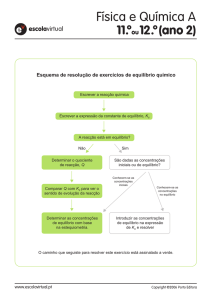
")
Faculdade de Medicina de Lisboa – Metabolismo e Endocrinologia 07/08
Catabolismo Proteico
(a) Renovação proteica – etapas e intervenientes, transportador e locais de
degradação celular;
Todas as proteínas eventualmente têm de ser degradadas, quer porque a sua
“vida” chegou ao fim, quer devido a problemas estruturais. A degradação proteica é,
assim, um passo essencial para prevenir o desenvolvimento de proteínas com anomalias
estruturais, incapazes de executar a sua função correctamente.
Dependendo da sua sinalização, as proteínas seguem um de dois caminhos
aquando da sua degradação. As proteínas extracelulares, membranares e com maior
peso molecular, ou seja, maior período de semi-vida, possuem muitas vezes grupos
açucarados (glicoproteínas) que permitem a marcação das mesmas. Aquando do seu
envelhecimento, ocorre uma marcação destas com ácido siálico, tornando-se acilo
glicoproteínas, que funciona como sinal para serem endocitadas. São então degradadas
nos lisossomas onde depois se reciclam os seus aminoácidos, num processo que não
recorre a ATP. As enzimas que tratam da degradação lisossomal são as enzimas
proteolíticas – Catepsinas –, activadas pelo pH ácido do lisossoma, e dividem-se em
endopeptidases, se reconhecem sequências de aminoácidos no interior da cadeia
proteica, e exopeptidases, se actuam nos terminais da cadeia peptídica. Estas últimas
podem ainda reconhecer e actuar exclusivamente no terminal carboxil ou amina de uma
cadeia. Este mecanismo é também utilizado quando as proteínas têm malformações que
lhes conferem grande instabilidade nas estruturas terciárias (quando analisadas
independentemente umas das outras).
Já as proteínas defeituosas ou com peso molecular médio ou baixo, isto é tempo
de semi-vida curto, são degradadas no citosol através de mecanismos dependentes de
ATP, nomeadamente o sistema Ubiquitina-Proteossoma. Este tipo de degradação, em
células eucariotas, envolve a proteína Ubiquitina e três enzimas (E1, E2 e E3), para
além do centro onde se dá a degradação, o
proteossoma. O processo de ubiquitinação dá-se no
citosol e tem que ser repetido no mínimo quatro
vezes para que o proteossoma reconheça a proteína a
ser degradada. Esta marcação da proteína divide-se
em três etapas:
1º - A enzima E1, recorrendo à energia libertada pela
hidrólise de ATP a AMP, une-se à ubiquitina através
da sua extremidade carboxilo;
2º - A enzima E2 toma o lugar da enzima E1;
3º - A enzima E3 permite que a enzima E2 unida à
ubiquitina seja substituída pela proteína a degradar,
Fig.1 - Proteossoma
estabelecendo-se uma ligação entre a extremidade
carboxilo da ubiquitina e um grupo amina de um
resíduo de lisina da proteína alvo.
Após este processo, a proteína ubiquitinada segue, finalmente, para o
Proteossoma 26S. Estes proteossomas localizam-se no citosol e são constituídos por três
subcomplexos: no centro o subcomplexo 20S e nas extremidades superior e inferior o
subcomplexo 19S (regulador). É num destes subcomplexos 19S que a proteína
ubiquitinada se liga e começa, então, a desenrolar-se por acção das cerca de 18
subunidades, seis das quais ATPases, que constituem cada complexo regulador do
Trabalho realizado por: João Carreiras (58404), Frederico Ribeiro (58449) e João Veiga (58757)
21 de Abril de 2008 Mestrado Integrado em Engenharia Biomédica
Faculdade de Medicina de Lisboa – Metabolismo e Endocrinologia 07/08
proteossoma. Por fim, a proteína já desenrolada é processada no subcomplexo 20S por
acção de proteases presentes em cada três das sete subunidades β que constituem a
unidade central do proteossoma.
No fim da sua degradação, o que era uma proteína passou a ser apenas um
conjunto de aminoácidos e péptidos mais simples que podem ser reciclados, fechando
um ciclo de renovação proteica, ou catabolisados.
(b) Catabolismo dos aminoácidos, destino dos grupos aminados, etapas,
componentes e mecanismos de transferência e transporte dos grupos aminados;
Quando uma célula degrada os seus aminoácidos usa apenas os esqueletos de
carbono para gerar energia, ou seja, tem de remover o grupo amina dos mesmos quer
por transaminção quer por desaminação. No entanto, para quase todos os aminoácidos,
as transaminações são as reacções por excelência no catabolismo destes grupos
aminados.
Debrucemo-nos um pouco mais sobre estas reacções. Elas consistem na troca de
um grupo aminado entre duas espécies químicas e são dirigidas pelas enzimas
aminotransferases que conseguem passar o grupo aminado de um aminoácido para o αcetoglutarato, resultando então glutamato e o cetoácido correspondente. Como os
substratos são os vários aminoácidos, haverá uma enzima diferente para cada
aminoácido (alanina aminotransferase, aspartato aminotransferase). No entanto, o mais
importante nestas reacções enzimáticas prende-se com a energia livre da reacção:
ΔG≈0kJ/mol, que não só garante a livre reversibilidade da reacção como também
permite a troca dos grupos funcionais, cetona e amina, quase sem dispêndio de energia.
Para que tudo isto seja possível, a peça chave de todas as aminotranferases é a coenzima
PLP (piridoxal fosfato), proveniente da vitamina B6. Esta está ligada covalentemente ao
centro activo da enzima e consegue passar reversivelmente de aldeído (piridoxal fosfato,
capaz de receber um grupo aminado) a amina (piridoxamina fosfato, capaz de doar um
grupo aminado). Trata-se portanto de um transportador intermédio como demonstra o
esquema. É de notar que graças ao anel da PLP, a estabilização por ressonância das
moléculas intermédias fica garantida, podendo os electrões circular sem que se formem
espécies intermédias demasiado reactivas.
Fig.2 – Transferência do grupo aminado pela PLP
Durante a actividade celular, formam-se grupos aminados (-NH2/-NH3+) e
amónio (NH4+) nos tecidos, por degradação dos aminoácidos. Estes grupos são
transaminados para o glutamato, que os armazena na célula. Uma vez que são tóxicos,
os grupos aminados e o ião amónio vão ser transportados para o fígado para serem
processados. Em vez de glutamato, a forma preferencial para transportar estes
compostos em meio extracelular é a glutamina. O amónio é então combinado com
glutamato para formar glutamina sob a acção da glutamina sintetase. Esta reacção
Trabalho realizado por: João Carreiras (58404), Frederico Ribeiro (58449) e João Veiga (58757)
21 de Abril de 2008 Mestrado Integrado em Engenharia Biomédica
Faculdade de Medicina de Lisboa – Metabolismo e Endocrinologia 07/08
consome ATP e ocorre em dois passos: primeiro, o glutamato e o ATP reagem para
formar ADP e γ-glutamil fosfato. Esta molécula intermediária da reacção vai então
reagir com o amónio dando finalmente a glutamina e fosfato inorgânico. O destino final
da glutamina será a mitocôndria dos hepatócitos, onde liberta os grupos aminados e o
amónio para serem processados.
No entanto, existem excepções a este mecanismo geral. Aquando do metabolismo do
músculo esquelético, os aminoácidos sofrem uma transaminação entre o glutamato e o
piruvato para formar alanina e α-cetoglutarato, por acção da alanina aminotransferase. A
alanina encaminha-se, posteriormente, para o fígado por via sanguínea. Tal como todos
os aminoácidos que chegam ao fígado, a alanina é submetida a uma nova transminação
só que agora em sentido inverso de modo a doar o grupo amina ao α-cetoglutarato
devolvendo glutamato e piruvato. Numa posterior gliconeogénese, o piruvato pode
voltar ao tecido original sob forma de glicose completando assim o ciclo da glicosealanina.
Formação do amoníaco (NH3) e ião amónio (NH4+) e ciclo da ureia - etapas e
regulação.
O ciclo da ureia compreende todas as reacções e respectivas enzimas que
intervêm na formação da ureia(NH2CONH2), uma molécula que possibilita excretar o
ião amónio através da urina. O amoníaco e o seu respectivo ião são compostos tóxicos,
que têm de ser eliminados.
Entre as diferentes fontes de amónio e amoníaco destacam-se os grupos amina
resultantes do catabolismo dos diferentes aminoácidos, mas também a formação de
amónio puro, por parte do cérebro, resultante da degradação das bases azotadas dos
nucleótidos.
O amónio/grupos aminados são, como foi anteriormente referido, transportados
até ao fígado para serem processados. A glutamina entra para a matriz mitocondrial dos
hepatócitos, sendo desaminada pela glutaminase para formar glutamato libertando assim
um NH4+. Ficamos apenas com glutamato na matriz que, sob a acção da enzima
glutamato desidrogenase, devolve α-cetoglutarato e mais um NH4+.
Em conjugação com o CO2 presente na matriz mitocondrial, o NH4+ vai originar
a molécula de carbamoíl-fosfato. Esta reacção requer a hidrólise de dois ATP a ADP e é
catalisada pela carbamoíl-fosfato sintetase I.
A partir do carbamoíl-fosfato entramos então no ciclo da ureia, via composta por
quatro passos enzimáticos. O primeiro é a reacção de transferência do grupo carbamoíl
do carbamoíl-fosfato para a ornitina, pela acção da transcarbamoílase, o que origina
citrulina. De notar que este passo ainda ocorre no interior da mitocôndria, mas logo de
seguida a citrulina é transportada para o citosol do hepatócito, onde ocorrerão os
seguintes passos do ciclo. O segundo passo é a reacção de condensação do aspartato
com a citrulina com a formação de argininosuccinato e a enzima que o catalisa é a
argininosuccinato sintetase. Esta reacção passa por um estado intermediário onde se
forma citrulil-AMP, que resulta da clivagem de um ATP a AMP que reage com a
citrulina. É importante referir que, além do NH4+ já utilizado que originou o carbamoílfosfato no início da via, o aspartato aqui utilizado como reagente proveio da reacção de
transaminação entre o oxaloacetato e o glutamato pelo que, em cada ciclo, são
eliminados dois NH4+. O terceiro passo da via é a clivagem do argininosuccinato a
fumarato e arginina, sendo que apenas a segunda permanece no ciclo. Esta reacção é
catalisada pela argininosuccinase e é a única reacção reversível do ciclo. O último
passo, catalisado pela enzima arginase, consiste na hidrólise da arginina para originar
Trabalho realizado por: João Carreiras (58404), Frederico Ribeiro (58449) e João Veiga (58757)
21 de Abril de 2008 Mestrado Integrado em Engenharia Biomédica
Faculdade de Medicina de Lisboa – Metabolismo e Endocrinologia 07/08
uma molécula de ureia e de ornitina. A ureia é então encaminha para os rins através da
corrente sanguínea enquanto que a ornitina reentra na mitocôndria onde será utilizada
para iniciar um novo ciclo.
A regulação desta via é feita essencialmente a dois níveis, que podemos
distinguir de um ponto de vista temporal. Existe uma regulação da via a longo prazo,
através de uma alteração na produção das quatro enzimas do ciclo e da carbamoílfosfato sintetase I e existe uma regulação a curto prazo, através da regulação alostérica
da carbamoíl-fosfato sintetase I. É fácil ver o porquê de ser este o passo regulador da via
já que é a etapa responsável por dar o primeiro substrato do ciclo. A actividade da
carbamoíl-fosfato sintetase I é regulada pelo N-acetilglutamato, um composto que
resulta da acetil-CoA e do glutamato. No entanto, os níveis de N-acetilglutamato são
controlados pela arginina, uma vez que esta molécula é activadora da N-acetilglutamato
sintetase, a enzima responsável pela conversão da acetil-CoA e glutamato nesta
molécula. Podemos afirmar então que a arginina é um activador do ciclo da ureia. Existe
ainda uma regulação alostérica protagonizada pela ornitina que, ao ligar-se à carbamoílfosfato sintetase I, aumenta a sua actividade. Uma alteração nas concentrações dos
substratos intervenientes neste passo é um factor que, naturalmente, influenciará
também a actividade desta enzima. É ainda de assinalar que quanto mais vezes for
percorrido o ciclo, mais arginina é produzida e portanto mais carbamoíl-fosfato sintetase
I estará a funcionar, garantindo assim uma excreção cada vez mais rápida dos grupos
aminados.
Bibliografia
Principles of Biochemistry 4th Edition - D.L. Nelson, Cox Lehninger - W.H. Freeman 2004
Colour Atlas of Biochemistry 2nd Edition - J.Koolman, K.H.Roehm – Thieme 2005
http://www.bmb.leeds.ac.uk/illingworth/metabol/amino.htm
http://en.wikipedia.org/wiki/Proteasome
http://en.wikipedia.org/wiki/Urea_cycle
http://en.wikipedia.org/wiki/Carbamoyl_phosphate_synthetase_I
http://www2.ufp.pt/~pedros/bq/ureia.htm
http://www.hhmi.org/biointeractive/media/proteasome-lg.mov
Trabalho realizado por: João Carreiras (58404), Frederico Ribeiro (58449) e João Veiga (58757)
21 de Abril de 2008 Mestrado Integrado em Engenharia Biomédica
")