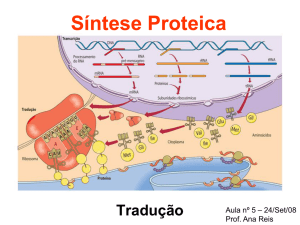
Tradução
Tradução: refere-se a todo o processo pelo qual a sequência
de bases de um mRNA é usada como molde para unir
aminoácidos para a formação de uma proteína.
O DNA guarda as informações para a síntese protéica;
O RNA leva estas informações para o citoplasma, local onde
ocorre efetivamente a síntese das proteínas.
A ordem dos aminoácidos ao longo da cadeia protéica
determina a função desta nova proteína; portanto o
mecanismo que mantém esta ordem durante a síntese é crítico.
3 tipos de RNA estao envolvidos no processo da
traducao:
rRNAs
tRNAs
mRNA
Gene IX, 2008
Um aa e trazido ao ribossomo pelo aminoacyl-tRNA
O mesmo e adicionado a cadeia polipeptidica em
crescimento pela interacao com o tRNA que trouxe o aa
anterior
Cada um desses tRNA ocupam um sitio distinto no
ribossomo
Sitio A: aminoacyl-tRNA (novo aa + novo codon)
Sitio P: peptidyl-tRNA (tRNA carregando a cadeia
polipeptidica em crescimento)
1
Tradução:
A tradução é subdividida em 3 estágios:
Iniciação: compreende o conjunto de reações que precedem a
formação da primeira ligação peptídica da proteína.
-o acoplamento da subunidade menor do ribossomo ao mRNA,
-A união do primeiro tRNA ao códon de início da protéina (AUG),
-A junção das duas subunidades do ribossomo.
Elongação: inclui todas as reações que ocorrem desde a
formação da primeira ligação peptídica até a incorporação do
último aa à proteína.
Terminação: abrange os processos envolvidos na liberação
do polipeptídeo já pronto. O ribossomo separa-se do mRNA
e suas duas subunidades dissociam-se.
Iniciacao
Alongacao
Terminacao
2
Iniciação em Procariotos
Sitio de ligacao do ribossomo – Shine Dalgarno:
35 bases no mRNA – complementar ao rRNA
Um tRNA “iniciador” especial inicia a cadeia
polipeptidica:
A sintese de proteina tem inicio com o aa
metionina (codon AUG)
tRNAs diferentes estao envolvidos na
iniciacao e na alongacao
O tRNA iniciador tem estrutura unica que o
distingue de todos os outros tRNAs
fMet-tRNA pode iniciar síntese protéica:
metionina formilada
fMet-tRNA: reconhece o codon AUG ou GUG
3
A energia necessária
aos diferentes passos
da síntese protéica é
fornecida pela
hidrólise de GTP.
Neste caso, GTP é
necessário para o
posicionamento
correto do códon AUG
na subunidade
ribossômica.
IF-2 liga o iniciador
fMet-tRNA e permite
sua entrada no sitio P
da subunidade 30S
Um mRNA contem muitos codons AUG: Como o codon de
iniciacao eh reconhecido como o ponto de inicio da traducao?
Sitio de ligacao do ribossomo: como eh a sequencia do
mRNA bacteriano?
AUG (menos frequente GUG ou UUG) sempre
presente
10 baes antes do AUG eh a sequencia:
5’…AGGAGG…3’: poli-purinas = sequencia
Shine-Dalgarno
Complementar a uma sequencia altamente
conservada com a extremidade 3’ do rRNA 16S.
Mutacoes na sequencia Shine-Dalgarno podem impedir um
mRNA de ser traduzido.
4
Quando um mRNA bacteriano eh
policistronico:
Cada regiao a ser traduzida apresenta um
sitio de ligacao do ribossomo
Cistron
Iniciação da sintese de proteinas em Eucariotos
Processo de iniciacao semelhante ao de bacteria
No que difere?
Ordem dos eventos diferente e
Numero de fatores de iniciacao maior
O mecanismo de reconhecimento do sinal de iniciação do
mRNA em eucariotos é diferente: o passo mais importante é o
reconhecimento da extremidade 5' do mRNA.
O terminal 5' dos mRNAs eucarióticos têm uma modificação
especial, um resíduo metilado de guanilato, ligado ao primeiro
nucleotídeo normal da cadeia por uma ligação 5' - 5', chamado
"cap".
a iniciação da tradução em eucariotos envolve o
reconhecimento do "cap", seguido da localização da seqüência
de consenso que envolve o códon AUG.
5
Ao contrário dos três fatores de iniciação bem definidos que
iniciam a tradução em bactérias, em eucariontes existe uma
multiplicidade de fatores de iniciação (chamados"eIFs“ – fatores
de iniciacao eucariontes).
Onde atuam?
Formacao do complexo de iniciacao com a extremidade 5’
Na formacao do complexo com o Met-tRNA
No escaneamento do mRNA pelo ribossomo a partir da
extremidade 5’ ate o primeiro AUG
Deteccao da ligacao do tRNA iniciador no sitio AUG
Conexao da subunidade 60S
A subunidade pequena (40S) é a primeira a se complexar com o
mRNA e com o tRNA iniciador. O GTP, também em eucariotos,
é hidrolisado após o acoplamento da subunidade grande (60S) ao
complexo.
Entretanto, há vários aspectos em que a iniciação de eucariotos
difere de iniciação em procariotos:
Met-tRNA iniciador é diferente daquele utilizado na
fase de alongamento, mas a metionina não é
formilada no grupo N-terminal;
Em levedura: o Met-tRNA apresenta estrutura
terciaria diferente e uma fosforilacao na base 64
há um número muito maior de fatores de iniciação
em eucariotos (12 eIFs registrados em levedura) que
em procariotos,
A presença de um terminal 5' com "cap" no mRNA é
necessária para a iniciação.
6
Elongação
Depois que a região de iniciação do mRNA foi corretamente
ligada ao ribossomo, ele sempre se move em uma direção fixa
durante a síntese protéica.
O ribossomo não tem a opção de mover-se para a direita ou para
a esquerda, refletindo o fato do mRNA ter uma direção, definida
pela orientação dos seus terminais 5' e 3'.
A entrada de um novo aminoacyl-tRNA no sitio A é mediada
pelo “Fator de elongacao” (EF-Tu em bacteria)
EF-Tu é uma proteina altamente conservada entre as bacterias e
eucariotos.
EF-Tu esta associada ao ribossomo somente durante o processo
de entrada do aminoacyl-tRNA (associacao ciclica)
Ha cerca de ~70.000 moleculas de EF-Tu por bacteria (~ 5% da
proteina total bacteriana)
Exemplo aplicado:
Kirromicina é um antibiotico que inibe a
função do EF-Tu.
Quando EF-Tu liga-se a kirromicina,
permanece capaz de ligar o aminoacyltRNA ao sitio A, MAS
Não se solta do sitio A
Assim o ribossomo fica “parado” no mRNA
sem prosseguir com a síntese da proteina
Cada ribossomo tem dois sítios para ligação de tRNA. São os
sítios peptidil (P) e Aminoacil (A).
A especificídade para um determinado aminoacil-tRNA é dada
pelo códon específico do mRNA, isto é, embora a cavidade do
ribossomo possa aceitar qualquer aminoacil-tRNA, a superfície
contendo mRNA a torna específica para uma molécula específica.
Met-tRNA i met entra no sítio P.
Todos os aminoacil-tRNAs subsequentes entram no sítio A.
Depois que o segundo aminoacil-tRNA é colocado, corretamente,
no sítio A, uma ligação peptídica se forma entre a carboxila do
primeiro aminoácido e o grupo amina do segundo.
O dipeptídeo resultante fica, portanto, ligado, através da carboxila
do segundo aminoácido, ao segundo tRNA.
7
Então, numa etapa chamada translocação, o peptidil-tRNA e o
códon do mRNA ao qual ele está acoplado, movem-se
coordenademente para o sítio P do ribossomo.
Este processo de adição de aminoácido se repete várias vezes,
até que a cadeia completa se forme.
Alguns pontos devem ser enfatizados:
A formação da ligação peptídica move o ponto de ligação do
sítio P para sítio A (através da enzima peptidil transferase)
O novo tRNA terminal move-se, então, do sítio A para o
sítio P. Ao mesmo tempo, o molde de mRNA ligado à
subunidade pequena do ribossomo move-se para colocar o
códon n+l na posição ocupada anteriormente pelo códon n;
Conjuntamente com a etapa (3), a molécula de tRNA que
liberou seu aminoácido para a formação da ligação peptídica, é
ejetado do sítio P,
O sítio A fica, então, livre para aceitar um novo aminoaciltRNA, cujo especificidade é determinada pelo pareamento
correto de bases entre o anticódon e o códon do mRNA.
8
Terminação
Duas condições são necessárias para a terminação da
síntese protéica:
Uma é a presença de um códon que dá o sinal para o
processo de terminação.
A outra é a presença de uma proteína, o "releasing
factor" (RF), que lê o sinal de terminação de cadeia.
Como a cadeia polipeptídica, após ter sido completada,
ainda está ligada a um peptidyl-tRNA, a etapa de
terminação envolve a quebra de ligação com este tRNA.
Quando isto acontece, a cadeia nascente se dissocia
rapidamente, uma vez que sua ligação com o ribossomo
ocorria através desta ligação.
Há três sinais que significam o final da tradução: UAG,
UAA e UGA.
estes códons são reconhecidos por proteínas, os "releasing
factors“ (RF).
Em E. coli, há dois RFs bem caracterizados:
RF l, que reconhece os codons UAG e UAA, e
RF2, que reconhece UGA e UAA.
Em eucariotos, há apenas um fator de terminação RF, que
reconhece os três codons de terminação.
Eles funcionam induzindo a peptidil transferase a transferir
a cadeia crescente para água, ao inves do aminoacil-tRNA.
Após a terminação da cadeia, a subunidade maior do ribossomo
(50S em procariotos e 60S em eucariotos), é liberada do mRNA
antes da subunidade menor (30S em procariotos, 40S em
eucariotos).
As subunidades ficam, então, disponíveis para iniciar a síntese
de outro polipeptídeo.
9