Bioenergética
Uma via metabólica é um conjunto de reacções em cadeia catalisadas por enzimas. O
metabolismo celular é portanto organizado por enzimas.
O metabolismo celular depende de dois tipos de reacções químicas: catabólicas e anabólicas.
As primeiras consistem na quebra de macromoléculas, sendo fonte de energia e de pequenhas
moléculas. As últimas, por sua vez, utilizam energia derivada do catabolismo para sintetizar
moléculas essenciais para a célula.
Para que as enzimas funcionem precisão de uma região activa, á qual se vai ligar uma
molécula. Essa região activa tem 2 partes, uma que reconhece e liga o substrato e outra que
cataliza a reacção após o substrato se ter ligado (a energia livre da reacção diminui). Dada a
reacção, o subtrato passa a produto, libertando-se da enzima e passando esta novamente a ter
a região activa pronta a voltar a ligar outro subtrato.
Para manter a homeostasia, cada célula tem que efectuar inúmeras reacções químicas.
Assim, pequenas moléculas orgânicas ligam-se por ligações covalentes para construir uma
grande diversidade de proteínas, ácidos nucleicos e outras macromoléculas. Conjuntos de
macromoléculas são ligados por ligações não covalentes.
A biossíntese necessita de moléculas carrier activadas pois a energia libertada pela oxidação de
moléculas energéticas tem de ser utilizada na construção de moléculas maiores e mais
complexas. Na maioria dos casos a energia é armazenada em moléculas carrier que possuem
uma ou mais ligações covalentes ricas em energia facilmente utilizável. As moléculas carrier
mais importantes são o ATP, NADH, NADPH. No Anabolismo, por as reacções serem
energeticamente desfavoráveis são activadas moléculas carrier. A formação destas moléculas
activadas está portanto acoplada a reacções energeticamente favoráveis do catabolismo. A
oxidação da glucose está acoplada a reacções enzimáticas que asseguram grande parte da
energia livre libertada pela oxidação seja capturada sob forma quimicamente utilizável, em vez
de ser perdida em calor. Este processo depende de reacções paralelas e acopladas: a reacção
energeticamente favorável fornece energia para a reacção energeticamente desfavorável a
qual produz uma molécula carrier activa ou outra forma de armazenamento de energia.
As moléculas obtêm energia através da oxidação de moléculas orgânicas. Assim, a fonte de
energia das células são as ligações químicas das mléculas orgânicas. A extracção de energia dáse por oxidação gradual. Na presença de oxigénio, a forma energeticamente mais estável do
carbono é o dióxido de carbono e do hidrogénio é a água. Assim, uma célula pode obter
energia a partir de açucares e outras moleculas orgânicas através da combinação dos átomos
de C e H com o O e produção de dióxido de carbono e água. A este processo dá-se o nome de
respiração.
A oxidação redução é outra reacção que ocorre nas células e que envolve a transferência de
electrões. Porém, o metabolismo envolve uma série de reacções enzimáticas, raramente
envolvendo a adição directa de oxigénio. A oxidação consiste na reacção em que os electrões
são transferidos de um átomo para outro (perda). Por outro lado, a redução é a reacção
inversa, consistindo no ganho ou adição de electrões. Numa reacção química o número de
electrões é conservado, ocorrendo estas reacções sempre simultaneamente.
O ATP é uma molécula versátil que funciona como armazenamento ou troca de energia para
desencadear várias reacções químicas celulares. O ATP é sintetizado pela adição de um grupo
fosfato a uma molécula de ADP (reacção desfavorável). Quando necessário, liberta a energia
armazenada através da hidrólise em ADP+Pi (reacção favorável). ADP libertado pode ser
novamente convertido em ATP. As ligações entre os grupos fosfato de ATP são ligações de alta
energia pois a sua hidrólise resulta numa diminuição de energia livre do sistema. O ATP pode
ser hidrolisado no descrito anteriormente ou em AMP e pirofosfato.
O NADH e NADPH também são moléculas carrier importantes. São carriers especializados no
armazenamento e transporte de electrões de alta energia e átomos de hidrogénio. Nas
reacções, o NAD+ e o NADP+ recebem um packet of energy correspondente a electrões de alta
energia e um protão (H+). Tal como o ATP, o NADPH participa em muitas reacções que de outro
modo seriam desfavoráveis energeticamente.
Resumindo, existem várias moléculas carrier activas que participam no metabolismo, levando
um grupo ligado numa ligação de alta energia. O ATP liga o fosfato, o NADH, NADPH e FADH2
ligam electrões e hidrogénios, o acetyl CoA liga um grupo acetil numa ligação transferível, a
uridina difosfato glucose liga a glucose, entre outros.
As células necessitam de fornecimento constante de energia para viver. A energia provém de
energia química de ligações das moléculas de alimento que servem de combustível para a
célula. Os açucares são moléculas energéticas muito importantes, que são oxidadas em CO2 e
H2O.
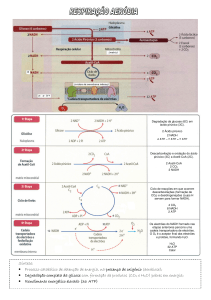
A principal etapa do catabolismo dos açucares é a glicólise. A glicólise é um conjunto de
reacções que levam à degradação da glicose até ao piruvato (no citosol) e que tem como
objectivo a obtenção de energia. Pode dividir-se este processo em 2 etapas: uma primeira em
que h+a investimento energético e que engloba as 4 primeiras reacções e a segunda etapa em
que se dá o ganho energético. Dá-se no citoplasma de células cerebrais, músculos esqueléticos
e eritrócitos.
Reacção global da glicólise:
Esquema geral da glicólise:
Na degradação completa da glicose ocorre tanto a degradação de GAP como de DHAP,
surgindo portanto como produto final 2 piruvatos. O balanço energético é a produção de 8
ATP. A 4ª reacção da glicólise é portanto a responsável pelos 2 piruvatos finais.
Na 1º reacção da glicólise há a transferência de um grupo fosfato de ATP para a glucose
originando G6P. Esta reacção é catalisada pela hexocinase. Ocorre a hidrólise espontânea do
ATP (reacção com muita tendência a ocorrer) porque o citoplasma é praticamente água.
Quando o ATP se liga ao centro activo da enzima catalisadora, há menos espaço para a água
interagir como ATP, havendo uma menor tendência para ocorrer a sua hidrólise.
Destino do produto final da glicólise (condições anaeróbias versus condições aeróbias):
O nosso organismo não sintetiza a enzima necessária á fermentação alcoólica. Porém, a
fermentação a lactato ocorre nos músculos em actividade intensa e em alguns
microorganismos. As condições anaeróbicas implicam a presença de dióxido de carbono.
Em condições aeróbias, o piruvato formado na glicólise é rapidamente descarboxilado por um
conjunto de 3 enzimas mitocondriais: complexo piruvato desidrogenase. Desta reacção formase dióxido de carbono, 1 NADH e Acetil-CoA. Estes processos já não ocorrem no citosol mas
sim na mitocôndria. A posterior degradação da acetil-CoA ocorre num ciclo, o de Krebs, ou do
ácido cítrico. Consiste em 8 reacções durante as quais se oxidam os grupos acetil provenientes
de diferentes vias metabólicas. Grande parte da energia de oxidação do piruvato é transferida
para transportadores electrónicos. Total do ciclo: 24 ATP. A passagem do piruvato a acetil-CoA
implica ainda o ganho de 6 ATP. As enzimas que degradam ácidos gordos também produzem
acetil-CoA na mitocôndria. Cada molécula de ácido gordo é degradada completamente num
ciclo de reacções. Por cada volta do ciclo produzem-se 1 acteil CoA, 1 NADH e 1 FADH2.
Esquema resumido do ciclo de Krebs:
É no último passo da degradação de uma molécula energética que a maior parte da energia
química é libertada. No processo final, os transportadores de electrões NADH e FADH2
transferem os electrões que ganharam na oxidação de outras moléculas para a cadeia
transportadora de electrões.
À medida que os electrões passam ao longo de uma cadeia de aceitadores de electrões e de
moléculas dadoras vão passando para níveis energéticos cada vez menores. Neste processo
gera-se um gradiente de H+ - fonte de energia para outras reacções, tais como a formação de
ATP pela fosforilação do ADP. No final da série transferência de electrões, os electrões passam
para moléculas de oxigénio gasoso que difundem para a mitocôndria, combinando-se
simultaneamente com o H+ e gerando moléculas de H2O.
Os electrões atingem o nível de energia mais baixo e consequentemente toda a energia
disponível foi extraída pela oxidação das moléculas energéticas- fosforilação oxidativa.
A maioria dos transportadores electrónicos encontram-se inseridos em complexos
multienzimáticos localizados na membrana interna da mtocôndria. Os electrões ganhos da
oxidação de outras moléculas são transferidos pelos transportadores para a cadeia
transportadora de electrões. À medida que os electroões passam para níveis energéticos
menores, gerando um gradiente de protões (fonte de energia para reacções como a formação
de ATP).
O complexo I é o principal ponto de entrada de electrões na cadeia respiratória mitocôndrial.
Dá-se produção de ATP. Reacção:
. O complexo II é o
2º ponto de entrada de electões. Contém, succinato desidrogenase. Tanto este complexo
como o I catalisam a transferência de NADH e succinato para a ubiquinona. Reacção de II:
.
O complexo III transfere electrões da ubiquinona para o citocromo c. Há também produção de
ATP. Reacção:
.
No complexo IV também há produção de ATP. Este complexo catalisa a transferência de
electrões do citocromo c para o oxigénio. Reacção:
.
O gradiente electroquímico resultante é utilizado para formar ATP pela ATPsintase ou
complexo V. Os protões regressam posteriormente à matriz mitocondrial dada a
impermeabilidade da membrana. O fluxo de protões provindo daí leva à síntese de ATP.
Os protões são bombados da matriz para o espaço intermembranar.
Quando o NADH (ao qual a membrana interna da mitocôndria é impermeável) é transportado
pelo malato-aspartato origina 3 ATP (células de fígado, rins e coração). Quando é utilizado o
transportador glicerol fosfato (músculos esqueléticos) origina 2 ATP.
A oxdiação do piruvato e dos ácidos gordos é acoplada à redução dos transportadores de
electrões NAD+ e FAD. A transferência de electrões até o oxigénio, ligada à geração do
gradiente de protões. Utilização da energia, armazenada no gradiente de concentração
de protões transmembranar, para a síntese de ATP.
O total da degradação total da glicose é 32 ATP (38 mas com 6 de investimento
energético). Não se conta com a afosforilação oxidativa para este balanço. Considerase o malato aspartato como transportador.