")
UNIVERSIDADE FEDERAL DE LAVRAS
DEPARTAMENTO DE BIOLOGIA
SETOR DE FISIOLOGIA VEGETAL
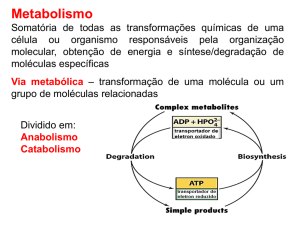
RESPIRAÇÃO
Amauri Alves de Alvarenga
Lavras – MG
2004
RESPIRAÇÃO
AMAURI ALVES DE ALVARENGA
A fotossíntese fornece substratos orgânicos básicos dos quais dependem
as plantas. Com seu metabolismo do C associado, a respiração libera, de
forma controlada, a energia armazenada nos compostos de carbono para o
uso celular.
A respiração aeróbica, comum a quase todos os organismos eucarióticos,
é um processo biológico pelo qual os compostos orgânicos reduzidos são
mobilizados e subseqüentemente oxidados de maneira controlada. A energia
livre produzida é transitoriamente armazenada na forma de ATP, podendo ser
prontamente utilizada nos processos celulares vitais e no desenvolvimento da
planta.
Os carboidratos produzidos pelas plantas durante a fotossíntese,
constituem a principal fonte de energia para os organismos vivos. Embora os
lipídeos, ácidos orgânicos e ocasionalmente as proteínas possam servir como
substrato respiratório, praticamente toda a energia produzida através do
catabolismo advém dos carboidratos. Do ponto de vista químico, a respiração
vegetal pode ser expressa como sendo a oxidação da molécula de sacarose
(12 átomos de C) e a redução de 12 moléculas de O2, resultando na seguinte
reação líquida:
C12H22O11 + 12O2
12CO2 + 11H2O
A completa oxidação de uma molécula de sacarose rende uma energia
livre correspondente a 1.380 kcal, a qual é utilizada em no metabolismo celular
para a biossíntese de inúmeras moléculas essenciais ao processo do
desenvolvimento e a produção vegetal.
Esta expressão simplificada da respiração aeróbica representa uma
seqüência de inúmeras reações metabólicas, em quatro etapas distintas e
agrupadas: glicólise, ciclo dos ácidos tricarboxílicos (ciclo do ácido cítrico ou
ciclo de Krebs), rota das pentoses fosfato e fosforilação oxidativa.
1. Glicólise
De ocorrência citoplasmática, a glicólise tem por definição clássica a
conversão de uma molécula de glicose em duas de piruvato através de uma
seqüência de 10 reações que podem ocorrer tanto na ausência quanto na
presença de O2 (figura 3.1), porém, sem sua participação.
Durante a etapa glicolítica, substratos de diferentes origens são
canalizados para triose fosfato. Cada molécula de sacarose (C12H22O11)
metabolizada no citosol produz quatro moléculas de triose-fosfato. È
importante observar que a glicólise inicia-se pela fosforilação de hexoses
(glicose e frutose) provenientes da hidrólise da sacarose. Os produtos da
fotossíntese tais como hexose e triose-fosfato também constituem substratos
respiratórios que abastecem a glicólise. Os amiloplastos, a partir da hidrólise
do amido, fornecem glicose que a ser fosforilada (G-6-P), deixa a organela,
indo para o citoplasma para ser oxidada, inicialmente pela via glicolítica.
CITOSOL
PLASTÍDEO
Amido
GLICÓLISE
Sacarose
UDP
Invertase
PPi
AT
P
Hexoquinas
e
AD
P
Glicose
ADP
Glicose
quinase
Fosfoglico
-mutase
Hexose fosfato
isomerase
Fosfofrutoquinas
e
PPi-dependente
Fosfoglico
-mutase
Glicose-1-P
Frutose-6-P
Glicose-6-P
Hexose
fosfato
H2O
AT
P
UDP-Glicose
pirofosforilas
e
UT
P
AT
P
Hexoquinas
e
AD
P
Pi
Glicose-1-P
UDP-Glicose
Frutose
Glicose
Amilase
Amido
fosforilase
Sacarose
sintase
Glicose-6-P
Hexose fosfato
isomerase
PPi
AT
P
Pi
Glicose-6-P
AMILOPLASTOS
Fosfofrutoquinase
ATP-dependente
AD
P
Frutose-1,6-bifosfato
Aldolase
Trioses
fosfato
Gliceraldeido3-fosfato
Triose fosfato
isomerase
Diidroxiacetona
fosfato
Trioses fosfato
CLOROPLASTOS
Gliceraldeído-3fosfato
desidrogenase
Pi
Gliceraldeido3-fosfato
1,3-Bifosfoglicerato
ADP
NAD+
NADH
ATP
Fosfoglicerato
quinase
3-Fosfoglicerato
Fosfoglicerato
mutase
2-Fosfoglicerato
Enolase
H2O
PEP
carboxilase
HCO3-
Fosfoenolpiruvato
Oxaloacetato
ADP
Piruvato
quinase
ATP
Pi
NAD+
Piruvato
Malato
CO2
Piruvato
descarboxilase
Acetaldeído
NADH
NAD+
Etanol
Álcool
desidrogenase
NADH
NADH
Lactato
desidrogenase
NAD+
Lactato
Para a
MITOCÔNDRIA
Reações de
fermentação
Figura 3.1: Reações da glicólise e via fermentativa em plantas superiores
(adaptado de Taiz & Zeiger, 2004).
Na fase de conversão de energia da glicólise, a triose fosfato é
convertida a piruvato. A forma oxidada do NAD+ é reduzida a NADH pela
gliceraldeido-3-fosfato desidrogenase. O ATP é sintetizado nas reações
catalisadas pela fosfoglicerato quinase e piruvato quinase. Um produto
final alternativo, fosfoenolpiruvato, pode ser convertido a malato para
oxidação mitocondrial; podendo o NADH ser reoxidado durante a
fermentação ou pela lactato desidrogenase ou pela álcool desidrogenase.
Embora seja uma das funções da glicólise gerar ATP, a primeira fase
representa uma fase de gasto.
Nos animais, a glicose é o principal substrato da respiração, enquanto
na maioria das plantas, é a sacarose. Nos vegetais, por ação da sacarose
sintase no citosol, a sacarose na presença de UDP é convertida em frutose
e UDP-glicose. Na primeira fase da glicólise, por ação da UDP-glico
pirofosforilase, a UDP-glicose + 2Pi se transforma em glicose-6-fosfato. Em
alguns tecidos, a sacarose por ação da invertase citosólica é hidrolisada
em hexoses (frutose e glicose). Nesta fase, a glicose é fosforilada pela
enzima hexoquinase, produzindo glicose-6 fosfato, que em seguida vai a
frutose-6-fosfato por ação de uma isomerase hexose-fosfato. A geração de
frutose 1,6 bifosfato se dará pela ação da fosfofrutoquinase dependente de
ATP ou por quinase dependente de PPi. Aqui reside um ponto de controle
da glicólise. Da frutose 1,6 bifosfato, há formação de duas moléculas com
3 átomos de carbono cada uma (triose-fosfato), a dihidroxiacetona fosfato
e gliceraldeído 3-fosfato. Nota-se então que nesta fase são gastas duas
moléculas de ATP.
Na segunda fase, ocorre a produção de energia que é conservada
através da fosforilação de quatro moléculas de ADP a ATP, por ocasião da
conversão das duas moléculas de gliceraldeído-3-fosfato em duas
moléculas de ácido pirúvico. Nesta fase, ocorre também a redução de
duas moléculas de NAD+ a NADH.
A expressão abaixo representa a equação geral da glicólise:
Glicose + 2NAD+ + 2 ADP + 2Pi
2Piruvato + 2NADH + 2ATP + 2H2O
Embora 4 moléculas de ATP sejam formadas na segunda fase da
glicólise, o rendimento líquido final é de apenas 2 moléculas de ATP por
molécula de glicose degradada, uma vez que duas moléculas de ATP foram
gastas na primeira fase. Sob condições aeróbicas, as duas moléculas de
NADH são reoxidadas na membrana interna da mitocondria produzindo 4
moléculas de ATP. Desse modo, sob condições aeróbicas a produção líquida
de ATP por molécula de glicose degradada é igual a 6. A figura 3.1
apresentada anteriormente mostra o rendimento energético na via glicolítica.
Respiração anaeróbica
Na ausência de oxigênio, o ciclo de Krebs e a fosforilação oxidativa não
operam, logo, a glicólise não continua a operar pelo fato do suprimento de
NAD ser limitado, ficando todo ele na forma reduzida (NADH). Para superar tal
problema, plantas e outros organismos prosseguem o metabolismo do piruvato
utilizando as rotas fermentativas.
Desta maneira, sob condições anaeróbicas, o produto final da
respiração é o etanol ou lactato (Figura 3.1).
O termo fermentação anaeróbica significa degradação anaeróbica da
glicose.
Na fermentação alcoólica, mais comum em plantas (mais conhecida
como fermento de cervejaria) as enzimas piruvato descarboxilase álcool
desidrogenase atuam sobre o piruvato produzindo ao final, etanol e CO2 e
oxidando o NADH a NAD+. Na fermentação do ácido láctico (comum em
músculos de mamíferos), mas encontrada também em plantas, a lactato
desidrogenase utiliza o NADH para reduzir o piruvato a lactato, regenerando
assim, o NAD+. Esta situação é comum quando plantas são submetidas a
condições de solos alagados com pouco ou nenhum O 2, levando as células
radiculares ao metabolismo fermentativo. No caso do milho, por exemplo, sob
baixas concentrações de oxigênio no meio de cultivo, é comum acumular ácido
láctico no início e, posteriormente, etanol, que possivelmente seje menos
fitotóxico que o lactato que acumula no citosol, acidificando-o.
Do ponto de vista de rendimento energético, pode-se dizer que a
fermentação é um processo de baixo rendimento energético na forma de ATP.
Para se ter o mesmo rendimento energético por mol de glicose oxidada em
relação à respiração aeróbica, serão necessárias 18 moléculas de glicose.
O balanço da glicólise em condições anaeróbica passa a ser então:
2 lactato + 2ATP + 2H2O + 2 H+
Glicose + 2ADP + 2Pi
Nota-se então que a redução de uma molécula de glicose gera energia
suficiente para síntese de 2 moléculas de ATP (4 moléculas a menos que em
condições aeróbicas). Veja a figura 3.1 apresentada anteriormente.
1. Rota das Pentoses Fosfato
A partir dos anos 50 ficou estabelecido que os tecidos vegetais
utilizam a via oxidativa das pentoses fosfato para a oxidação de carboidratos
no citosol (Figura 3.2). Nas células vegetais, a glicólise não é a única rota para
a oxidação de açúcares. Partilhando metabólitos comuns, a rota oxidativa das
pentoses fosfato pode realizar com eficiência tal tarefa (figura 3.2). As reações
são catalizadas por enzimas solúveis presentes nos plastídeos e citosol.
Geralmente, a via plastídica predomina em relação a via citosólica.
As duas primeiras reações representam um evento oxidativo onde a
glicose 6-fosfato (6 carbonos) é convertida em ribulose 5-fosfato (5 carbonos)
com a liberação de CO2 e formação de NADPH. A partir da ribulose são
produzidos pentoses, hexoses (glicose P e frutose P) que poderão ser
utilizadas na via glicolítica.
Estudos com
14C-glucose
indicam que a glicólise representa uma via
predominante, contabilizando de 80-95% do fluxo total de C na maioria dos
tecidos vegetais. Em tecidos já diferenciados, a rota da RPF aumenta
normalmente em relação aos tecidos meristemáticos. Em plantas atacadas por
doenças tem sido observado um aumento na degradação de glicose via RPF.
Essa rota desempenha diversos papéis no metabolismo vegetal, tais como a
produção de poder redutor extramitocondrial e extracloroplastídeo, pois aquele
produzido pelas reações fotoquímicas da fotossíntese é insuficiente para cobrir
a demanda celular. O NADPH pode ser mobilizado para as vias de biossíntese
de lipídios, esteróides, aminoácidos e componentes da parede celular. A
redução de nitrato, especialmente em tecidos aclorofilados como raízes é
dependente de NADPH. Parte da força redutora dessa rota pode ainda
contribuir para a redução do O2, gerando ATP.
A ribose produzida na RPF participa da biossíntese de nucleotídeos
(ácidos nucléicos), o mesmo acorrendo com a eritrose-4-P, importante
composto para a síntese de compostos fenólicos, aminoácidos aromáticos,
lignina, flavonóides, fitoalexinas e reguladores de crescimento. A RPF pode
ser considerada também como um atalho alternativo para a glicólise.
Tem sido constatado também que plantas deficientes em boro
apresentam um aumento na degradação de açúcar via RPF.
Acredita-se que nos estádios iniciais do enverdecimento da plântula,
fase ainda heterotrófica, a rota oxidativa das pentoses fosfato esteja envolvida
em gerar intermediários do ciclo de Calvin.
Ribulose – 5 – fosfato
Pentose fosfato
isomerase
Pentose fosfato
epimerase
Ribose-5-fosfato Xilulose- 5-fosfato
Glicose-6-fosfato
NADP+
Glicose-6fosfatodesidrogenase
Trancetolase
NADPH
6-Fosfogluconato
NADP+ Gliceraldeído-3-fosfato Sedo- heptulose-7Gluconato-6fosfatodesidrogenase
fosfato
NADP
H
Trancetolase
Ribulose-5-fosfato
Frutose-6-fosfato Eritrose- 4-fosfato
Hexose fosfato
isomerase
Xilulose-5-fosfato
Trancetolase
Gliceraldeído-3-fosfato
Figura 3.2: Reações da rota das pentoses fosfato em plantas superiores
(adaptado de Taiz & Zeiger, 2004).
2. Ciclo de Krebs
O ciclo de Krebs, também conhecido como ciclo do ácido cítrico ou
ciclo dos ácidos tricarboxílicos ocorre integralmente na mitocondria sob
condições aeróbicas. O nome ciclo de Krebs foi dado em homenagem ao
bioquímico inglês Hans A. Krebs, ao postular em 1937, a via de oxidação do
piruvato em músculo do peito de pomba. Posteriormente à sua descoberta, o
ciclo do ácido cítrico tem sido observado em todos os tecidos de animais,
plantas e microorganismo aeróbicos (Figura 3.3).
A semelhança dos cloroplastos, as mitocôndrias são organelas semiautônomas constituídas por ribossomos, RNA e DNA que permitem a síntese
de um certo número de enzimas e a transmissão de informações genéticas. As
mitocôndrias se proliferam por divisões do tipo fissão e não por biogênese de
novo. O número por células varia de acordo com a atividade do tecido,
refletindo seu papel no metabolismo energético.
O ciclo de Krebs constitui o segundo estádio da respiração (oxidação
da glicose), iniciando com a entrada do piruvato citosólico no interior da
mitocôndria através de uma proteína transportadora específica. Na matriz
mitocondrial por ação de um complexo piruvato desidrogenase, em três
etapas, o piruvato é descarboxilado, gerando CO2, NADH e acetil-CoA.
Obseva-se que o ciclo de Krebs funciona de maneira circular onde uma volta
no ciclo, inicia-se pela doação do grupo acetil do acetil-CoA ao composto de 4
carbonos (oxalacetato) que por ação da citrato sintase, resulta na formação
de citrato, um composto de 6 carbonos.
Após 9 passos seqüenciais catalizados enzimaticamente, regenerase o oxalacetato, que por sua vez está pronto para reagir com outro grupo
acetil da molécula de acetil-CoA e iniciar uma nova volta no ciclo.
Conclui-se então que não ocorre consumo líquido de oxalacetato.
Teoricamente apenas uma molécula desse ácido é suficiente para a oxidação
de um número infinito de piruvato. O piruvato citosólico pode ser regenerado
via ciclo de Krebs, a partir do malato na presença de malato desidrogenase,
via descarboxilação, produzindo também poder redutor (NADH). Uma
característica do ciclo dos ácidos tricarboxílicos nas plantas, às vezes ausente
em outros organismos, é a atividade significativa da enzima málica
desidrogenase dependente de NAD+.
Considerando-se que duas moléculas de piruvato são produzidas por
molécula de glicose, a equação geral do ciclo de Krebs fica assim
esquematizada:
2 Piruvato + 8 NAD + 2ADP + 2FAD
6CO2 + 8 NADH + 2 FADH2 + 2 ATP
Piruvato
CO2
NADH
Enzima
málica
NAD+
CoA
NAD+
A enzima málica
descarboxila o malato a
piruvato, permitindo as
mitocôndrias vegetais
oxidarem o malato.
NADH
Piruvato
Desidrogenase
CO2
Acetil-CoA
Malato
desidrogenase
Oxalacetato
NADH
Citrato
sintase
CoA
Citrato
Aconitase
NAD+
Isocitrato
Malato
NAD+
NADH
Ciclo do
Ácido Cítrico
Fumarase
H2O
Isocitrato
desidrogenase
CO2
Fumarato
2-Oxoglutarato
FADH2
Succinato
desidrogenase
Uma molécula de ATP é
sintetizada por uma reação
fosforilativa ao nível de
substrato, durante a reação
catalisada pela succinil-CoA
sintetase.
FAD
Succinat
o
CoA
NAD+
CoA
NADH
Succinil-CoA
ATP
ADP
H2O
CO2
2-Oxoglutarato
desidrogenase
Succinil-CoA
sintetase
Figura 3.3: Ciclo dos Ácidos Tricarboxílicos, ou de Krebs (adaptado de Taiz e
Zeiger, 2004).
3. Rendimento Energético da Respiração
Ao final do ciclo de Krebs, pode-se calcular o número de ATP
formado quando uma molécula de glicose é oxidada à CO2 via glicólise e ciclo
de Krebs (Figura 3.4).
Figura 3.4: Rendimento energético da respiração aeróbica (Buchanan, 2000).
Na glicólise há um consumo inicial de 2 ATP e uma produção
posterior de 4 ATP e 2 NADH, dando um rendimento líquido de 6 ATP. No
ciclo de Krebs são produzidos mais 30 ATP. Conclui-se assim que na
respiração aeróbica há produção de 36 ATP, equivalente a 288 Kcal (36X 8
Kcal), dando um rendimento energético de aproximadamente 42% (288 Kcal /
686 Kcal), sendo o restante da energia perdida na forma de calor. Por outro
lado, na respiração anaeróbica o rendimento energético é muito baixo, ou seja,
cerca de 2% (2 ATP X 8 Kcal / 686 Kcal).
4. Cadeia de Transporte de Elétrons (CTE)
Como foi visto anteriormente, para cada molécula de glicose oxidada
envolvendo a glicólise e ciclo de Krebs, são geradas no citoplasma 2
moléculas de NADH e 8 moléculas de NADH e 2 moléculas de FADH 2 na
mitocôndria.
Esses compostos, ao serem reoxidados repassam seus elétrons ao
oxigênio que é o aceptor final no processo via cadeia de transporte de elétrons
(CTE). Veja a figura 3.5.
A CTE consiste de uma série de proteínas transportadoras (Figura
3.5) que recebem e doam elétrons, através de reações de óxido-redução.
Neste sistema, cada componente da cadeia recebe elétrons do componente
precedente, transferindo-os para o seguinte em uma seqüência específica até
o O2. Os elétrons, ao fluírem pela CTE perdem parte de sua energia que é
conservada na forma de ATP, por meio da fosforilação oxidativa, onde há
participação do O2.
Figura 3.5: Cadeia transportadora de elétrons mitocondrial
Nota-se por esta figura que existem 3 sítios de formação de ATP. Os
pares de elétrons doados pelo NADH mitocondrial passam por todos esses
sítios conservadores de energia formando um máximo de 3 ATP. Por outro
lado, os elétrons doados pelo NADH (citoplasmático) e FADH 2 entram na CTE
ao nível da ubiquinona, ou seja, um sítio adiante do primeiro, levando
portanto, a formação de 2 ATP.
5. Cociente Respiratório (CR)
Denomina-se cociente respiratório a relação entre o CO 2 liberado e o O2
consumido durante o processo respiratório, caracterizado ou identificado pela
expressão CR = CO2 liberado / O2 consumido. A principal fonte de reserva de
tecidos ou órgãos como sementes, por exemplo, pode ser identificada pelo seu
CR. Veja os exemplos a seguir. O CR de cereais como milho, arroz, aveia e
trigo é aproximadamente igual a 1 (um), indicando que o AMIDO é a principal
reserva encontrada nas sementes; em sementes de oleaginosas como
girassol, algodão, amendoin por serem ricas em lipídeos, o seu CR é menor ou
igual a 0,7. Entretanto, órgãos vegetais com CR superior a 1 (um) é indicativo
de oxidação predominante de ácidos orgânicos ou de oxidação por via
fermentativa, onde o consumo de O2 é muito baixo, ou quase nulo. Vejamos
alguns exemplos.
(A) C6H12O6 + 6O2
6CO2 + 6H2O + x ATP (CR=1)
(glicose)
(B) C18H36O2 + 26O2
18CO2 +18H2O + x ATP (CR=0,69)
(Ácido oléico)
(C) C4H4O5 + 2,5O2
4CO2 + 2H2O (CR=1,6)
(Ácido oxalacético)
Medindo-se o CR de uma semente ou de um outro órgão da planta, pode
ser obtida a informação de qual o principal composto de reserva está sendo
oxidado. A identificação do substrato que está sendo oxidado não é tão
simples, pois vários tipos de compostos podem ser oxidados ao mesmo
tempo. Portanto, o CR é tão somente um valor médio dependente da
contribuição de cada substrato e de seu conteúdo relativo de C, H,e O.
6. Produção de Compostos Intermediários na respiração de Plantas
A primeira vista, pode-se imaginar que a respiração tem como única
função à produção de energia. Na realidade é um processo complexo que
envolve uma série de reações que não só levam a produção de energia, mas,
também a compostos intermediários imprescindíveis para a produção de
aminoácidos, pigmentos fotossintéticos, celulose, fitohormônios, alcalóides,
ácidos graxos, ácidos nucléicos e inúmeras outras biomoléculas essenciais às
células (Figura 3.6). Em casos das células apresentarem um suprimento
adequado de ATP, os substratos respiratórios serão completamente oxidados,
gerando esqueletos de carbono para a biossíntese de outras moléculas.
Outro aspecto importante é o fato da remoção de ácidos orgânicos
do ciclo de Krebs prevenir a regeneração do oxalacetato. Neste caso, o ciclo é
interrompido ou diminui sua velocidade até que haja, por exemplo, outros
mecanismos capazes de regenerar estes importantes ácidos orgânicos. A
esses mecanismos enzimáticos especiais pelos quais os intermediários do
ciclo podem ser repostos, denominam-se “Reações anapleróticas”.
Amido
ADP
ATP
NAD
NADP
FMN
CoA
Citocininas
Ácidos Nucléicos
Glicose-1-fosfato
Nucleotideo
s
Ácido Indolacético
(auxina)
Alcalóides
Flavonóides
Lignina
Triptofano
Tirosina
Fenilalanin
a
Proteínas
Pentose
fosfato
Celulose
Glicose-6-fosfato
Eritrose-4-fosfato
Gliceraldeido-3-fosfato
Ácido chiquímico
Fosfoenolpiruvat
o
Alanin
a
Glicerol-3-fosfato
Lipídeos e
substâncias
relacionadas
Piruvato
Acetil-CoA
Aspartato
Oxaloacetato
Malato
Ácidos graxos
Citrato
Ciclo do
Ácido
Cítrico
Diidroxiacetona
fosfato
Giberelinas
Carotenóides
Esteróis
Ácido abscísico
Isocitrato
Proteínas
Fumarat
o
2 - Oxoglutarato
Succinat
o
Clorofilas
Ficocianinas
Fitocromo
Citocromo
Catalase
Glutamato
Outros
aminoácidos
Figura 3.6: Produção de compostos intermediários na respiração de plantas
(adaptado de Taiz & Zeiger, 2004).
7. Respiração em plantas e órgãos vegetais
A grande maioria dos estudos de respiração foram desenvolvidos em
organelas isoladas e em extratos livres de células. O questionamento que se
faz é, como o processo respiratório se realiza em nível de planta ou em
ecossistemas naturais e agrícolas.
As plantas de uma maneira geral, respiram aproximadamente a metade da
produção fotossintética diária. Fatores relevantes incluem a espécie, hábito de
crescimento da planta, tipo e idade do órgão, além de variáveis ambientais,
como concentração de oxigênio, temperatura, suprimento de nutrientes e
água.
Estudos em plantas herbáceas revelam que 30 a 60% do ganho diário de
carbono na fotossíntese é perdido pela respiração. Em espécies tropicais,
essa perda de carbono pela respiração pode chegar a valores na ordem de 70
a 80%, devido a temperaturas noturnas elevadas.
De maneira geral, quanto maior for a atividade metabólica de um tecido,
maior sua taxa respiratória. Gemas em desenvolvimento, por exemplo,
mostram elevadas taxas de respiratórias por unidade de matéria seca. Em
regiões mais distantes dos meristemas essa taxa tende a decrescer
significativamente. Tecidos vegetativos maduros e em caules geralmente são
os que apresentam as menores taxas de respiração. Normalmente a atividade
respiratória é mais intensa nas regiões de crescimento ativo, pois, além da
demanda considerada normal de energia metabólica, há aquela demanda
adicional para formação dos mais diversos componentes celulares. O principal
substrato utilizado no processo respiratório é a sacarose, um dissacarídeo
oriundo da fotossíntese e transportado via floema para as mais diversas
regiões de consumo da planta.
7.1 Respiração de Frutos
O desenvolvimento dos frutos encontra-se nas dependência dos
carboidratos produzidos pela fotossíntese nas folhas vizinhas e dos nutrientes
minerais absorvidos pelas raízes e translocados das folhas. Logo após a
fertilização, há uma intensa divisão celular acompanhada de forte atividade
respiratória que declina nas fases subseqüentes do desenvolvimento do fruto
até a sua senescência.
Certos frutos como banana, abacate, mamão, maçã, dentre outros,
apresentam um aumento na atividade respiratória e depois um decréscimo, no
fim da fase de maturação. A esse fenômeno, denomina-se de “Climatério”,
fase essa antecedida por uma maior produção de Etileno. Parece que na
maioria dos frutos, há pelo menos um pequeno aumento no teor de etileno,
antes de se iniciar o aumento na respiração. Nestes frutos, acredita-se que o
etileno seja a causa do climatério, este se antecipa e ocorre a “Maturação
Induzida” (Figura 3.7). Nesses frutos, a aplicação exógena de substâncias que
liberam etileno como o Ethrel (ácido 2-cloro-etilfosfônico) promove o mesmo
tipo de efeito, fazendo com a maturação seja mais precoce. Os frutos não
climatérios como os citrus, exibem padrões diferentes de respiração.
O metabolismo do etileno, atualmente é bem conhecido, especialmente a
sua biossíntese, o que vem a facilitar a compreensão do seu modo de ação no
processo de maturação dos frutos climatérios, muito embora, estejam
envolvidos outros fitohormônios como ácido indolacético (AIA), giberelinas e
ácido abscísico. Isto tem permitido o desenvolvimento de metodologias para o
controle da maturação desses frutos, como o uso de inibidores da síntese de
etileno Sabe-se que estes fitohormônios podem estar ligados direta ou
indiretamente com a produção de enzimas hidrolíticas e respiratórias. Durante
o processo de maturação dos frutos, ocorre uma diminuição dos ácidos
orgânicos e um acúmulo de açúcares livres (hexoses e sacaroses) devido,
neste último caso, à hidrólise de amido, pectinas e outros polissacarídeos.
Com o conhecimento dos fatores que afetam o climatério, pode-se retardar ou
acelerar o amadurecimento do fruto. Os frutos climatérios estão na fase ótima
para serem consumidos, próximo ao climatério.
Figura 3.7: Produção de etileno e respiração: amadurecimento de banana,
sugerindo a participação do etileno.
7.2 Respiração em Raízes
Nas raízes, a respiração é intensa, sendo a energia produzida
empregada na formação de novas raízes e no crescimento daquelas já
existentes. A absorção e o acúmulo de íons minerais no sistema radicular são
processos altamente dependentes de energia metabólica fornecida pelo
processo respiratório. Neste contexto, há de ressaltar a importância da rota
das pentoses monofosfato (RPF) nas raízes como geradora de poder redutor
(NADPH) utilizado no processo de redução do nitrato a nitrito. O estudo
respiratório merece uma atenção especial quando se trata de plantas
submetidas à condições de baixa disponibilidade de oxigênio, sobretudo, em
situações de enxarcamento e/ou inundação periódica comum em épocas de
chuva, uma vez que predominará o metabolismo respiratório fermentativo ou
anaeróbico.
7.3 Respiração nas Folhas
Nas folhas, a liberação de CO2 é mais ou menos constante, não
sofrendo alterações substanciais com a idade do tecido. Em tecidos
vegetativos maduros, a respiração varia com a espécie e as condições sob as
quais as plantas se desenvolvem. Antes que ocorra a abscisão foliar, a
respiração geralmente aumenta rapidamente, decrescendo em seguida
(senescência foliar) quando as folhas encontram-se praticamente mortas. Há
evidências de que parte do CO2 liberado seja proveniente de outros processos
metabólicos, como por exemplo, a Rota da Pentose Fosfato (RPF) e a
fotorrespiração, sobretudo, em plantas C3.
7.4 Respiração nas Sementes em Germinação
O processo de germinação de uma semente inicia-se com a
embebição de água, seguida pelo desencadeamento de uma série de
alterações
metabólicas
representadas
pela
hidrólise
de
compostos
armazenados na semente (Figura 3.8).
Os produtos de hidrólise destes compostos, sobretudo de carboidratos e
lipídeos, constituem os principais substratos para a respiração e a síntese de
componentes celulares essenciais ao desenvolvimento inicial da plântula.
Durante o processo de germinação, a massa seca da semente decresce
continuamente
devido
a
oxidação
de
compostos
ricos
em
energia
acompanhado pela liberação de CO2. Vários substratos armazenados nas
sementes podem ser utilizados na respiração como o polissacarídeo amido
(mais utilizado) e lipídeos, principalmente.
Malato
NAD+
desidrogenase
Triacilgliceróis são
hidrolisados para
produzir ácidos
graxos.
Lipase
Ácidos graxos são metabolizados
por β- oxidação a acetil-CoA no
glioxissomo.
OLEOSSOMO
Triacilgliceróis
Ácido Graxo
O malato é transportado
para o citosol e oxidado a
oxalacetato, o qual é
convertido
a
fosfoenolpiruvato
pela
enzima
PEP
carboxiquinase. O PEP
resultante
é,
então,
metabolizado
para
produzir sacarose pela
rota gliconeogênica.
Citrato
Citrato
sintase
Acil-CoA (Cn)
Acetil-CoA
n/2.O2
Aconitase
CoA
CoA
Citrato
Ciclo do
Oxaloacetato Glioxilato
NADH
Isocitrato
Β- oxidação
n. H2O
Malato
CoA
n.NAD+
n.NADH
Isocitrato
Malato
sintase
Isocitrato
liase
Glioxilato
n.FADH2
Succinato
n.FAD
O succinato move-se
para
dentro
da
mitocôndria
e
é
convertido a malato.
GLIOXISSOMO
Fosfoenolpiruvato
(PEP)
ADP
Succinato
CITOSOL
ATP
PEP
Carboxiquinase
Frutose-6-P
Malato
Fumarato
NADH
NAD+
MITOCONDRIA
Oxaloacetato
SACAROSE
Malato
desidrogenase
Malato
Figura 3.8: Mobilização de reservas durante a germinação de sementes
(adaptado de Taiz & Zeiger, 2004).
")