Volume 23
Série de Publicações ILSI Brasil
Funções Plenamente
Reconhecidas de Nutrientes
Vitamina E
Célia Cohen
Nutricionista Doutoranda do Departamento de Clínica Médica da Faculdade de Medicina de Ribeirão Preto – USP
Camila Siqueira Silva
Nutricionista Mestre e Doutora pela Faculdade de Medicina de Ribeirão Preto – USP
Helio Vannucchi
Professor Titular da Faculdade de Medicina de Ribeirão Preto – USP
Força-tarefa Alimentos Fortificados e Suplementos
Comitê de Nutrição
ILSI Brasil
Setembro 2014
© 2014 ILSI Brasil International Life Sciences Institute do Brasil
ILSI BRASIL
INTERNATIONAL LIFE SCIENCES INSTITUTE DO BRASIL
Rua Hungria, 664 - conj.113
01455-904 - São Paulo - SP - Brasil
Tel./Fax: 55 (11) 3035 5585 e-mail: [email protected]
© 2014 ILSI Brasil International Life Sciences Institute do Brasil
ISBN: 978-85-86126-48-2
Funções Plenamente Reconhecidas de Nutrientes - Vitamina E / ILSI Brasil (2014)
3
1. ESTRUTURA
O termo vitamina E é usado para uma família de oito moléculas de estrutura semelhante. Os quatro
tocoferóis consistem de um anel cromanol com padrões diferentes de substituição de grupos metil
nas posições 5,7 e 8 do grupo principal (α, β, δ, γ) e uma cadeia lateral fitil com 16 carbonos
saturados. Os tocoferóis possuem três centros quirais nos carbonos 2, 4’ e 8’, e os isômeros de
ocorrência natural possuem a configuração R nas 3 posições. Os tocotrienóis possuem o mesmo
padrão de substituição no anel cromanol com uma cadeia lateral isoprenoide com 16 carbonos
insaturados e ligações duplas nas posições 3’, 7’ e 11’ (Figura 1) (1).
Figura 1: Estrutura química de tocoferóis e tocotrienóis (1).
2. METABOLISMO
A vitamina E é captada na forma alcoólica livre (6-hidroxil) pelo intestino sem discriminação dos
isômeros individuais, não havendo diferença na cinética de absorção entre o alfa-tocoferol livre
e seus ésteres acetato ou succinato após a administração oral (2). O alfa-tocoferol absorvido é
secretado pelo intestino em quilomícrons e os remanescentes de quilomícron são capturados pelo
fígado, que secreta o alfa-tocoferol em VLDL (3). O metabolismo do VLDL resulta na incorporação
de alfa-tocoferol pelo LDL e HDL (4, 5).
Dos 4 tocoferóis e 4 tocotrienóis (designados como alfa-, beta-, gama- e sigma-) encontrados nos
alimentos, apenas o alfa-tocoferol supre as necessidades humanas de vitamina E (6). Apesar de todas
essas vitaminas Es apresentarem funções antioxidantes similares (taxas constantes para doação de
átomos H dentro de uma ordem de magnitude), não-alfa-tocoferóis são pouco reconhecidos pela
proteína transportadora de alfa-tocoferol (alfa-TTP). Desta forma, o alfa-TTP é responsável pela
manutenção das concentrações de alfa-tocoferol plasmáticas (7).
Na ausência de alfa-TTP, a via padrão resulta no acúmulo lisossomal de alfa-tocoferol e na sua
excreção, em vez da secreção de alfa-tocoferol no plasma, como mostrado em pacientes com
defeito na alfa-TTP (8).
4
Funções Plenamente Reconhecidas de Nutrientes - Vitamina E / ILSI Brasil (2014)
Os metabólitos da vitamina E são os CEHC (2’-carboxietil-6-hidroxicromano) produtos das
respectivas formas de vitamina E, isto é, alfa-, beta-, gama- e sigma-CEHC. As várias formas
de vitamina E são w-oxidadas pelos citocromos P450s (CYPs), seguidos da beta-oxidação e são
conjugados e excretados na urina (9) ou bile (10). De forma similar a outros xenobióticos, CEHCs
são sulfatadas (11) ou glicuronidadas (12-14). A excreção urinária de alfa-CEHC é positivamente
correlacionada com a concentração plasmática de alfa-TOH e pode refletir a quantidade de alfaTOH que excede o requerimento corporal ou capacidade de fatores envolvidos (15).
3. FUNÇÃO
A principal atividade fisiológica da vitamina E é a sua ação antioxidante, sendo o alfa-tocoferol a
isoforma com maior atividade (16). A vitamina E está presente de forma abundante nas membranas
biológicas, onde protege os ácidos graxos poli-insaturados da peroxidação, contribuindo assim
para a manutenção da integridade e estabilidade de estruturas celulares (17, 18). O a-tocoferol
reage com radicais peroxila e hidroxila, impedindo a propagação das reações em cadeia induzidas
pelos radicais livres (19).
Além de possuir propriedades de captura de radicais, o alfa-tocoferol pode agir como um potente
redutor e um agente eletrofílico em reações químicas (20). O alfa-tocoferol e seus isômeros
podem estar envolvidos na progressão da oxidação de LDL e na aterosclerose, não apenas por suas
propriedades anti ou pró-oxidantes, mas também por sua capacidade de regular a expressão gênica.
A captação dietética de vitamina E pelo intestino é inespecífica, mas seu transporte pela VLDL
é regulado por proteínas citosólicas envolvidas no tráfego intracelular de tocoferóis hidrofóbicos
(1). Todas as formas de vitamina E ativam a expressão gênica pelo receptor pregnano X (21). Esse
receptor nuclear é conhecido por regular enzimas metabolizadoras de drogas como CYP3A4 (20).
Mecanismos alternativos envolvendo efeitos anti-inflamatórios e na sinalização intracelular têm
sido o foco de investigações intensas (22).
O alfa-tocoferol atua como regulador da expressão gênica por meio de duas vias centrais de
transdução de sinais, a proteína C quinase e a fosfatidilinositol 3-quinase, que regulam inúmeros
fatores de transcrição. Dessa forma, mudanças na atividade destas quinases podem influenciar uma
série de eventos altamente relevantes às funções celulares (23-25). Apesar de a sua capacidade de
modular a transdução de sinal e expressão gênica ser descrita em vários estudos, os mecanismos
moleculares ainda devem ser mais bem elucidados. Embora nem todos os genes sejam diretamente
modulados pelo sistema tocoferol-proteína C-quinase, todos os fenômenos correlacionados tornam
ainda mais difícil a análise da expressão gênica ou eventos sob controle direto do alfa-tocoferol.
A ingestão elevada de vitamina E, seja por alimentos ou suplementos, está envolvida na modulação
do sistema imune e interações das células inflamatórias com tecidos-alvo por meio de vários
mecanismos (26-28), incluindo a inibição da oxidação de LDL-colesterol, redução da liberação de
citocinas, inibição da agregação plaquetária, alteração na proliferação das células do músculo liso
e controle do tônus vascular, bem como redução da interação do endotélio vascular com células do
sistema imune e inflamatórias (29).
Funções Plenamente Reconhecidas de Nutrientes - Vitamina E / ILSI Brasil (2014)
5
4. AVALIAÇÃO DO ESTADO NUTRICIONAL DE VITAMINA E
Segundo Sauberlich (30), os parâmetros mais utilizados para a avaliação do estado nutricional
de vitamina E são o grau de hemólise dos eritrócitos, a concentração de tocoferol no plasma ou
soro, a concentração de tocoferol em eritrócitos, linfócitos ou plaquetas e as medidas de produtos
de peroxidação (31). Assim, são considerados deficientes indivíduos que possuam concentração
sérica de alfa-tocoferol inferior a 5ug/mL e hemólise eritrocitária acima de 20% (30).
5. DEFICIÊNCIA DE VITAMINA E
A vitamina E é amplamente encontrada na alimentação, e por isso sua deficiência é rara, sendo
as principais causas representadas por anormalidades nas lipoproteínas, defeitos no gene da alfaTTP e síndromes de má-absorção por insuficiência pancreática, doença de Crohn, doença celíaca,
indivíduos desnutridos com AIDS, entre outras (32, 33). Baixos níveis de vitamina E no plasma
podem ocorrer em condições clínicas agudas, como traumas, sepse e processos inflamatórios
(33). Além disso, há maior exigência desta vitamina antioxidante em pacientes queimados (34). A
deficiência de vitamina E induz a peroxidação lipídica em diferentes condições como nefrectomia,
processos inflamatórios, alcoolismo agudo (35-37). Indivíduos com deficiência de vitamina E e
ataxia possuem uma neuropatia periférica caracterizada pela morte dos axônios de grande calibre
nos nervos sensoriais (38). Pacientes com essa síndrome possuem concentração reduzida de
vitamina E em seus nervos previamente ao aparecimento da função nervosa anormal (39).
6. RECOMENDAÇÕES
O nível recomendado de ingestão média diária para adultos de ambos os gêneros foi aumentado
pelo Food and Nutrition Board (FNB) da Academia Nacional de Ciências (6) e leva em
consideração apenas a forma alfa-tocoferol (AT) da vitamina E. No passado, a atividade de
vitamina E nos alimentos era expressa como equivalentes de alfa-tocoferol (ATEs) porque
pensava-se que as outras formas de ocorrência natural (β, γ, δ-tocoferol e tocotrienóis) também
contribuíam para tal (40). As recomendações de ingestão de vitamina são descritas na tabela 1.
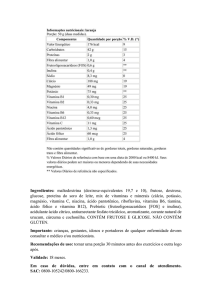
Tabela 1: Recomendação de ingestão de vitamina E (6).
Idade
AI (mg/dia)
EAR (mg/dia)
RDA (mg/dia)
UL (mg/dia)
0-6 meses
4
7-12 meses
5
1-3 anos
5
6
200
4-8 anos
6
7
300
9-13 anos
9
11
600
14-18 anos
12
15
800
19- > 70 anos
12
15
1000
Gestação
12
15
800
Lactação
16
19
800
6
Funções Plenamente Reconhecidas de Nutrientes - Vitamina E / ILSI Brasil (2014)
7. FONTES
A vitamina E é um nutriente essencial e, portanto, deve ser obtido por alimentos e suplementos
(41). Os oito isômeros são amplamente distribuídos na natureza, sendo a vitamina E detectada
em composições diversas nas plantas (4-160μg/g de peso fresco) (42). As fontes mais ricas desta
vitamina são os lipídeos do látex (8% peso/v), seguido por óleos vegetais (43).
8. CONSUMO NO BRASIL
No Brasil, a ingestão de vitamina E é inadequada em adolescentes de ambos os sexos entre 10 e 18
anos, entre adultos de 19 a 59 anos e em idosos de 60 anos ou mais, tanto em áreas urbanas quanto
rurais (44).
Funções Plenamente Reconhecidas de Nutrientes - Vitamina E / ILSI Brasil (2014)
7
9. BIBLIOGRAFIA
1. Schneider C. Chemistry and biology of vitamin E. Molecular nutrition & food research 2005
Jan;49(1):7-30.
2. Cheeseman KH, Holley AE, Kelly FJ, Wasil M, Hughes L, Burton G. Biokinetics in humans of
RRR-alpha-tocopherol: the free phenol, acetate ester, and succinate ester forms of vitamin E. Free
radical biology & medicine 1995 Nov;19(5):591-8.
3. Traber MG, Ingold KU, Burton GW, Kayden HJ. Absorption and transport of deuteriumsubstituted 2R,4’R,8’R-alpha-tocopherol in human lipoproteins. Lipids 1988 Aug;23(8):791-7.
4. Traber MG, Lane JC, Lagmay NR, Kayden HJ. Studies on the transfer of tocopherol between
lipoproteins. Lipids 1992 Sep;27(9):657-63.
5. Kayden HJ, Traber MG. Absorption, lipoprotein transport, and regulation of plasma concentrations
of vitamin E in humans. Journal of lipid research 1993 Mar;34(3):343-58.
6. Monsen ER. Dietary reference intakes for the antioxidant nutrients: vitamin C, vitamin E,
selenium, and carotenoids. Journal of the American Dietetic Association 2000 Jun;100(6):637-40.
7. Traber MG, Sokol RJ, Burton GW, Ingold KU, Papas AM, Huffaker JE et al. Impaired ability
of patients with familial isolated vitamin E deficiency to incorporate alpha-tocopherol into
lipoproteins secreted by the liver. The Journal of clinical investigation 1990 Feb;85(2):397-407.
8. Traber MG, Sokol RJ, Kohlschutter A, Yokota T, Muller DP, Dufour R et al. Impaired
discrimination between stereoisomers of alpha-tocopherol in patients with familial isolated vitamin
E deficiency. Journal of lipid research 1993 Feb;34(2):201-10.
9. Brigelius-Flohe R, Traber MG. Vitamin E: function and metabolism. Faseb J 1999
Jul;13(10):1145-55.
8
Funções Plenamente Reconhecidas de Nutrientes - Vitamina E / ILSI Brasil (2014)
10. Kiyose C, Saito H, Kaneko K, Hamamura K, Tomioka M, Ueda T et al. Alpha-tocopherol
affects the urinary and biliary excretion of 2,7,8-trimethyl-2 (2’-carboxyethyl)-6-hydroxychroman,
gamma-tocopherol metabolite, in rats. Lipids 2001 May;36(5):467-72.
11. Jiang Q, Freiser H, Wood KV, Yin X. Identification and quantitation of novel vitamin E
metabolites, sulfated long-chain carboxychromanols, in human A549 cells and in rats. Journal of
lipid research 2007 May;48(5):1221-30.
12. Swanson JE, Ben RN, Burton GW, Parker RS. Urinary excretion of 2,7, 8-trimethyl-2-(betacarboxyethyl)-6-hydroxychroman is a major route of elimination of gamma-tocopherol in humans.
Journal of lipid research 1999 Apr;40(4):665-71.
13. Stahl W, Graf P, Brigelius-Flohe R, Wechter W, Sies H. Quantification of the alpha- and
gamma-tocopherol metabolites 2,5,7, 8-tetramethyl-2-(2’-carboxyethyl)-6-hydroxychroman and
2,7, 8-trimethyl-2-(2’-carboxyethyl)-6-hydroxychroman in human serum. Analytical biochemistry
1999 Nov 15;275(2):254-9.
14. Pope SA, Burtin GE, Clayton PT, Madge DJ, Muller DP. Synthesis and analysis of conjugates
of the major vitamin E metabolite, alpha-CEHC. Free radical biology & medicine 2002 Sep
15;33(6):807-17.
15. Eichhorn JC, Lee R, Dunster C, Basu S, Kelly FJ. Alpha- and gamma-tocopherol plasma and
urinary biokinetics following alpha-tocopherol supplementation. Annals of the New York Academy
of Sciences 2004 Dec;1031:339-40.
16. Traber MG, Packer L. Vitamin E: beyond antioxidant function. The American journal of
clinical nutrition 1995 Dec;62(6 Suppl):1501S-9S.
17. Vannucchi H, Jordao Junior AA, Iglesias AC, Morandi MV, Chiarello PG. Effect of different
dietary levels of vitamin E on lipid peroxidation in rats. Archivos latinoamericanos de nutricion
1997 Mar;47(1):34-7.
18. Vannucchi H, Araujo WF, Bernardes MM, Jordao Junior AA. Effect of different vitamin E levels
on lipid peroxidation in streptozotocin-diabetic rats. International journal for vitamin and nutrition
research. Internationale Zeitschrift fur Vitamin- und Ernahrungsforschung 1999 Jul;69(4):250-4.
Funções Plenamente Reconhecidas de Nutrientes - Vitamina E / ILSI Brasil (2014)
9
19. Traber MG. Cellular and molecular mechanisms of oxidants and antioxidants. Mineral and
electrolyte metabolism 1997;23(3-6):135-9.
20. Stocker A. Molecular mechanisms of vitamin E transport. Annals of the New York Academy of
Sciences 2004 Dec;1031:44-59.
21. Landes N, Pfluger P, Kluth D, Birringer M, Ruhl R, Bol GF et al. Vitamin E activates gene
expression via the pregnane X receptor. Biochemical pharmacology 2003 Jan 15;65(2):269-73.
22. Azzi A, Stocker A. Vitamin E: non-antioxidant roles. Progress in lipid research 2000
May;39(3):231-55.
23. Azzi A, Gysin R, Kempna P, Munteanu A, Negis Y, Villacorta L et al. Vitamin E mediates cell
signaling and regulation of gene expression. Annals of the New York Academy of Sciences 2004
Dec;1031:86-95.
24. Azzi A. Molecular mechanism of alpha-tocopherol action. Free radical biology & medicine
2007 Jul 1;43(1):16-21.
25. Zingg JM. Modulation of signal transduction by vitamin E. Molecular aspects of medicine
2007 Oct-Dec;28(5-6):481-506.
26. Ozer NK, Boscoboinik D, Azzi A. New roles of low density lipoproteins and vitamin E in
the pathogenesis of atherosclerosis. Biochemistry and molecular biology international 1995
Jan;35(1):117-24.
27. Monteiro TH, Silva CS, Cordeiro Simoes Ambrosio LM, Zucoloto S, Vannucchi H. Vitamin
E alters inflammatory gene expression in alcoholic chronic pancreatitis. Journal of nutrigenetics
and nutrigenomics;5(2):94-105.
28. Silva CS, Monteiro TH, Simoes-Ambrosio LM, Sunaga DY, Cardoso JF, Furtado KS, et
al. Effects of alpha-tocopherol supplementation on liver of rats chronically exposed to ethanol.
Journal of nutrigenetics and nutrigenomics;6(3):125-36.
29. Steiner M. Influence of vitamin E on platelet function in humans. Journal of the American College
of Nutrition 1991 Oct;10(5):466-73.
30. Sauberlich H. Laboratory tests for the assessment of nutritional status. Boca Raton: CRC-Press
1999.
31. Cozzolino MBS. Vitamina E (Tocoferol). In: SMF C, ed. Biodisponibilidade de nutrientes. 3 ed.
Barueri, SP: Manole 2009:319-39.
32. Pontes Monteiro J, Ferreira da Cunha D, Freire Carvalho Cunha S, Modesto dos Santos V, Jordao
AA, Correia D et al. Nutritional assessment of vitamin E in malnourished patients with AIDS. Nutrition
(Burbank, Los Angeles County, Calif. 2000 May;16(5):339-43.
33. Asakura L. Vitamina E. In: Vannucchi H, ed. Nutrição Humana. Rio de Janeiro: Guanabara Koogan
2006.
34. Vinha PP, Martinez EZ, Vannucchi H, Marchini JS, Farina JA, Jr., Jordao AA, Jr. et al. Effect of
acute thermal injury in status of serum vitamins, inflammatory markers, and oxidative stress markers:
preliminary data. J Burn Care Res. Mar-Apr;34(2):e87-91.
35. Cordeiro MB, Coimbra TM, Costa RS, Meirelles MS, Jordao AA, Jr., Vannucchi H. Lipid peroxidation
in vitamin E-deficient rats submitted to subtotal nephrectomy. Renal failure 2002 Jul;24(4):407-19.
36. Vannucchi H, Vianna SM, Meirelles MS, Campos AD, Sarti W, Jordao Junior AA. Lipid peroxidation
and urinary excretion of vitamin E in rats submitted to an immunological inflammatory process. Drug
and chemical toxicology 2003 Nov;26(4):285-93.
37. Jordao AA, Jr., Chiarello PG, Arantes MR, Meirelles MS, Vannucchi H. Effect of an acute dose of
ethanol on lipid peroxidation in rats: action of vitamin E. Food Chem Toxicol 2004 Mar;42(3):459-64.
38. Sokol R. Vitamin E deficiency and neurological disorders. In: Packer L FJ, ed. Vitamin E in Health
and Disease. New York: Marcel Dekker 1993:815-49.
39. Traber MG, Sokol RJ, Ringel SP, Neville HE, Thellman CA, Kayden HJ. Lack of tocopherol in
peripheral nerves of vitamin E-deficient patients with peripheral neuropathy. The New England journal
of medicine 1987 Jul 30;317(5):262-5.
40. Murphy SP, Barr SI, Poos MI. Using the new dietary reference intakes to assess diets: a map to the
maze. Nutrition reviews 2002 Sep;60(9):267-75.
41. Burton GW, Cheeseman KH, Doba T, Ingold KU, Slater TF. Vitamin E as an antioxidant in vitro and
in vivo. Ciba Foundation symposium 1983;101:4-18.
42. Threlfall DR. The biosynthesis of vitamins E and K and related compounds. Vitamins and hormones
1971;29:153-200.
43. Azzi A, Breyer I, Feher M, Pastori M, Ricciarelli R, Spycher S et al. Specific cellular responses to
alpha-tocopherol. The Journal of nutrition 2000 Jul;130(7):1649-52.
44. IBGE. Pesquisa de orçamentos familiares 2008-2009: análise do consumo alimentar pessoal no
Brasil, 2011.
ILSI Brasil
International Life Sciences Institute do Brasil
Rua Hungria, 664 - conj. 113 - 01455-904 - São Paulo - SP - Brasil
Tel./Fax: 55 (11) 3035-5585 - e-mail: [email protected]