Resumo - capítulos 1 e 3
Pedro Ivo Gomes de Faria
Sumário
1 Capítulo 1 - A Evolução da Célula
1.1 De moléculas à primeira célula . . . . . . . . . . . . . . . . . .
1.2 De procariontes a eucariontes . . . . . . . . . . . . . . . . . .
1.3 De organismos unicelulares a multicelulares . . . . . . . . . . .
2
2
3
3
2 Capítulo 3 - Macromoléculas: Estrutura,
mação
2.1 Ácidos nucleicos . . . . . . . . . . . . . . .
2.2 Estrutura proteica . . . . . . . . . . . . .
2.3 Proteínas como catalisadores . . . . . . . .
4
4
6
7
1
Formato e Infor. . . . . . . . . . .
. . . . . . . . . . .
. . . . . . . . . . .
1
1.1
Capítulo 1 - A Evolução da Célula
De moléculas à primeira célula
Todos os organismos vivos são constituídos de células que (submetidas à
seleção natural e à variação) evoluíram a partir de um ancestral comum. Segundo a hipótese de Oparin, os primeiros compostos orgânicos (como aminoácidos e açúcares) da Terra foram originados a partir de reações químicas entre os compostos inorgânicos existentes, promovidas pela alta temperatura
do planeta e pela ocorrência de descargas elétricas na atmosfera. O experimento de Miller comprovou que a hipótese era razoável, pois tais moléculas
foram obtidas no sistema fechado que criou para simular as condições da
Terra primitiva.
Moléculas orgânicas simples (aminoácidos e nucleotídeos) podem se unir
para formar cadeias de polímeros, que são vistos como alguns dos principais constituintes da célula. Em particular, polinucleotídeos são capazes de
orientar a síntese de outros polinucleotídeos (através da afinidade existente
entre os pares de bases (Adenina,Timina) , (Citosina, Guanina) e (Adenina,
Uracila) ). O RNA (um tipo de polinucleotídeo) transporta toda a informação que o caracteriza em sua cadeia de nucleotídeos, entre as quais seu
comportamento químico (determinado pela sua estrutura tridimensional).
Existem evidências de que (entre 3.5 e 4 bilhões de anos atrás) o processo
de evolução começou a partir de moléculas de RNA capazes de autorreplicação, entre as quais foram selecionadas as que também conseguiam catalisar
a reação. Porém, polipeptídeos possuem eficiência catalítica muito maior que
a de nucleotídeos; e são através de proteínas especializadas (enzimas) que as
reações são catalisadas em células atuais. Logo, parece provável que o RNA
tenha guiado (de um modo primitivo) a síntese de proteínas.
Além da síntese de proteínas, seria necessário que o RNA que a produziu
as mantivesse em suas adjacências (pois, em caso contrário, as proteínas produzidas poderiam beneficar qual espécie de RNA) . A partir do surgimento
espontâneo de membranas fosfolipídicas (que isolaram determinados filamentos de RNA do meio externo), os RNAs que sintetizavam enzimas mais eficientes sofreram seleção natural positiva (e assim surgiram as primeiras células). Um dos últimos estágios evolucionários (o das céluas atuais) sugere que
células que desenvolveram outro polinucleotídeo (o DNA) para armazenar a
infomação hereditária (em vez de fazê-lo no RNA) novamente sofreram seleção positiva, pois a estrutura em dupla hélice do DNA torna a célula mais
robusta e estável (e também permite um mecanismo de reparação que opera
usando o filamento intacto para corrigir o filamento danificado associado estima-se que esse mecanismo corrija 99,9% dos erros).
2
1.2
De procariontes a eucariontes
As células atuais são classificadas em eucariontes (com uma membrana denominada carioteca envolvendo o núcleo) ou procariontes (sem núcleo diferenciado), e acredita-se que aquelas tenham evoluído a partir dessas. Apesar
de serem simples, células procariontes (como as de bactérias) são capazes de
se reproduzir rapidamente (por fissão binária), ocupam vários nichos ecológicos (vivendo até mesmo em fontes termais) e compõem a maior parte das
células existentes.
Como não havia oxigênio na Terra primitiva, acredita-se que as primeiras
células (eucariontes) obtinham energia (na forma de ATP) através da quebra
anaeróbica de glicose, cuja via metabólica evoluiu com o surgimento gradual
de enzimas capazes de catalisar diferentes transformações de metabólitos.
Entre os organismos mais autossuficientes estão as cianobactérias, que
conseguem transformar o gás carbônico (por fotossíntese) e o nitrogênio atmosférico em moléculas orgânicas (que podem ser metabolizadas). Com o
aumento do oxigênio atmosférico gerado pela fotossíntese, ocorreu um aumento na população de células capazes de fazer a glicólise aeróbia (respiração), processo mais eficiente energeticamente do que a anaeróbia.
Já as células eucariontes são mais complexas que as procariontes, possuindo ossuem membrana nuclear individualizada e vários tipos de organelas
(cada uma desempenhando uma função). Elas dependem das mitocôndrias
para obtenção de energia (através da respiração celular), sendo que as plantas
também possuem organelas capazes de produzir glicose a partir de água e gás
carbônico (através da fotossíntese, que ocorre nos cloroplatos). É provável
que tais organelas tenham surgido através da fagocitose de células procarióticas, que passaram a viver em simbiose com a célula hospedeira anaeróbica.
Seu material genético aparece em quantidade muito maior que o de célulasprocarióticas, e por isso é mantido no núcleo na forma (condensada) de
cromossomos. Os cromossomos se mnatém isolados do citoplasma graças à
carioteca, impedindo que interajam com as reações químicas que lá ocorrem.
1.3
De organismos unicelulares a multicelulares
Apesar de seres unicelulares comporem a maior parte da biomassa da Terra, a
“minoria” multicelular consegue utilizar muito melhor os recursos à sua volta
através da divisão de trabalho (cooperação) entre células especializadas. É
provável que o primeiro passo evolucionário na direção dos multicelulares
tenha sido a formação de colônias de unicelulares (que se beneficiam da concentração de enzimas digestórias assim obtida).
Uma das especializações mais importantes foi a formação de células epiteliais.
3
Graças ao tecido epitelial, foi possível desenvolver um ambiente interno no
qual as enzimas digestórias pudessem ser secretadas sem que se perdessem
ao meio externo.
O desenvolvimento de padrões complexos nos seres multicelulares só é
possível devido à memória celular: células eucarióticas persistem em seus
estados especializados mesmo que as influências que orientaram sua diferenciação tenham desaparecido. Isso é algo que se reflete no desenvolvivmento
embrionário, pois embriões de várias espécies diferentes (como um humano
e uma galinha, por exemplo) possuem estágios iniciais semelhantes, mas se
diferenciam radicalmente conforme alcançam os últimos estágios.
Apesar de grande parte das células de um mesmo organismo possuírem
o mesmo material genético (já que são provenientes de um mesmo zigoto),
a regulação gênica (ativação ou inibição de grupos de genes em resposta
a sianis internos ou externos) é a responsável por permitir que exista uma
grande variedade de especializações celulares (principalmente em plantas ou
animais superiores).
2
2.1
Capítulo 3 - Macromoléculas: Estrutura,
Formato e Informação
Ácidos nucleicos
Por um longo tempo (a partir de 1864, com os trabalhos de Mendel), a
genética trabalhou com o conceito abstrato de que as células possuíam fatores
(genes) que eram passados de geração em geração, responsáveis por transmitir
toda a informação hereditária. Foi apenas em 1953 que Watson e Crick
mostraram que a base física dos genes estava no DNA (ou no RNA, no caso
de alguns vírus).
Uma molécula de DNA consiste de duas cadeias poliméricas (formando
uma dupla hélice), cujos monômeros são os nucleotídeos (cada um deles formado por uma desoxirribose, um fosfato e uma base nitrogenada). A ligação
entre cada nucleotídeo é do tipo fosfodiéster, entre o carbono 5’ de uma desoxirribose e o 3’ da outra. Uma de suas principais características é que as
bases nitrogenadas de cadeia estão dentro da hélice, e formam pares complementares (adenina com timina e citosina com guanina) que mantém as
cadeias unidas (através de ligações de hidrogênio).
A própria estrutura do DNA sugere como a transferência de informação
genética é feita: como cadeia possui uma sequência de nucleotídeos complementar à cadeia associada, as duas carregam a mesma informação. Sendo A
e A’ duas cadeias complementares de uma mesma molécula, basta separá-las
4
para que A sirva como molde de uma outra cadeia A’ (analogamente para A’),
o que permite a formação de duas novas moléculas de modo semiconservativo
(duplicação do DNA).
Apesar da elevada precisão com que é feito o processo de duplicação
(menos de um erro a cada bilhão de nucleotídeos copiados), um único erro
(mutação) é suficiente para causar a morte da célula ou criar um gene com
uma função vantajosa. Por outro lado, também é possível que nenhuma
mudança ocorra (caso seja afetada uma região não codificante ou caso a
mudança não cause alteração da seqüência de aminoácidos codificada - devido
à degeneração do código genético).
A síntese de proteínas envolve mapear regiões específicas do DNA (os
genes) para sequências de nucleotídeos de natureza diferente (possuem uma
ribose no lugar da desoxirribose e uma nova base chamada uracila no lugar
da timina), que constituem o RNA (transcrição do DNA). Os transcritos
que orientam a síntese de proteínas são os RNAm (RNA mensageiros), os
que carregam aminoácidos são os RNAt (RNA transportadores) e os que
compõem os ribossomos são os RNAr (RNA ribossômicos).
Em seres eucariontes, ainda ocorre a excisão (ou splicing) do RNA recémtranscrito, no qual são algumas partes do RNA são removidas e degradadas
(os íntrons) e as partes restantes (os éxons) são reunidas e formam o RNA
maduro que irá para o citoplasma. Esse processo permite que vários tipos
de RNA (e, consequentemente, várias proteínas) sejam formados(as) a partir
do mesmo gene (splicing alternativo).
Cada cadeia de RNA é lida (pelos ribossomos) uma trinca de nucleotídeos
(códons) por vez, sendo que cada uma delas codifica (indiretamente) um
aminoácido (tradução do RNA). Por outro lado, um aminoácido pode ser
codificado por mais de um códon (degeneração do código genético), e alguns códons não codificam aminoácidos (e sim o início ou fim da tradução).
Quando um ribossomo encontra um códon de parada, tanto ele quanto a
proteína se desacoplam na extremidade 3’ do RNAm.
A molécula que de fato carrega um aminoácido e reconhece um determinado códon do RNAm (através da complementaridade de bases com seu
anticódon) é o RNAt (que se conecta a um dos sítios do ribossomo, adicionando o aminoácido que carrega no final da cadeia polipeptídica).
Algo surpreendente sobre o RNA é que alguns tipos de moléculas são
capazes de atuar como catalisadores: moléculas de RNAr do protozoário
Tetrahymena (por exemplo) são capazes de catalisar o seu próprio splicing
(além disso, os íntrons liberados também podem catalisar splicings de outros
RNAs). Outro exemplo é o RNAr, que catalisa a formação de novas ligações
peptídicas durante a tradução do RNAm.
5
2.2
Estrutura proteica
A maioria das cadeias polipeptídicas dobra-se em uma única disposição determinada pela sua seqüência de aminoácidos, devido às ligações não-covalentes
que a cadeia forma com si mesma e com a água (através de ligações de
hidrogênio). Outro tipo de ligação estabilizante da estrutura é a dissulfureto
(S-S), que ocorre quando dois grupos -SH de cisteínas vizinhas sofrem oxidação. Cada um dos aminoácidos que constitui uma proteína possui uma
cadeia lateral (polar ou apolar) que contribui para a sua estrutura tridimensional: em meio aquoso, as cadeias polares tendem a ficar viradas para
fora (devido à afinidade com a água, que também é polar) e as apolares
(hidrofóbicas) ficam viradas para dentro.
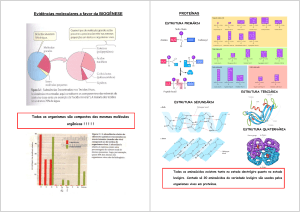
Proteínas possuem diferentes níveis de organização, que são as estruturas
primárias, secundárias, terciárias e quaternárias.
A estrutura primária é a sequência de aminoácidos ao longo da cadeia
polipeptídica, que é específica para cada proteína.
A estrutura secundária descreve as estruturas espaciais regulares formadas por segmentos da cadeia, e é constituída de hélices-alfa e folhas-beta.
Uma folha-beta é um padrão no qual regiões vizinhas da cadeia polipeptídica associam-se por meio de ligações de hidrogênio, resultando em uma
estrutura achatada e rígida. O núcleo da maioria das proteínas globulares
contém extensas regiões formadas por folhas-beta.
Já uma hélice-alfa é gerada quando uma única cadeia polipeptídica se
contorce em torno de si própria, formando uma “escada” em espiral. A estabilização se dá pela presença das ligações de hidrogênio entre os grupamentos
NH e CO da cadeia principal. Em ambientes aquosos uma alfa-hélice normalmente não é estável por si própria, e se entrelaça ao redor de outras (como
tranças de uma corda) formando uma estrutura chamada "hélice superenrolada".
A estrutura terciária descreve o dobramento final da cadeia por interação
de regiões com estrutura regular ou sem estrutura definida (é neste nível que
segmentos distantes da estrutura primária podem aproximar-se e interagir).
Aqui podem distinguir-se subunidades denominadas domínios: cada domínio
tem uma organização espacial compacta, com o interior hidrofóbico e a superfície polar (geralmente, cadeias com mais de 200 aminoácidos se dobram
em dois ou mais domínios).
Um domínio pode ser visto como a unidade básica estrutural de uma proteína, com núcleo composto de hélices-alfa e/ou folhas-beta interconectadas.
Como existem formas limitadas de combiná-las, certas combinações (motifs)
ocorrem repetidamente em muitas proteínas não-relacionadas.
A estrutura quaternária descreve a associação de duas ou mais cadeias
6
para compor uma proteína funcional. Ela é mantida por ligações não-covalentes
entre as subunidades, dos mesmos tipos que mantêm a estrutura terciária.
Apesar do número (téorico) relativamente alto de proteínas de n aminoácidos (20n ), apenas uma pequena fração possui uma estrutura tridimensional
estável (a maioria possui várias conformações de enrgia semelhante e diferentes propriedades químicas). As proteínas atuais apenas são viáveis justamente por causa de sua estabilidade e propriedades de dobramento únicas.
Assim, como algumas confomrações são recorrentes, é útil comparar proteínas em diversos níveis: muitos anos de experimentação podem ser poupados
ser uma dada sequência de aminoácidos for similar à uma sequência cuja
estrutura ou função forem conhecidas.
Outra característica das proteínas é que algumas possuem um sítio de
ligação compatível com alguma parte de sua própria superfície, formando
vários tipos de compelxos. As vantagens de se formar estruturas maiores a
partir de subunidades residem na redução do material genético necessário,
no controle facilitado da montagem e desmontagem dessas estruturas (pois
as ligações entre as subunidades são de baixa energia) e no menor índice de
erros na sua síntese (pois subunidades mal formadas podem ser descartadas).
Muitas estruturas em células são capazes de de automontagem (se os
componentes individuais forem colocados - sob condições apropriadas - num
tubo de ensaio, são capazes de formar a estrutura original). Isso acontece
com o vírus do mosaico do tabaco (uma molécula de RNA envolta por um
complexo proteico cilíndrico) e com ribossomos bacteriais. Por outro lado,
alguns processos de montagem são irreversíveis (como alguns vírus bacteriais)
e portanto as estruturas finais não consegume se formar a partir de suas
subunidades.
2.3
Proteínas como catalisadores
As propriedades químicas de uma proteína dependem da partes que ficam
expostas superficialmente, interagindo com outras moléculas através de ligações não-covalentes. As outras moléculas são denominadas ligantes (se a
proteína for uma enzima, o ligante é chamado de substrato), e a região que
se associa ao ligante é o sítio de ligação. A principal importância das partes
internas da proteína é manter o formato e a rigidez da parte externa.
O primeiro passo na catálise enzimática é a ligação do substrato do sítio
de ligação. Em seguida, a enzima orienta os átomos apropriados do modo
correto para a reação que está prestes a ocorrer. As moléculas de substrato
passam por várias formas intermediárias instáveis (estados de transição) antes
de chegar ao produto final da reação, e é justamente por essas formas que a
enzima possui maior afinidade (diminuindo drasticamente a energia de ati7
vação). Apesar disso, é importante lembrar que a enzima catalisa tanto a
reação no sentido de formação de produto quanto a reação inversa, e portanto o efeito da enzima é fazer com que o sistema chegue ao equilíbrio mais
rapidamente (a velocidade da reação em um sentido ou no outro depende entre outros fatores - da concentração dos substratos e dos produtos).
Alguns dos meios utilizados pela célula para acelerar a velocidade de formação de produtos envolve a montagem de complexos multienzimáticos (estruturas que reúnem enzimas que participam de uma mesma via metabólica)
e formação de compartimentos membranosos (em eucarióticos) ao redor de
moléculas de substrato (aumentando a sua concentração, o que favorece a
formação da reação direta).
8