Deriva genética, tamanho efetivo
populacional e teoria neutra
Instituto de Biociências
USP
Diogo Meyer
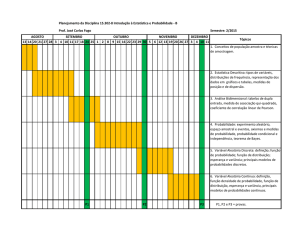
Ridley, 6.5, 6.6, 6.7, 7.1, 7.2 7.3
(não cubro quadro 7.1 e 7.2)
Deriva para o cenário 3
2
Frequências alélicas (Noturno)
Média freq. alélicas
0.6
0.5
Frequência
0.4
Média cen. I
0.3
Média cen. IV
Média cen. III
0.2
Média cen. II
0.1
0
1
2
3
4
5
6
7
8
9
10
11
12
Gerações
3
Variância de frequências
alélicas (Noturno)
Variância freq. alélicas
0.3
0.25
Variância
0.2
Variância cen. I
0.15
Variância cen. IV
Variância cen. III
0.1
Variância cen. II
0.05
0
1
2
3
4
5
6
7
8
9
10
11
12
Gerações
4
Taxa de heterozigose
Média taxa Heterozigose
0.6
Média Heterozigose
0.5
0.4
H cen. I
0.3
H cen. IV
H cen. III
0.2
H cen. II
0.1
0
1
2
3
4
5
6
7
8
9
10
11
12
Gerações
5
Taxa de heterozigose
H observada v.s. H esperada
0.6
Heterozigose (H)
0.5
0.4
H. cen. I (N=4)
0.3
H. cen. III (N=8)
Linha Teo. N=8
0.2
Linha Teo. N=4
0.1
0
1
2
3
4
5
6
7
Gerações
8
9
10
11
12
6
Conceitos sobre deriva genética
- Aumenta variância entre populações
- Diminui diversidade dentro de populações
- processos são mais rápidos em populações
pequenas
Embora a maioria dos alunos já conheça os pressupos
Probabilidade
de fixação de um
menos da disciplina de Genética Populacional e Filogeo
tenham a noção exacta de que os principais co
alelo
populações estão vinculados a modelos concret
demográfico (Ewens, 2004; p. 35-38). Por isso, come
revisão do modelo W-F baseada, entre outras, na defin
(2005 p. 13-14).
http://www.biology.arizona.edu/evolution/act/drift/frame.html
A figura 4 mostra um exemplo da evolução da iden
modelo W-F, em que se procura relacionar intuit
oscilação de frequências (deriva genética), fixação de linha
ancestral
comum.
de cada
alelo: 1/2N
•Probabilidade de fixação
Esta intuição é aproveitada para abordar a evolução da
descendência, F, com a dedução da expressão
•Probabilidade de fixação de classe de alelos com i cópias: i/2N
•Fórmula geral para aumento de identidade (por descendência(:
Ft = 1! (1!
1 t
) [1]
2N
W-F, em que se procura relacionar intuitivamente as noções de
reverso da modelo
teoria
do coalescente (Templeton, 2006; p. 140-141)
oscilação de frequências (deriva genética), fixação de linhagens e coalescência num
equivalênciaancestral
ilustracomum.
bem a relação entre abordagens com diferentes sent
fluxo do tempo.
Esta intuição é aproveitada para abordar
a evolução
da probabilidade identidade por
Probabilidade
de
identidade
descendência, F, com a dedução da expressão
Ft = 1! (1!
!#,"
!#+"
!#*"
!#)"
!#("
1 t
) [1]
2N
N=20
em que Ft é a probabilidade média de identidade por descendência na geração t contada
a partir da geração de referência, e N é o número de indivíduos da população
(Hartl e Clark 2007, p. 116-117, por exemplo). A probabilidade média de
amostrar ao acaso na geraçãoN=50
t duas linhagens que não provenham do mesmo
ancestral na geração de referência é 1- Ft.
!#'"
N=100
!#&"
1!
N=200
0.8!
!#%"
!#$"
!"
$"
(!"
Frequência
Probabilidade de identidade por descendência
$"
0.6!
0.4!
Tempo (gerações)
,,"
9
$'+"
A deriva é um processo bem
documentado:
Variação genética em humanos
A deriva é um processo bem
documentado:
Variação genética em humanos
Prugnolle et al., 2005
Variância entre frequências alélicas
Variância freq. Alélicas
0.25
Variância
0.2
0.15
Variância cen. I
Variância cen. IV
0.1
Variância cen. III
Variância cen. II
0.05
0
1
2
3
4
5
6
7
8
9
10
11
12
Gerações
12
from one
ments. T
polymorp
testing. A
the polym
data such
obtained
more dif
because,
frequent
therefore
1991). In
phisms
variation
Deriva
aumenta
variância
entre
in this tree can be compared
Major separations
to dates of humanpopulações
migrations, estimated from archaeological
data (see Cavilli-Sforza
et al. (1988)
and
with archaeological
(b) Comparison
data
figure 2). Anatomically
modern humans are assumed
to have migrated Aumento
from da
Africa
100 ka ago
roughly
variância
(Brauer 1989; Clarkentre
populações
1989;
Stringer 1990). Expansion
humanas
250
African vs.
non-African
X 200,
iZ 150
.~
t 100
I
Amerind vs.
Northeast Asian
X~
50
0._.
0
,,w'
20
,,
Southeast Asian vs.
1Australian & New Guinean
-
Caucasoidvs. Northeast
Asian & Amerind
40
60
time/ ka
80
(a) Data
Previou
morphism
al. 1991a
100
Mountain et al.,
1991
populatio
A deriva é um processo bem
documentado:
Evidências experimentais
Buri, 1956
•107 populações
(garrafas) de
drosophila
•16 indivíduos em
cada
•Alelo bw75 visível,
início p = 0.5
rozygosity
A deriva é um processo bem
documentado:
Taxa de heterozigose (H)
Evidências experimentais
Declínio de H
para N=9
Declínio de H
para N=16
Declínio de H observado
Geração
Tamanho Populacional Efetivo (Ne)
Definição: Tamanho de uma população idealizada
(Wright-Fisher) que perde variação na mesma taxa
que a população sob estudo
Porque Ne pode ser diferente de N?
- períodos de tamanho pequeno
- variância na reprodução
- assimetria na razão sexual dos sexos
Tamanho de roedores ao longo do
tempo (bank voles e )
Myodes
Apedemus
17
Tamanho Populacional Efetivo (Ne)
3.6
Testes da teoria n
1. Variação em tamanho populacional ao longo do tempo
por exemplo, supondo que há 5 gerações com mudanças de
ex. 5 gerações com tamanho que varia:
N usando:
•se oH
proporcional
a
N
⇣
⌘
e
1
Ne
=
1
5
1
N1
+
1
N2
+
1
N3
+
1
N4
+
1
N5
uma população
tem relógio molec
• Para
k igual
a u,quelogo
• 9 gerações com tamanho 1000
• seleção principalmente purifica
1
N
• 1 geração com tamanho 10
=
9 9 1
1
+
10
10 10
1000
1
9 1
=
Nee 10 1000
9 1
Ne = ( 10
1000 +
+
9 1
1
10 10 )
1 1
10 10
= 91.4
Ne = ( 10 1000 +
)
10 10
1
= 91.4
Tamanho
Populacional
Efetivo
(N
e)
Outro caso envolvem assimetr
população
2. Variação na razão sexual
Ne =
4Nm Nf
Nf +Nm
Po
19
Ne = ( 10 1000 +
)
10 10
1
= 91.4
Tamanho
Populacional
Efetivo
(N
e)
Outro caso envolvem assimetr
população
2. Variação na razão sexual
Ne =
!
!
4Nm Nf
Nf +Nm
Po
Em elefantes marinhos, nas Ilhas Falkland, encontrou-se:
550 fêmeas e 75 machos. O tamanho da população é de
625 indivíduos.
Usando marcadores genéticos Fabiani et al. (2004) viram
que só 28% dos machos reproduziram ao longo de duas
estações reprodutivas (21 machos reproduzem)
Nm = 21, Nf = 550
A taxa de perda de diversidade
genética depende do tamanho efetivo
populacional
NM = 21, NF = 550
Ne = 4NMNF / (NM + NF )
Ne = 4NMNF / (NM + NF ) ~ 81
22
O modelo
Wright-Fisher com mutação
Parâmetro do modelo
evolutivo
Pressuposto
Tamanho da população
Finito
Forma de cruzamento
Aleatório
Sobrevivência dos genótipos
Igual para todos (i.e., sem
seleção)
Introdução de novos alelos
(mutação e migração)
Mutação ocorre com taxa µ
por geração
Tipos de mutações
-
vantajosa: aumenta chances de
reprodução e sobrevivência
-
deletéria: reduz chances de reprodução
e sobrevivência
-
neutra: não altera chances de
reprodução e sobrevivência
24
Teoria neutra e de seleção
Teoria de seleção
Teoria neutra
Removidas
Deletérias
Vantajosas
Sofrem deriva
Se fixam
Neutras
25
Teoria Neutra
- Deriva pode explicar evolução?
- Teoria neutra propõe que sim:
a grande maioria da variação dentro de
espécies e
a grande maioria das diferenças entre
espécies
resultam de deriva genética
Motoo Kimiura
1924-1994
“Precisamos reconhecer a grande importância
da deriva genética aleatória, resultante do
tamanho finito de populações, na formação da
estrutura biológica de populações. A importância
da deriva tem sido desvalorizada na última
década. Essa atitude tem sido influenciada pela
opinião de que quase nenhuma mutação é
neutra, e também que o número de indivíduos
que formam uma espécie é geralmente tão
grande que a amostragem aleatória de gametas
teria efeito ínfimo no curso da evolução”. (Kimura, 1968, Nature).
Motoo Kimiura
1924-1994
Teoria neutra
Se evolução resulta só de mutação e de
deriva perguntamos:
- Quanta variação haverá numa população?
- Quantas diferenças haverá entre espécies?
Variação intrapopulacional sob
neutralidade
µ
Crow e Kimura, 1964
H pode ser estimado a
partir de dados
Podemos testar a
hipótese neutra:
- N previsto faz sentido?
1/2N
0.6
0.4
0.2
0.0
(H)
Taxa de Heterozygosity
Heterozigose
(H)
0.8
Equilíbrio entre mutação e deriva.
0
2000
4000
6000
8000
Population
size (N)
Tamanho
populacional
(N)
10000
Taxa de evolução molecular
Humano vs Camundongo
- Ancestral comum: 80 milhoẽs da anos atrás
- Proteína com 100 amino-ácidos e 16 diferenças
0,1 mudanças por proteína milhão de anos
1 mudança por amino-ácido por milhão de anos
-6
(16/100)/160 milhões = 1 x 10 mudança/aminoácido/ano
As diferenças entre espécies
surgem da variação populacional
Tempo
alelo A
alelo a
As diferenças entre espécies
surgem da variação populacional
Tempo
alelo A
alelo a
As diferenças entre espécies
surgem da variação populacional
Tempo
alelo A
alelo a
Teste da teoria neutra
• µ = 10-6 mutações por ano (1 a cada 106 anos)
• logo k= 10-6 substituições por ano (1 a cada 106 anos)
• Separação há 20 milhões de anos: 40 substituições
• Separação há 40 milhões de anos: 80 substituições
Teste da teoria neutra 1:
taxa de evolução
• Previsão da teoria neutra:
κ=μ
Teste da teoria neutra 2:
variabilidade populacional
Interação de mutação e deriva
0.6
0.4
0.2
0.0
(H)
Taxa deHeterozygosity
Heterozigose
0.8
• Previsão da teoria neutra:
0
2000
4000
6000
8000
Population size (N)
Tamanho
populacional
10000
Conceitos chave sobre teoria
neutra
• Teoria neutra: evolução por deriva genética
• Teoria neutra requer seleção negativa (ou
purificadora)
• Previsões da teoria neutra:
– Diversidade (H) proporcional ao tamanho
populacional
– divergência entre espécies proporcional ao tempo de
separação
40