Carlos Capela
GLÚCIDOS OU GLÍCIDOS
GLÚCIDOS OU GLÍCIDOS ......................................................1
A.
INTRODUÇÃO ........................................................................6
1.
FUNÇÕES.................................................................................... 8
1.a.
1.b.
1.c.
B.
Energética.................................................................................................. 8
Estrutural .................................................................................................. 8
Reserva Energética ................................................................................... 8
AS OSES OU MONOSSACÁRIDOS ..........................................9
1.
NOMENCLATURA .......................................................................... 9
2.
ISÓMEROS ÓPTICOS ..................................................................... 9
2.a.
2.b.
3.
Enantiómeros .......................................................................................... 10
Epímeros.................................................................................................. 10
ESTRUTURA E DIAGRAMAS ........................................................ 11
3.a.
3.b.
Projecções de Fisher ............................................................................... 11
Estruturas cíclicas................................................................................... 11
3.B.I.
3.B.II.
4.
4.a.
4.b.
4.c.
4.d.
4.e.
5.
5.a.
5.b.
5.c.
6.
6.a.
6.b.
6.c.
6.d.
ESTRUTURA CÍCLICA DE TOLLENS .................................................................... 12
ESTRUTURA CÍCLICA DE HAWORTH .................................................................. 13
REACÇÕES DOS MONOSSACÁRIDOS .......................................... 15
Muta-rotação ........................................................................................... 15
Reacções de Redóx .................................................................................. 16
Isomerização............................................................................................ 16
Esterificação............................................................................................ 17
Formação de Glicósidos ......................................................................... 17
MONOSSACÁRIDOS IMPORTANTES ............................................ 18
Glicose ..................................................................................................... 18
Fructose ................................................................................................... 18
Galactose ................................................................................................. 18
DERIVADOS DAS OSES ................................................................ 19
Desoxioses ............................................................................................... 19
Osaminas ................................................................................................. 19
Ácidos Aldónicos ..................................................................................... 20
Ácidos Urónicos ...................................................................................... 20
Glúcidos página 1 de 122
Carlos Capela
6.e.
6.f.
6.g.
6.h.
6.i.
6.j.
6.k.
C.
Ácidos Aldáricos...................................................................................... 21
Ácidos Siálicos......................................................................................... 21
Lactonas................................................................................................... 22
Ésteres das Oses ...................................................................................... 23
Glicósidos................................................................................................. 23
Alditóis ..................................................................................................... 24
Ciclitóis .................................................................................................... 24
ÓSIDOS ................................................................................25
1.
CLASSIFICAÇÃO.......................................................................... 25
2.
HOLÓSIDOS ................................................................................. 26
2.a.
Dissacáridos ............................................................................................ 26
2.A.I.
2.A.II.
2.A.III.
2.A.IV.
2.b.
2.c.
Oligossacáridos ....................................................................................... 27
Polissacáridos.......................................................................................... 27
2.C.I.
2.c.i.1.
2.c.i.2.
2.c.i.3.
2.c.i.4.
2.C.II.
3.
D.
SACAROSE (GLICOSE+FRUTOSE)....................................................................... 26
LACTOSE (GALACTOSE+GLICOSE) ................................................................... 26
MALTOSE (GLICOSE+GLICOSE) ........................................................................ 27
CELOBIOSE (GLICOSE+GLICOSE) ..................................................................... 27
HOMOPOLISSACÁRIDOS ..................................................................................... 27
Amido ........................................................................................................................... 27
Glicogénio.................................................................................................................... 27
Celulose........................................................................................................................ 27
Dextrinas...................................................................................................................... 27
HETEROPOLISSACÁRIDOS .................................................................................. 27
HETERÓSIDOS ............................................................................. 27
METABOLISMO DOS GLÍCIDOS ..........................................27
1.
HIDRÓLISE ENZIMÁTICA – DIGESTÃO ...................................... 27
2.
GLICÓLISE OU VIA DE EMBDEN-MEYERHOF ........................... 27
2.a.
2.b.
Introdução ............................................................................................... 27
A Glicólise propriamente dita................................................................. 27
2.B.I.
REACÇÃO Nº 1 – FOSFORILAÇÃO ....................................................................... 27
REACÇÃO Nº 2 E 3 – DA GLICOSE-6-FOSFATO À FRUTOSE-1,6-BISFOSFATO .... 27
2.B.II.
2.B.III. REACÇÃO Nº 4 – CISÃO DA FRUTOSE 1,6-BISFOSFATO EM TRIOSES-FOSFATO 27
2.B.IV. REACÇÃO Nº 5 – GLICERALDEIDO-3-FOSFATO É OXIDADO A ÁCIDO 1,3BISFOSFOGLICÉRICO .......................................................................................................... 27
2.B.V.
REACÇÃO Nº 6 – TRANSFORMAÇÃO DO ÁCIDO 1,3-BISFOSFOGLICÉRICO EM
ÁCIDO 3-FOSFOGLICÉRICO ................................................................................................ 27
2.B.VI. REACÇÃO Nº 7, 8 E 9 – FORMAÇÃO DO ÁCIDO PIRÚVICO ................................. 27
Glúcidos página 2 de 122
Carlos Capela
2.c.
Regulação da glicólise ............................................................................ 27
2.C.I.
2.c.i.1.
2.c.i.2.
2.C.II.
2.c.ii.1.
2.c.ii.2.
2.d.
2.e.
2.f.
3.
REGULAÇÃO DA ENTRADA DE GLICOSE NA VIA: .............................................. 27
Glicogénio Fosforilase:............................................................................................... 27
Hexocinase: ................................................................................................................. 27
REGULAÇÃO DA VIA PROPRIAMENTE DITA: ...................................................... 27
Fosfofrutocinase I:..................................................................................................... 27
Piruvato-cinase: ......................................................................................................... 27
Ciclo de Rapaport-Luebering ................................................................. 27
Obtenção de energia no músculo ........................................................... 27
Destino da Di-hidroxiacetona-fosfato.................................................... 27
REOXIDAÇÃO DO NADH............................................................ 27
3.a.
Em Aerobiose .......................................................................................... 27
3.A.I.
3.A.II.
3.A.III.
3.b.
Em Anaerobiose ...................................................................................... 27
3.B.I.
3.B.II.
3.B.III.
3.B.IV.
4.
TRANSPORTE PELO GLICEROL-3-FOSFATO ....................................................... 27
TRANSPORTE PELO ÁCIDO MÁLICO OU SHUTTLE DO ÀCIDO MÁLICO ............ 27
RENDIMENTO ENERGÉTICO EM AEROBIOSE ...................................................... 27
FERMENTAÇÃO LÁCTICA ................................................................................... 27
FERMENTAÇÃO ALCOÓLICA .............................................................................. 27
REDUÇÃO DA DI-HIDROXIACETONA-FOSFATO A GLICEROL ............................ 27
BALANÇO ENERGÉTICO EM ANAEROBIOSE ........................................................ 27
VIA DAS PENTOSES-FOSFATO .................................................... 27
4.a.
4.b.
4.c.
4.d.
4.e.
Introdução ............................................................................................... 27
Fase oxidante .......................................................................................... 27
Fase Não-oxidante .................................................................................. 27
Balanço energético.................................................................................. 27
Regulação:............................................................................................... 27
5. DESCARBOXILAÇÃO OXIDANTE DO ÁCIDO PIRÚVICO A
ACETIL-COA ....................................................................................... 27
5.a.
5.b.
5.c.
5.d.
6.
Introdução ............................................................................................... 27
Descarboxilação Oxidante do Ácido Pirúvico ....................................... 27
Balanço energético.................................................................................. 27
Regulação ................................................................................................ 27
CICLO DE KREBS ........................................................................ 27
6.a.
6.b.
Introdução ............................................................................................... 27
Ciclo de Krebs propriamente dito ........................................................... 27
6.B.I.
6.B.II.
FORMAÇÃO DO ÁCIDO CÍTRICO ........................................................................ 27
FORMAÇÃO DE ÁCIDO ISOCÍTRICO ................................................................... 27
6.B.III.
6.B.IV.
6.B.V.
6.B.VI.
FORMAÇÃO DE ÁCIDO α-CETOGLUTÁRICO ...................................................... 27
FORMAÇÃO DE SUCCINIL-COA.......................................................................... 27
FORMAÇÃO DE ÁCIDO SUCCÍNICO .................................................................... 27
REGENERAÇÃO DO ÁCIDO OXALOACÉTICO ...................................................... 27
6.c.
6.d.
Balanço Energético................................................................................. 27
Regulação ................................................................................................ 27
Glúcidos página 3 de 122
Carlos Capela
6.D.I.
REGULAÇÃO DA ENTRADA DE ACETIL-COA: 2 ENZIMAS: ................................ 27
6.d.i.1.
6.d.i.2.
6.D.II.
Complexo da Piruvato-Desidrogenase: ...................................................................... 27
Citrato-Sintase: ........................................................................................................... 27
REGULAÇÃO DA VIA PROPRIAMENTE DITA: 3 ENZIMAS:................................... 27
6.d.ii.1.
Isocitrato-Desidrogenase: .......................................................................................... 27
6.d.ii.2.
6.d.ii.3.
6.D.III.
6.e.
7.
α-Cetoglutarato-Desidrogenase: ........................................................................................ 27
Succinato desidrogenase: .......................................................................................... 27
REACÇÕES ANAPLERÓTICAS: ............................................................................ 27
O ciclo de Krebs como placa giratória do metabolismo ........................ 27
CADEIA TRANSPORTADORA DE ELECTRÕES ............................ 27
7.a.
Conceito:.................................................................................................. 27
7.A.I.
7.b.
7.c.
A Cadeia de Transportadores:................................................................ 27
Organização Multimolecular dos Transportadores de Electrões:........ 27
7.C.I.
7.C.II.
7.C.III.
7.C.IV.
8.
EQUAÇÃO TERMODINÂMICA DA REOXIDAÇÃO DAS COENZIMAS:..................... 27
COMPLEXO I:...................................................................................................... 27
COMPLEXO II: .................................................................................................... 27
COMPLEXO III: .................................................................................................. 27
COMPLEXO IV:................................................................................................... 27
FOSFORILAÇÃO OXIDATIVA ...................................................... 27
8.a.
8.b.
8.c.
Conceito:.................................................................................................. 27
A Energia: ............................................................................................... 27
A Enzima ATPase: .................................................................................. 27
8.C.I.
8.C.II.
8.d.
A FRACÇÃO F1:................................................................................................... 27
A FRACÇÃO FO: .................................................................................................. 27
Acoplamento Entre Cadeia Respiratória e Fosforilação Oxidativa:.... 27
8.D.I.
8.D.II.
8.D.III.
INTERNA
A HIPÓTESE QUIMIOSMÓTICA: ......................................................................... 27
SUPORTE EXPERIMENTAL DA TEORIA DE MITCHELL:...................................... 27
TRANSPORTE DE SUBSTRATOS ATRAVÉS DA MEMBRANA MITOCONDRIAL
............................................................................................................................. 27
8.d.iii.1.
8.d.iii.2.
8.d.iii.3.
8.e.
9.
Transporte de Pi:....................................................................................................... 27
Transporte de ATP/ADP: ......................................................................................... 27
Transporte de Equivalentes Redutores: ................................................................... 27
Rendimento da Respiração Celular: ...................................................... 27
METABOLISMO DO GLICOGÉNIO .............................................. 27
9.a.
9.b.
9.c.
9.d.
Introdução ............................................................................................... 27
Síntese – Glicogénese.............................................................................. 27
Degradação – Glicogenólise ................................................................... 27
Regulação do metabolismo do glicogénio.............................................. 27
9.D.I.
9.d.i.1.
9.d.i.2.
9.D.II.
9.d.ii.1.
9.d.ii.2.
9.d.ii.3.
9.d.ii.4.
REGULAÇÃO HORMONAL DO METABOLISMO DO GLICOGÉNIO NO MÚSCULO . 27
A Epinefrina ou Adrenalina ....................................................................................... 27
A insulina .................................................................................................................... 27
REGULAÇÃO DO METABOLISMO DO GLICOGÉNIO NO FÍGADO ......................... 27
O Glucagón ou Glucagina ......................................................................................... 27
Cálcio.......................................................................................................................... 27
A insulina ................................................................................................................... 27
Glicose ........................................................................................................................ 27
Glúcidos página 4 de 122
Carlos Capela
10. NEOGLICOGÉNESE ..................................................................... 27
10.a.
10.b.
Introdução............................................................................................ 27
Substratos da Neoglicogénese............................................................. 27
10.B.I. AMINOÁCIDOS GLICOGÉNICOS OU GLICOFORMADORES................................... 27
10.B.II. LÍPIDOS ............................................................................................................... 27
10.B.III. OUTROS AÇÚCARES ........................................................................................... 27
10.c.
Neoglicogénese ou Gliconeogénese .................................................... 27
10.C.I. TRANSFORMAÇÃO DO ÁCIDO PIRÚVICO EM ÁCIDO FOSFOENOLPIRÚVICO ..... 27
10.C.II. CONVERSÃO DA FRUTOSE-1,6-BISFOSFATO À FRUTOSE-6-FOSFATO E
HIDRÓLISE DA GLICOSE-6-FOSFATO ................................................................................. 27
10.d.
10.e.
10.f.
Balanço energético .............................................................................. 27
Regulação............................................................................................. 27
Ciclos dos Cori e de Felig.................................................................... 27
10.F.I.
10.F.II.
CICLO DOS CORI................................................................................................. 27
CICLO DA ALANINA OU CICLO DE FEHLIG ........................................................ 27
11. HOMEOSTASE DA GLICOSE ........................................................ 27
11.a.
Regulação Hormonal .......................................................................... 27
11.A.I. O CAMP ............................................................................................................. 27
11.A.II. OS GLICOCORTICÓIDES – O CORTISOL ............................................................. 27
11.A.III. A INSULINA ........................................................................................................ 27
11.A.IV. A GLUCAGINA E A ADRENALINA ....................................................................... 27
11.b.
11.c.
Fígado e Rim........................................................................................ 27
Outros órgãos e tecidos ....................................................................... 27
11.C.I. O MÚSCULO........................................................................................................ 27
11.C.II. O TECIDO ADIPOSO ............................................................................................ 27
11.C.III. CÉREBRO ............................................................................................................ 27
12. METABOLISMO DAS OUTRAS OSES ............................................ 27
12.a.
12.b.
12.c.
A frutose ............................................................................................... 27
A Galactose .......................................................................................... 27
O Ácido Glicurónico............................................................................ 27
12.C.I.
12.C.II.
E.
SÍNTESE ............................................................................................................... 27
CATABOLISMO .................................................................................................... 27
ANEXOS ...............................................................................27
1.
1.a.
1.b.
1.c.
2.
2.a.
2.b.
2.c.
PARA SABER MAIS … OS TRANSPORTADORES DE GLICOSE .... 27
Introdução ............................................................................................... 27
Transportadores de Glicose .................................................................... 27
A absorção de glúcidos ........................................................................... 27
PARA SABER MAIS … A DIABETES MELLITUS .......................... 27
Introdução ............................................................................................... 27
Diabetes Mellitus Insulino-Dependente................................................. 27
Diabetes Mellitus Insulino-Independente.............................................. 27
Glúcidos página 5 de 122
Carlos Capela
A. INTRODUÇÃO
Os carbohidratos (também chamados sacáridos, glúcidos, oses, hidratos de carbono ou
açúcares), são definidos, quimicamente, como poli-hidroxicetonas (cetoses) ou poli-hidroxialdeídos
(aldoses), ou seja, compostos orgânicos com, pelo menos três carbonos onde todos os carbonos
possuem um hidroxilo, com excepção de um, que possui o carbonilo primário (grupo aldeído) ou o
carbonilo secundário (grupo cetona).
Possuem fórmula empírica Cn(H2O)m, desde os mais simples (os monossacáridos, onde n =
m) até aos mais complexos. Mas alguns carbohidratos, possuem na sua estrutura nitrogénio, fósforo
ou enxofre não se adequando, portanto, à fórmula geral.
A grande informação subjacente a esta fórmula geral é a origem fotossintética dos
carbohidratos nas plantas, podendo-se dizer que os carbohidratos contêm na sua molécula a água, o
CO2 e a energia luminosa que foram utilizados na sua síntese. A conversão da energia luminosa em
energia química faz com que esses compostos fotossintetizados funcionem como um verdadeiro
combustível celular, libertando uma grande quantidade de energia térmica quando quebrada as
ligações dos carbonos das suas moléculas, libertando, também, a água e o CO2 que se encontravam
ligados.
Glúcidos página 6 de 122
Carlos Capela
A relação entre a fotossíntese e a função energética dos carbohidratos é indiscutível. De
facto, a clorofila presente nas células vegetais é a única molécula da natureza que não emite energia
na forma de calor após ter os seus electrões excitados pela luz: ela utiliza esta energia para unir
átomos de carbono do CO2 absorvido, “armazenando-a” nas moléculas de glicose sintetizadas neste
processo fotossintético.
Os animais não são capazes de sintetizar carbohidratos a partir de substratos simples não
energéticos, precisando obtê-los através da alimentação, produzindo CO2 (excretado para a
atmosfera), água e energia (utilizados nas reacções intracelulares).
Nos animais, há um processo chamado neoglicogénese que corresponde a uma síntese de
glicose a partir de percursores não glucídicos. Um outro processo de síntese endógena de glicose dáse através da glicogenólise do glicogénio sintetizado no fígado e músculos (glicogénese). Esses
processos, entretanto, só são possíveis a partir de substratos provenientes de um prévio metabolismo
glucídico, o que obriga a obtenção de carbohidratos pela alimentação, facto que torna os animais
dependentes das plantas em termos de obtenção de energia.
A energia térmica contida na molécula de glicose é libertada nas mitocôndrias e, por fim,
convertida em ligações altamente energéticas de fosfato na molécula de ATP (adenosina tri-fosfato)
durante o processo de respiração celular (fosforilação oxidativa). As duas primeiras ligações
libertam elevada energia (± 10 Kcal) quando quebradas, ao contrário da primeira que possui baixa
energia de ligação em relação às primeiras (± 6 Kcal).
Note que o ATP corresponde, enfim, a um verdadeiro armazém da energia solar que foi
conservada durante todo este fantástico processo biológico.
Glúcidos página 7 de 122
Carlos Capela
1.FUNÇÕES
1.a. Energética
São os principais produtores de energia sob a forma de ATP, cujas ligações ricas em energia
(±10 Kcal) são quebradas sempre que as células precisam de energia para as reacções bioquímicas. É
a principal função dos carbohidratos. Todos os seres vivos (com excepção dos vírus) possuem um
metabolismo adaptado ao consumo de glicose como substrato energético. Algumas bactérias
consumem dissacáridos (p.ex.: a lactose) na ausência de glicose, porém a maioria dos seres vivos
utiliza a glicose como a principal fonte energética.
1.b. Estrutural
A parede celular das plantas é constituída por um polímero de glicose – a celulose; a carapaça
dos insectos contém quitina, um polímero que fornece extrema resistência ao exo-esqueleto; as
células animais possuem uma série de carbohidratos na membrana plasmática responsáveis pelo
reconhecimento celular, pela agregação das células num tecido e por alguma actividade enzimática –
o glicocálice.
1.c. Reserva Energética
Nas plantas, há o amido, polímero de glicose; nos animais, há o glicogénio, também polímero
de glicose porém com uma estrutura mais compacta e ramificada.
Glúcidos página 8 de 122
Carlos Capela
B. AS OSES OU MONOSSACÁRIDOS
1.NOMENCLATURA
O nome genérico de um monossacárido inclui o tipo de função, um prefixo que indica o
número de átomos de carbono e a terminação -ose. Por exemplo:
•
Aldohexose é um aldeído de 6 carbonos;
•
Cetopentose é uma cetona de 5 carbonos.
2.ISÓMEROS ÓPTICOS
Isómeros de monossacáridos rodam a luz polarizada em direcções diferentes. O isómero que
faz rodar o plano de luz polarizada no sentido dos ponteiros do relógio é designado por dextrogiro
(+). Se o isómero rodar o plano de luz polarizada no sentido contrário ao dos ponteiros do relógio é
designado por levrogiro (–). Este dado só é observado experimentalmente, através de um
polarimetro.
A aldotriose gliceraldeido é usada como referência para todas as aldoses. Um monossacárido
é designado por D ou L dependendo do arranjamento dos átomos rodeando o carbono assimétrico
(neste caso C2). Muitos dos monossacáridos possuem mais do que um carbono quiral, o que
influencia a rotação da luz polarizada. Monossacáridos de cadeia longa possuem grupos adicionais
H-C-OH entre o carbono carbonil e o carbono quiral considerado para a designação D ou L, o que
pode promover designações opostas D/L e +/–. A maioria dos monossacáridos biológicos
importantes possui configuração D.
Mas vejamos como se classifica um isómero em D ou L. A configuração absoluta dos
monossacáridos é determinado pela estereoquimica do átomo de carbono quiral mais afastado do
carbono carbonil (numero 1 para os aldeídos e número mais baixo para uma cetona que geralmente é
sempre o carbono 2). Com base na posição do OH do carbono quiral de número mais alto, um
monossacárido é D se o OH se projectar para a direita, e L, se projectar-se para a esquerda.
Glúcidos página 9 de 122
Carlos Capela
D-gliceraldeido
L-gliceraldeido
Ao aumentar o número de carbonos quirais, o número de isómeros possíveis aumenta. Se n é
o número de carbonos quirais, o número possível de isómeros é 2n.
2.a. Enantiómeros
Estereoisómeros que são a imagem uma da outra num espelho plano são denominados por
enantiómeros. São exemplos o L e D-gliceraldeido ou a L e D-Ribose.
2.b. Epímeros
Estereoisómeros que diferem na configuração em torno de apenas um carbono assimétrico
são designados por epímeros. São exemplo a Glicose e Manose em C2.
Glúcidos página 10 de 122
Carlos Capela
3.ESTRUTURA E DIAGRAMAS
Monossacáridos são moléculas tridimensionais por isso vários métodos de desenho em 2
dimensões foram desenvolvidos.
3.a. Projecções de Fisher
O carbohidrato é desenhado com o esqueleto carbonado verticalmente e com os grupos –H e
–OH. As linhas verticais representam ligações para trás do plano do papel enquanto as horizontais
representam ligações acima do plano do papel.
3.b. Estruturas cíclicas
Em soluções aquosas (ou seja no organismo), aldeídos e cetonas reagem reversivelmente com
grupos hidroxilos para formar Hemiacetais. Somente 0,02% dos monossacáridos em solução aquosa
estão presentes na sua forma aberta. Esta ciclização ocorre, após hidratação, por eliminação de uma
molécula de água entre o OH (que ficou ligado ao carbono 1 das aldoses ou geralmente o carbono 2
das cetoses) e o OH ligado ao penúltimo ou antepenúltimo carbono da estrutura. Conforme a posição
do segundo OH envolvido, tratar-se-á de uma piranose ou de uma furanose, ou seja estruturas em
Glúcidos página 11 de 122
Carlos Capela
anel de 5 membros são denominadas furanose, enquanto estruturas em anel de 6 membros são
designadas por piranose.
Furanose
Piranose
Após ocorrer a ciclização, é gerado um novo carbono quiral (o carbono carbonil), designado
por carbono anomérico. Isto possibilita a existência de 2 formas isómeras designadas por
anómeros.
Na projecção de Fisher, conforme a posição do OH ligado ao carbono anomérico – do mesmo
lado ou do lado contrário ao OH que determina a classificação D ou L – teremos, respectivamente, o
anómero α ou o anómero β.
3.b.i. ESTRUTURA CÍCLICA DE TOLLENS
Na representação cíclica de Tollens, e para a série D, os anómeros α serão aqueles em que o
OH ligado ao carbono anomérico está à direita (isto é, do mesmo lado da ponte oxídica) enquanto
que os anómeros β são representados com este à esquerda.
O prefixo anomérico α ou β apenas deve ser utilizado em conjugação com o prefixo
configuracional e precede-o imediatamente. Ex: α-D-glicose.
Glúcidos página 12 de 122
Carlos Capela
3.b.ii. ESTRUTURA CÍCLICA DE HAWORTH
Na representação cíclica de Haworth, os grupos OH que figuravam à direita nas
representações de Tollens, são representados para baixo do plano e os que figuravam à esquerda são
representados para cima.
Nesta configuração, o isómero é designado por α se o grupo OH e o grupo CH2OH nos 2
átomos de carbono ligados pelo oxigénio estiver em trans um em relação ao outro e β se estiverem
em cis.
Glúcidos página 13 de 122
Carlos Capela
Existe ainda a possibilidade de se dividir as estruturas em anel em 2 grupos, conforme a sua
configuração espacial:
•
Estrutura em cadeira (mais comum pois é a mais estável)
•
Estrutura em barco
Glúcidos página 14 de 122
Carlos Capela
4.REACÇÕES DOS MONOSSACÁRIDOS
4.a. Muta-rotação
A interconversão em solução aquosa entre as formas α e β, piranose e furanose é dinâmica e
denomina-se Muta-rotação.
Exemplo: Para a molécula da glicose, em solução aquosa, temos as seguintes proporções:
•
β-D-Glicopiranose: 62%
•
α-D-Glicopiranose: 38%
•
α-D-Glicofuranose: menos de 0,5%
•
β-D-Glicofuranose: menos de 0,5%
•
Forma aberta: menos de 0,02%
Glúcidos página 15 de 122
Carlos Capela
4.b. Reacções de Redóx
Os monossacáridos podem sofrer uma variedade de reacções redóx, na presença de Cu2+, de
agentes oxidantes e de certas enzimas.
•
Ácidos Aldónicos: resultam da oxidação de um grupo aldeído.
•
Ácidos Aldónicos: resultam da oxidação do grupo terminal CH2OH.
•
Ácidos Aldáricos: resultam da combinação das duas reacções prévias.
•
Lactonas: são ésteres cíclicos que resultam da reacção do carbono carboxilo de um ácido
aldónico ou urónico com um grupo hidroxilo interno. Ex: Ácido L-ascórbico.
•
Alditóis: açucares álcoois resultantes da redução de um grupo aldeído ou cetona. Ex:
glicerol.
4.c. Isomerização
Monossacáridos
convertem-se
facilmente
nos
seus
isómeros,
quimicamente
ou
enzimaticamente. Muitas reacções de isomerização requerem o rearranjo dos átomos de hidrogénio e
das ligações duplas com a formação de intermediários enediol.
Glúcidos página 16 de 122
Carlos Capela
4.d. Esterificação
Os grupos hidroxilos dos monossacáridos podem reagir com ácidos formando ésteres. Esteres
de fosfato e de sulfato são uns dos mais comuns na natureza. Açúcares fosforilados são mais
reactivos do que os normais, o que releva especial importância nas substituições nucleofílicas, pois
os grupos hidroxilos são grupos de saída fracos.
4.e. Formação de Glicósidos
Hemiacetais reagem com álcoois para formar acetais. A ligação formada é designada por
ligação glicosídica, e o composto é denominado glicósido. A formação de acetais “fecha” a estrutura
cíclica, prevenindo a oxidação redução e a muta-rotação. Glicósidos de um ou mais monossacáridos
produzem carbohidratos complexos. A reacção de formação de glicósidos é uma reacção de
condensação, que liberta uma molécula de água.
Glúcidos página 17 de 122
Carlos Capela
5.MONOSSACÁRIDOS IMPORTANTES
5.a. Glicose
D-glicose é um dos mais comuns monossacáridos. É o
combustível primário para as células vivas. Os neurónios e os
eritrócitos usam quase exclusivamente glucose como fonte de
energia.
α- D-glucopiranose
5.b. Fructose
D-fructose é uma cetose encontrada em grandes
quantidades na fruta e no mel. Nos animais, é produzido em
grandes quantidades como componente do sémen, sendo
usado como combustível para os espermatozóides.
β- D-fructofuranose
5.c. Galactose
D-galactose
é
usada
como
percursora
de
muitas
macromoléculas (glicolípidos, proteoglicanos, fosfolípidos e
glicoproteínas) bem como da lactose (componente do leite).
Uma desordem molecular denominada galactosemia é
devido à incapacidade de metabolisar a galactose. A galactose e os
seus derivados concentram-se em certas regiões do organismo,
α- D-galactopiranose
provocando danos hepáticos, cataratas e atraso mental.
Glúcidos página 18 de 122
Carlos Capela
6.DERIVADOS DAS OSES
Modificações dos monossacáridos resultam em compostos que são de extrema importância
no metabolismo.
6.a. Desoxioses
Quando um grupo oxidrilo (OH) de um monossacárido é substituído por um átomo de
hidrogénio. Em sistemas biológicos, isto geralmente ocorre em C2. A 2-desoxi-β-D-ribose é a aldose
que intervêm na estrutura dos ácidos nucleicos (DNA).
2-desoxi-β-D-ribose
6.b. Osaminas
É um monossacárido em que um grupo OH foi substituído por um grupo amina (NH2),
geralmente acetilado. Nos sistemas biológicos, isto ocorre novamente em C2.
D-glucosamina
D-galactosamina
N-acetil-D-glucosamina
Glúcidos página 19 de 122
Carlos Capela
6.c. Ácidos Aldónicos
Resultam da oxidação do grupo aldeído do monossacárido a COOH. São designados
substituindo o sufixo “ose” por “ónico” e antepondo a palavra ácido.
6.d. Ácidos Urónicos
São formados pela oxidação do grupo terminal CH2OH das aldoses, a COOH. O respectivo
nome é formado por substituição do sufixo “ose” por “urónico”, antepondo a palavra ácido ao nome
da ose. O sílaba “ur” tem o significado de ω.
Ácido D-glucurónico
Ácido L-idurónico
Glúcidos página 20 de 122
Carlos Capela
6.e. Ácidos Aldáricos
Resultam da combinação das duas reacções prévias, ou seja, oxidação das aldoses nos 2
átomos de carbono terminais. São designadas substituindo o sufixo “ose” por “árico” e antepondo a
palavra ácido.
6.f. Ácidos Siálicos
Os ácidos siálicos ou neuramínicos são derivados (em geral acetilados) do ácido
neuramínico, formado pela condensação de uma molécula de ácido pirúvico (carbonos 1, 2, 3) com
uma molécula de D-manosamina (carbonos 4 a 9).
A acetilação do grupo amina do ácido neuramínico origina o ácido N-acetil-neuramínico. As
outras acetilações, que conduzem a diferentes ácidos siálicos, incidem em oxidrilos (em particular
em 4 e 7). Os ácidos siálicos são constituintes de diversas glicoproteínas e glicolípidos.
Glúcidos página 21 de 122
Carlos Capela
6.g. Lactonas
São ésteres cíclicos que resultam da reacção do carbono carboxilo de um ácido aldónico ou
urónico com um grupo hidroxilo interno. Ex: Ácido L-ascórbico.
O termo vitamina C deve ser usado como termo genérico para todos os compostos que
apresentam qualitativamente a actividade biológica do ácido ascórbico.
Glúcidos página 22 de 122
Carlos Capela
6.h. Ésteres das Oses
Os grupos hidroxilos dos monossacáridos podem reagir com ácidos formando ésteres. Esteres
de fosfato e de Sulfato são uns dos mais comuns na natureza. Açúcares fosforilados são mais
reactivos do que os normais, o que releva de especial importância nas substituições nucleofílicas,
pois os grupos hidroxilos são grupos de saída fracos.
6.i. Glicósidos
Hemiacetais reagem com álcoois para formar acetais. A ligação formada é designada por
ligação glicosídica, e o composto é denominado glicósido. A formação de acetais “fecha” a estrutura
cíclica, prevenindo a oxidação-redução e a muta-rotação. Glicósidos de um ou mais monossacáridos
produzem carbohidratos complexos. A reacção de formação de glicósidos é uma reacção de
condensação, que liberta uma molécula de água.
O nome forma-se mudando o “e” final do nome do monossacárido pelo sufixo “ido” e
colocando antes dessa palavra o nome do substituinte orgânico.
Glúcidos página 23 de 122
Carlos Capela
6.j. Alditóis
São açucares álcoois resultantes da redução de um grupo aldeído ou cetona. Ex: glicerol. O
nome forma-se mudando o sufixo “ose” para “itol”.
6.k. Ciclitóis
São poliálcoois cíclicos, existentes sobretudo nos tecidos vegetais. O seu principal
representante é o mioinositol, que ocorre frequentemente associado aos fosfolípidos.
Glúcidos página 24 de 122
Carlos Capela
C. ÓSIDOS
1.CLASSIFICAÇÃO
Oligossacáridos
(2-10)
Homo-polissacáridos: somente oses
Holósidos
Polissacáridos
(>10)
Heterósidos
Homo-polissacáridos: oses + derivados de oses
Por hidrólise originam, além das oses, compostos não glucídicos ou
aglicanos
Glúcidos página 25 de 122
Carlos Capela
2.HOLÓSIDOS
2.a. Dissacáridos
Os dissacáridos são glicósidos compostos por dois monossacáridos. Em alguns dissacáridos,
um dos monossacáridos mantêm o grupo carbonil livre, podendo sofrer muta-rotação e oxidaçãoredução. Estes dissacáridos são redutores e como exemplo temos a maltose e a lactose.
Outros não possuem carbonilos livres, e portanto estão encerrados na sua forma anomérica,
sendo não reductores. Como exemplo temos sacarose.
2.a.i. SACAROSE (GLICOSE+FRUTOSE)
Resulta de uma ligação glicosídica α,β(1,2) entre os dois carbonos anoméricos da glicose e
da frutose. Portanto é um açúcar não redutor. É o comum açúcar de mesa.
Sacarose
Lactose
2.a.ii. LACTOSE (GALACTOSE+GLICOSE)
Também conhecido como açúcar do leite, resulta de um ligação glicosídica β(1,4) entre a
galactose e a glicose.
Indivíduos com deficiências na enzima Lactase, possuem uma condição fisiológica
denominada por intolerância à lactose. A lactose que é ingerida não é hidrolisada e absorvida no
intestino delgado, sendo aproveitada pelas bactérias da flora intestinal do intestino grosso que a
fermentam, produzindo quantidades elevada de gás.
Glúcidos página 26 de 122
Carlos Capela
2.a.iii. MALTOSE (GLICOSE+GLICOSE)
É um intermediário na hidrólise do amido. Resulta de uma ligação glicosídica α(1,4) entre
dois resíduos de glicose. Não surge geralmente livre na natureza.
Maltose
Celobiose
2.a.iv. CELOBIOSE (GLICOSE+GLICOSE)
É um intermediário na hidrólise da celulose. Resulta de uma ligação glicosídica β(1,4) entre
dois resíduos de glicose. Tal como a Maltose, não surge geralmente livre na natureza.
2.b. Oligossacáridos
São pequenos polímeros que consistem em 2 a 10 unidades de monossacáridos. Muitos são
encontrados como grupos prostéticos de glicoproteínas e glicolípidos.
•
N-ligação – o oligossacárido encontra-se ligado ao polipéptido através de uma ligação Nglicosídica com o grupo amida da Asparagina;
•
O-ligação – o oligossacárido encontra-se ligado ao polipéptido através de uma ligação Oglicosídica com o grupo hidroxil da serina ou treonina; ou com um grupo hidroxilo do
lípido.
Glúcidos página 27 de 122
Carlos Capela
2.c. Polissacáridos
São constituídos por grande número de moléculas da mesma ose – Homopolissacáridos – ou
de oses diferentes – Heteropolissacáridos.
2.c.i. HOMOPOLISSACÁRIDOS
2.c.i.1. Amido
O Amido é formado por uma cadeia α-glicosídica que por hidrólise fornece sempre glicose,
por isso é denominado de glicosana ou glicana. É a fonte alimentar mais importante de hidratos de
carbono, sendo encontrado nos cereais, batatas, legumes e outros vegetais.
Os 2 constituintes principais são a Amilose (15-20%) de estrutura helicoidal não ramificada,
e a Amilopectina (80-85%), constituída por cadeias ramificadas formadas por 24-30 resíduos de
glicose unidos por ligações α(1,4) nas cadeias e por ligações α(1,6) nos pontos de ramificação. As
ramificações impossibilitam a formação de uma hélice.
Amilose
2.c.i.2. Glicogénio
É o homopolissacárido de armazenamento do organismo humano. Possui uma estrutura
idêntica à da amilopectina, sendo mais ramificado, tendo ramificações (ligações α(1,6)) a cada 11-18
resíduos de glicose (ligações α(1,4)).
Glúcidos página 28 de 122
Carlos Capela
Amilopectina
Ramo
Glicogénio
Ligação α(1,6) – ponto de ramificação
Ligação α(1,4)
2.c.i.3. Celulose
A celulose é um dos compostos orgânicos mais abundantes da biosfera e a principal
substância responsável pela estrutura das paredes celulares das plantas. Faz aproximadamente um
terço da biomassa de uma planta.
Não é hidrolisável pelas enzimas presentes no aparelho digestivo do humano ou de outros
mamíferos, devido à ausência de uma hidrolase que actue sobre a ligação β. É por isso importante na
formação do bolo alimentar. A celulase é uma enzima microbial, portanto os ruminantes alojam no
seu tracto digestivo, bactérias comensais que digerem a celulose.
A celulose é constituída por cadeias muito longas, formadas por resíduos de β-D-glicose,
ligadas por ligações glicosídicas β(1,4). O monómero estrutural é a celobiose.
Celobiose
Glúcidos página 29 de 122
Carlos Capela
As cadeias de celulose podem encontrar-se estreitamente associadas através de ligações de
hidrogénio ou de tipo Van der Walls, formando microfibrilhas. As fibras de celulose consistem em
aproximadamente 40 microfibrilhas. Estas estruturas complexas formam estruturas complexas,
praticamente insolúveis, que constituem a base de utilização industrial da celulose (fibras de papel,
tecidos, etc.). Mas, além das pontes de hidrogénio que se vão estabelecer entre cadeias, também
dentro de cada cadeia ocorrem estas ligações.
A ligação β confere às cadeias uma linearidade e uma resistência tênsil que as adequa então à
construção de fibras e a servirem de material de construção nas plantas.
Microfibrilhas de Celulose
2.c.i.4. Dextrinas
São glucosanas resultantes das α-amilases sobre a amilopectina e glicogénio. Contêm em
média 8 unidades de glicose, com uma ou mais ligações glicosídicas α(1,6).
Glúcidos página 30 de 122
Carlos Capela
2.c.ii. HETEROPOLISSACÁRIDOS
Glicósidos composto por múltiplos monossacáridos de pelo menos dois tipos. Podemos
também encontrar derivados dos monossacáridos.
Devido à sua importância e abundância destaco os glicosaminoglicanos (GAGs) que
consistem em cadeias de hidratos de carbono complexos caracterizados pelo seu teor em osaminas e
ácidos urónicos.
Os GAGs são classificados tendo em atenção os resíduos de açúcar, tipos de ligações,
presença e localização dos grupos sulfato. A ligação glicosídica do dissacárido base pode ser do tipo
α (Heparina, Heparina sulfato) ou do tipo β (os restantes).
O carácter ácido resulta da presença de grupos carboxílicos, sulfúricos, ou ambos. No pH
fisiológico, estão todos carregados negativamente, o que produz repulsão entre eles. Este carácter
poli-aniónico é também aproveitado para atrair e reter cargas positivas, em especial o Na+,
desempenhando assim um papel muito importante na hidratação do meio biológico, pois a água
acompanha o Na+ por osmose.
Os GAGs são geralmente encontrados como grupos prostéticos em lípidos e proteínas,
formando os glicoconjugados.
•
Ácido hialurónico – encontrado no humor vítreo do olho, no fluido sinovial das
articulações e nas matrizes dos tecidos.
•
Condroitina-6-sulfato – é um componente da cartilagem.
•
Dermatano sulfato – componente do tecido de sustentação, cuja concentração aumenta
com a idade.
•
Heparina/Heparano sulfato – anticoagulante encontrado nos mastócitos.
•
Queratano sulfato – encontrado na córnea, cartilagem e discos intervertebrais.
Glúcidos página 31 de 122
Carlos Capela
Ácido Hialurónico
Ácido D-glicurónico + GlcNAc
Ligação β(1, 3)
Dermatano sulfato
Ácido L-idurónico + GalNAc-4-sulfato
Ligação β(1, 3)
Condroitina-4 ou 6-sulfato
Ácido D-glicurónico + GalNAc-6-sulfato
Ligação β(1, 3)
Heparina/Heparano sulfato
Ácido D-glicurónico-2-sulfato (ou Ácido Lidurónico) + N-sulfo-D-glucosamina-6-sulfato
Ligação α(1, 4)
Heparanos tem menos sulfatos que as Heparinas
Queratano sulfato
Galactose + GlcNAc-6-sulfato
Ligação β(1, 4)
Glúcidos página 32 de 122
Carlos Capela
3.HETERÓSIDOS
Os heterósidos resultam de uma ose, através da sua função semi-acetálica, com um composto
que não é nem uma ose nem um derivado de ose.
A porção não glucídica é designado por aglicano, enquanto a porção glucídica é denominada
por glicano. Como exemplo, temos os proteoglicanos, que são heterósidos constituídos por resíduos
glucídicos – os glicosaminoglicanos – ligados a uma cadeia proteica.
Glúcidos página 33 de 122
Carlos Capela
D. METABOLISMO DOS GLÍCIDOS
As OSES, em particular a Glicose, devem a sua importância ao facto de a sua oxidação fornecer
aos organismos vivos grande parte da energia que lhes é necessária. Porém, outro aspecto não menos
relevante é que os átomos de carbono da glicose vão encontrar-se num grande número de compostos
– aminoácidos, ácidos gordos, esteróis, glicerol, etc.
Os glícidos presentes nos alimentos são geralmente dissacáridos, como a lactose e a sacarose, e
polissacáridos como o amido e glicogénio, que têm de ser hidrolisados antes de poderem atravessar
as membranas celulares.
1.HIDRÓLISE ENZIMÁTICA – DIGESTÃO
A digestão dos glícidos inicia-se na cavidade bucal. O amido e o glicogénio são parcialmente
hidrolisados por amilases que catalisam a ruptura das ligações glicosídicas α-1,4. Nos animais as αamilases são enzimas da saliva e do suco pancreático.
No caso das cadeias lineares – a amilose – atinge-se a hidrólise completa em unidades de
maltose e de glicose.
AMILOSE
α-amilase
MALTOSE + GLICOSE
Porém, no caso da amilopectina e do glicogénio, a hidrólise das ligações glicosídicas α-1,4,
realiza-se com dificuldade na proximidade dos pontos de ramificação, e as ligações glicosídicas α1,6 aí existentes não são atacadas pela α-amilase. Obtêm-se assim uma mistura de maltose, de
maltotriose e de α-dextrina (oligossacárido constituído por unidades de glicose unidas por ligações
α-1,4 e α-1,6).
A hidrólise, nas dextrinas residuais, das ligações glicosídicas α-1,6 entre os pontos de ligação
é efectuada pela oligo-1,6-glicosidase segregada pelas células da mucosa intestinal. A hidrólise é
completada por uma α-glicosidase, a maltase, que quebra as unidades de maltose (provenientes da
acção da α-amilase e da oligo-1,6-glicosidase) originando-se 2 moléculas de glicose.
MALTOSE
Maltase
GLICOSE + GLICOSE
Glúcidos página 34 de 122
Carlos Capela
No intestino do Homem encontra-se ainda outras enzimas que atacam os dissacáridos:
•
A β-frutosidase ou Sacarase catalisa a hidrólise da Sacarose em Glicose e Frutose.
SACAROSE
•
Sacarase
FRUTOSE + GLICOSE
A β-galactosidase ou Lactase, catalisa a hidrólise da Lactose em Galactose e Glicose.
LACTOSE
Lactase
GALACTOSE + GLICOSE
Portanto, a digestão dos glícidos alimentares conduz predominantemente à glicose, mas
também à galactose e à frutose. Porém, o metabolismo da frutose e da galactose entroca no da
glicose.
Nas células intestinais verifica-se também, o transporte activo de glicose que depois é
fosforilada a glicose-6-fosfato pela hexocinases ou fosforilases. A glicose-6-fosfato é depois
hidrolisada, obtendo-se glicose livre no sangue, sendo esta reacção catalisada pela Glicose-6fosfatase.
Como já referi, a maior parte da glicose passa, através das células do tracto intestinal, para o
sangue portal e, depois para a circulação geral, para ser usada pelos outros tecidos. O fígado é o 1º
órgão a ter a oportunidade de remover glicose do sangue portal. Quando a glicemia (concentração de
glicose no sangue) é alta, o fígado remove a glicose, para os processos de Glicogénese e Glicólise,
que consomem glicose. Quando a glicemia baixa, o fígado fornece glicose ao sangue pelos processos
produtores de glicose, a Glicogenólise e a Neoglicogénese.
O fígado é também, o 1º órgão exposto ao sangue que flúi directamente do pâncreas, e,
portanto, está exposto às concentrações mais elevadas de hormonas libertadas pelo pâncreas
endócrino – Glucagina e Insulina. Estes importantes reguladores hormonais dos níveis de glicose
sanguínea têm efeitos sobre as etapas catalisadas por enzimas no fígado.
Glúcidos página 35 de 122
Carlos Capela
2.GLICÓLISE OU VIA DE EMBDEN-MEYERHOF
2.a. Introdução
A Glicose é o principal hidrato de carbono que é absorvido no intestino e aproveitado pelas
células do corpo que dela retiram energia, sendo a única fonte de energia para algumas células
(como os eritrócitos e células do SNC). A Glicose é tão importante para estas células que vários
outros tecidos do corpo funcionam em conjunto para assegurar a utilização contínua desta substância
(como o fígado).
É uma via singular, porque pode funcionar quer na presença de oxigénio se este estiver
presente (glicólise aeróbia), ou na ausência deste (glicólise anaeróbia). Assim a glicólise permite ao
músculo-esquelético níveis bastante elevados de actividade, mesmo não dispondo de oxigénio e
permite que tecidos com capacidade glicolítica significativa sobrevivam a episódios anóxios.
A Glicólise processa-se no citosol uma vez que as enzimas participantes também se
encontram neste compartimento celular. Consiste em 9 reacções ou passos:
2.b. A Glicólise propriamente dita
2.b.i.
REACÇÃO Nº 1 – FOSFORILAÇÃO
A concentração de glucose na corrente sanguínea é mantida a níveis sensivelmente constantes
de cerca de 4-5 mM. A glucose entra nas células por difusão facilitada. Este processo não permite a
acumulação na célula de concentrações de glucose superiores às existentes no sangue, pelo que a
célula deve ter um processo para acumular glucose no seu interior. Isto é feito por modificação
química da glucose pela enzima hexocinase. No entanto, nas células parenquimatosas do fígado e
nos ilhéus de Langerhans do pâncreas, este processo é catalisado pelas glucoquinases ou hexocinase
VI.
Glúcidos página 36 de 122
Carlos Capela
Hexocinase
Mg2+
A membrana celular é impermeável à glucose-6-fosfato, que pode por isso ser acumulada na
célula. A glucose-6-fosfato será utilizada na síntese do glicogénio (uma forma de armazenamento de
glucose), para produzir outros compostos de carbono na via das pentoses fosfato, ou degradada para
produzir energia – glicólise. Nesta etapa é necessária a utilização de ATP como dador de fosfatos
que é transformado em ADP. A reacção é acompanhada por perda significativa de energia livre sob a
forma de calor, o que torna a reacção irreversível em condições fisiológicas. A hexocinase possui
uma elevada afinidade para a glicose fosforilando toda a glicose que entra na célula, mesmo quando
as suas concentrações sanguíneas são baixas. Esta enzima sofre retro-inibição alostérica pelo produto
desta reacção: Glicose 6-fosfato.
2.b.ii. REACÇÃO Nº 2 E 3 – DA GLICOSE-6-FOSFATO À FRUTOSE-1,6BISFOSFATO
Para poder ser utilizada na produção de energia, a glucose-6-fosfato é primeiro isomerizada a
frutose-6-fosfato pela fosfohexoisomerase. A frutose-6-fosfato é depois fosforilada a frutose-1,6bisfosfato, com gasto de ATP pela fosfofrutocinase. Este é o ponto de não-retorno desta via
metabólica: a partir do momento em que a glucose é transformada em frutose-1,6-bisfosfato já não
pode ser usada em nenhuma outra via.
Glúcidos página 37 de 122
Carlos Capela
Isomerase
Fosfofrutocinase
Glucose-6-P
2.b.iii. REACÇÃO Nº 4 – CISÃO DA FRUTOSE 1,6-BISFOSFATO EM
TRIOSES-FOSFATO
Seguidamente, a frutose-1,6-bisfosfato é clivada em duas moléculas de três carbonos cada,
pela aldolase:
Aldolase
Isomerase
Estas duas moléculas (dihidroxiacetona fosfato e gliceraldeído-3-fosfato) são facilmente
interconvertíveis por isomerização catalisada pela fosfotriose-isomerase. Portanto, basta uma via
Glúcidos página 38 de 122
Carlos Capela
metabólica para degradar as duas. É por esta razão que a glucose-6-P foi isomerizada a frutose-6-P: a
clivagem da glucose daria origem a duas moléculas bastante diferentes, de dois e quatro átomos de
carbono, respectivamente, que exigiriam duas vias metabólicas diferentes para a sua degradação.
A Di-hidroxiacetona-fosfato é convertida a gliceraldeido-3-fosfato, continuando a via
glicolítica a partir do último composto.
2.b.iv. REACÇÃO Nº 5 – GLICERALDEIDO-3-FOSFATO É OXIDADO A
ÁCIDO 1,3-BISFOSFOGLICÉRICO
Os aldeídos têm potenciais de oxidação-redução bastante baixos (cerca de -600 a -500 mV).
A reacção de oxidação do gliceraldeído-3-fosfato pelo NAD+ (E0=-320 mV) é portanto bastante
espontânea. Dá-se a oxidação do gliceraldeído-3-fosfato à Ácido 1,3-Bisfosfoglicérico, devido à
Gliceraldeido-3-fosfato-desidrogenase que é NAD+ dependente, transformando-se este em NADH
+ H+, por transferência dos equivalentes redutores removidos na oxidação. Por fosforólise é
adicionado um fosfato inorgânico (Pi). Ocorre aqui a única reacção de oxidação da Glicólise.
2.b.v. REACÇÃO
Nº
6
–
TRANSFORMAÇÃO
DO
ÁCIDO
1,3-
BISFOSFOGLICÉRICO EM ÁCIDO 3-FOSFOGLICÉRICO
Os ácidos fosforilados têm grupos fosfatos bastante energéticos: a saída do grupo fosfato dá
origem a espécies muito mais estabilizadas por ressonância. O grupo fosfato do carbono 1 do 1,3bisfosfoglicerato pode por isso ser transferido para o ADP, produzindo ATP, pela acção da
fosfoglicerato-cinase, formando-se Ácido 3-fosfoglicérico. Visto que se formam 2 moléculas de
triose-fosfato por molécula de glicose, nesta etapa são formadas 2 moléculas de ATP por molécula
de glicose (mas recordemos que já gastamos também 2, ou seja, o saldo energético é nulo).
Gliceraldeido-3-P-desidrogenase
Fosfoglicerato-cinase
Glúcidos página 39 de 122
Carlos Capela
2.b.vi. REACÇÃO Nº 7, 8 E 9 – FORMAÇÃO DO ÁCIDO PIRÚVICO
O Ácido 3-fosfoglicérico é convertido a Ácido 2-fosfogliceríco pela fosfoglicerato-mutase,
que depois de desidratado pela acção de uma enolase dá origem a um fosfoenol, o Ácido fosfoenolpirúvico.
Fosfoglicerato-mutase
Enolase
Devido ao seu elevado potencial de transferência de fosfato o Ácido fosfoenol-pirúvico pode
transferir um fosfato ao ADP através da enzima Piruvato-cinase. Neste estágio formam-se 2
moléculas de ATP e 2 moléculas de ácido pirúvico por glicose oxidada.
Piruvato-cinase
Glúcidos página 40 de 122
Carlos Capela
2.c. Regulação da glicólise
São 4 as enzimas reguladoras da via glicolítica; 2 regulam a entrada de glicose na via, e outras 2
regulam a via propriamente dita.
2.c.i.
REGULAÇÃO DA ENTRADA DE GLICOSE NA VIA:
2.c.i.1. Glicogénio Fosforilase:
•
Enzima que catalisa a hidrólise do glicogénio celular em glicose-1-fosfato.
•
Sofre regulação covalente e alostérica:
o Regulação Covalente: Fosfo e Defosforilação:
Fosforilase A – Activa
Fosforilada
↓
Fosforilase B – Inactiva
Desfosforilada
↓
Fosforilase B – Activa
Desfosforilada (na presença de AMP)
o Regulação Alostérica: A forma “B”, normalmente inactiva, pode ser activada pela
presença do modulador alostérico positivo AMP, cuja concentração aumenta no músculo
após a quebra do ATP.
2.c.i.2. Hexocinase:
•
Catalisa a fosforilação da glicose a glicose-6-fosfato – Primeira reacção da via glicolítica.
•
As hexocinase I, II e III, ao contrário da IV, são inibidas pelo produto da reacção glicose-6fosfato. Se a metabolização da glicose-6-fosfato é menor que a sua síntese, esta acumula-se
inibindo a hexocinase.
Glúcidos página 41 de 122
Carlos Capela
2.c.ii.
REGULAÇÃO DA VIA PROPRIAMENTE DITA:
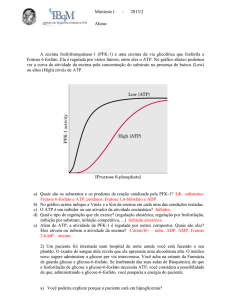
2.c.ii.1. Fosfofrutocinase I:
•
Enzima muito complexa que catalisa a fosforilação da frutose-6-fosfato – terceira etapa da
via. É unifuncional pois é incapaz de catalisar a reacção inversa que se efectua na
neoglicogénese pela acção da frutose-1,6-bisfosfatase.
•
É alostérica: possui vários activadores e inibidores, tais como os activadores AMP, ADP
(que sinalizam a falta de energia disponível) e fosfato, e os inibidores ATP, frutose-1,6bisfosfato e ácido cítrico (que sinaliza a abundância de intermediários do ciclo de Krebs). É
também inibida por H+, o que é importante em situações de anaerobiose (a fermentação
produz ácido láctico, que faz baixar o pH). Provavelmente este mecanismo impede que
nestas situações a célula esgote toda a sua reserva de ATP na reacção da fosfofrutocinase, o
que impediria a activação da glucose pela hexocinase.
•
Dentro do sistema regulador desta enzima a frutose-2,6-bisfosfato desempenha um papel
crucial pois é o efector positivo mais poderoso desta enzima. É sintetizada pela fosforilação
da Frutose-6-fosfato pela acção da fosfofrutocinase II – enzima bifuncional pois também
possui uma actividade frutose-2-6-bisfosfatase. A cinase corresponde a um domínio Nterminal e a fosfatase a um domínio carboxi-terminal. A proteína fosforilada actua como
uma bisfosfatase e desfosforilada como cinase. Os mecanismos de fosforilação dependem
duma proteína-cinase dependente de cAMP e a desfosforilação duma fosfatase. A
fosfofrutocinase II é então uma enzima bifuncional e está sob o controlo alostérico da
frutose-6-fosfato, que pelo aumento da concentração de glicose no estado bem alimentado
activa a quinase e inibe a fosfatase, acelerando a glicólise. Por outro lado, quando a glicose
estiver baixa, a glucagina estimula a produção de cAMP, activando a proteína quinase
dependente dele, que por sua vez inactiva a fosfofrutocinase II e activa a frutose-2,6bisfosfatase, através da fosforilação.
Glúcidos página 42 de 122
Carlos Capela
2.c.ii.2. Piruvato-cinase:
•
Catalisa a última reacção da via, a conversão do ácido PEP em ácido pirúvico.
•
É alostérica e inibida por ATP, Acetil-CoA e Ácidos Gordos.
Glúcidos página 43 de 122
Carlos Capela
Resumo da regulação
Glúcidos página 44 de 122
Carlos Capela
2.d. Ciclo de Rapaport-Luebering
Como no eritrócito não há mitocondrias, não pode haver nem ciclo de Krebs, nem cadeia
respiratória. Deste modo, toda a energia terá que ser fornecida pela glicólise.
Todavia, no eritrócito há uma grande concentração de ácido 2,3-bisfosfoglicérico (2,3-BPG) que
se forma por um “desvio” da glicólise, o ciclo de Rapaport-Luebering, podendo-se formar, quer pela
acção de uma mutase sobre o ácido 1,3-bisfosfoglicérico, quer pela acção de uma quinase sobre o
ácido 3-fosfoglicérico.
Mas qual é o papel do ácido 2, 3-bisfosfoglicérico? As suas cargas negativas unem-se a cargas
positivas das duas cadeias da hemoglobina, atraindo-as e aumentando, assim, a expulsão de oxigénio
para os tecidos (deslocamento da curva de dissociação da oxihemoglobina para a direita), o que é um
benefício em situações de falta de oxigénio.
Gliceraldeido-3-fosfato
Gliceraldeido-3-fosfato
desidrogenase
Ácido 1,3-
Ácido 1,3-Bisfosfoglicérico
mutase
Bisfosfoglicérico
Ácido 1,3Bisfosfoglicérico
cinase
Ácido 2,3-
Bisfosfoglicérico
Ácido-3Fosfoglicérico
Ácido 2,3-Bisfosfoglicérico
cinase
fosfatase
Ácido 3-Fosfoglicérico
Ácido Pirúvico
Glúcidos página 45 de 122
Carlos Capela
2.e. Obtenção de energia no músculo1
A forma mais simples e mais directa de obtenção de energia é a hidrólise do ATP. De notar, que
o ATP que existe normalmente no tecido muscular apenas chega para aproximadamente 1 segundo
de actividade contráctil. É por isso importante, ressintetizá-lo de imediato. Para este objectivo o
músculo contem um outro composto com uma ligação fosfato de alta energia, a fosfocreatina ou
creatina-fosfato que irá ser utilizado, nestas ocasiões. A fosfocreatina pode transferir o seu grupo
fosfato para o ADP, num processo catalisado pela creatina fosfocinase.
No entanto, se o esforço se prolongar, os músculos podem obter ATP (a partir de substratos
como a glicose e ácidos gordos):
•
Por fosforilação oxidativa, um processo energeticamente eficaz que utiliza o oxigénio
molecular, mas que é algo lento;
•
Por glicólise anaeróbia, um processo rápido, mas que esgota facilmente as reservas de
glicose.
Acabado o esforço torna-se necessário refazer as reservas. Quando o ATP não é tão necessário,
este vai decompor-se em ADP regenerando a fosfocreatina a partir da creatina.
1
Ver derivados de aminoácidos: creatina
Glúcidos página 46 de 122
Carlos Capela
2.f. Destino da Di-hidroxiacetona-fosfato
Na glicólise, vimos que na reacção catalisada pela Frutose-1,6-bisfosfato aldolase, se
formava, a partir da Frutose-1,6-bisfosfato, Gliceraldeido-3-fosfato e Di-hidroxiacetona-fosfato (2
trioses-fosfato). Esta última é em princípio totalmente convertida em Gliceraldeido-3-fosfato pela
Triose-fosfato isomerase. Assim uma molécula de glicose é convertida em 2 moléculas de
Gliceraldeido-3-fosfato.
Aldolase
Isomerase
Quando isto não acontece, a Di-hidroxiacetona-fosfato pode ser convertida em Glicerol-3fosfato (que é um percursor dos lípidos) numa reacção reversível catalisada pela Glicerol-3-fosfato
desidrogenase, em que o NADH é oxidado a NAD+.
O Glicerol-3-fosfato é, a par da Acetil-Coenzima A, o principal ponto de contacto entre os
metabolismos lipídicos e glucídicos.
Por outro lado, o glicerol pode ser fosforilado pela Glicerolcinase em Glicerol-3-fosfato, e
deste em Di-hidroxiacetona-fosfato.
Glúcidos página 47 de 122
Carlos Capela
Estas vias exprimem a relação entre o glicerol dos lípidos e o metabolismo da glicose.
Glúcidos página 48 de 122
Carlos Capela
3.REOXIDAÇÃO DO NADH
A reacção de oxidação do Gliceraldeido-3-fosfato requer NAD+, mas o teor deste é baixo nas
células, pelo que o NADH tem de ser reoxidado a NAD+.
A reoxidação pode ser realizada em condições aeróbias ou anaeróbias:
3.a. Em Aerobiose
Em aerobiose, esta reoxidação processa-se através da Cadeia Transportadora de Electrões (CTE).
Porém, enquanto a glicólise é um processo citoplasmático, o transporte electrónico é um processo
mitocondrial. E sucede que o NADH citoplasmático não consegue atravessar a membrana
mitocondrial interna.
O transporte dos electrões do NADH para a mitocôndria terá, portanto, de realizar-se através de
um transportador que os transfira do citosol até à membrana mitocondrial interna e aí os entregue a
um aceitador do CTE. Por outras palavras, tem que ser um transportador ao qual a membrana
mitocondrial interna seja permeável.
Existem vários transportadores, sendo o Glicerol-3-fosfato e o Ácido Málico os que intervêm
com mais frequência. Este sistema é conhecido como Shuttle.
3.a.i. TRANSPORTE PELO GLICEROL-3-FOSFATO
No citoplasma, o NADH vai reduzir a Di-hidroxiacetona-fosfato a Glicerol-3-fosfato, por acção
de uma desidrogenase, a Glicerol-3-fosfato desidrogenase citoplasmática. O Glicerol-3-fosfato
atravessa a membrana e, já na mitocôndria, é reoxidado a Di-hidroxiacetona-fosfato por uma
desidrogenase dependente de FAD, a Glicerol-3-fosfato desidrogenase mitocondrial.
A reacção global consiste, assim, na transferência de 2 electrões do NADH, do citosol para a
CTE mitocondrial. Porém, como o aceitador é o FAD, a reoxidação do FADH2 formado apenas
permite a síntese de 2 ATP’s.
Glúcidos página 49 de 122
Carlos Capela
3.a.ii. TRANSPORTE PELO ÁCIDO MÁLICO OU SHUTTLE DO ÀCIDO
MÁLICO
Neste caso o NADH citoplasmático transfere os seus electrões para o Ácido Oxaloacético,
reduzindo-o a Ácido Málico. Já na mitocondria este composto é reoxidado a Ácido Oxaloacético por
uma desidrogenase dependente de NAD+ do ciclo de Krebs. Trata-se de um sistema de transporte
mais eficiente, mas também mais complexo. É utilizado pelos mamíferos ao nível dos rins, fígado e
coração.
Assim quando o transportador é o Ácido Málico não há perda no rendimento energético da
Glicólise.
Glúcidos página 50 de 122
Carlos Capela
3.a.iii. RENDIMENTO ENERGÉTICO EM AEROBIOSE
Reacções
Glicose → Glicose-6-fosfato
Frutose-6-fosfato → Frutose-1,6-bisfosfato
ATP
consumido
1
ATP
formados
1
2 Ácido-1,3-bisfosfoglicérico → Ácido-3-fosfoglicérico
2
2 Ácido Fosfoenolpirúvico → 2 Ácido Pirúvico
2
4 ou 62
Reoxidação de 2 NADH pela CTE
Total
Saldo energético
8 ou 103
2
6 ou 84
2
Consoante o transporte de e- do NADH citoplasmático seja realizado através do Glicerol-3-fosfato ou do Ácido Málico.
Idem 1
4
Idem 1
3
Glúcidos página 51 de 122
Carlos Capela
3.b. Em Anaerobiose
Quando a glicólise decorre na ausência de oxigénio, e portanto a CTE não funciona (pois o
ultimo aceitador de electrões é o oxigénio molecular), a célula tem de utilizar outras reacções para
reoxidar o NADH. Veremos 3 vias, das quais as duas primeiras são as mais utilizadas.
3.b.i. FERMENTAÇÃO5 LÁCTICA
Nas células musculares (quando o oxigénio é utilizado mais rapidamente do que é fornecido
às células ocorrem aí, muitas vezes, condições de anaerobiose) ou nas bactérias lácticas (que vivem
em anaerobiose), o Ácido Pirúvico é reduzido a Ácido Láctico, enquanto o NADH é oxidado a
NAD+. Esta reacção reversível é catalisada por uma Lactato-desidrogenase.
3.b.ii. FERMENTAÇÃO ALCOÓLICA
Nomeadamente em leveduras, o Ácido Pirúvico é numa primeira etapa, descarboxilado a CO2
e Acetaldeído por uma Piruvato-descarboxilase que possui como coenzima o Pirofosfato de Tiamina
(vit. B1) e que contêm Zn2+.
Em seguida, numa reacção catalisada por uma álcool-desidrogenase, o Acetaldeído é
reduzido a etanol, enquanto o NADH é oxidado a NAD+.
5
Fermentação é um processo em que o aceitador final dos electrões provenientes da degradação é um produto orgânico
da própria degradação.
Glúcidos página 52 de 122
Carlos Capela
A reacção global da glicólise anaeróbia com fermentação alcoólica será:
Glicose + 2ADP + 2Pi + 2H+ → 2etanol + 2ATP + 2CO2 + 2H2O
Tanto a álcool-desidrogenase como a Lactato-desidrogenase têm o NADH como coenzima e
são enzimas alostéricas (tetrâmeros).
A Piruvato-descarboxilase da fermentação alcoólica não existe nos tecidos dos vertebrados
ou nos organismos que realizam a fermentação láctica.
No fígado humano, a álcool-desidrogenase catalisa a oxidação do etanol (ingerido ou
produzido pelos microorganismos intestinais), com a redução do NAD+ a NADH.
3.b.iii. REDUÇÃO DA DI-HIDROXIACETONA-FOSFATO A GLICEROL
A fermentação da glicose pelas leveduras é sempre acompanhada pela formação de pequenas
quantidades de glicerol. Uma Glicerol-3-fosfato-desidrogenase catalisa a redução da Dihidroxiacetona-fosfato a Glicerol-3-fosfato, enquanto o NADH é oxidado a NAD+. Em seguida, uma
fosfatase específica pode cindir a ligação éster e origina o glicerol.
3.b.iv. BALANÇO ENERGÉTICO EM ANAEROBIOSE
Visto que não há intervenção da CTE forma-se apenas 4 ATP (tendo-se gasto 2) por
molécula de glicose. O Saldo é de 2 ATP.
Glúcidos página 53 de 122
Carlos Capela
4.VIA DAS PENTOSES-FOSFATO
Esta série de reacções é também conhecida “Shunt” Hexose-monofosfórico, ou Via do
Fosfogluconato, ou Via de Dickens-Horecker.
4.a. Introdução
Importância Biológica:
•
Para realizar o seu anabolismo, a célula não precisa apenas de energia (ATP): também
precisa de poder redutor, sob a forma de NADPH. O NADPH é produzido durante a
oxidação da glucose-6-P por uma via distinta da glicólise, a via das pentoses-fosfato.
Esta via é muito activa em tecidos envolvidos na biossíntese de colesterol e de ácidos
gordos (fígado, tecido adiposo, córtex adrenal, glândulas mamárias).
•
Esta via também produz ribose-5-P, o açúcar constituinte dos ácidos nucleicos.
•
Permite também às células, se for caso disso metabolisar a glicose-6-fosfato com
produção de ATP sem utilizar a via da glicólise.
Ao contrário do que sucede na Glicólise, não consome ATP, e é um processo essencialmente
aeróbio, pois a reoxidação das coenzimas reduzidas só é possível através da CTE ou de reacções de
biossíntese, que utilizem o NADPH e gerem, portanto, NADP+.
As enzimas envolvidas nesta via estão localizadas no citosol. Esta via divide-se em 2 etapas:
1. A glicose-6-fosfato é descarboxilada a Ribulose-5-fosfato, precedida por 2 reacções de oxidação,
com a formação de NADPH – Fase Oxidante.
2. Interconversão das pentoses-fosfato e das hexoses-fosfato por transaldolização e transcetolisação
– Fase Não-oxidante.
Glúcidos página 54 de 122
Carlos Capela
4.b. Fase oxidante
A glucose-6-P é primeiro oxidada no seu carbono 1, dando origem a uma lactona (um ácido
carboxílico cíclico). Os electrões libertados são utilizados para reduzir uma molécula de NADP+. O
anel é então aberto por reacção com água:
Glicose-6-fosfato
desidrogenase
6-fosfo-glucolactonase
A descarboxilação do gluconato liberta dois electrões, que vão reduzir outra molécula de
NADP+. Obtém-se assim um açúcar de 5 carbonos, a ribulose-5-fosfato, que por isomerização é
transformado em ribose-5-P6.
Fosfopentose
isomerase
6-fosfogluconato
desidrogenase
6
Fosfopentose
epimerase
Na figura assinalam-se a verde as diferenças entre os isómeros
Glúcidos página 55 de 122
Carlos Capela
4.c. Fase Não-oxidante
Nesta fase ocorrem transferências de grupos com 3 átomos de carbono – Transaldolisação –
e com 2 átomos de carbono – Transcetolisação. A enzima responsável pela transaldolisação é a
Transaldolase enquanto que pela transcetolisação é a Transcetolase7.
Esta etapa depende das necessidades da célula: se a célula só precisar de NADPH e não
precisar de ribose-5-P, esta poderá ser reaproveitada. Isto é feito através de 3 reacções. Na primeira,
a ribose-5-P recebe dois carbonos da xilulose-5-P (obtida por epimerização da ribulose-5-P):
Seguidamente, são transferidos três carbonos da sedoeptulose-7-P para o gliceraldeído-3-P:
7
A Transcetolase é controlada pela vitamina B1 na sua forma activa, TPP (Tiamina de Pirofosfato). A TPP é essencial
para a indução da síntese de transcetolase. Este facto é tão importante que se pode ter uma ideia do grau de carência de
vit. B1, através do doseamento da transcetolase dos eritrócitos.
Glúcidos página 56 de 122
Carlos Capela
Por transferência de dois carbonos da xilulose-5-P para a eritrose-4-P, forma-se outra
molécula de frutose-6-P e uma molécula de gliceraldeído-3-P:
O balanço das reacções da fase não oxidante é:
2 Xilulose-5-P + Ribose-5-P → 2 frutose-6-P + gliceraldeído-3-P
A frutose-6-P e o gliceraldeído-3-P podem ser utilizados na glicólise para produção de
energia, ou reciclados pela Neoglucogénese para formar novamente glicose-6-P.
Quando as necessidades de ribose-5-P são superiores às de NADPH, esta pode ser produzida
por estas reacções a partir de frutose-6-P e gliceraldeído-3-P.
4.d. Balanço energético
Ocorrem 2 oxidações, com a formação de NADPH, cujos electrões poderão ser transferidos
para a CTE com a formação de ATP. Já se esclareceu que o destino habitual do NADPH produzido
pela via das pentoses-fosfato não é a produção de ATP, mas sim contribuir com o seu poder redutor
nas biossínteses.
Numa volta de ciclo o equivalente a uma molécula de glicose em cada seis é completamente
oxidada. Através de seis ciclos da via das pentoses-fosfato e da gluconeogénese, por cada molécula
de glucose-6-P completamente oxidada a seis moléculas de CO2 são reduzidas 12 moléculas de
NADP+ com a formação de 12 NADPH.
6 Glicose-6-P + 12 NADP+ → 5 Glicose-6-P + 6 CO2 + 12 NADPH + 12 H+ + Pi
Glúcidos página 57 de 122
Carlos Capela
4.e. Regulação:
O factor de regulação mais importante da via das pentoses é ao nível do NADP+, que é o
aceitador de electrões na oxidação da glicose-6-fosfato a ácido-6-fosfoglucónico.
Devido ao efeito do teor em NADP+, no citosol, sobre a velocidade da fase oxidante da via,
fica assegurada uma relação muito estreita entre a produção de NADPH e a sua utilização nas
reduções metabólicas e, desta forma, regulando o valor do quociente NADP+/NADPH.
Assim, se o organismo requer maior quantidade de ribose-5-fosfato do que de NADPH – isto
é, se a biossíntese das proteínas predomina sobre a dos lípidos, apenas funcionará a fase não
oxidante da via. Nestas condições, a frutose-6-fosfato e o gliceraldeído-3-fosfato (formados pela via
da glicólise a partir da glucose-6-fosfato) são transformados em ribose-5-fosfato sem formação de
NADPH.
No caso contrário, e em alternativa à fase inversa da via das interconversões, a ribose-5fosfato formada na fase oxidante, pode ser convertida em frutose-6-fosfato e em gliceraldeído-3fosfato, e daí em ácido pirúvico. Por este processo gera-se ATP e NADPH, e cinco dos seis átomos
de carbono da glucose-6-fosfato vão formar ácido pirúvico.
Glúcidos página 58 de 122
Carlos Capela
As inter-relações das vias da glicólise e das pentoses-fosfato permitem ajustar às
necessidades celulares os teores de NADPH, de ATP e de compostos centrais como a ribose-5fosfato e o ácido pirúvico.
O peróxido de hidrogénio é removido pela glutatião peroxidase. O glutatião oxidado é
reduzido pela glutatião redutase, na presença de NADPH, cuja concentração diminui, activando a
via. A via das pentoses fosfato é muito importante no eritrócito, para manter o glutatião reduzido,
para que este remova o peróxido de hidrogénio, lesivo para o eritrócito, em especial para a
membrana celular.
Glúcidos página 59 de 122
Carlos Capela
5.DESCARBOXILAÇÃO OXIDANTE DO ÁCIDO
PIRÚVICO A ACETIL-COA
5.a. Introdução
O ácido pirúvico formado no final da glicólise pode ter vários destinos metabólicos:
5.b. Descarboxilação Oxidante do Ácido Pirúvico
Mas aquele que nos importa salientar é a sua descarboxilação oxidante (que é o elo de ligação
entre a glicólise e o ciclo de Krebs).
Antes que o ácido pirúvico possa entrar no ciclo de Ácido cítrico, ele deve ser transportado
para dentro da mitocôndria através de um transportador especial de ácido pirúvico que ajuda a sua
passagem através da membrana mitocondrial interna, o que envolve um mecanismo de simporte no
qual um protão é co-transportado.
Já dentro da mitocôndria, o ácido pirúvico sofre descarboxilação oxidante, formando-se
acetil-CoA. Esta reacção é catalisada por várias enzimas diferentes que operam sequencialmente
num complexo multienzimático denominado por Complexo Piruvato-desidrogenase. A coenzima da
Glúcidos página 60 de 122
Carlos Capela
reacção é o Pirofosfato de Tiamina (TPP), ligado à enzima por interacções não covalentes. O Mg2+ é
o cofactor da reacção.
O ácido pirúvico é descarboxilado a um derivado hidroxietil do anel tiazólico do difosfato de
tiamina ligado à enzima, o qual por sua vez reage com o Ácido Lipóico, formando Acetil-lipoato. Na
presença da Dihidrolipoil-transacetilase (uma enzima do complexo), o Acetil-lipoato reage com a
Coenzima-A, formando-se Acetil-CoA e Ácido Lipóico reduzido. O ciclo da reacção termina quando
o Ácido Lipóico é reoxidado por uma flavoproteina na presença da Ácido Lipóico-desidrogenase.
Glúcidos página 61 de 122
Carlos Capela
5.c. Balanço energético
A flavoproteina reduzida é oxidada pelo NAD+, o qual por sua vez, transfere os equivalentes
redutores para a cadeia respiratória, correspondendo-lhe a formação de 3 ATP. Deve-se ter em
atenção que por cada molécula de glicose são originadas duas de ácido pirúvico e assim também 2
moléculas de Acetil-CoA, formando-se 6 ATP.
5.d. Regulação
A Piruvato-desidrogenase é inibida pelos seus produtos (retro-inibição): Acetil-CoA e
NADH. O complexo é então regulado pelo jogo quinase/fosfatase. A quinase vai ser activada pelo
aumento das razões (Acetil-CoA/CoA), (NADH/NAD+) e (ATP/ADP), fosforilando o complexo,
inactivando-o. A Piruvato-desidrogenase não é portanto apenas inibida por um alto potencial
energético, mas também nas condições de oxidação dos ácidos gordos que conduzem ao aumento
destas razões.
Por outro lado a diminuição destas razões, inibe a cinase, estimula a fosfatase que
desfosforila o Complexo Piruvato-desidrogenase, activando-o.
Glúcidos página 62 de 122
Carlos Capela
O Acetil-CoA é o composto de partida para a síntese de todos os lípidos.
Glúcidos página 63 de 122
Carlos Capela
6.CICLO DE KREBS
6.a. Introdução
Também é designado por Ciclo dos Ácidos Tricarboxílicos ou Ciclo do Ácido Cítrico.
O ciclo de Krebs ocorre na mitocôndria e compreende essencialmente, a combinação de uma
molécula de Acetil-CoA com o ácido dicarboxílico de 4 carbonos, o ácido oxaloacético, resultando a
formação de um ácido tricarboxílico de 6 carbonos, o ácido cítrico. Segue-se um conjunto de
reacções através das quais 2 moléculas de dióxido de carbono se perdem, e é regenerado o ácido
oxaloacético.
Visto que não são precisas mais do que pequenas quantidades de ácido oxaloacético para
transformar grande número de unidades acetílicas a CO2, considera-se que o ácido oxaloacético
desempenha o papel catalítico.
Este processo necessita de O2 como receptor final dos equivalentes redutores que são produzidos
sob a forma de H+ e e-. Assim, a ausência (anóxia) ou a deficiência parcial (hipóxia) de O2 determina
a inibição total ou parcial do ciclo.
As enzimas do ciclo do Ácido cítrico estão localizadas na matriz mitocondrial, livres ou ligadas à
superfície interna da membrana mitocondrial.
Glúcidos página 64 de 122
Carlos Capela
6.b. Ciclo de Krebs propriamente dito
6.b.i. FORMAÇÃO DO ÁCIDO CÍTRICO
A 1ª reacção do ciclo consiste na condensação do Acetil-CoA com o Ácido Oxaloacético para
formar o Ácido Cítrico. Esta reacção é catalisada pela Citrato sintetase, que efectua uma ligação
carbono-carbono entre o carbono-metílico do Acetil-CoA e o carbono carbonil do ácido
oxaloacético. A reacção de condensação a Citroil-SCoA, é acompanhada pela hidrólise da ligação
tioéster da CoA, perdendo-se grande parte da energia livre como calor, o que garante que a reacção
ocorra até ao fim.
Ácido Oxaloacético
Citroil-SCoA
Ligado à enzima
Ácido Cítrico
Acetil-CoA
Citrato Sintetase
6.b.ii. FORMAÇÃO DE ÁCIDO ISOCÍTRICO
O ácido cítrico é isomerizado a Ácido Isocítrico pela acção da enzima Aconitase (aconitohidratase) que contém ferro no estado Fe2+, na forma de uma proteína ferro-enxofre (Fe:S). Esta
conversão ocorre em 2 etapas: desidratação a Ácido Cis-aconítico, e a rehidratação a Ácido
Isocítrico.
A reacção é inibida pelo Ácido flúor-acético, o qual, na forma de fluoracetil-CoA condensa-se
com o ácido oxaloacético para formar o ácido fluorcítrico. Este último inibe a aconitase,
ocasionando a acumulação de ácido cítrico.
É possível que o ácido cis-aconítico não seja um intermediário obrigatório entre o ácido cítrico e
o ácido isocítrico, sendo na realidade uma ramificação lateral da via principal do ciclo.
A isomerização produz um composto de carbono ramificado mais fácil de metabolizar.
Glúcidos página 65 de 122
Carlos Capela
Ácido Cítrico
cis-Aconitato
Ácido Isocítrico
Aconitase
6.b.iii. FORMAÇÃO DE ÁCIDO α-CETOGLUTÁRICO
O ácido isocítrico sofre desidrogenação em presença da Isocitrato-desidrogenase para formar
ácido Oxalosuccínico.
Foram descritas 3 enzimas. Uma, que é NAD+-dependente, é encontrada somente nas
mitocôndrias. As outras 2 enzimas são NADP+-dependentes e são encontradas nas mitocondrias e no
citosol. A oxidação, ligada à cadeia respiratória do ácido isocítrico ocorre quase exclusivamente
através da enzima NAD+-dependente.
Segue-se uma descarboxilação a Ácido α-cetoglutárico, também catalisada pela Isocitratodesidrogenase. É possível que o ácido Oxalosuccínico permaneça ligado à enzima como
intermediário na reacção global.
Ácido Isocítrico
Ácido Oxalosuccínico
Ácido α-cetoglutárico
Isocitrato Desidrogenase
6.b.iv. FORMAÇÃO DE SUCCINIL-COA
Tal como o ácido pirúvico, o ácido α-cetoglutárico é um α-cetoácido (isto é, possui um grupo
carbonilo adjacente ao grupo ácido carboxílico). É portanto de prever que reaja exactamente como o
ácido pirúvico, isto é, que a sua descarboxilação forneça energia suficiente para que se forme uma
ligação tioéster com a coenzima A. E é isto que de facto ocorre. A enzima responsável por esta
Glúcidos página 66 de 122
Carlos Capela
reacção, a α-cetoglutarato desidrogenase, é aliás bastante análoga à piruvato desidrogenase na sua
composição e cofactores: Pirofosfato de Tiamina (TPP), ácido lipóico, NAD+, FAD e CoA.
A reacção resulta na formação de Succinil-CoA (um tioéster com uma ligação de alta energia). O
equilíbrio desta reacção é de tal modo a favor da formação de Succinil-CoA que, fisiologicamente,
ela deve ser considerada unidireccional. O Arsenito inibe a reacção, causando a acumulação do
substrato, o ácido α-cetoglutárico.
α-cetoglutarato
desidrogenase
Succinil-CoA
Ácido α-cetoglutárico
6.b.v. FORMAÇÃO DE ÁCIDO SUCCÍNICO
Para continuar o ciclo, o Succinil-CoA é convertido a Ácido Succínico pela Succinil-CoA
Sintetase. Esta reacção requer GDP, que é convertido em GTP, em presença de fosfato inorgânico.
Este é o único exemplo, no ciclo do ácido cítrico, da formação de um fosfato de alta energia ao nível
do substrato, e surge em virtude da libertação oxidativa do ácido α-cetoglutárico. O ATP pode ser
formado a partir do GTP por meio de uma nucleósido-difosfato-cinase: ADP + GTP → ATP + GDP.
Succinil-CoA
Sintetase
Succinil-CoA
Ácido Succínico
Glúcidos página 67 de 122
Carlos Capela
6.b.vi. REGENERAÇÃO DO ÁCIDO OXALOACÉTICO
O ácido succínico sofre metabolização adicional passando, primeiro por uma desidrogenação,
seguida pela adição de H2O, e subsequentemente, por uma nova desidrogenação, que regenera o
ácido oxaloacético.
A 1ª reacção de desidrogenação é catalisada pela Succinato-desidrogenase, que está ligada à
superfície interna da membrana mitocondrial interna, ao contrário das outras. Esta é a única
desidrogenação do ciclo de Krebs que envolve a transferência directa de hidrogénio do substrato sem
a participação do NAD+, uma vez que o aceitador é o FAD. Forma-se Ácido Fumárico como
resultado da desidrogenação. A adição de Ácido Málico ou Oxaloacético inibe competitivamente a
Succinato-desidrogenase provocando a acumulação de ácido succínico.
Sobre a influência da Fumarase, a água é adicionada ao ácido fumárico para originar-se Ácido
Málico.
O ácido málico é convertido a ácido oxaloacético pela Malato-desidrogenase, reacção que requer
NAD+. Embora o equilíbrio desta reacção seja muito favorável ao ácido málico, o fluxo efectivo é na
direcção do ácido oxaloacético porque esse composto, juntamente com outro produto da reacção, o
NADH + H+, é removido nas reacções subsequentes.
Succinato
Desidrogenase
Malato
Desidrogenase
Fumarase
Ácido Succínico
Ácido Fumárico
Ácido Málico
Ácido Oxaloacético
As enzimas que participam no ciclo do ácido cítrico podem ser também encontradas fora da
mitocôndria, excepto a α-cetoglutarato desidrogenase e a Succinato-desidrogenase.
Glúcidos página 68 de 122
Carlos Capela
Visão geral
Glúcidos página 69 de 122
Carlos Capela
6.c. Balanço Energético
Os equivalentes redutores formados como resultado da acção das desidrogenases são transferidos
para a cadeia respiratória na membrana interna da mitocôndria.
Os equivalentes redutores formados são 3 moléculas de NADH + H+ e uma de FADH2 por cada
molécula de Acetil-CoA catabolisada numa volta completa do ciclo.
Durante a passagem, pela cadeia transportadora de electrões, os equivalentes de redução do
NADH + H+ origina 3 ATPs. Contudo, o FADH2 produz somente 2 ATPs, visto que transfere o seu
equivalente redutor directamente à coenzima Q, o que significa que entra na cadeia respiratória
depois do NADH, perdendo a 1ª bomba de protões8.
Um outro fosfato de alta energia é formado no próprio ciclo, durante a conversão do SuccinilCoA a ácido succínico.
Assim 12 ATPs são formados por cada ciclo de ácido cítrico, e como cada molécula de glicose
origina 2 ciclo, consideram-se 24 ATPs.
Equivalentes redutores
8
ATPs formados
3 NADH + H+
3 ATPs
1 FADH2
2 ATPs
Reacção: Succinil-CoA a ácido succínico
1 ATP
Total:
12 ATPs
1 Molécula de glicose → 2 Acetil-CoA
2 × 12 = 24 ATPs
ver fosforilação oxidativa
Glúcidos página 70 de 122
Carlos Capela
6.d. Regulação
O ciclo de Krebs é controlado fundamentalmente pela disponibilidade de substratos, inibição
pelos produtos e por outros intermediários do ciclo.
A actividade é imediatamente dependente do fornecimento dos co-factores oxidados das
desidrogenases, os quais, por sua vez, devido à forte ligação entre a oxidação e a fosforilação na
CTE, são dependentes da disponibilização de ADP e, portanto, da velocidade de utilização de ATP.
Portanto, se houver suficiente fornecimento de O2, a velocidade de realização do trabalho através da
utilização de ATP determina tanto a velocidade da reacção quanto a actividade do ciclo do ácido
cítrico.
Algumas enzimas, pelas suas propriedades, indicam também que o controlo pode ser efectuado
ao nível do próprio ciclo. Estas são responsáveis pelo estado de energia expresso pelas relações
ATP/ADP e NADH/NAD+:
São 5 as enzimas que regulam a velocidade do Ciclo de Krebs, actuando na regulação do
fornecimento de combustível para a via – Acetil-CoA – e no ciclo propriamente dito.
6.d.i. REGULAÇÃO DA ENTRADA DE ACETIL-COA: 2 ENZIMAS:
6.d.i.1. Complexo da Piruvato-Desidrogenase:
•
Sofre regulação alostérica e covalente.
•
A regulação covalente: fosfo e defosforilação:
Piruvato Desidrogenase Activa
(Defosforilada)
↓
Piruvato Desidrogenase Inactiva
(Fosforilada)
•
A regulação alostérica: Inibida por ATP, Acetil-CoA e NADH + H+ → “feed back” negativo.
6.d.i.2. Citrato-Sintase:
•
Catalisa a 1ª etapa do ciclo;
•
Sofre regulação alostérica: é inibida pelo ácido cítrico, succinil-CoA e NADH + H+;
•
A concentração de ácido oxaloacético também é um factor importante de regulação da
actividade desta enzima.
Glúcidos página 71 de 122
Carlos Capela
6.d.ii. REGULAÇÃO DA VIA PROPRIAMENTE DITA: 3 ENZIMAS:
6.d.ii.1. Isocitrato-Desidrogenase:
•
Cataliza a 3ª etapa do ciclo;
•
Sofre regulação alostérica: é activada por ADP e inibida por NADH + H+ e NADPH.
6.d.ii.2. α-Cetoglutarato-Desidrogenase:
•
Cataliza a etapa 4 do ciclo;
•
Também é alostérica e inibida por succinil-CoA.
Estas desidrogenases mencionadas são estimuladas pelo ião cálcio.
6.d.ii.3. Succinato desidrogenase:
•
É inibida pelo ácido oxaloacético.
6.d.iii. REACÇÕES ANAPLERÓTICAS:
Ou “reacções que completam”. São reacções que completam as concentrações de intermediários
do Ciclo de Krebs, quando a concentração de um deles diminui na célula, garantindo assim a
continuidade da via.
Exemplo: carboxilação do ácido pirúvico a ácido oxaloacético:
Ácido Pirúvico + CO2 + ATP + H2O → ácido Oxaloacético + ADP + Pi + H+
Enzima: Piruvato-Carboxilase
Glúcidos página 72 de 122
Carlos Capela
6.e. O ciclo de Krebs como placa giratória do metabolismo
Importantes vias metabólicas tem como produto final um constituinte do ciclo, enquanto outros
se originam no ciclo. Estas vias compreendem processos como a Neoglicogénese, Transaminação,
Desaminação e Síntese de ácidos gordos. Portanto, o ciclo do ácido cítrico desempenha funções
tanto nos processos catabólicos quanto nos anabólicos, sendo por isso designado Anfibólico.
Todos os metabólitos intermediários do ciclo desde o ácido cítrico até ao ácido oxaloacético são
potencialmente glicogénicos, visto que podem originar uma produção efectiva de glicose no fígado
ou no rim, uma vez que estes órgãos são os únicos que possuem as enzimas necessárias à
neoglicogénese9. A enzima chave que permite a transferência efectiva para fora do ciclo, de um dos
seus componentes, para a via da neoglicogénese, é a fosfoenolpiruvato-carboxicinase, que catalisa a
descarboxilação do ácido oxaloacético a ácido fosfoenolpirúvico, funcionando o GTP como fonte de
energia.
As reacções de transaminação produzem ácido pirúvico a partir da alanina, ácido oxaloacético a
partir do ácido aspártico e ácido α-cetoglutárico a partir do ácido glutâmico. Pelo facto de estas
reacções serem reversíveis, o ciclo fornece também os esqueletos carbonados para a síntese de
alguns dos aminoácidos não essenciais. Outros aminoácidos contribuem para a neoglicogénese,
porque a totalidade ou parte dos seus esqueletos carbonados são introduzidos no ciclo depois da
desaminação ou transaminação.10
O acetil-CoA, formado a partir do ácido pirúvico, pela acção da piruvato-desidrogenase, é o
principal composto que inicia a síntese dos ácidos gordos. No entanto, o acetil-CoA não consegue
atravessar a membrana mitocondrial interna, e portanto tem que ser transformado em ácido cítrico
para ser transportado para fora da mitocôndria, onde se refaz a acetil-CoA, uma vez que as enzimas
responsáveis pela síntese dos ácidos gordos são extramitocondriais e a piruvato-desidrogenase é
exclusivamente mitocondrial.11
9
Ver Neoglicogénese
Rever Metabolismo dos Aminoácidos
11
Ver síntese de ácidos gordos no capítulo dos Lípidos
10
Glúcidos página 73 de 122
Carlos Capela
Por tudo isto, o ciclo de Krebs é a placa giratória do metabolismo intermediário.
Glúcidos página 74 de 122
Carlos Capela
7.CADEIA TRANSPORTADORA DE ELECTRÕES
7.a. Conceito:
Também denominada por cadeia respiratória, é a via de convergência de todo o metabolismo
aeróbio da célula; é formada por uma sequência de compostos transportadores de electrões
localizados na membrana mitocondrial interna, e dirige um fluxo de pares de electrões das
coenzimas captadoras – NADH + H+, FADH2 – ao oxigénio molecular, com grande libertação de
energia.
O oxigénio, ao receber o par de electrões, reduz-se a água, e a energia libertada é dirigida para a
síntese do ATP, num processo acoplado ao transporte de electrões chamado Fosforilação Oxidativa.
½ O2 + 2 e- + 2H+ → H2O
7.a.i. EQUAÇÃO TERMODINÂMICA DA REOXIDAÇÃO DAS COENZIMAS:
NADH + H+ + ½ O2 → NAD+ + H2O
∆G = - 52,6 Kcal/Mol
FADH2 + ½ O2 → FAD + H2O ∆G = - 43,4 Kcal/Mol
7.b. A Cadeia de Transportadores:
Os transportadores de electrões da cadeia respiratória e a sua sequência estão descritos a seguir:
•
NADH-Desidrogenase: É o primeiro transportador da sequência; recebe os pares de
electrões do NADH e transfere-os para a Ubiquinona ou Coenzima Q. Possui um grupo
prostético FMN – Flavina Mononucleotídeo – que intermedia o processo.
NADH + H+ + FMN → NAD+ + FMNH2
•
Succinato-Desidrogenase: Actua no Ciclo de Krebs, e tem o FAD como grupo prostético.
Transfere os electrões do FADH2 directamente para a Ubiquinona.
Glúcidos página 75 de 122
Carlos Capela
•
Ubiquinona: Recebe os pares de electrões do NADH e do FADH2 e transfere-os para uma
sequência de Hemeproteínas denominadas Citrocomos.
•
Os Citocromos distribuem-se em 3 classes principais: Citocromos a, b e c:
o Citocromo b: É o primeiro citocromo da sequência a reduzir. Transfere os electrões da
ubiquinona para o citocromo c1.
o Citocromo c1: Recebe os electrões do b e transfere-os para o citocromo c.
o Citocromo c: Transfere os electrões do c1 para o citocromo a. Difere dos outros
citocromos por ser uma proteína hidrossolúvel.
o Citocromo a: Transfere os electrões de c para o citocromo a3.
o Citocromo a3: É o último citocromo da sequência, transferindo o par de electrões para o
oxigénio, que o reduz, formando uma molécula de água.
•
Oxigénio: É o aceitador final de electrões da cadeia respiratória. A sua redução a água é a
última etapa da respiração celular.
NADH + H+
↓
FMN (NADH-Desidrogenase)
↓
Ubiquinona ← FADH2 ← Ácido Succínico
↓
Citocromo b
↓
Citocrocromo c1
↓
Citocromo c
↓
Citocromo a
↓
Citocromo a3
↓
Oxigénio
Glúcidos página 76 de 122
Carlos Capela
7.c. Organização Multimolecular dos Transportadores de
Electrões:
Experiências através da actuação de detergentes sobre a membrana mitocondrial interna
demonstraram que os transportadores de electrões, com excepção da ubiquinona e do citocromo c,
estão organizados em 4 grandes complexos multimoleculares, a saber:
•
Complexo da NADH-Desidrogenase, ou Complexo I;
•
Complexo da Succinato-Desidrogenase ou Complexo II;
•
Complexo Citocromo bc1 ou Complexo III;
•
Complexo da Citocromo-Oxidase ou Complexo IV.
Glúcidos página 77 de 122
Carlos Capela
7.c.i. COMPLEXO I:
É o maior dos 4 complexos; formado por 26 cadeias polipeptídicas, incluindo 7 centros de
enxofre/ferro e a flavoproteina ligada ao FMN;
O sítio de ligação com o NADH está voltado para a matriz mitocondrial, favorecendo a
transferência de electrões.
A ubiquinona reduzida pelo complexo I difunde-se pela bicamada lipídica da membrana
mitocondria interna até o complexo III. A ubiquinona reduzida é a mediadora da transferência dos
electrões dos complexos I e II ao complexo III.
Os protões que acompanham os electrões são transferidos da matriz para o espaço
intermembranar.
7.c.ii. COMPLEXO II:
Formado principalmente pela succinato-desidrogenase, a única enzima do ciclo de Krebs que se
situa na membrana mitocondrial interna.
Possui 4 sub-unidades, incluindo 2 proteínas com grupos enxofre/ferro, e uma delas ligada ao
FAD.
Os sítios de oxi-redução do ácido succínico do complexo II, estão voltados para a matriz
mitocondrial.
7.c.iii. COMPLEXO III:
Formado por 2 tipos de citocromo b (bL e bH), pelo citocromo c1, uma proteína enxofre/ferro e
entre 4 a 6 proteínas adicionais.
O caminho dos electrões através do complexo III é sinuoso e complexo; o citocromo c que os
recebe está localizado na camada fosfolipídica da membrana mitocondria interna do lado do espaço
intermembranar da mitocôndria. O citocromo c, por ser hidrossolúvel, tem baixa afinidade para a
bicamada lipídica da membrana mitocondria interna e difunde-se através desta, mediando a ligação
entre os complexos III e IV.
O processo leva ao transporte de electrões até o citocromo c, e ao bombeamento de protões da
matriz mitocondrial para o espaço intermembranar.
Glúcidos página 78 de 122
Carlos Capela
7.c.iv. COMPLEXO IV:
Contém cerca de 13 sub-unidades polipeptídicas, e 2 átomos de cobre, além dos grupos heme
característicos dos citocromos a e a3.
Os electrões doados pelo citocromo c são transportados através dos átomos de cobre e ferro até
ao lado da matriz mitocondrial, onde vão reduzir o O2 em H2O.
A redução incompleta do O2 pode levar à geração de espécies reactivas de oxigénio como o ião
superóxido, o peróxido de hidrogénio e o radical hidroxilo, todos muito reactivos e tóxicos para as
células.
Glúcidos página 79 de 122
Carlos Capela
8.FOSFORILAÇÃO OXIDATIVA
8.a. Conceito:
Processo metabólico de síntese de ATP a partir da energia libertada pelo transporte de electrões
na cadeia respiratória.
•
Depende de alguns factores:
o Energia Livre: obtida do transporte de electrões;
o Uma enzima transmembranar denominada ATPase.
8.b. A Energia:
Durante o fluxo de electrões ocorre a libertação de energia livre suficiente para a síntese de ATP
em 3 locais da cadeia respiratória: Complexos I, III e IV. Estes locais são denominados “Sítios de
Fosforilação Oxidativa”.
Nestes locais a libertação de energia livre é em quantidade semelhante à necessária para a síntese
do ATP.
8.c. A Enzima ATPase:
Também denominada por ATP Sintetase, ou F1FoATPase, ou ainda, oxissoma.
É uma enzima de estrutura muito complexa, formada por 16 sub-unidades polipeptídicas
distribuídas em 2 fracções funcionais: As fracções Fo e F1.
8.c.i. A FRACÇÃO F1:
É semelhante a uma maçaneta cujo cabo seria a fracção Fo. Está ligada à membrana mitocondrial
interna, sempre voltada para o lado da matriz mitocondrial.
Glúcidos página 80 de 122
Carlos Capela
Possui 9 unidades polipeptídicas de 5 tipos diferentes – 3 α, 3 β, 1 γ, 1 δ e 1 ε – e vários sítios de
ligação com o ATP, ADP e fosfato.
Tem actividade de síntese do ATP, mas para isso precisa estar associada à fracção Fo. Quando
dissociada de Fo, só é capaz de hidrolisar o ATP.
8.c.ii. A FRACÇÃO FO:
Actua como um canal de protões através da membrana mitocondrial interna.
É formada por um conjunto de 9 a 12 polipéptidos localizados através da membrana mitocondrial
interna, e está ligada à F1 sempre no lado da matriz mitocondrial.
O “o” subscrito não é um zero, mas sim a letra inicial da palavra “oligomicina”, um potente
inibidor desta enzima e, por consequência, da fosforilação oxidativa.
Glúcidos página 81 de 122
Carlos Capela
8.d. Acoplamento
Entre
Fosforilação Oxidativa:
Cadeia
Respiratória
e
“Como é que a energia libertada no transporte de electrões é utilizada pela célula para a
síntese de ATP?”
Várias hipóteses já foram apresentadas para tentar explicar o processo;
Hipóteses como o “acoplamento químico”, ou o “acoplamento conformacional”, mas foram
abandonadas por falta de evidências experimentais concretas.
Actualmente, a hipótese aceita foi descrita por Peter Mitchell em 1961, e é designada por
“Hipótese Quimiosmótica de Mitchell”.
8.d.i. A HIPÓTESE QUIMIOSMÓTICA:
Segundo Mitchell, as condições necessárias para que a fosforilação oxidativa ocorra são:
•
Uma bomba de protões na cadeia respiratória, criando um fluxo da matriz para o citosol;
•
Uma membrana mitocondrial interna íntegra e impermeável a protões;
A partir desta situação, Mitchell prevê os seguintes eventos na membrana mitocondrial interna:
•
A Cadeia Respiratória, ao transportar os electrões utiliza a energia libertada para bombear
protões da matriz para o citosol;
•
A membrana mitocondrial interna, por ser impermeável a protões, impede o retorno destes à
matriz;
•
Gera-se assim um Gradiente Duplo – de pH e electrostático – através da membrana
mitocondrial interna, que gera uma situação de elevada instabilidade e, por consequência,
uma força que atrai os protões de volta à matriz;
•
Esta força, denominada Força Protão-Motriz, dirige o refluxo de protões para a matriz
mitocondrial através dos canais de protões da enzima ATPase;
•
A passagem dos protões pela ATPase determina a síntese do ATP.
Glúcidos página 82 de 122
Carlos Capela
8.d.ii. SUPORTE EXPERIMENTAL DA TEORIA DE MITCHELL:
Não existe nenhum intermediário rico em energia na Cadeia Respiratória e a fosforilação
oxidativa requer uma membrana mitocondrial interna intacta.
A membrana mitocondrial interna é impermeável a protões e outros iões como Cl-, OH- e K+.
A fosforilação oxidativa pode ser inibida por agentes ionóforos (transportadores de iões) e
desacopladores (transportadores de H+ através da membrana mitocondrial interna).
O fluxo de electrões na cadeia respiratória ejecta protões da matriz mitocondrial para o espaço
intermembranar da mitocondria.
Glúcidos página 83 de 122
Carlos Capela
8.d.iii. TRANSPORTE
DE
SUBSTRATOS
ATRAVÉS
DA
MEMBRANA
MITOCONDRIAL INTERNA
Como a membrana mitocondrial interna é altamente selectiva, existem através dela sistemas de
transporte de ATP, ADP, Pi e equivalentes redutores do NADH que permitem a troca destes
substratos entre a mitocondria e o citosol.
8.d.iii.1. Transporte de Pi:
Realizado por uma proteína específica que promove a troca de fosfato, que entra na matriz na
forma de iões fosfórico H2PO4-, com hidroxilos OH-.
8.d.iii.2. Transporte de ATP/ADP:
A saída do ATP da matriz para o citosol está condicionada com a entrada do ADP. Este processo
ocorre através da mesma proteína transportadora.
8.d.iii.3. Transporte de Equivalentes Redutores:
Os electrões do NADH que são obtidos em vias oxidativas citosólicas – como a cadeia
glicolítica, por exemplo, entram na mitocondria através de um sistema de transporte conhecido como
o Shuttle Ácido Málico/Acido Aspártico.
Através deste processo, o ácido oxaloacético é reduzido a ácido málico no citosol, este atravessa
a membrana mitocondrial interna para ser reoxidado a ácido oxaloacético com a redução do NAD+,
agora na matriz mitocondrial. O processo ocorre com gasto de energia.
Glúcidos página 84 de 122
Carlos Capela
8.e. Rendimento da Respiração Celular:
Ao calcularmos o rendimento em ATPs da oxidação total de uma molécula de glicose, e
considerando que cada par de electrões do NADH rende 3 ATPs, e cada par de electrões do FADH2
rende 2 ATPs na fosforilação oxidativa, temos:
Fenómeno
Saldo Energético (ATPs)
Glicólise
6 ou 8
Descarboxilação oxidante do ácido pirúvico
6
Ciclo de Krebs
24
Total:
36 ou 38
Este número pressupõe gasto de ATP zero em processos paralelos, o que não ocorre na prática.
Aceita-se como um número mais realista 30 ATPs/Glicose o rendimento real, considerando-se a
energia gasta durante todo o processo.
Glúcidos página 85 de 122
Carlos Capela
9.METABOLISMO DO GLICOGÉNIO
9.a. Introdução
O glicogénio é a principal forma de armazenamento de hidratos de carbono nos animais,
correspondendo ao amido nas plantas.
Localiza-se preferencialmente no fígado e músculos, sendo a quantidade superior nos músculos,
uma vez que a massa muscular é muito superior à massa do fígado.
O glicogénio age como fonte rapidamente disponível de unidades de hexose para a glicólise, no
músculo. O glicogénio hepático, por sua vez, encontra-se sobretudo relacionado com o
armazenamento e libertação de unidades de glicose para a manutenção da glicose sanguínea –
glicemia – particularmente no intervalo das refeições.
9.b. Síntese – Glicogénese
A Glicogénese é o processo pelo qual a glicose vai ser transformada em glicogénio.
Logo que entra na célula, a glucose é fosforilada a glucose-6-P pela enzima hexocinase:
A membrana celular é impermeável à glucose-6-fosfato, que pode por isso ser acumulada na
célula. A glucose-6-fosfato será utilizada na síntese do glicogénio (uma forma de armazenamento de
glucose), na síntese de outros compostos de carbono na via das pentoses fosfato, ou degradada para
produzir energia – glicólise.
Glúcidos página 86 de 122
Carlos Capela
Grandes quantidades de glucose-6-P dentro da célula provocam um aumento da pressão
osmótica. Nessas condições a água terá tendência a entrar para dentro da célula, provocando um
aumento do seu volume e eventual lise. Por isso, a glucose-6-P vai ser armazenada sob a forma de
um polímero: o glicogénio. O glicogénio é um polissacárido pouco solúvel (e que portanto não
provoca aumento da pressão osmótica), bastante ramificado e constituído exclusivamente por
monómeros de glucose unidos entre si por ligações α-1,4 e α-1,6 (nas ramificações):
Para poder ser utilizada na síntese do glicogénio, a glucose-6-fosfato é primeiro isomerizada a
glucose-1-fosfato, pela enzima Fosfoglucomutase.
A adição de glucose-1-P ao carbono 4’ de uma extremidade da cadeia de glicogénio não é uma
reacção favorecida termodinamicamente em condições fisiológicas, uma vez que o potencial de
transferência de fosfato das ligações C-O-P normais é bastante baixo. Por isso, a glucose-1-P vai ser
activada, isto é, vai ser transformada numa espécie com alto potencial de transferência de fosfato,
Glúcidos página 87 de 122
Carlos Capela
pela acção da UDP-Glicose pirofosforilase. Isto é conseguido por reacção com uridina trifosfatada
(UTP, uma molécula análoga do ATP, mas com uridina no lugar da adenina).
Esta reacção, só por si, não parece ser termodinamicamente favorável, pelo que se poderia pensar
que não teria utilidade. No entanto, o pirofosfato (PPi) que se forma nesta reacção pode ser
hidrolisado, numa reacção bastante exoenergética. A eliminação do PPi impele o equilíbrio no
sentido de formação da UDP-glucose, ilustrando mais uma vez o princípio da utilização de uma
reacção bastante exoenergética para tornar espontânea uma outra reacção que de outra forma não
seria favorecida termodinamicamente.
A UDP-glucose tem um elevado potencial de transferência de fosfato, o que lhe permite doar
glucose à extremidade 4’ de uma cadeia de glicogénio (ligações α-1,4), numa reacção catalizada pela
Glicogénio sintetase:
Glúcidos página 88 de 122
Carlos Capela
A glicogénio sintetase só consegue adicionar glucose a cadeias de glicogénio pré-existentes ou
primer, isto é, não é capaz de começar a síntese de uma nova molécula de glicogénio. A síntese do
glicogénio é iniciada pela adição de uma molécula de glucose a um resíduo de tirosina de uma
proteína denominada glicogenina.
As ramificações (ligações α-1,6) são realizadas por uma “enzima ramificadora”. Esta actua
sobre cadeias lineares de glicogénio com pelo menos 11 glicoses. A enzima ramificadora (amilo
(1,4→1,6)-transglicosilase) transfere segmentos terminais de glicogénio de cerca de 7 resíduos de
glicose para o grupo OH do carbono 6 de um resíduo de glucose (que pode estar na mesma ou noutra
cadeia). As ramificações devem estar a pelo menos 4 resíduos de distância uma da outra.
7 UDP-Glicose
Glicogénio
sintetase
7 UDP
Enzima
ramificadora
Glúcidos página 89 de 122
Carlos Capela
9.c. Degradação – Glicogenólise
A glicogenólise consiste na degradação de glicogénio a glicose. O glicogénio é degradado pela
acção conjunta de três enzimas:
•
Glicogénio fosforilase (é uma cinase), que cliva uma ligação α-1,4 com fosfato inorgânico
(Pi). Esta enzima remove os resíduos glicosil-1,4 das cadeias mais externas da molécula de
glicogénio até restarem, aproximadamente, 4 resíduos de glucose por ramificação. Utiliza
fosfato de piridoxal, um derivado da vitamina B6, como cofactor. É o passo limitante da
glicogenólise e nele forma-se glicose-1-P.
Uma molécula de glicogénio com ramos de apenas 4 glicoses (o que se denomina uma “dextrinalimite”) não pode ser degradada apenas pela glicogénio fosforilase. Necessita da acção da enzima
seguinte:
•
Enzima desramificadora do glicogénio: transfere três resíduos de glicose de um ramo
limite para outro ramo. O último resíduo da ramificação (com uma ligação α-1,6) é eliminado
por hidrólise, dando como resultado glucose livre e glicogénio desramificado. A hidrólise é
catalizada pela mesma enzima desramificadora.
Glúcidos página 90 de 122
Carlos Capela
A glicogénio fosforilase é bastante mais rápida do que a enzima desramificadora, pelo que os
ramos exteriores do glicogénio são degradados muito rapidamente no músculo em poucos segundos
quando é necessária muita energia. A degradação do glicogénio para lá deste ponto exige a enzima
desramificadora e é portanto mais lenta, o que explica em parte o facto do músculo só poder exercer
a sua máxima força durante poucos segundos.
•
Fosfoglucomutase: cataliza a isomerização de glucose-1-P a glucose-6-P, e vice-versa:
Glúcidos página 91 de 122
Carlos Capela
A glucose 6-fosfato pode então ser utilizada na glicólise. Ao contrário do músculo, o fígado (e
em menor extensão, o rim) possui glucose-6-fosfatase, uma enzima hidrolítica que cataliza a
desfosforilação da glucose 6-fosfato, o que lhe permite fornecer glucose ao resto do organismo:
9.d. Regulação do metabolismo do glicogénio
A regulação do metabolismo do glicogénio faz-se, essencialmente, através de duas enzimas
fundamentais, a glicogénio sintetase e a glicogénio fosforilase. O cAMP (é um sinalizador de baixos
níveis energéticos) desempenha, um papel fundamental na regulação destas enzimas, pois medeando
a fosforilação destas enzimas, inibe a sintetase e estimula a fosforilase, actuando, assim, no sentido
da glicogenólise.
A glicogénio sintetase existe sob duas formas. A forma a, não fosforilada, activa, e a forma b,
fosforilada, inactiva. A forma a é fosforilada por uma quinase. A quinase tem, por sua vez, uma
forma inactiva (I) e uma forma activa (A). A forma I é activada pelo cAMP.
Glúcidos página 92 de 122
Carlos Capela
A glicogénio fosforilase também existe sob duas formas: a e b. A forma a é activa e a forma b
não. A forma b converte-se na forma a pela acção da quinase, na presença de ATP e Mg2+. Como
vimos anteriormente, a quinase existe sob duas formas – activa (A) e inactiva (I) – intervindo na
activação o cAMP formado pela adenilciclase.
As fosforilações geralmente dão-se nos resíduos de serina, originando-se fosfoserina.
A desfosforilação é assegurada principalmente pela fosfoproteína fosfatase I, que pode
desfosforilar a glicogénio sintetase, a glicogénio fosforilase e a quinase. Esta converte a glicogénio
fosforilase a na forma b, a glicogénio sintetase b na glicogénio sintetase a, e inactiva a quinase.
A fosfoproteína fosfatase é inibida pelo inibidor proteico I, que se forma por fosforilação da sua
forma inactiva pela proteína quinase formada pelo cAMP. O inibidor é inactivado ao ser
desfosforilado por uma fosfoproteína fosfatase I.
Glúcidos página 93 de 122
Carlos Capela
Podemos concluir que o principal regulador do metabolismo do glicogénio é o cAMP, pois
estimula a glicogenólise e inibe a glicogénese dependendo, portanto, o sentido do metabolismo do
glicogénio do balanço dos mecanismos estimuladores e inibidores do cAMP.
9.d.i. REGULAÇÃO HORMONAL DO METABOLISMO DO GLICOGÉNIO NO
MÚSCULO
9.d.i.1. A Epinefrina ou Adrenalina
Em situações de stress, a medula supra-renal liberta para a circulação grandes quantidades de
adrenalina e alguma nor-adrenalina. Estas hormonas estimulam a produção de cAMP e
consequentemente a glicogenólise. A adrenalina combina-se com uma proteína específica existente
no interior da membrana, o receptor adrenérgico. O complexo adrenalina-receptor na presença de
GTP combina-se com a proteína G, que traduz o sinal capaz de activar a adenilciclase.
Glúcidos página 94 de 122
Carlos Capela
9.d.i.2. A insulina
A insulina facilita o transporte da glicose para o interior da célula, estimula a glicogénio sintetase
favorecendo, assim, a glicogénese, uma vez activa a fosfoproteína fosfatase I. É uma hormona
hipoglicemiante.
9.d.ii. REGULAÇÃO DO METABOLISMO DO GLICOGÉNIO NO FÍGADO
A regulação do metabolismo do glicogénio no fígado utiliza os mesmos mecanismos gerais que
descrevemos para o músculo, embora haja diferenças quanto às hormonas intervenientes.
9.d.ii.1. O Glucagón ou Glucagina
No fígado, a glucagina toma o lugar da adrenalina. Como resposta a uma descida da glicemia
(hipoglicémia), as células do pâncreas (células α dos ilhéus de Langerhans) secretam glucagina que,
ao combinar-se a um receptor específico da membrana celular, estimula a adenilciclase (por um
mecanismo semelhante ao da adrenalina), favorecendo assim, a glicogenólise. A glicose-6-fosfato,
formada pela glicogenólise no fígado, pode transformar-se em glicose pela acção da glicose-6fosfatase, enzima que não existe no músculo. A glicose é exportada para a corrente sanguínea,
repondo assim os valores normais de glicemia. É uma hormona hiperglicemiante.
9.d.ii.2. Cálcio
O aumento do cálcio citoplasmático tem também um efeito regulador. O aumento dos níveis de
cálcio deve-se a 2 hormonas: a vasopressina ou ADH e a adrenalina.
Tanto a vasopressina como a adrenalina combinam-se com receptores específicos que, no caso
da adrenalina são os receptores adrenérgicos. O complexo hormona-receptor vai activar um lípido da
membrana, o fosfatidil-inositol-4,5-bisfosfato, que se irá cindir em dois mensageiros, o inositol1,4,5-trifosfato (IP3) e o 1,2-diacilglicerol. O IP3 liberta cálcio do retículo endoplasmático, que por
sua vez, irá activar a quinase. O 1,2-diacilglicerol activa directamente a quinase. Em suma, a acção
destes dois mensageiros conduz à glicogenólise.
Glúcidos página 95 de 122
Carlos Capela
9.d.ii.3. A insulina
A insulina estimula a glicogénio sintetase favorecendo, assim, a glicogénese, uma vez que activa
a fosfoproteína fosfatase I. É uma hormona hipoglicemiante.
9.d.ii.4. Glicose
A glicose tem uma acção reguladora, uma vez que se combina com a glicogénio fosforilase a
convertendo-a num substrato óptimo para as fosfatases. Por outro lado, activa a glicogénio sintetase.
Em suma, o excesso de glicose favorece a glicogénese.
Glúcidos página 96 de 122
Carlos Capela
10. NEOGLICOGÉNESE
10.a. Introdução
Existem duas formas principais de manter os níveis de glucose no sangue entre as refeições: a
degradação do glicogénio e a Neoglicogénese ou Gliconeogénese.
Define-se Neoglicogénese como a formação de glicose a partir de material não glucídico e
do ácido láctico.
Os órgãos com elevada capacidade neoglicolítica são o Fígado e o Rim. Estes processos
realizam-se em situações de fome prolongada.
As reacções irreversíveis da glicose impedem que a neoglicogénese seja uma simples
reversão do processo. Há 3 reacções que, por razões de ordens termodinâmica, não são reversíveis
nas condições fisiológicas:
•
Transformação do Ácido Fosfoenolpirúvico em Ácido Pirúvico – Piruvato-cinase
•
Fosforilação da Frutose-6-fosfato a Frutose-1,6-Bisfosfato – Fosfofrutocinase I
Glúcidos página 97 de 122
Carlos Capela
•
Fosforilação da glicose a glicose-6-fosfato – Glicocinase ou Hexocinase
A estes níveis, a neoglicogénese utiliza reacções catalisadas por enzimas diferentes.
Glicólise
Neoglicogénese
•
Hexocinase
•
Glucose-6-fosfatase
•
Fosfofrutocinase I
•
Frutose-1,6-bisfosfatase
•
Piruvato cinase
•
Piruvato-carboxilase
•
Fosfoenolpiruvato-carboxicinase
10.b. Substratos da Neoglicogénese
10.b.i. AMINOÁCIDOS GLICOGÉNICOS OU GLICOFORMADORES
Aminoácidos que por desaminação ou transaminação originam ácido pirúvico ou
intermediários do Ciclo de Krebs:
•
Ácido α-cetoglutárico: ácido glutâmico, glutamina, histidina, arginina e prolina;
•
Ácido Oxaloacético: ácido aspártico e asparagina;
•
Ácido Pirúvico: alanina, triptofano, serina, treonina, cisteína e glicina;
•
Succinil-CoA: treonina, valina, isoleucina e metionina;
•
Ácido Fumárico: tirosina e fenilalanina.
Glúcidos página 98 de 122
Carlos Capela
10.b.ii. LÍPIDOS
•
GLICEROL: é um dos produtos do metabolismo do tecido adiposo e só os tecidos que
possuem a enzima activadora, a Glicerolcinase, podem utilizá-lo. Esta requer ATP e
catalisa a conversão de Glicerol a Glicerol-3-fosfato. Este vai ser oxidado a Dihidroxiacetona fosfato pelo NAD+, na presença da Glicerol-3-P-desidrogenase. Esta
enzima é encontrada entre outros tecidos, especialmente no fígado e nos rins.
•
ÁCIDOS GORDOS: os ácidos gordos com um número par de carbonos originam AcetilCoA. Por outro lado, o Acetil-CoA activa a Piruvato-carboxilase e inibe a Piruvatodesidrogenase. Os ácidos gordos com um número impar de carbonos, para além de
originarem Acetil-CoA, também originam Proprionil-CoA, que depois se transforma em
Succinil-CoA.
10.b.iii. OUTROS AÇÚCARES
A frutose, a galactose e a manose podem entrar na via da neoglucogénese.12
10.c. Neoglicogénese ou Gliconeogénese
Na gluconeogénese, cada um dos passos irreversíveis da glicólise, é substituído por reacções
termodinamicamente favoráveis.
10.c.i. TRANSFORMAÇÃO
DO
ÁCIDO
PIRÚVICO
EM
ÁCIDO
FOSFOENOLPIRÚVICO
Dos três passos, a síntese do Ácido fosfoenolpirúvico a partir do ácido pirúvico é o mais
exigente em termos energéticos, por ter um ∆G bastante positivo. Para ultrapassar a barreira
termodinâmica, esta reacção vai ser acoplada a uma descarboxilação, uma estratégia usada
frequentemente pela célula para impelir um equilíbrio no sentido da formação de produtos, como se
viu em várias reacções do ciclo de Krebs. Como quer o ácido pirúvico quer o ácido
fosfoenolpirúvico (PEP) são compostos com três carbonos, isto implica uma carboxilação prévia,
cuja energia provém da hidrólise do ATP. A descarboxilação do ácido oxaloacético assim formado
produz a energia necessária para a fosforilação do carbono 2 pelo GTP, dando origem ao PEP (numa
reacção catalizada pela fosfoenolpiruvato carboxicinase – PEPCK).
12
Ver o metabolismo das outras oses
Glúcidos página 99 de 122
Carlos Capela
Ácido Oxaloacético
Ácido pirúvico
Piruvato-carboxilase
Ácido Málico
Malato-desidrogenase (mit.)
Ácido Oxaloacético
Ácido Málico
Fosfoenolpiruvato-carboxicinase (cit.)
Malato-desidrogenase (cit.)
Ácido Fosfoenolpirúvico
A enzima responsável pela carboxilação do ácido pirúvico (a piruvato carboxilase) existe na
matriz mitocondrial, e contém biotina (uma coenzima transportadora de CO2 retirado ao HCO3-); ao
passo que as outras enzimas da neoglicogénese são citoplasmáticas. Por outro lado o ácido
oxaloacético (OAA) formado nesta reacção é incapaz de atravessar a membrana interna da
mitocôndria. Como resolve o organismo este problema?
Pode sair da mitocôndria apenas depois de transformado em ácido Málico ou Aspártico. A
escolha do processo depende da disponibilidade de NADH (necessário para a gluconeogénese) no
citoplasma. Se houver NADH suficiente no citoplasma (por exemplo: se se estiver a realizar
gluconeogénese a partir do ácido láctico) o OAA é transaminado a ácido Aspártico. Caso contrário,
Glúcidos página 100 de 122
Carlos Capela
o OAA é reduzido a ácido málico, que sai da mitocôndria para o citoplasma, onde é novamente
oxidado a OAA com produção simultânea de NADH. O OAA é então descarboxilado a PEP pela
PEPCK citoplasmática. Em humanos, existe também uma PEPCK mitocondrial.
10.c.ii. CONVERSÃO
DA
FRUTOSE-1,6-BISFOSFATO
À
FRUTOSE-6-
FOSFATO E HIDRÓLISE DA GLICOSE-6-FOSFATO
As reacções catalizadas pela fosfofrutocinase I e pela hexocinase são substituídas na
gluconeogénese por reacções hidrolíticas. Neste ponto, em vez de fosforilar ADP a ATP (o inverso
da glicólise, mas desfavorecido termodinamicamente em condições fisiológicas), ocorre a libertação
do fosfato por hidrólise. Esta é a enzima chave, no sentido que a sua presença determina se um
tecido é ou não é capaz de ressintetizar o glicogénio ou glicose a partir do ácido pirúvico e das
triose-fosfato. Julga-se que não existe nos músculos cardíacos e liso.
A frutose 1,6-bisfosfatase existe em quase todos os tecidos, mas a glucose-6-fosfatase
existe apenas no fígado e no rim, o que lhes permite fornecer glucose ao resto do organismo. A
glicose só pode passar através da membrana celular para o meio exterior depois de desfosforilada.
Esta enzima encontra-se na membrana do Retículo endoplasmático.
Glúcidos página 101 de 122
Carlos Capela
10.d. Balanço energético
A síntese de glicose requer ATP. São necessárias 6 moléculas de ATP para a síntese de uma
molécula de glicose a partir de 2 moléculas de ácido láctico. O ATP de que as células hepáticas
necessitam para a síntese de glicose é fornecido em grande parte pela oxidação dos ácidos gordos.
As condições metabólicas sob as quais o fígado deve sintetizar a glicose, favorecem, geralmente, um
aumento na disponibilidade de ácidos gordos no sangue provenientes das reservas adiposas. Esses
ácidos gordos são oxidados, pelas mitocôndrias hepáticas a corpos cetónicos com a produção
concomitante de grandes quantidades de ATP. Esse ATP é utilizado para suportar as necessidades
energéticas da neoglicogénese, independentemente do substrato usado como fonte de carbono para o
processo.
10.e. Regulação
A glicólise e a neoglicogénese são controladas pelos mecanismos para que seja possível que
apenas uma das vias funcione. A inibição da glicólise nos seus pontos principais, ou a repressão da
síntese das enzimas envolvidas nesses pontos, favorece a efectividade das enzimas neoglicogénicas
opostas.
A fosfofrutocinase é estimulada pelo AMP e inibida pelo ATP e ácido cítrico. Estes têm uma
acção oposta sobre a frutose-1,6-bisfosfatase. Assim, quando há um baixo nível energético, indicado
por concentrações baixas de ATP e elevadas de AMP, a frutose-1,6-bisfosfatase é inibida e a
glicólise favorecida. Por outro lado, quando o nível energético é elevado, indicado por elevadas
concentrações de ATP, frutose-1,6-bisfosfatase é activada enquanto a fosfofrutocinase é inibida,
favorecendo assim a neoglicogénese.
A frutose-2,6-bisfosfato também tem uma acção regulatória.13
A piruvato-cinase é inibida pelo ATP e alanina, ao contrário da carboxicinase, que é inibida
pelo ADP e estimulada pelo ATP e acetil-CoA. Aqui, também, os elevados níveis energéticos
favorecem a neoglicogénese e os baixos níveis a glicólise.
Para melhor explicitar esta regulação, temos como exemplo, a oxidação de ácidos gordos.
Esta oxidação faz mais do que simplesmente fornecer ATP para o processo. Promove a síntese de
glicose, através do aumento da concentração no estado estacionário de Acetil-CoA mitocondrial, um
13
Ver regulação da glicólise: regulação da via propriamente dita.
Glúcidos página 102 de 122
Carlos Capela
efector alostérico positivo da Piruvato-carboxilase mitocondrial. A elevação de acetil-CoA e da
actividade da Piruvato-Carboxilase resulta numa maior síntese de ácido cítrico, um efector negativo
da fosfofrutocinase e positivo da frutose-1,6-bisfosfatase. A inibição desta enzima provoca uma
diminuição na concentração de Frutose-1,6-bisfosfato, um activador da piruvato-cinase. Assim,
reduz-se o fluxo do ácido fosfoenolpirúvico a ácido pirúvico, pela acção da piruvato-cinase,
aumentando por outro lado os esforços combinados da piruvato-carboxilase e da fosfoenolpiruvatocarboxicinase, na conversão do ácido pirúvico a ácido fosfoenolpirúvico.
Em suma, um aumento dos níveis de ATP, com o consequente decréscimo dos níveis de
AMP, favorece a neoglicogénese através da inibição da fosfofrutocinase e da piruvato-cinase, e da
activação da frutose-1,6-bisfosfatase.
Para terminar, é de referir que esta regulação faz-se não só de um modo passivo, regulandose a actividade das enzimas, mas também a sua síntese, alterando-se, essencialmente, a velocidade de
transcrição. Por exemplo, a insulina, estimula a síntese de fosfofrutocinase e da piruvato-cinase,
enquanto a glucagina têm uma acção oposta.
Glúcidos página 103 de 122
Carlos Capela
Glúcidos página 104 de 122
Carlos Capela
10.f. Ciclos dos Cori e de Felig14
Dois ciclos importantes entre tecidos que envolvem a neoglicogénese são conhecidos, o ciclo
dos Cori e o ciclo de Felig ou da Alanina. Estes ciclos dependem da neoglicogénese no fígado,
seguida da libertação da glicose e o seu uso num tecido periférico.
10.f.i. CICLO DOS CORI
No músculo, no decorrer de um esforço muscular intenso, forma-se ácido láctico. Este tecido,
não tem a capacidade de fazer a reacção inversa, para que o ácido láctico se converta em ácido
pirúvico. Por isso, o ácido láctico é transportado para o fígado ou para o rim, para aí ser oxidado a
ácido pirúvico.
10.f.ii. CICLO DA ALANINA OU CICLO DE FEHLIG
Como o músculo não tem enzimas neoglicogénicas chave, o ácido pirúvico não se poderá
converter em ácido fosfoenolpirúvico. Todavia, no músculo, o ácido pirúvico pode sofrer uma
transaminação, originando alanina (uma das formas de transporte da amónia), que pode ir para o
fígado e aí ser reconvertida em ácido pirúvico e entrar na neoglicogénese.
14
Para os exemplos, utilizou-se o músculo, mas pode ser qualquer tecido que não tenha as enzimas neoglicogénicas
chave, ou seja, a capacidade de converter o ácido láctico a ácido pirúvico; e/ou este ultimo a ácido fosfoenolpirúvico.
Glúcidos página 105 de 122
Carlos Capela
Em suma, como o músculo não tem enzimas neoglicogénicas chave, o ácido pirúvico é
transportado ao fígado sob a forma de ácido láctico (ciclo dos Cori), ou como alanina (ciclo de
Fehlig).
Glúcidos página 106 de 122
Carlos Capela
11. HOMEOSTASE DA GLICOSE
O fígado dos mamíferos é o directo responsável pela manutenção dos níveis normais da
glicemia.
Quando o teor em glicose do sangue portal é elevado, o fígado aumenta a absorção de glicose, e
quando é baixo, liberta glicose. Por outras palavras, quando há excesso de glúcidos, funciona a
glicólise e a glicogénese; quando há falta, funciona a glicogenólise e a neoglicogénese.
11.a. Regulação Hormonal
O papel das hormonas já foi referido anteriormente. Aqui, iremos aborda-las numa visão de
conjunto.
11.a.i. O CAMP
Muitas hormonas utilizam o cAMP como segundo mensageiro através do sistema da
adenilciclase, e algumas podem actuar na sua destruição. O cAMP sinaliza um baixo nível
energético.
A acção do cAMP está relacionada com a formação de glicose pois estimula a neoglicogénese e
a glicogenólise, inibindo a glicólise.
11.a.ii. OS GLICOCORTICÓIDES – O CORTISOL
Os glicocorticóides, sintetizados e secretados pelo cortéx da supra-renal, dos quais o seu
representante mais quantitativo é o cortisol, actuam pelos seguintes mecanismos:
•
Permitem a neoglicogénese a partir das proteínas e lípidos, uma vez que aceleram os
catabolismos proteicos e lipídicos;
•
Ao activarem o metabolismo dos lípidos, forma-se mais acetil-CoA que activa a piruvato
carboxilase e a frutose-1,6-bisfosfatase, inibindo pelo outro lado, a enzimas glicolíticas
chave;
•
Induz a síntese das enzimas neoglicogénicas, uma vez que acelera a sua transcrição.
Em suma, estes mecanismos estimulam a neoglicogénese e inibem a glicólise. Os
glicocorticóides são hormonas hiperglicemiantes.
Glúcidos página 107 de 122
Carlos Capela
11.a.iii. A INSULINA
A insulina tem uma acção fundamental sobre o metabolismo glucídico. Para além de promover a
entrada de glicose nas células. Para além desta importante acção, possui outras como:
•
Estimulação e indução das enzimas glicolíticas chave;
•
Inibição das enzimas neoglicogénicas chave;
•
Indução das enzimas da lipogénese;
•
Diminuição da lipólise.
Em suma, a acção da insulina está ligada à diminuição da glicose, estimulando a glicogénese e
muito especialmente, a glicólise e estimulando a transformação de glúcidos em lípidos. É uma
hormona hipoglicemiante.
11.a.iv. A GLUCAGINA E A ADRENALINA
A glucagina e a adrenalina têm uma acção oposta à da insulina pois estimulam o cAMP. A
adrenalina tem um efeito mais poderoso do que a glucagina.
11.b. Fígado e Rim
O fígado é o órgão fundamental na regulação da glicemia. Funciona como uma reserva de
glicose na forma de glicogénio. É o órgão que possui a maior reserva de glicose, só superado pelo
tecido muscular, uma vez que a massa deste é bastante superior à sua.
Basicamente, tem como função descer a glicemia quando está elevada, e elevá-la quando está
baixa, mantendo-a dentro dos parâmetros biológicos recomendados, necessários à homeostase do
organismo.
O rim desempenha um importante papel, pois é responsável pela reabsorção de glicose até
valores séricos de cerca de 180 mg/100 ml, a partir dos quais o rim elimina a glicose na urina
(glicosúria). A glicosúria é um dos sinais da diabetes mellitus.
Glúcidos página 108 de 122
Carlos Capela
11.c. Outros órgãos e tecidos
11.c.i. O MÚSCULO
A insulina facilita a entrada de glicose nas células musculares, sendo esta passagem um factor
limitante e constituindo, assim, um sistema de regulação do metabolismo da glicose, criando
limitações à quantidade de glicose disponível para a glicólise e glicogénese.
A adrenalina estimula a glicogenólise com a formação final de glicose-6-fosfato, mas como o
músculo não possui a enzima glicose-6-fosfatase, toda a glicose-6-fosfato seguirá a via glicolítica.
11.c.ii. O TECIDO ADIPOSO
A insulina promove no tecido adiposo a lipogénese, devido ao excesso de acetil-CoA e ATP
fornecidos pela glicose. Como iremos ver no capítulo dos lípidos, os glícidos são passíveis de ser
transformados a ácidos gordos. É por esta razão que a ingestão de grandes quantidades de glúcidos
também provoca um acréscimo do tecido adiposo.
11.c.iii. CÉREBRO
Ao contrário das outras células do nosso organismo, que consomem, para a formação de energia,
não só glicose, mas também ácidos gordos livres (FFA) e corpos cetónicos, as células cerebrais
utilizam unicamente a glicose. Em situações de hipoglicémia prolongada, podem também utilizar
corpos cetónicos como fonte de energia.
A glicose entra nas células cerebrais em função de um gradiente de concentração, sem qualquer
acção da insulina (o Glut 3 é independente da insulina).
Glúcidos página 109 de 122
Carlos Capela
12. METABOLISMO DAS OUTRAS OSES
A maior parte das transformações das oses em energia, faz-se através do metabolismo do
glicogénio e da glicose. As outras hexoses transformam-se em glicose ou intermediários da glicólise,
assim como a glicose pode originar várias hexoses.
12.a. A frutose
O metabolismo da frutose pode seguir dois caminhos:
1. A hexocinase pode transformar a frutose em frutose-6-fosfato que será catabolisada a
glicose-6-fosfato. Esta via é pouco significativa.
2. A frutocinase transforma a frutose em frutose-1-fosfato, que seguidamente, pela acção da
frutose-1-fosfato aldolase, se cinde em gliceraldeído e dihidroxiacetona fosfato. O
gliceraldeído é fosforilado pela tioquinase, originando o gliceraldeído-3-fosfato. A triose
isomerase converte dihidroxiacetona fosfato em gliceraldeído-3-fosfato. Desta maneira a
frutose entra na glicólise através de 2 moléculas de gliceraldeído-3-fosfato.
Glúcidos página 110 de 122
Carlos Capela
12.b. A Galactose
A galactose, pela acção da galactocinase, é fosforilada em galactose-1-fosfato, que combinandose com a UDP-glicose, forma a UDP-galactose, que seguidamente, se epimeriza em UDP glicose
pela acção da UDP glicose epimerase.
Glúcidos página 111 de 122
Carlos Capela
12.c. O Ácido Glicurónico
12.c.i. SÍNTESE
A primeira etapa é a formação de UDP-Glicose. A glicose-6-fosfato será isomerizada em
glicose-1-fosfato pela fosfoglucomutase. Pela acção da UDP-Glicose pirofosforilase, a glicose-1fosfato combina-se com o UTP originando UDP-Glicose.
A UDP-glicose pela oxidação do álcool primário em C6 formará o UDP-glicuronato, que em
seguida, dará o ácido glicurónico.
12.c.ii. CATABOLISMO
O ácido glicurónico transforma-se em L-xilulose. A L-xilulose isomerisar-se-á em D-xilulose
que entrara no ciclo de Dickens-Horecker.
Glúcidos página 112 de 122
Carlos Capela
E. ANEXOS
1.PARA SABER MAIS … OS TRANSPORTADORES
DE GLICOSE
1.a. Introdução
A insulina é produzida em resposta à hiperglicémia. Esta quando captada pelos receptores
específicos na membrana plasmática das células promove a libertação de transportadores do interior
da célula para a membrana, sob a forma de vesícula de secreção.
O número ou a afinidade dos receptores de insulina – ou ambos – são afectados pela insulina e
por outras hormonas. A exposição a quantidades aumentadas de insulina diminui a concentração de
receptores (“down-regulation”), enquanto a exposição a níveis diminuídos de insulina aumenta a
afinidade e o número.
1.b. Transportadores de Glicose
A glicose penetra nas células por difusão facilitada, ou no intestino e nos rins por transporte
activo secundário com o Na+. No músculo, no tecido adiposo e em alguns outros tecidos, ela facilita
a sua própria entrada nas células, aumentando o número de transportadores de glicose nas
membranas celulares.
Os transportadores de glicose responsáveis pela sua difusão facilitada através das membranas
celulares constituem uma família de proteínas estreitamente relacionadas que atravessam a
membrana celular 12 vezes.
Glúcidos página 113 de 122
Carlos Capela
Orientação da proteína transportadora de
glicose na membrana plasmática. Existem 12
domínios
que
atravessam
a
membrana,
indicados pelas áreas sombreadas, e um local
de
glicosilação
(CHO)
no
exterior
da
membrana. As extremidades aminoterminal e
carboxiterminal localizam-se no citoplasma no
interior da célula.
Diferem dos transportadores de glicose dependentes de sódio SGLT 1 e SGLT 2, com os quais
não tem qualquer homologia, embora os SGLT também apresentem 12 domínios transmembranares;
o SGLT 1 e o SGLT 2 são responsáveis pelo transporte activo secundário de glicose para fora do
intestino e dos túbulos renais.
Particularmente nos segmentos transmembranares helicoidais 3, 5, 7 e 11, os aminoácidos dos
transportadores facilitadores parecem circundar canais pelos quais a glicose pode penetrar. Supõe-se
que a conformação modifica-se e a glicose é libertada no interior da célula.
Foram caracterizados sete transportadores diferentes de glicose, designados pela sua ordem de
descoberta GLUT 1 a GLUT 7. Eles contêm 492 a 524 aminoácidos e sua afinidade pela glicose
varia. Cada transportador parece ter evoluído para exercer tarefas especiais. O GLUT 4 é o
transportador no tecido muscular e adiposo, sendo estimulado pela insulina. O reservatório de
moléculas de GLUT 4 é mantido no citoplasma das células sensíveis à insulina e, quando essas
células são expostas à insulina, os transportadores deslocam-se rapidamente para a membrana
celular, aparentemente por exocitose. Quando cessa a estimulação pela insulina, os transportadores
retomam ao citoplasma, provavelmente por endocitose, ficando prontos para a próxima exposição à
insulina. Os outros transportadores GLUT parecem permanecer na membrana celular.
Nos tecidos em que a insulina aumenta o número de transportadores de glicose na membrana
celular, a fosforilação da glicose uma vez no interior da célula é regulada por outras hormonas.
Tanto a hormona do crescimento (GH) quanto o cortisol inibem a fosforilação em certos tecidos.
Entretanto, o processo normalmente é tão rápido que só constitui uma etapa limitadora da velocidade
do metabolismo da glicose quando a entrada de glicose está elevada.
A insulina também aumenta a entrada da glicose nos hepatócitos, porém não exerce esse efeito
por aumento do número de transportadores GLUT 4 nas membranas celulares. Na verdade, ela induz
a hexocinase, que aumenta a fosforilação da glicose de modo que a concentração intracelular de
glicose livre permanece baixa, facilitando a sua entrada no interior da célula.
Glúcidos página 114 de 122
Carlos Capela
Km (mM)15 Principais locais de expressão
Função
Transporte activo secundário (co-transporte de Na+-Glicose)
SGLT 1
Absorção de Glicose
0,1 a 1,0
SGLT 2
Absorção de Glicose
1,6
Intestino delgado, túbulos renais
Túbulos renais
Difusão Facilitada
GLUT 1
Captação basal de glicose
1a2
Placenta,
barreira
hematoencefálica,
cérebro, eritrócitos, rins, cólon, e muitos
outros órgãos.
GLUT 2
Sensor de glicose das células β;
12 a 20
transporte para fora das células
Células β dos ilhéus de Langerhans, fígado,
células epiteliais do intestino delgado, rins
epiteliais intestinais e renais
GLUT 3
Captação basal de glicose
<1
Cérebro, placenta, rins, e outros órgãos
GLUT 4
Captação
5
Músculo esquelético e cardíaco, tecido
de
glicose,
estimulada pela insulina
GLUT 5
Transporte de frutose
GLUT 6
Nenhuma
GLUT 7
Transportador
fosfato
de
no
adiposo, e outros tecidos
1a2
glicose-6-
Jejuno, esperma
−
Pseudogene
−
Fígado, outros órgãos?
retículo
endoplasmático
15
Km é a concentração de glicose em que o transporte corresponde à metade do valor máximo.
Glúcidos página 115 de 122
Carlos Capela
1.c. A absorção de glúcidos
As hexoses e pentoses são rapidamente absorvidas através da parede do intestino delgado.
Praticamente todas as hexoses são removidas antes de alcançarem a porção terminal do íleon. As
moléculas de açúcar passam das células da mucosa para o sangue, alcançando posteriormente a veia
porta.
O transporte da maioria das hexoses é singularmente afectado pela quantidade de Na+ no lúmen
intestinal; a presença de elevada concentração de Na+ na superfície das células da mucosa facilita o
influxo de açúcar para as células epiteliais, sendo inibido por baixas concentrações de Na+, porque a
glicose e o Na+ compartilham o mesmo co-transportador ou simporter, o transportador de glicose
dependente de sódio (SGLT, sodium-dependent glucose transporter, co-transportador de Na+glicose). Os membros dessa família de transportadores, SGLT 1 e SGLT 2, assemelham-se aos
transportadores de glicose responsáveis pela difusão facilitada, uma vez que cruzam a membrana
celular 12 vezes, possuindo as suas extremidades –COOH e –NH2 terminais no lado citoplasmático
da membrana. Entretanto não existe qualquer homologia com a série GLUT de transportadores, o
SGLT 1 e o SGLT 2 também são responsáveis pelo transporte da glicose para fora dos túbulos
renais.
Como a concentração intracelular de Na+ é baixa nas células intestinais e renais, como nas outras
células, o Na+ penetra na célula pelo seu gradiente de concentração. A glicose desloca-se com o Na+,
sendo libertada no interior da célula. O Na+ é transportado para os espaços intercelulares laterais,
enquanto a glicose é transportada pelo GLUT 2 para os capilares. Por conseguinte, o transporte da
glicose constitui um exemplo de transporte activo secundário, pois a energia para o transporte de
glicose é fornecida indirectamente pelo transporte de Na+ para fora da célula, o que mantém a
concentração de Na+ baixa no interior da célula, e como consequência entra mais Na+ e, portanto,
mais glicose.
O mecanismo que serve para a glicose também transporta a galactose. A frutose utiliza um
mecanismo diferente. A sua absorção é independente do Na+ ou do transportador de glicose e
galactose; com efeito, é transportada por difusão facilitada do lúmen do intestino para os enterócitos
pelo GLUT 5 e dos enterócitos para os capilares pelo GLUT 2. Parte da frutose é convertida em
glicose nas células da mucosa. O GLUT 5 pode também transportar glicose e galactose, mas a sua
afinidade para estas hexoses é extremamente baixa.
A insulina exerce pouco efeito sobre o transporte intestinal de açúcares. Nesse aspecto, a
absorção intestinal assemelha-se à reabsorção de glicose nos túbulos contornados proximais dos rins;
Glúcidos página 116 de 122
Carlos Capela
nenhum desses processos requer fosforilação e ambos continuam praticamente normais na diabetes.
A intensidade máxima de absorção de glicose pelo intestino é de cerca de 120 g/hora.
A reabsorção de glicose nos rins assemelha-se à que ocorre no intestino. A glicose e o Na+
ligam-se ao transportador comum SGLT 2 na membrana luminal, e a glicose é transportada para o
interior da célula à mediada que o Na+ se desloca para o interior devido ao seu gradiente eléctrico e
químico. Em seguida, o Na+ é bombeado para fora da célula, para os espaços intercelulares laterais,
enquanto a glicose é transportada pelo GLUT 2 para o líquido intersticial. Por conseguinte, o
transporte de glicose nos rins, bem como no intestino, é um exemplo de transporte activo secundário.
Glúcidos página 117 de 122
Carlos Capela
2. PARA SABER MAIS … A DIABETES MELLITUS
2.a. Introdução
É uma doença metabólica hereditária, caracterizada pela insuficiência da acção hormonal da
insulina, frequentemente pela diminuição ou ausência da secreção pelas células β dos ilhéus de
Langerhans do pâncreas, ou raramente por ineficácia no sistema receptor celular para a insulina. É
influenciada por múltiplos e complexos factores genéticos e ambientais, que interagem
potencializando a sua expressão patológica.
O conhecimento da diabetes é muito antigo, sendo uma das doenças metabólicas com um
historial bem definido na história da medicina. Para se classificar a diabetes mellitus, deve-se levar
em consideração factores clínicos importantes, sendo que a classificação mais correntemente
utilizada (e por isso talvez a menos correcta) divide os pacientes em dois grupos:
1. Diabetes do tipo I ou Insulino-Dependente – também denominada de diabetes infantojuvenil porque, geralmente, aparece na infância ou na adolescência, mas não é limitada a
estes pacientes;
2. Diabetes do tipo II ou Insulino-Independente – também denominada diabetes do adulto
obeso, por ocorrer, geralmente, em indivíduos obesos, de meia-idade.
O carácter hereditário da diabetes mellitus está relacionado com um gene regulador da produção
de anticorpos anti-células β, localizado no braço curto (p) do cromossoma 6, devendo existir,
provavelmente, factores ambientais que estimulam a sua expressão génica mais precoce ou tardia, o
que justifica as diferentes faixas etárias de manifestação da sintomatologia.
Glúcidos página 118 de 122
Carlos Capela
2.b. Diabetes Mellitus Insulino-Dependente
Em contraste com a diabetes insulino-independente, há uma ausência completa da produção de
insulina pelo pâncreas nesta doença.
Devido à produção defeituosa de insulina pela células β, os níveis de insulina sanguínea não
aumentam em resposta aos níveis elevados de glicose sanguínea (Hiperglicémia)16. Mesmo quando a
glicose da alimentação está sendo absorvida pelo intestino, a relação insulina/glucagina não pode
aumentar e o fígado continua neoglicogénico e cetogénico. Como é impossível, mudar para
processos de glicólise, glicogénese e lipogénese, o fígado não pode regular apropriadamente os
níveis de glicemia. De facto, como a neoglicogénese é contínua, o fígado contribui para a
hiperglicémia, no estado bem alimentado. A incapacidade de alguns tecidos, especialmente o
músculo, de captar glicose na ausência de insulina, contribui ainda mais para a hiperglicémia. A
neoglicogénese acelerada, sustentada pela proteólise tecidular mantém a hiperglicémia, mesmo no
estado de jejum.
Paralelamente, há a extrapolação do limiar renal da glicose (a partir ± 160 mg/dl de glicemia) e a
sua libertação na urina (Glicosúria). Devido à hiperglicemia há perda osmótica de água a nível
tubular renal, promovendo perda excessiva de urina (Poliúria), o que induz um processo de
desidratação, levando o diabético a beber água exageradamente (Polidipsia).
A ausência da insulina provoca também lipólise acentuada no tecido adiposo, e
consequentemente um aumento dos níveis plasmáticos de ácidos gordos, e numa acelerada produção
de corpos cetónicos pelo fígado. Se os corpos cetónicos não forem usados tão rapidamente quanto
são formados, desenvolve-se cetoacidose diabética, devido ao acumulo de corpos cetónicos. O
excesso de corpos cetónicos provoca a sua eliminação pela respiração, dando ao hálito um cheiro
adocicado (hálito cetónico), e pela urina (cetonúria). O carácter ácido dos corpos cetónicos é
responsável pela queda acentuada do pH sanguíneo, que acarretará consequências nefastas ao
equilíbrio ácido-base, podendo levar, inclusive, à morte, associado a outras complicações clínicas
envolvidas no processo. O baixo pH plasmático estimula o centro respiratório, produzindo a rápida
respiração profunda, descrita por Kussmaul como “fome de ar” e denominada em sua homenagem
por respiração de Kussmaul.
Mas, nem todos os ácidos gordos captados pelo fígado, podem seguir a via da oxidação e da
cetogénese. O excesso é esterificado e direccionado para a síntese de VLDL. Assim, como resultado,
vamos ter uma hipertriacilgliceridémia porque as VLDLs são sintetizadas e libertadas pelo fígado
16
Valores normais: 70-110 mg/dl
Glúcidos página 119 de 122
Carlos Capela
mais rapidamente do que são depuradas do sangue pela lipoproteina lipase. A quantidade desta
enzima é dependente do nível de insulina no sangue. O defeito da lipoproteina lipase também resulta
numa hiperquilomicranémia, um vez que esta enzima também é necessária para o catabolismo das
quilomicras, no tecido adiposo.
Em suma, na diabetes insulino-dependente, cada tecido continua a executar o seu papel
catabólico para o qual foi designado no jejum, apesar da absorção de combustível adequada, ou
mesmo em excesso, no intestino. Isto resulta numa elevação de todos os combustíveis no sangue,
com severa perda dos tecidos corporais e, finalmente, morte, a menos que a insulina seja
administrada. A insulina exógena promove a captação de glicose pelos tecidos e inibe a
neoglicogénese, a lipólise e a proteólise.
Glúcidos página 120 de 122
Carlos Capela
2.c. Diabetes Mellitus Insulino-Independente
Em contraste com a diabetes insulino-dependente, a insulina não está ausente na diabetes
insulino-independente.
De facto, níveis normais a elevados de insulina podem ser observados nesta forma de diabetes e
o problema é, principalmente, resistência à acção da insulina. A obesidade muitas vezes precede o
desenvolvimento da diabetes insulino-independente e parece ser o principal factor contribuinte.
Pacientes obesos, são, geralmente, hiperinsulinêmicos. A resistência à insulina é um fenómeno
pouco entendido no qual os tecidos não respondem à insulina. O número ou a afinidade dos
receptores de insulina está reduzido em alguns pacientes; outros apresentam ligação normal da
insulina, porém respostas pós-receptores anormais, como a activação do transporte de glicose. Como
regra geral, quanto maior a massa de tecido adiposo num organismo, maior a resistência das células
normalmente insulino-sensíveis à acção da insulina. Dados bem recentes sugerem que níveis
aumentados da expressão do factor de necrose tumoral α (TNF-α “Tumor Necrosis Factor-α”), em
adipócitos de indivíduos obesos, contribuam para a resistência. Quanto maior a massa do tecido
adiposo, maior a produção de TNF-α, que actua prejudicando o funcionamento do receptor de
insulina.
Como consequência, os níveis de insulina plasmática estão muito elevados no sangue de um
indivíduo obeso. Enquanto as células β do pâncreas produzirem a insulina suficiente para superar a
resistência à insulina, um indivíduo obeso terá níveis sanguíneos relativamente normais de glicose e
lipoproteinas. Mas, embora os níveis de insulina de pacientes diabéticos insulino-independente
possam estar, muitas vezes elevados, não são tão elevados quanto num indivíduo não diabético,
porém igualmente obeso. As células β dos ilhéus de Langerhans desses pacientes diabéticos não
produzem insulina suficiente para superar a resistência à insulina, induzida pela sua obesidade. Por
isso, esta forma de diabetes é também uma forma de falha das células β dos ilhéus de Langerhans.
Desta forma, a doença não é causada somente pela resistência à insulina, mas também por
funcionamento prejudicado das células β.
A hiperinsulinémia constante pode agravar a situação, pois a partir de um certo nível deixa de
estimular o receptor de insulina (e a consequente transdução de sinal, exocitose das vesículas
contento os transportadores de glicose, e logicamente a entrada de glicose na célula), tendo mesmo
um carácter inibitório, provocando a diminuição dos receptores de insulina.
Glúcidos página 121 de 122
Carlos Capela
A dieta, por si só, já é capaz, frequentemente, de controlar a doença em diabéticos obesos. Se o
paciente puder ser motivado a perder peso, os receptores de insulina aumentarão em número e as
anormalidades pós-receptor melhoram, o que aumentará a sensibilidade tecidual à insulina.
Outra solução é a administração de insulina exógena que reduzirá a hiperglicémia, esta deve ser
administrada frequentemente para controlar os níveis de glicemia de pacientes diabéticos insulinoindependente.
A hiperglicémia resulta, principalmente, da captação insuficiente de glicose pelos tecidos
periféricos, especialmente o músculo. Em contraste com a diabetes insulino-dependente, a
cetoacidose não se desenvolve porque os adipócitos permanecem sensíveis aos efeitos da insulina
sobre a lipólise. Hipertriacilgliceridémia é característica da diabetes insulino-independente, mas
geralmente resulta de um aumento nas VLDLs, sem hiperquilomicronemia (uma vez que a
lipoproteina lipase é activada pela acção da insulina). Isto é provavelmente explicado pelas
velocidades rápidas da síntese hepática de novo de ácidos gordos, estimulada pela hiperglicémia e
hiperinsulinémia.
Glúcidos página 122 de 122