TD DE BIOLOGIA (ATUALIDADES – Nº 06) – 2017
Colégio Nossa Senhora das Graças
Congregação das Filhas do Coração Imaculado de Maria
Rua Monsenhor Otávio de Castro, 535 - Bairro de Fátima - Fortaleza-Ceará
CEP: 60.050-150 – Telefone: (0xx85)4012-0858/Fax: (0xx85)4012-0853
Home Page: www.cnsg.com.br E-mail: [email protected]
ALUNO(A): ___________________________________________________________________________________________________________ Nº: ________
SÉRIE: 3ª
CURSO: EM
TURMA: ______________
TD DE BIOLOGIA (ATUALIDADES – Nº 06) – 2017
TURNO: ______________
DATA: ___/___/___
PROFESSOR: LINCOLN TAVARES
Corpos Cetônicos
Por Débora Carvalho Meldau
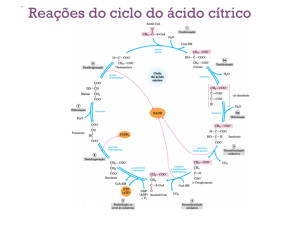
Durante o processo de oxidação dos ácidos graxos no
fígado dos seres humanos e da maioria dos outros
mamíferos, o acetil-CoA (acetilcoenzima A) formado
pode entrar no ciclo do ácido cítrico ou pode ser
convertido nos denominados “corpos cetônicos”, ou seja,
favorecida.
A produção do composto em questão pelo fígado e sua
exportação para os tecidos extra-hepáticos, em geral,
permitem a oxidação continuada dos ácidos graxos no
fígado, mesmo quando não há a oxidação do acetil-CoA
por meio do ciclo do ácido cítrico.
em acetoacetato, D- -hidroxibutirato e acetona, que
são exportados para outros tecidos através da circulação
sanguínea.
Acetona
O primeiro evento para a formação do acetoacetato a
nível hepático é a condensação enzimática de duas
moléculas de acetil-CoA, catalizada pela enzima tiolase.
Então, há a condensação do acetoacetil-CoA em acetil-
Acetilcoenzima A
A acetona, que é produzida em menor quantidade do
que os outros compostos, é exalada. O acetoacetato e o
CoA para originar o -hidroxi- -metilglutaril-CoA, o
D- -hidroxibutirato são transportados pelo sangue até
qual é quebrado para formar acetoato livre e acetil-CoA.
alcançarem os tecidos extra-hepáticos (por exemplo,
músculos esqueléticos, cardíaco, córtex renal), onde
ocorre a oxidação desses compostos por meio da via do
ciclo do ácido cítrico para fornecer grande parte da
energia requerida por esses mesmos tecidos. O cérebro,
que normalmente usa apenas a glicose como
combustível, em condições de necessidade (fome),
quando a glicose não está disponível, pode adaptar-se
para utilizar o acetoacetato ou o D- -hidroxibutirato na
obtenção de energia.
O acetoato livre é reduzido em D- -hidroxibutirato,
através de uma reação reversível catalisada pela enzima
D- -hidroxibutirato desidrogenase. O acetoato é
facilmente descarboxilado; o grupo carboxila pode ser
perdido espontaneamente ou pela ação da acetoacetato
descarboxilase.
D- -hidroxibutirato
Nos tecidos extra-hepáticos o D- -hidroxibutirato é
Acetoacetato
A disponibilidade de oxalacetato para iniciar a entrada
do acetil-CoA no ciclo do ácido cítrico é o principal fator
determinante da via metabólica que será tomada pelo
acetil-CoA na mitocôndria do fígado. Em certas
circunstâncias, como no jejum, as moléculas de
oxalacetato são retiradas do ciclo do ácido cítrico e
utilizadas na síntese de moléculas de glicose
(gliconeogênese). Quando a concentração de oxalacetato
está muito baixa, pouco acetil-CoA entra no ciclo de
Krebs e, assim, a formação de corpos cetônicos é
oxidado até acetoacetato pela D- -hidroxibutirato
desidrogenase. O acetoacetato é ativado para dar origem
ao éster da coenzima A por transferência do CoA do
succinil-CoA (intermediário do ciclo do ácido cítrico),
em uma reação catalisada pela -cetoacil-CoA
transferase. O acetil-CoA é então clivado pela enzima
tilose, liberando suas duas moléculas de acetil-CoA que,
por sua vez, entram no ciclo do ácido cítrico.
A produção e a exportação dos corpos cetônicos pelo
fígado permitem a oxidação continuada dos ácidos
RN 1443 ID/17
1
TD DE BIOLOGIA (ATUALIDADES – Nº 06) – 2017
graxos, mesmo com uma mínima oxidação do acetil-CoA
a nível hepático. Isso ocorre, por exemplo, quando os
intermediários do ácido cítrico estão empregados para a
síntese de glicose, através da gliconeogênese, a oxidação
dos intermediários do ciclo do ácido cítrico diminui e o
mesmo ocorre com a oxidação do acetil-CoA. Além disso,
o fígado possui uma quantidade limitada de coenzima A
e, quando a maior parte dela está ligada nas moléculas
do acetil-CoA, a -oxidação dos ácidos graxos é reduzida
de velocidade devido à falta desta coenzima livre. A
produção e a exportação dos corpos cetônicos liberam a
coenzima A, permitindo que a oxidação dos ácidos graxos
continue.
Casos como jejum prolongado, ou diabetes melito nãotratado, resultam em uma superprodução de corpos
cetônicos, à qual se associam sérios problemas médicos.
Durante o jejum, a gliconeogênese retira a maior parte
dos intermediários do ciclo de Krebs, redirecionando o
acetil-CoA para a produção de corpos cetônicos. No
diabetes não-tratado, a insulina está presente em ínfimas
quantidades, e os tecidos extra-hepáticos não
conseguem captar a glicose da corrente sanguínea de
forma eficiente. Para aumentar o nível de glicose no
sangue, a gliconeogênese hepática é acelerada, o que
também ocorre com a oxidação dos ácidos graxos no
fígado e na musculatura, gerando uma produção de
corpos cetônicos acima da capacidade de sua oxidação
pelos tecidos extra-hepáticos.
O aumento nos níveis sanguíneos do acetoacetato e D-
-hidroxibutirato diminuem o pH sanguíneo, resultando
em uma acidose, condição que pode provocar o coma,
em casos extremos, e até evoluir para a morte. Os corpos
cetônicos no sangue e na urina de indivíduos diabéticos
não-tratados podem atingir níveis muito altos, condição
denominada cetose.
Fontes:
http://pt.wikipedia.org/wiki/Corpos_cetônicos
Lehninger Princípios da Bioquímica – David L. Nelson,
Michel M. Cox. 3 edição, 2002.
RN 1443 ID/17
2