Resposta: Isso é que nunca encontrou o estimulador dele que é o
antígeno. Ele só vê aquilo. Você muda até a subclasse IgG quanto mais a subclasse. Então
isso é porque se adapta melhor, reagem melhor, deve ter uma explicação. Cada patologia vem
sendo estudada...
Agora vamos ver porque a gente tem a diversidade de anticorpos, vamos estudar isso,
aí que entra a recombinação. A localização cromossomal das cadeias que formam a
imunoglobulina é diferente, então a gente tem no cromossoma humano e em alguns animais
que já foram estudados, a gente tem a localização e varia de animal para animal. Aí vocês tem
dois exemplos em humanos e em camudongos (melhor acompanhar olhando os slides). A
cadeia que tem a ----- e a lambda, no caso de humanos a cadeia lambda ela é formada o gene
produtor dela está no cromossomo número 22 e a cadeia ---- de número 2 e a cadeia pesada
toda ela com as possibilidades de a gente fazer mudanças de classe está no cromossomo de
número 14 em humanos.
Se vocês olharem por exemplo o camundongo já muda é 16, 6 e 12 é diferente, então
para humanos que é o que interessa você tem 3 cromossomos responsáveis pela produção
dos 3 tipos de cadeias que nós vamos gerar nossas imunoglobulinas. Aí vocês vão dizer tem
uma diversidade tão grande e só tem 3 cromossomos. A célula só com esses 3 cromossomos
faz todas as imunoglobulinas que nós precisamos com algumas variantes ainda. Vamos olhar o
processo de recombinação.
O que a gente chama de recombinação? Isso aqui é uma seqüência de dna que
formara a cadeia lambda, aqui uma seqüência de dna que originará a cadeia ---- e aqui a
cadeia pesada. 14, 22 e 2 ou 2 e 22. É uma seqüência bem grande que você tem e daqui você
vai tirar todas as imunoglobulinas que você precisa. Então os estudos que foram feitos não em
humanos mas em rato, camundongo que aí você consegue tirar o dna, estudar...
Então os estudos experimentais são mais em animais experimentais.
Na cadeia --- você tem ao longo do cromossomo seqüências que tem um certo número
de pares de --- separando uma da outra e a conseqüência disso no camundongo, foram
identificadas 300 seqüências. Então espalhadas ao longo dos cromossomos nós temos essas
seqüências que a gente chama de V. V de variável. Elas vão... elas tem uma parte constante
esse l que tem na frente a gente chama de seqüência líder que também é separada da v por
um trecho menor de segmento de dna então lvlvlvlvlv ao longo do cromossomo. Quando a
célula precursora aquela célula da medula óssea essa célula percussora no seu dna quando
você vai diferenciar para surgir diferentes células e o lifóncito vai ser formado, vai ser escolhido
de certa forma uma das seqüências das 300 seqüências pra ser recombinando bem mais
adiante com outro grupo de seqüências de bases chamado j. Então essa seqüência vai ser
escolhida normalmente em camundongo nós temos cinco seqüências de 300 seqüências de lv.
vamos supor que eu pegue uma seqüência dessas lv e eu vou aproximar de uma j então
quando eu faço esse processo que eu chamo de recombinação pra criação do dna do linfócito
e a toda essa parte aqui aproximar o v dessa j escolhido aqui. Então o dna agora que está
presente no linfócito não contém mais esse trecho entre o v escolhido e j escolhido. Isso é uma
recombinação porque se forma como se fosse um anel no que você aproximou esse trecho
aqui que era o v com l.Aí eu botei junto do j que eu quero, ai eu aproximei. Eu vou perder todo
esse trecho que esta aqui, e se eu decidir combinar o de numero 300 com o j primeiro você so
vai perder um intervalo aqui, ai fica o resto fica mas ai eu só vou transcrever em uma cadeia
polipeptídica, o que combinou o resto vai ser esquecido. Quem é j? Quando você tem uma
cadeia leve vai ser formada por duas partes uma variável e outra constante, e aqui nessa
variável tem um j então na cadeia leve eu tenho mais ou menos cento e vinte cinco
aminoácidos na parte variável e 125 aminoácidos na parte constante. Então a parte variável é
formada por dois genes, o v e o j. Porque eu coloco j no meio?
Porque me dá mais poder de reconhecimento, se eu combinar meus 115 com 10 aa
que tenha diferença do j1,j2,j3. Isso me da mais poder de reconhecimento só tem uma coisa na
hora que o linfócito for formado com determinado j é pro resto da vida. Mas eu posso também a
partir da mesma linhagem eu posso combinar o v com j1 ou com j2 ou j4, então é um processo
de recombinação.
Eu teria 300 seqüências v na cadeia ----que posso combinar com 5 seqüências j, que
eu posso dar origem a 1500 seqüências diferentes. Na cadeia pesada isso aqui é c1,c2 então
depois agente, depois agente ver. Na cadeia pesada eu tenho j aqui, eu tenho trecho d e um v.
Então na cadeia j que tem mais ou menos o mesmo numero de aminoácidos eu vou ter duas
recombinações, veja só o detalhe esses lv aqui são em numero de mil, d 12 e j aparentemente
5 ou 6 . Então recombinação desses trechos 1000X12 pra que posso recombinar com
d1,d2,d3,d4 ate d12, e vezes j que vamos dizer que seja 5. Então quanto seria 60 mil. Então eu
vou ter 60 mil X 1500, porque eu vou juntar a leve com a pesada, então eu posso juntar 60 mil
com 1500, que vai dar 9000000. Aqui você tem duas recombinações o v se aproximando do d
e o d se aproxima do j e estão próximo do c, então o c leva sua parte variável já presa a
constante eu posso mudar depois, mas a parte variável vai junto, é um processo de
recombinação. No caso da cadeia lambda é diferente da ------ não no sentido que tem a mesma
seqüência de recombinação porque ele tem de constante possibilidades também de variar a
constante deles. Então ele pode fabricar mais de uma constante coisa que aqui no caso dessa
mais simples que é a -----, não faz porque so tem uma região então esses aqui tem a constante
variável. Eu tenho uma cadeia leve a linha de células precursora onde eu tenho o dna da
cadeia ----------no linfócito depois eu tenho a formação do rna primário depois eu tenho, tenho
a formação do rna mesnsageiro. Como vocês sabem todos os eucariontes tem intróns então
eles vão ser processados pra formar o mensageiro e depois do mensageiro depois a gente tem
o polipeptídeo nascente e a cadeia ----- perde esta seqüência livre.
Lá em cima eu tenho um trecho de dna com seqüências lv depois seqüência j e a parte
constante quando eu recombino criei, rearranjei a seqüência de tal maneira que agora eu vou
constituir o meu linfócito. Qual é a seqüência que o linfócito escolheu? Esta seqüência aqui que
ta marcada para aproximar do j, ele escolheu aquele penúltimo j aqui. O linfócito tem este dna,
não interessa o que vier antes, não rearranjou não transcreve. Quando chega aqui que eu já
tenho o dna pronto ocorre a transcrição. Então a seqüência líder começa a ser transcrita a
partir dela até o g. Eu tenho um rna que não é o rna mensageiro, portanto vai precisar ter uma
etapa de processamento, ou seja, retirada das partes não codificadoras, os chamados intróns.
Isso é feito já depois desta síntese com a retirada deste trecho que fica entre este j escolhido e
o c. E esta seqüência líder também tem um intrón aqui que aproxima. Então ficamos com isso.
O que é isso? O rna mensageiro. Quando este rna mensageiro é traduzido em proteína, você
então tem um polipeptídeo nascente que é totalmente produzido a partir desta informação no
rna mensageiro completo, só que a nossa cadeia não tem essa seqüência líder. A seqüência
líder admite-se que ela seja direcionadora durante todo processo, quando a proteína está feita
não interessa mais ficar com essa seqüência líder. Ela não faz parte realmente da cadeia ---nem fará parte das outras. Então isso daqui constitui o peptídeo que é formado por v. Quem é
v? é formado por uma seqüência de aminoácidos originados a partir dos genes v e j e aqui eu
tenho a parte constante da cadeia.
Pergunta?????
Resposta: Nesse aí ele recombinou o l e o v, recombinou lv 23 com j4
Pergunta????
Resposta: Ele não volta não... A parte que não recombinar e tiver anterior a ele
continua lá, mas não tem mais capacidade de produzir, ele só tem condição de produzir o que
recombinar com o j. o que não recombinou não tem mais. O linfócito não faz o rearranjo o
problema é este. Quem faz o rearranjo é a célula precursora para originar o linfócito, mas
depois de ele feito ele não faz mais rearranjo molecular para gerar novas diversidades
Parte 9 – Fernando Jorge
O que o outro faz na cadeia pesada? Coisa muito semelhante, só que com um (Felipão
interrompe e n da pra escutar). O Dna lá de cima é dela, a precursora, ai quando tira já vem
recombinado (Felipão de novo) rearranjo molecular. Isso é dado em genética, forma aquilo que
eu disse assim, forma como que uma bolsa de enzimas chamadas recombinases que fazem
isso: cortam e ai o DNA é unido. Tam enzimas que fazem todo esse processo, mas como
vocês vêem a aula não dá pra dar detalhes. Bem, ai entra a cadeia pesada. A pesada, ela pesa
um pouquinho mais, pra entender somente a parte de cá, mas o resto é igual. Ta lá em cima
novamente a célula precursora, o DNA bem grande e com toda a informação. Vejam que nesse
caso além da seqüência líder ( eu acho) e (outra que eu n consegui entender) eu tenho a “d”
que, fica aqui no meio, e tenho a “j”. então esse “d” a gente chama diversidade. Por que?
Porque na medida que eu combinasse mil com cinco eu teria cinco mil diferentes cadeias, mas
se eu criei mais uma zona de diversidade, eu invés de cinco passo a ter setenta mil. Então isso
aqui age propositadamente para aumentar a diversidade dos anticorpos gerados. Então
quando a gente faz na transformação, passando da célula precursora até chegar ao DNA
primário, que é o RNA da cadeia pesada, você tem duas recombinações , você vai aproximar o
...... e o “d” do “j”, ou seja, no início, ao que tudo indica, primeiro o “d” e o “j” se juntam, e o “d” e
o “j” já juntos teria a recombinação na ordem lógica dos passos o “d” se aproxima rearranjando
primeiro d-j e depois o reajanjo....... Pronto, formado o DNA do linfócito, você tem essa
seqüência, novamente é valido a mesma coisa, o que não tiver re combinado não transcreve.
Então vem pra cá. Mas vejam que nesse DNA do linfócito eu ainda tenho introns aqui. E ainda
tenho um passo a mais: entre essas seqüências toda aqui, qu vão me originar a parte variável
da cadeia, eu tenho pra cá vários trechinhos aqui que representam o que? Cadeia mi ( letra
grega), seqüência da cadeia mi, a cadeia delta, gama, gama 3, gama 1, gama 2b, gama 2a, eu
falei pra vocês que tinha dicidido em classes, eta, e alfa. Então cada seqüência dessa vai ser
levada para o linfócito, porque aí é o DNA do linfócito. A variável já ta feita, mas as
possibilidades de eu mudar a classe das imunoglobulinas continua existindo no DNA do
linfócito. Ai quando o linfócito vai ser produzido o que é que acontece? Eu vou ter um RNA
primário transcrito, e nesse caso, ele ta mostrando aqui, ou você faz com um ou faz com outro.
Então vamos dizer que de início eu vou transcrever esse daqui. Então quando eu transcrever
eu tenho um mi aqui, né? Então eu transcrevo, processo pra tirar os introns e crio esse RNA
aqui, ou, do contrario, eu tenho o gana aqui e transcrevo. Então isso vai depender, como eu
disse a vocês, de alguma mensagem de alguma coisa ainda não muito bem explicada de quem
é que regila essa mudança de classes. Porque essa mudança de classes, como eu disse, você
pode produzir IgG de subclasses diferentes dependendo da patologia. E o que é que explica
isso? Não sei. Agora que tem diferenças, tem de uma doença pra outra e não interessa, por
exemplo, a doença. Você pode mudar o paciente, mas se uma determinada patologia induz à
produção de um tipo de IgE, Por exemplo, pode ser outro paciente mas ele vai ter uma
alteração no IgE se a patologia for a mesma. Bem, então feito isso vai ter a formação do
polipeptídeo depois sai essa parte líder e aí eu tenho a proteína. Então nesse caso você tem
mudança de classe, portyanto lembrem que na mudança de classe o linfócito pode fazer, só
não pode mais mudar a parte variável dele, mais nunca. Vamos ver agora aqui uma coisa
interessante: olhe uma coisa ainda que muita gente diz: a diversidade antigânica é só a única
responsável pela diversidade antigânica é o problema de rearranjo molecular? Eu posso dizer a
vocês que não. Hoje em dia já se sabe que as células, o próprio linfócito pode se melhorar a
combinação dele se ele começar a enxergar de diferentes maneiras o antígeno. Então, como
ele pode melhorar? Hoje em dia a gente sabe que no ponto de corte você pode ter aqui o
finalzinho do “v” o começinho do “j”. você vai recombinar. Essas terminações aqui, as
seqüências de bases nitrogenadas que esse DNA tem, tem determinadas possibilidades que
ele pode, por exemplo, vamos dizer que eu tenha nessa terminação aqui dez bases e dez
bases aqui. Eu posso interagir fechando isso, combinando oito bases daqui com dez daqui, ou
eu posso botar dez daqui com cinco daqui. Entendeu? Então essas pequenas possibilidades
de você variar essa recombinação que voe faz no ponto de ligação mesmo, são pequenas mas
podem gerar um, dois aminoácidos diferentes ali e que esse um dois aminoácidos diferentes ali
favoreçam às vezes a melhorar a resposta imunológica de um determinado anticorpo. É
mínimo, você diz, mudou um aminoácido só. Mas é um só, a gente vê em enzima, você vê que
um só aminoácido, se for essencial pro sítio ativo, ele pode simplesmente inutilizar a enzima,
ela não tem mais atividade, vocês sabem disso. Então basta uma mutação pra você ter isso.
Então nesse caso essas terminações, o que ta sendo mostrado aqui, depois eu quero que
vocês leiam, é que essas terminações ai possibilitam ainda que haja pequenas modificações
na seqüência da variável daquele linfócito. E isso aumenta ainda mais o poder de fogo dos
anticorpos.
(pergunta q n da p entender) a célula disso? Não. Já é linfócito. Eu to chamando o
linfócito disso porque ele ainda não encontrou o padrinho dele, o anticorpo. (outra pergunta).
Ele já tem sim, já ta nessa posição. Quando o linfócito é criado mesmo que ele ainda não tenha
ou não venha a encontrar na vida todinha os anticorpos, ele já existe com a conformação
própria de produção de uma variável. (mais pergunta). Não, tem que ser pelo específico. Né
por isso que eu digo que a resposta adaptativa é em conseqüência da estimulação pelo
antígeno. Então não existe resposta adaptativa se você não tiver estimulação pelo antígeno.
(perguntas). Eu chamo de virgem porque eu quero dizer assim: é o linfócito que ainda não
encontrou o antígeno que ele reconhece, por isso que eu to dizendo virgem. Ele ainda não
identificou nada que apareça lá. Então é nesse sentido que eu to dizendo que ele ainda é
imaturo no sentido de produzir memória, etc. ele existe lá, ta apto a de repente aparecer um
corpo estranho lá qualquer que ele reconheça, mas enquanto ele não reconhece a gente
costuma usar esse termo, chamar ele de linfócito virgem. Quer dizer: ele ainda não foi
estimulado.
Parte 10 – Filippe Moura
Filipão (ou seja, eu... é meio estranho falar de mim na 3ª pessoa, mas vamo lá...) faz
uma pergunta que é basicamente o seguinte:
- Professora, o que é que vai sinalizar inicialmente para o linfócito que tipo de anticorpo
vai ser o produzido por ele?
Primeiramente ela não entende a pergunta e responde outra coisa. Repete-se a
pergunta e ela responde:
- Ah! Isso não tem... é o que eu disse no início, você cria uma população de linfócitos,
ao nascer, que foram originados de vários processos de recombinação e acredita-se, pelo que
se estudou até hoje, que tenham essas possibilidades que eu disse a você. Porque foram
encontrados na parte variável, no VL de camundongos, mil seqüências daquela e admite-se
que você pode recombinar mil com doze. Espera-se que seja isso, agora dizer a você o que é
que ordena esse processo de recombinação, agente não sabe.
Filipão:
- Porque eu pensava que era uma resposta, entendeu?
Profa.:
- Não! Veja, a resposta aparece já no linfócito B, mas pra você gerar população com
diversidade capaz de identificar diferentes antígenos, isso é aleatório. Eu não sei quem regula.
Alguém pergunta (acho que Júlio):
- Não é o contato com o antígeno que regula?
Profa.:
- Não. O contato com o antígeno vai estimular ele a produzir o plasmócito, porque o
contato é feito através desses receptores de superfície.
Outra pergunta:
- Como é que os receptores vão conhecer os antígenos?
Profa.:
- Mas ele não precisa conhecer, ele precisa é ter uma estrutura química capaz de
interagir com a estrutura química do antígeno. É tudo químico.
Pergunta que eu não consegui decifrar.
Profa.:
- Certo, pode haver! Aí é que é o problema. Não tem gente que morre de doença autoimune? É porque perde essa capacidade de reconhecer.
Outra pergunta que não dá pra entender (acho que é sobre a infecção por um antígeno
que não tenha receptor compatível nos nossos linfócitos, agente morreria?).
Profa.:
- Olhe, teoricamente morreria; na prática você tem ainda processos de alteração
somática nas células que pode melhorar a especificidade. Então, é de se esperar que os
organismos tenham algum tipo de reação que possa melhorar essa interação, mas não é mais
por rearranjo. Pode ser por mutações, alterações de alguma coisa, ou até, sei lá, no tipo de
ligação, de intron que você possa tirar. Deve ter alguma coisa porque já são conhecidas
reações para aprimorar a resposta imune. Agora, o que se espera realmente e o que se vê é
que agente tem uma infinidade tão grande de diferentes linfócitos que agente reage, não é?
Você vê que dia a dia agente está em contato com bactéria, com todo o tipo de patógeno e
ninguém está morrendo por causa disso.
Complexo Principal de Histocompatibilidade
- Em humanos agente chama de HLA. O complexo principal de histocompatibilidade,
MHC por causa do inglês (major histocompatibility complex), quer dizer o que? Todos nós
temos na nossa superfície celular receptores que são capazes de externalizar proteínas que
foram internalizadas. Que proteínas seriam essas? Somente fagócitos poderiam ter esse
complexo? Não. Qualquer célula não pode ser infectada por um vírus? Não é? Têm vírus que
infectam células da pele, do pulmão etc. Teoricamente todas as nossas células têm
possibilidade de serem infectadas por patógenos. Você tem tipos de parasitas, como o Cruzi,
que ele adere na superfície da célula cardíaca, entra e se multiplica, como se fosse um vírus. É
por isso que as pessoas que têm doença de chagas incham o coração e acabam morrendo por
insuficiência cardíaca. Por que? Porque agente até hoje ainda não descobriu uma maneira de
parar essa invasão.
Como teoricamente agente tem qualquer célula com capacidade de ser infectada por
um patógeno, a célula tem que, no caso de ela ser infectada, sinalizar, porque quando o
patógeno está no fluido do sangue é fácil; os linfócitos estão lá, circulando, identificam, fazem
interação, são estimulados e combatem. Mas quando está dentro da célula, como é que esta
célula poderia sinalizar? Essas células todas do nosso organismo têm proteínas de superfície,
que conjuntamente agente chama complexo principal de histocompatibilidade, que
externalizam o que for internalizado. Quando você internaliza, por exemplo, um vírus, o que
acontece? Você vai usar a maquinaria da célula para formar novas partículas virais. Só que
dentro da sua célula você tem enzimas que são capazes, parcialmente, de degradar o vírus.
Então no momento que está sendo degradado, aquela célula infectada, através desses
receptores do MHC, vai externalizar um fragmento daquilo. Essa é a forma que as células têm
de mostrar ao exterior que algum elemento estranho está nas imediações.
Agente tem dois tipo de proteínas de histocompatibilidade. Uma é a classe 1, que
aparece em qualquer tipo de célula e que normalmente externaliza materiais virais, ou seja,
materiais obtidos endogenamente. Por que eu digo endógeno? É porque quando uma célula é
infectada por vírus, a própria maquinaria celular produz proteínas e essas proteínas que já
foram produzidas dentro da célula serão degradadas parcialmente. E então o que você vai
externalizar são materiais que já foram produzidos pela célula. Nesse caso, a classe 1 desse
complexo é quem externaliza. Aí tem a outra classe de proteínas, que agente chama classe 2.
Esta não está em qualquer tipo de célula, ela está somente nas células que agente chama
apresentadoras de antígeno, mas na realidade são células fagocíticas ou linfócitos. O linfócito
também tem a classe 2 porque quando ele interage, parte do antígeno é internalizado para
estimular a divisão celular. O macrófago, por exemplo, quando ele internaliza o que é que ele
faz? Ele sintetiza novas partículas? Não. Ele internaliza o microorganismo por fagocitose e
dentro da célula ele quebra, através das enzimas lisossomais, aquele material que foi
internalizado mas que é externo e o que ele mostra não foi o que ele fabricou, é o que é
externo mesmo, que foi somente fagocitado. Então a classe 2 só aparece nas células do
sistema imune que têm capacidade de mostrar esses antígenos, de fagocitá-los para poder
mostrar. A classe 2 só está presente nas células apresentadoras de antígeno.
Pergunta.
Profa.:
- Todas as duas têm. A classe 1 está em todas as células, inclusive macrófagos. Se
você internaliza um vírus você não vai produzir novas partículas dentro da sua célula? Então a
classe 1 é a que pode mostrar antígenos que foram produzidos pela célula, embora que a
mensagem tenha vindo de um vírus.
Pergunta.
Profa.:
- Qualquer célula. É aí que cai a questão da rejeição. Você tem um padrão de MHC
seu, que é diferente do meu e de todos os seus colegas, a não ser que você tenha um gêmeo
univitelino. No momento que você tem padrões diferentes, se você pega um órgão qualquer,
um fígado meu, eu morri e você leva ele e bota em você, porque o seu está ruim. Na hora que
você fizer esse transplante a superfície das minhas células, que têm esse complexo (alguém
pergunta “é o glicocalix é?”). Está presente junto com o glicocálix mas não é. São proteínas
transmembrana. Então quando o órgão é transplantado, como o seu padrão de proteína é
distinto, você vai ver o meu padrão diferente do seu. É por isso que se chama complexo de
histocompatibilidade, porque é preciso haver compatibilidade para isso não ser visto como
antígeno de histocompatibilidade. Por isso quando vai fazer o transplante faz-se um teste de
histocompatibilidade. Às vezes são parentes próximos, mas às vezes há compatibilidade entre
pessoas, familiarmente falando, diferentes mas com padrões muito semelhantes.
Parte 11 – Felipe Mendonça
Pergunta...
Resposta: Nesse caso, das quatro, funcionariam como antígeno, mas aqui eu to
falando só da preservadora (o áudio começa com essa resposta, que eu não entendi nada)
Pergunta (alguém pergunta algo sobre rejeição e medicamentos utilizados para
diminuir a rejeição)
Resposta: não, não, não, ele vai atuar diminuindo mais a produção de citocinas,
bloqueando a sua expressão gênica, e de uma série de coisas pra evitar que o sistema imune
desenvolva aquela cadeia de reações que nós estamos vendo.
Pergunta de novo sobre isso (sobre a diminuição da imunidade com a utilização desses
medicamentos)
Resposta: Fica sim com imunidade baixa. (Aqui ela começa a contar a história de uma
conhecida que morreu depois de receber um órgão transplantado)
Pergunta ainda sobre esse tema
Resposta: É, tem que ficar tomando pro resto da vida, por isso que transplante só se
deve fazer como última opção.
(Ela começa a conversar sobre a época que ela trabalhou no hospital e começa a
discutir sobre a validade de se utilizar como parâmetro de morte a ausência de atividade
cerebral e tal, sobre tachar uma pessoal que não tem mais chances de sobreviver etc)
Pergunta (Filippão pergunta algo)
Resposta: Veja só, esse complexo principal de histocompatibilidade (MHC), a classe I e
a classe II, são formados por proteínas distintas. A classe I tem uma longa proteína aqui,
transmembrânica, chamada cadeia alfa, que aqui você tem a terminação carboxílica e aqui a
terminação amina, como a maioria das proteínas transmembrana o grupo carboxila na porção
citoplasmática e o grupo amina na porção extracelular. Percebem que essa cadeia alfa
apresenta três domínios: domínio 1,2 e 3. Ainda no MHC de classe I, além da longa cadeia
alfa, temos também, apenas no lado extracelular da membrana, uma proteína que forma a
cadeia beta, sendo chamada de Beta-2 microglobulina. Esse complexo MHC é sintetizado,
acredita-se, em conseqüência da presença, dentro da célula, dos vírus e fragmentos virais.
Isso levaria a transcrição e produção dessas proteínas. Essas proteínas, por sua vez, trariam,
ligados aquela parte ali de cima que vocês tão vendo (precisa acompanhar os slides),
fragmentos que seriam externalizados do lado de fora da célula.
Isso tudo no MHC de classe I. Já no caso do MHC de classe II, que está presente
apenas nas células do sistema imune que apresentam antígenos e atuam em processos de
fagocitose, ela é constituída por duas cadeias transmembrânicas diferentes. Tanto a cadeia
alfa como a cadeia beta da classe II tem dois domínios extracelulares. A região apresentadora
de antígeno localiza-se entre alfa1 e beta1, ao contrário do MHC de classe I que tinha essa
região entre os domínios alfa1 e alfa2. Esse material (o MHC) é sempre estimulado (no caso, a
produção) pela presença de antígeno dentro da célula, para ligar-se a este e apresentá-lo.
Vejam nesse slide que eles estão mostrando justamente isso, ali é a região onde o material
(antígeno) é exposto. Essa região de mostrar o fragmento é variável, assim como a gente viu
no caso das imunoglobulinas, que a parte superior pode ter recombinações para gerar
diversidade. São duas as razões para a existência dessa variabilidade na região apresentadora
do MHC: cria ,de certa forma, um perfil individual próprio e pode-se mostrar ,dessa forma,
diferentes materiais antigênicos. Então da mesma maneira que nós temos variabilidade nas
imunoglobulinas, nós temos na porção apresentadora do MHC, o que faz com que apenas
antígenos quimicamente compatíveis com a região de um determinado complexo sejam
apresentados.
Pergunta (algo sobre qual/quais a(s) classe(s) de MHC presente(s) no macrófago)
Resposta: O macrófago tem os dois, tanto classe I como classe II. E por que será que o
macrófago tem classe I e classe II, hein? Tem classe II porque ele é uma célula do sistema
imune que fagocita e apresenta antígenos e tem classe I porque ele é uma célula normal e
pode ser infectada por um vírus qualquer. Se lembrem que as células que não são fagocíticas
têm apenas classe I.
Pergunta de novo.
Resposta: A diferença entre classe I e classe II é que a primeira apresenta o que é
produzido pela célula (induzido por uma infecção viral, por exemplo) e a segunda apresenta o
que é endocitado e degradado pela célula.
Aqui para vocês entenderem, eu tenho um exemplo para mostrar como a coisa
funciona. Eu tenho uma célula qualquer e um vírus penetra nela. Uma vez penetrando esse
vírus, independendo da maneira que ele penetrou (se ele foi endocitado ou simplesmente
injetou o RNA, não interessa), vai utilizar o DNA ou RNA dele pra produzir novas partículas
virais. Essas proteínas que vão compor as novas partículas virais, parte delas, de fato, vão se
juntar para formar novas partículas, no entanto, uma outra parte sofre proteólise devido a
enzimas endógenas presentes no citoplasma que permanecem degradando essas proteínas
virais. Esses fragmentos peptídicos gerados vão ser expostos, externalizados por esse MHC
aqui que só tem um pezinho, o MHC de classe I. No momento em que essa célula está atacada
por um vírus e esta externalizando fragmentos, ela é reconhecida por um tipo de linfócito.
Esses linfócitos, que são do tipo T, eles tem receptores de superfície. Esses receptores são
proteínas com duas cadeias externalizadas, com porções transmembrânicas, mas totalmente
diferentes das imunoglobulinas. Botem na cabeça que linfócito T não produz nada de receptor
pra ser secretado. Ele interage célula à célula, diretamente. Então o linfócito T que tem
características de reconhecimento para aquele material externalizado, interage com esse
material. Uma vez interagindo, ele vai produzir citocinas. Esse linfócito que interage com
células infectadas é chamado de linfócito T citotóxico, ou simplismente linfócito TC. Esses
linfócitos T citotóxicos produzem citocinas, que levam a célula à apoptose. Esse processo
funciona como uma tentativa do sistema imune de reverter a atuação desse vírus em outras
células. Somente o linfócito citotóxico vai atuar nesse quadro? Não. Os fragmentos gerados
pela apoptose da células infectada vai atrair toda a gama de células do sistema imune,
inclusive o linfócito B também. Então a gente não pode excluir essa atuação em conjunto do
sistema imune. O linfócito citotóxico foi responsável por reconhecer... (alguma coisa bate no
gravador e não dá pra ouvir o final da frase).
Pergunta: O que impede as citocinas liberadas pelo linfócito TC de atuar nas células
vizinhas não infectadas? (Deu pra colocar a pergunta toda porque fui eu quem perguntei aí me
lembro : P)
Resposta: Ah, existem uma série de citocinas, não é uma só não. Algumas levam a
estimulação desse próprio linfócito T, outras levam a uma maior interação, outras ligam-se à
membrana para perfurá-la, tipo perforina. Tem muitas citocinas em jogo, não é uma única só
não. A produção dessas citocinas leva a célula a estourar.
Pergunta de novo (merma pergunta por que ela não respondeu o que impedia)
Resposta: Isso, ela vai atacar as infectadas, porque precisa dessa interação. Só que
lembrem que ao estourar essa célula um monte de outras células são encaminhadas pra lá. O
linfócito B canalizado para lá, os complementos etc e tal.
Pergunta de novo (merma coisa porque ela não explicou ainda)
Resposta: Ele só produz citocina devido a exposição de fragmentos pela célula
infectada...
Pergunta de novo (merma ladainha ainda e continua esse jogo de pergunta e resposta
até que Fred entra no jogo e tenta explicar pra ela a pergunta de fato)
Resposta: Ah, é porque ele (linfócito TC) lança geralmente na superfície e porque na
célula infectada tem receptores próprios para interagirem com essas citocinas. Tudo isso é na
base da atuação em receptores que limitam a ação em outras células.
MUITO BARULHO, NÃO SEI SE É PERGUNTA OU O QUE É (escutei Betinha falando
algo do tipo “tem que ter paciência, paciência”)
VOLTANDO AO SILÊNCIO DE NOVO...
(Ela fala algo do tipo quanto é cansativo tá escutando ela durante 4h seguidas e diz
que entende a gente e falou já com Eduardo, mas ele disse que não dava pra fazer nada:
Betinha é nossa heróia! \ o /)
No caso desse slide daqui, a gente tá vendo uma célula apresentadora de antígeno.
Uma proteína é internalizada, é degradada em vesículas lisossomais e os fragmentos são
externalizados no MHC classe II. E quem é que reconhece esse antígeno externalizado? É o
linfócito T auxiliar, ou helper. Então esse linfócito T helper tem um comportamento diferente do
linfócito T citotóxico, que através de citocinas, induz a apoptose da célula infectada. Esse
helper não, ele é auxiliar, ele auxiliar em um efeito maior no sistema imune. As citocinas
produzidas por ele vão atuar de diversas maneiras, vão promover a proliferação de linfóticos B,
do próprio linfótico T auxiliar, vão trazer macrófagos para aquela área, enfim, o linfócito T
auxiliar, que é o linfócito que o vírus da AIDS atua, é o grande mentor do sistema imune. Então
se o linfócito T auxiliar não está funcionando muito bem, as infecções a nível celular, aliás, não
só a nível celular, se dão com uma produção muito grande. Se, por exemplo, aquilo ali fosse
uma célula dendrítica (acho que ela aponta para a célula apresentadora de antígenos no slide)
e tivesse expondo um fragmento que foi fagocitado, o linfócito T auxiliar estaria estimulando,
via citocinas, a produção de diversos clones, os linfócitos B etc. Isso tudo mesmo no início de
uma infecção. A gente não pode pensar que isso é apenas os últimos dos últimos não.
Nesse slide aqui da pra vocês verem, perceberem que é um jogo de citocinas. O
Linfócito TH foi estimulado, etapa I, ao ser estimulado, na etapa II, ele estimula a própria célula
apresentadora de antígenos que o estimulou. A célula apresentadora de antígenos, por sua
vez, secreta um tipo de interleucina-1 que se liga a receptores que estão dentro do linfócito TH.
Essa ligação ativa a produção de interleucina-2 pelo linfócito TH. Essa interleucina-2 atua em
receptores do próprio linfócito TH, estimulando a divisão celular. Isso tudo aí eu só mostrei
para que vocês vissem como é o jogo de vai e vem de citocinas, e tudo dependendo de
receptores. Vejam que a manutenção da integridade da membrana celular é crucial, porque se
vocês tiverem falha na produção de determinados receptores, por exemplo, você pode
desencadear uma falha na resposta imune. Uma citocina liberada acaba não estimulando a
proliferação de um tipo celular ou coisa semelhante. Bom pessoal, eu não vou mais insistir não
que eu acho que vocês tão muito cansados já...
FIM