EFEITOS ANATOMOPATOLÓGICOS APÓS CONTAMINAÇÃO IN VITRO POR
CLORETOS DE MERCÚRIO E DE CÁDMIO EM TESTÍCULOS DO PEIXE
TELEÓSTEO Gymnotus carapo (tuvira) (LINNAEUS, 1758)
RENATA VASCONCELOS MOREIRA
UNIVERSIDADE ESTADUAL DO NORTE FLUMINENSE
DARCY RIBEIRO - UENF
CAMPOS DOS GOYTACAZES/RJ
NOVEMBRO DE 2010
EFEITOS ANATOMAPATOLÓGICOS APÓS CONTAMINAÇÃO IN VITRO POR
CLORETOS DE MERCÚRIO E DE CÁDMIO EM TESTÍCULOS DO PEIXE
TELEÓSTEO Gymnotus carapo (tuvira) (LINNAEUS, 1758)
RENATA VASCONCELOS MOREIRA
Dissertação apresentada ao Centro
de Biociências e Biotecnologia da
Universidade
Estadual
Fluminense
como
do
Norte
parte
das
exigências para obtenção do Título de
Mestre em Biociências.
Orientador: Prof. Dr. Edésio José Tenório de Melo
UNIVERSIDADE ESTADUAL DO NORTE FLUMINENSE
DARCY RIBEIRO - UENF
CAMPOS DOS GOYTACAZES/RJ
NOVEMBRO DE 2010
i EFEITOS ANATOMOPATOLÓGICOS APÓS CONTAMINAÇÃO IN VITRO POR
CLORETOS DE MERCÚRIO E DE CÁDMIO EM TESTÍCULOS DO PEIXE
TELEÓSTEO Gymnotus carapo (tuvira) (LINNAEUS, 1758)
Dissertação apresentada ao Centro
de Biociências e Biotecnologia da
Universidade
Estadual
Fluminense
como
do
Norte
parte
das
exigências para obtenção do Título de
Mestre em Biociências.
Aprovada em 12 de novembro de 2010.
Comissão examinadora:
_________________________________________________________________
Prof. Carlos Eduardo de Rezende (Dr. em Ciências Biológicas - Biofísica) - UENF
_________________________________________________________________
Prof. Eulógio Carlos Queiroz de Carvalho (Dr. em Patologia - Anatomia Patológica) UENF
_________________________________________________________________
Prof. Leonardo Gomes da Silva (Dr. em Biociências e Biotecnologia) - UFRJ
________________________________________________________________
Prof. Edésio José Tenório de Melo (Dr. em Ciências Biológicas - Biofísica) - UENF
(Orientador)
ii “Não é o desafio com que nos deparamos que
determina quem somos e o que estamos nos tornando,
mas a maneira com que respondemos ao desafio.
Somos combatentes, idealistas, mas plenamente conscientes.
Problemas para vencer, liberdade para provar.
E, enquanto acreditamos em nosso sonho,
nada é por acaso.”
(Henfil)
iii AGRADECIMENTOS
As nossas principais conquistas não são frutos de um trabalho isolado, tão
pouco individual, por isto agradeço, primeiramente, ao meu orientador Prof. Dr.
Edésio José Tenório de Melo, pela oportunidade de crescimento e por ter acreditado
em mim! Obrigada.
À minha querida mãe, pelo incentivo e apoio não só durante este período,
mas em toda minha vida.
Aos meus irmãos Alex e Alan, pela união, respeito e amizade.
Ao meu marido Leonardo pela cumplicidade e amor incondicional. Meu porto
seguro. Amo você!
À Juliana, minha “irmã”, pela amizade e, sobretudo, pelo apoio e
encorajamento para superar as dificuldades vividas.
Ao meu amigo Diogo, pelo carinho e por estar sempre me fazendo sorrir com
seu jeito espontâneo de ser.
A Cristiane Vergílio, pela ajuda indispensável durante a execução deste
trabalho, e, é claro, pelas sugestões e críticas.
Ao pescador César.
Ao Prof. Dr. Carlos Eduardo Veiga de Carvalho pela colaboração e incentivo.
Ao Prof. Eulógio Carlos Queiroz de Carvalho e aos técnicos Luciano Grillo de
Almeida, Luciana da Silva Lemos, Ricardo Guerreiro e Elizabeth Gonçalves Pires do
Setor de Morfologia e Anatomia Patológica do Laboratório de Sanidade Animal
(CCTA/UENF), por todo auxílio e disponibilidade.
iv Aos funcionários do Laboratório de Biologia Celular e Tecidual, especialmente
as técnicas: Beatriz Ferreira Ribeiro e Giovanna Alves de Moraes, pela ajuda no
desenvolvimento da pesquisa.
Ao Laboratório de Ciências Ambientais (LCA).
As meninas do Setor de Toxicologia Celular: Carol e Laís pela ajuda nos
aquários e pela amizade, que mesmo em pouco tempo, foi muito importante para
mim.
A todos os professores do Laboratório de Biologia Celular e Tecidual, por todo
conhecimento transmitido durante esses anos. Em especial ao Prof. Dr. Flávio Costa
Miguens.
Aos membros da banca examinadora: Carlos Eduardo de Rezende, Eulógio
Carlos Queiroz de Carvalho e Leonardo Gomes da Silva.
À CAPES, FAPERJ e UENF pelo apoio logístico e financeiro.
v SUMÁRIO
LISTA DE FIGURAS............................................................................................................... vii
LISTA DE TABELAS.............................................................................................................. viii
ABREVIATURAS ..................................................................................................................... ix
RESUMO ..................................................................................................................................x
ABSTRACT.............................................................................................................................. xi
1.INTRODUÇÃO.......................................................................................................................1
1.1.Metais Pesados ..................................................................................................................1
1.2.Mercúrio..............................................................................................................................3
1.3.Cádmio ...............................................................................................................................6
1.4.Contaminação in vitro e Exposição aguda .........................................................................8
1.5.Peixes nos estudos de Toxicologia ............................................................ .......................9
1.5.1.Espécie utilizada: Gymnotus carapo .............................................................................10
1.6.Testículos e espermatogênese em peixes .......................................................................14
1.7.Histopatologia.... ...............................................................................................................17
1.8.Efeito dos metais pesados nos testículos.........................................................................18
2.OBJETIVOS.........................................................................................................................20
2.1.OBJETIVO GERAL...........................................................................................................20
2.2.OBJETIVOS ESPECÍFICOS ............................................................................................20
3.MATERIAL E MÉTODOS ....................................................................................................21
3.1.Coleta dos exemplares de Gymnotus carapo...................................................................21
3.2.Preparação de soluções ...................................................................................................22
3.3.Contaminação dos exemplares de Gymnotus carapo por HgCl2 e CdCl2 ........................22
3.4.Preparação para Microscopia Óptica (MO) ......................................................................26
3.5.Preparação para Microscopia Eletrônica de Transmissão (MET) ....................................27
3.6.Preparação para Microscopia Eletrônica de Varredura (MEV).........................................27
4. RESULTADOS. ..................................................................................................................29
4.1.Efeitos tóxicos do HgCl2 e CdCl2 nos exemplares de Gymnotus carapo..........................29
4.2.Aspectos anatômicos do testículos de Gymnotus carapo ................................................30
4.3.Aspectos histológicos .......................................................................................................32
4.4. Aspectos ultraestruturais .................................................................................................40
4.5. Aspectos morfológicos dos espermatozóides de Gymnotus carapo ...............................43
5.DISCUSSÃO........................................................................................................................43
6.CONCLUSÕES....................................................................................................................48
7.REFERÊNCIAS BIBLIOGRÁFICAS ....................................................................................49 vi LISTA DE FIGURAS
Figura 1.Tabela periódica dos elementos químicos....................................................2
Figura 2. Principais alvos para toxicidade do Hg........................................................5
Figura 3. Foto da espécie Gymnotus carapo............................................................10
Figura 4. Localização do órgão elétrico em Gymnotiformes.....................................11
Figura 5. Localização dos testículos de Gymnotus carapo.......................................13
Figura 6. Testículos de Gymnotus carapo.................................................................14
Figura 7. Espermatogênese em peixes.....................................................................15
Figura 8. Localização da Lagoa de cima...................................................................21
Figura 9. Aspectos anatômicos do testículo de Gymnotus carapo............................31
Figura 10. Região do testículo de Gymnotus carapo onde foram realizados cortes
transversais................................................................................................................32
Figura 11. Organização estrutural do testículo de Gymnotus carapo (grupo
controle)......................................................................................................................35
Figura 12. Organização estrutural do testículo de Gymnotus carapo (grupo exposto
a 5µM e 10µM de HgCl2)............................................................................................36
Figura 13. Organização estrutural do testículo de Gymnotus carapo (grupo exposto
a 20µM e 30µM de HgCl2)..........................................................................................37
Figura 14. Organização estrutural do testículo de Gymnotus carapo (grupo exposto
a 10µM, 30µM e 40µM de CdCl2)...............................................................................38
Figura 15. Corte transversal dos testículos de Gymnotus carapo.............................39
Figura 16. Organização ultraestrutural do testículo de Gymnotus carapo (grupo
controle)......................................................................................................................41
Figura 17. Organização ultraestrutural do testículo de Gymnotus carapo (grupo
exposto ao HgCl2 e CdCl2).........................................................................................42
Figura 18. Fotomicrografia dos espermatozóides de Gymnotus carapo corados com
Giemsa.......................................................................................................................44
Figura 19. Fotomicrografia (MEV) dos espermatozóides de Gymnotus carapo........45
Figura 20. Porcentagem de espermatozóides do grupo controle e do grupo exposto
ao HgCl2 e CdCl2 com flagelo e sem flagelo..............................................................46
vii LISTA DE TABELAS
Tabela 1. Resumo das alterações histológicas nos testículos de peixes causados
por metais pesados....................................................................................................19
Tabela 2. Tabela de conversão das concentrações de HgCl2 em µM para µg/g...... 24
Tabela 3 Tabela de conversão das concentrações de CdCl2 em µM para µg/g........24
Tabela 4. Número amostral, comprimento padrão e peso dos exemplares de
Gymnotus carapo utilizados para o experimento de HgCl2........................................25
Tabela 5. Número amostral, comprimento padrão e peso dos exemplares de
Gymnotus carapo utilizados para o experimento de CdCl2........................................25
Tabela 6. Numero de exemplares de Gymnotus carapo que morreram durante a
exposição ao HgCl2....................................................................................................29
Tabela 7. Número de exemplares de Gymnotus carapo que morreram durante a
exposição ao CdCl2....................................................................................................29
Tabela 8. Número amostral, peso do testículo e o índice gonadossomático dos
exemplares de Gymnotus carapo expostos ao HgCl2................................................31
Tabela 9. Número amostral, peso do testículo e o índice gonadossomático dos
exemplares de Gymnotus carapo expostos ao CdCl2................................................32
viii ABREVIATURAS
Cd: Cádmio
CdCl2: Cloreto de cádmio
EODs: Descargas dos órgãos elétricos
Hg: Mercúrio
HgCl2: Cloreto de mercúrio
HgT: Concentração de mercúrio total
it: Tecido intersticial
L: Célula de Leydig
ppb: Parte por bilhão
ppm: Parte por milhão
PBS: Phosphate buffer solution
ROS: Espécies reativas de oxigênio
S: Célula de Sertoli
SPCI: Espermatócito primário
SPCII: Espermatócito secundário
SD: Espermátide
SPG: Espermatogônia
SPGI: Espermatogônia primária
SPGII: Espermatogônia secundária
SZ: Espermatozóide
ix RESUMO
Poluentes ambientais são lançados diariamente nos ecossistemas aquáticos e
podem trazer sérias conseqüências para os organismos, dentre eles destacam-se os
metais. Mercúrio e cádmio são metais reconhecidos por sua alta toxicidade e grande
persistência no ambiente. Estudos anteriores demonstraram que mesmo em baixas
concentrações, estes metais têm a capacidade de afetar o sistema reprodutor dos
peixes, inibindo o desenvolvimento das gônadas e a espermatogênese. Com isso, o
presente estudo tem como objetivo verificar se o HgCl2 e CdCl2 provocam alterações
nos testículos e espermatozóides de Gymnotus carapo (tuvira). A contaminação foi
realizada através de injeção intraperitoneal de solução de HgCl2 ou CdCl2
(5μM/10μM/20μM/30μM/40µM) por 24h, 48h, 72h e 96h. Alterações histológicas
foram identificadas nos testículos dos peixes expostos ao HgCl2 e CdCl2, porém no
grupo tratado com HgCl2 os efeitos foram considerados mais severos. Após
tratamento com 5µM de HgCl2, alterações histológicas foram observadas a partir de
72 horas, enquanto que para 10μM, 20µM e 30μM essas alterações foram
visualizadas em todos os tempos de exposição analisados. Em relação ao CdCl2, as
alterações foram observadas a partir de 72h de exposição com 10µM. As lesões
encontradas no tecido foram semelhantes em todas as concentrações
administradas, sendo os efeitos observados progressivos com o aumento da
concentração ao metal exposto. As alterações mais significativas identificadas no
testículo dos espécimes expostos ao HgCl2 e CdCl2, incluem fibrose intersticial,
hiperemia, desintegração intersticial e tubular, desorganização dos túbulos
seminíferos, degeneração e agregação espermática. Além disso, pela análise
ultraestrutural observamos as células germinativas em apoptose e em processo de
vacuolização. Mudanças morfológicas foram visualizadas em espermatozóides de G.
carapo. Após o tratamento com 20μM de HgCl2 e CdCl2 por 72/96h respectivamente,
foram observadas deformidades na porção da cabeça e ausência de flagelo.
Portanto, os resultados apresentados no presente estudo demonstram mudanças no
testículo e nos espermatozóides de G. carapo, após exposição ao HgCl2 e CdCl2.
Essas mudanças consequentemente vão afetar a reprodução desta espécie,
levando prejuízos à nível de população. Assim as informações obtidas neste estudo
demonstram que histopatologia gonadal pode ser um biomarcador de poluição por
metais pesados nesta espécie de peixe.
Palavras-chave: testículo, Gymnotus carapo, mercúrio, cádmio, histopatologia,
peixe.
x ABSTRACT
Environmental pollutants are released daily in aquatic ecosystems and can have
serious consequences for the organisms, among the polluents include the metals.
Metals mercury and cadmium are known for their high toxicity and great persistence
in the environment. Previous studies had show that even at low concentrations, these
metals are able to affect the reproductive system of fish, inhibiting gonad
development and spermatogenesis. Thus, this study aims to determine if the HgCl2
and CdCl2 can causes changes in the testes and spermatozoa of Gymnotus carapo
(tuvira). The contamination was performed by intraperitoneal injection of HgCl2 or
CdCl2 solution (5μM/10μM/20μM/30μM/40µM) for 24h, 48h, 72h e 96h. Histological
changes were identified in the testes of fish exposed to HgCl2 and CdCl2, but in the
group treated with HgCl2 effects were considered severe. After treatment with 5μM
HgCl2, histopathological changes were observed after 72 h, while for 10μM, 20μM
and 30μM these changes were seen at all exposure times examined. In relation to
CdCl2, changes were observed from 72 h exposure to 10µM. The lesions found in the
tissue were similar at all concentrations administered, and the progressive effects
observed with increasing concentration of the exposed metal. The most significant
changes identified in the testes of fish exposed to HgCl2 and CdCl2, includes fibrosis,
blood vessel congestion or hyperemia, interstitial and lobular disintegration,
disorganization of the seminiferous lobules degeneration, sperm aggregation.
Furthermore, by ultrastructural analysis we can observe the germ cells in apoptosis
and vacuolization. Morphological changes also were observed in spermatozoa of G.
carapo. After treatment with 20µM of HgCl2 and CdCl2 for 72/96h respectively,
deformities were observed on the head and also the flagellum absence. Therefore,
the results presented in this study show changes in the testis and spermatozoa of G.
carapo after exposure to HgCl2 and CdCl2. These changes will go affect the
reproductive success of this species, which may cause damage to the level of
population. Thus the information obtained during this study show that gonadal
histopathology may be a biomarker of heavy metal pollution in this fish species.
Keywords: testes, Gymnotus carapo, mercury, cadmium, histopathology, fish.
xi 1. INTRODUÇÃO
A ação antropogênica vem aumentando a concentração de vários compostos
químicos no ambiente aquático. Estes poluentes podem provocar graves
consequências em vertebrados e invertebrados e, com o passar do tempo, podem
comprometer sua permanência no ecossistema (Jobling,1995).
Dentre os contaminantes mais intensamente estudados nos ecossistemas
aquáticos estão os metais (Palenzuela et al., 2004). Os metais podem interromper
ou prejudicar a reprodução de animais aquáticos, afetando o sistema endócrino e
agindo nos órgãos reprodutivos e nos gametas desses organismos. Como resultado
desse distúrbio, o processo de desenvolvimento das gônadas é seriamente
prejudicado juntamente com a taxa de fertilização (Dietrich et al, 2004; Rurangwa,
1998).
1.1. METAIS PESADOS
A denominação “metal pesado” é aplicada a um grupo heterogêneo de
elementos, incluindo metais, semi metais e não metais (Figura 1) que possuem
número atômico maior do que 20 ou peso específico maior que 5 g cm-3 (Malavolta,
1994). São denominados tóxicos aqueles metais que produzem danos em pequenas
concentrações, dentre eles estão arsênio, cádmio, cobre, cromo, chumbo e
mercúrio. Em geral, metais tóxicos estão presentes em pequenas quantidades no
meio aquático por ação de fenômenos naturais, mas podem ser despejados em
quantidades significativas por atividades industriais, agrícolas e de mineração (Abel,
1989).
Um
organismo
aquático
pode
apresentar
dois
tipos
básicos
de
comportamento em relação aos metais: ou é sensível à ação tóxica de um
determinado metal, ou não é sensível, mas o bioacumula, potencializando seu efeito
nocivo através das cadeias alimentares de forma a colocar em risco organismos
situados nos topos destas cadeias (Viarengo, 1989).
1 Figura 1. Tabela periódica dos elementos químicos.
Fonte: grandeabobora.com
Os efeitos tóxicos dos metais podem incluir tanto a letalidade e efeitos
subletais, como alterações no crescimento, desenvolvimento, reprodução e
respostas farmacocinéticas, patológicas, bioquímicas e comportamentais (Baatrup,
1991).
A toxicidade dos metais está relacionada à sua capacidade de interferir em
reações enzimáticas, por meio do bloqueio ou deslocamento do íon essencial de
várias biomoléculas. Ainda possuem baixa mobilidade, em virtude das pequenas
dimensões e das cargas duplas e triplas, fazendo com que se acumulem por longo
período no ambiente (Conell & Miller, 1984).
A alta toxicidade de alguns metais para os organismos, mesmo em baixas
concentrações, reforça a importância de estudos relacionados com os efeitos
induzidos por esses elementos (Baatrup, 1991). Dentre os metais considerados
“pesados”, mercúrio e cádmio geram grande preocupação ambiental devido a sua
ampla utilização industrial, toxicidade e comportamento no ambiente (Búcio et al.,
1995).
2 Informações importantes sobre o potencial tóxico dos metais podem ser
obtidas de ambientes contaminados ou através de estudos de simulação em
laboratório, diferindo quanto ao objetivo e metodologias de avaliação. Estudos em
toxicologia ambiental devem considerar alguns aspectos importantes: modelo de
estudo, parâmetros analisados e desenho experimental (campo ou laboratório).
Deste modo, é possível contribuir com informações que possibilitem um melhor
entendimento do processo toxicológico do metais.
1.2. MERCÚRIO
O mercúrio é um elemento químico de número atômico 80, cujo símbolo Hg
deriva do nome grego Hidrargyrium, o qual significa prata líquida. Encontra-se no
grupo IIB da tabela periódica de classificação dos elementos e é o único metal em
estado líquido à temperatura ambiente (Hoffman et al., 1995).
O mercúrio é considerado altamente tóxico, devido a sua capacidade de
bioacumular nos organismos, biomagnificar através da cadeia alimentar e possuir
uma intensa ciclagem no ecossistema (Förstner & Wittman,1983). Dentre as formas
químicas destacam-se o mercúrio metálico Hg0 que é volátil e libera um gás
monoatômico: o vapor de mercúrio. Este é estável, podendo permanecer na
atmosfera por meses ou até anos, revelando-se, deste modo, muito importante no
ciclo do mercúrio, pois pode sofrer oxidação, originando o mercúrio mercuroso (Hg+1)
e o mercúrio mercúrico (Hg+2). Essas formas inorgânicas podem se ligar a elementos
como o cloro, enxofre ou oxigênio e originar compostos de mercúrio inorgânico
também designado como sais de mercúrio (sais mercurosos e mercúricos) (Malm et
al.,1997)
Um dos sais mais importantes industrialmente é o cloreto de mercúrio (I) ou
calomelano (Hg2Cl2) que é um composto branco, pouco solúvel em água e utilizado
como purgante, anti-helmíntico e diurético, e o cloreto de mercúrio (II) (HgCl2),
um sublimado corrosivo muito tóxico, empregado como desinfetante e utilizado em
muitas aplicações, incluindo as medicinais, entretanto, é uma das formas mais
tóxicas do mercúrio pela sua solubilidade em água em relação a outros compostos
de mercúrio (HSDB, 2000).
3 Quando um átomo de mercúrio se liga covalentemente a grupos metil (CH3),
dá origem a um composto orgânico conhecido como metilmercúrio. O metilmercúrio
(CH3Hg), por sua vez, é originado a partir do processo de metilação, através do
Hg2+,
mediado
por
diversos
microorganismos
em
ambientes
aquáticos,
particularmente no sedimento (Malm et al.,1997).
As fontes naturais mais significativas de mercúrio são gaseificação da crosta
terrestre, as emissões de vulcões e a evaporação de corpos aquáticos (Who, 1991).
As fontes antrópicas ocorrem através de indústrias que queimam combustíveis
fósseis, produção eletrolítica de cloro-soda, produção de acetaldeído, incineradores
de lixo, polpa de papel, tintas, pesticidas, fungicidas, lâmpadas de vapor de
mercúrio, baterias, produtos odontológicos, amalgamação de mercúrio em extração
de ouro, entre outros (Canela,1995).
Independente da origem, o mercúrio na forma orgânica ou inorgânica pode
sofrer transformações no ambiente, o que torna sua distribuição ambiental bastante
complexa. Em geral o ciclo biogeoquímico do mercúrio exibe um equilíbrio dinâmico,
mas ações humanas podem introduzir elementos ou compostos mais rápidos e em
nível maior que os processos naturais, alterando os padrões dos ciclos e as
condições às quais a flora e a fauna estão adaptadas (Mauro et al., 1999).
O mercúrio atinge os ecossistemas aquáticos através da precipitação e por
meio do descarte de resíduos agro-industriais na proximidade de rios, lagos e
lagoas. A absorção deste elemento nos peixes ocorre pela alimentação ou
respiração branquial e a sua distribuição acontece por meio da circulação
sanguínea. A absorção do mercúrio é influenciada pela concentração deste
elemento, pela taxa metabólica e pela eficiência de absorção (disponibilidade),
determinada pelas características do ambiente aquático. Os níveis de mercúrio na
biota aquática variam entre as espécies de uma mesma localidade e para uma
mesma espécie em diferentes localidades (Azevedo, 2003).
A toxicidade do mercúrio e seus compostos se deve a sua forte interação com
grupos sulfidrila, R-SH de proteínas, enzimas (Lacerda, 1997; Baatrup, 1991) e
proteínas de baixo peso molecular (como coenzima A, cisteína, glutationa, entre
outros) (Valle & Ulmer, 1972). Nas sulfidrilas, o Hg2+ substitui o átomo de hidrogênio,
sendo capaz de inativar enzimas sulfidriladas, interferindo no metabolismo e nas
funções celulares. O mercúrio também se liga, afetando a configuração de purinas e
pirimidinas e ácidos nucléicos podendo levar danos ao DNA (Bucio et al., 1999).
4 Além disso, pode reagir diretamente com vários constituintes de membrana,
interferindo na integridade estrutural e nas propriedades físicas das membranas
celulares (Baatrup, 1991). A capacidade dos metais pesados atravessarem a
membrana para o interior das células pode prejudicar suas funções, causando a
morte celular e a destruição de qualquer tecido com a qual entrem em contato
(Azevedo, 2003).
Além disso, em condições fisiológicas normais, os animais possuem um
balanço entre a geração e neutralização das espécies reativas de oxigênio (ROS).
Entretanto, quando os organismos estão expostos aos Hg e a outros metais
pesados, a taxa de produção das ROS excede a sua capacidade de neutralização
pelas defesas antioxidantes dos organismos (Figura 3), levando ao estresse
oxidativo (Stohs & Bagchi, 1995). Células sob estresse oxidativo desenvolvem várias
disfunções, em razão das lesões causadas pelas ROS aos lipídios, proteínas e DNA
(Ercal et al., 2001; Van Der Oost et al., 2003).
Assim, torna-se necessário conhecer os reais danos à saúde dos organismos
em diferentes níveis de organização biológica frente à exposição crônica e aguda a
este metal.
Figura 2. Principais alvos para toxicidade do Hg.
5 1.3. CÁDMIO
O cádmio é um elemento químico de símbolo Cd de número atômico 48 e
assim como o Hg está situado no grupo IIB (grupo 12) da tabela periódica de
classificação dos elementos. É um metal branco azulado que à temperatura
ambiente encontra-se no estado sólido.
O cádmio pode ser liberado no meio ambiente através de fontes naturais ou
antropogênicas, dentre as principais fontes naturais destaca-se o vulcanismo, spray
oceânico e queimadas de florestas (Hutton, 1987). Por outro lado, um aumento das
concentrações de Cd no ambiente tem ocorrido principalmente em função de suas
inúmeras aplicações industriais, como nos processos de galvanoplastia, além de ser
um elemento constituinte de baterias, pilhas, pigmentos, revestimentos elétricos,
estabilizantes e catalisadores (Fielder & Dale, 1983). Com isso, a produção,
utilização e emissões de Cd para o ambiente têm aumentado dramaticamente
durante o século 20, e consequentemente levado a contaminação dos ecossistemas
aquáticos (Burger, 2008).
A exposição ao cádmio nos humanos ocorre geralmente através de duas
fontes principais: a primeira é por via oral (por água e ingestão de alimentos
contamnados), e a segunda por inalação (através da fumaça do cigarro). Nesse
sentido, a inalação do fumo ou de aerossóis contendo cádmio pode causar doenças
pulmonárias crônicas ou agudas. Dentro de certas indústrias, a exposição
ocupacional é a fonte de contaminação predominante (Friberg, 1983). O cádmio
entra na corrente sanguínea, sendo absorvido pelo estômago ou intestino após a
ingestão do alimento ou da água, ou por absorção nos pulmões após a inalação. De
qualquer forma, uma vez que o cádmio é absorvido é fortemente retido, inclusive
baixas doses deste metal podem constituir um nível significativo no organismo
(Marettová et al., 2010).
Um vez absorvido, o cádmio é transportado pela corrente sanguínea até o
fígado.
Pequenas
quantidades
desse
complexo
proteína-cádmio
passam
continuamente do fígado para a corrente sanguínea, para ser transportado até os
rins. O rim excreta de 1 a 2% do cádmio obtido diretamente das fontes ambientais. A
concentração do metal nos rins é aproximadamente 10 mil vezes mais alta que a da
corrente sanguínea. A excreção fecal do metal representa uma mínima quantidade
6 do cádmio não absorvido no sistema gastrointestinal (Burger, 2008).
Além do fígado, rim e pulmões, outros órgãos também são danificados por
exposição ao cádmio em animais ou humanos, incluindo o sistema reprodutor, o
sistema imunológico e o sistema nervoso (Jarup; Akesson, 2009).
A toxicidade do cádmio está relacionada com a sua capacidade de substituir o
cálcio nas reações biológicas. Cádmio e cálcio são dois elementos intimamente
relacionados, com similaridades em muitos aspectos, devido a sua semelhança nos
raios iônicos. A captação celular de cádmio, ocorre principalmente através dos
canais de Ca +2 e Cd torna-se um potente bloqueador dos canais de Ca +2, inibindo a
captação celular de Ca +2 (Beyersmann & Hechtenberg, 1997).
O cádmio também é capaz de causar um aumento no estresse oxidativo pela
ligação a grupos sulfidrilas de proteínas e pela diminuição da glutationa (Valko et al.,
2005). Consequentemente, o estresse oxidativo pode promover alteração nos
mecanismos de reparo do DNA e indução da proliferação celular, que, por sua vez,
pode levar a tumorigênese (Beyersmann & Hartwig, 2008). Além disso, o cádmio se
liga preferivelmente a resíduos de cisteína de proteínas, em particular com a
metalotioneína e também pode inibir muitas enzimas. A metalotioneína é uma
enzima importante no transporte e armazenamento do cádmio e outros metais (Who,
1992).
Apesar de muitas pesquisas analisarem a concentração de cádmio em
tecidos internos dos organismos (Carignan & Villard, 2001), existe uma maior
necessidade de estudos em laboratório com este metal para avaliação dose-efeito
(tanto subletal como letal) tanto a nível celular como a nível tecidual (Burger &
Gochfeld, 2001; Stansley et al., 1991). Futuras descobertas nesse campo ajudariam
a aprofundar o conhecimento sobre os mecanismos básicos que determinam os
danos causados pelo cádmio, permitindo um maior conhecimento da sua toxicidade.
7 1.4. CONTAMINAÇÃO IN VITRO E EXPOSIÇÃO AGUDA
A realização de experimentos em laboratório é importante para a observação
dos efeitos nocivos do mercúrio e cádmio, na ausência de ações sinérgicas ou
antagônicas que podem ocorrer com outros elementos do ambiente. Nesse sentido,
os bioensaios de contaminação em laboratório oferecem à vantagem de avaliar a
toxicidade dos metais pesados em concentrações específicas, com isso as
mudanças induzidas por esses elementos utilizados podem ser empregadas para
observação da relação dose/resposta dos organismos expostos (Vergílio, 2009).
Segundo Rand e Petrocelli (1985), estudos em toxicologia aquática podem
ser qualitativos e quantitativos em relação aos efeitos tóxicos sobre os organismos
aquáticos. A toxicologia aquática também está relacionada com as concentrações
dos agentes químicos que podem ocorrer no ambiente aquático (água, sedimento e
alimento). A toxicidade de um composto químico depende da exposição, da
suscetibilidade do organismo, das características químicas do agente e de fatores
ambientais. A exposição é o contato/reação entre o organismo e o composto
químico, sendo que os fatores mais importantes relacionados à exposição são: o
tipo, a duração e freqüência da exposição e a concentração do agente químico.
Com relação à duração e a freqüência da exposição dos organismos ao
agente químico, Rand e Petrocelli (1985) afirmaram que na exposição aguda, os
organismos entram e contato com o composto químico num evento único ou em
eventos múltiplos que ocorrem num pequeno período de tempo, geralmente
variando de horas a dias. Nas exposições agudas, onde o agente químico é
rapidamente absorvido, geralmente os efeitos são imediatos, embora seja possível a
produção de efeitos retardados similares àqueles resultantes da exposição crônica.
A freqüência de exposição também afeta a toxicidade dos compostos
químicos. Uma exposição aguda pode resultar num efeito adverso imediato num
organismo, enquanto duas exposições sucessivas, com que a soma equivale à
exposição aguda única podem não ter efeito ou este ser pequeno, devido ao
metabolismo (desintoxicação) do composto químico pelo organismo entre as
exposições ou à adaptação do organismo ao composto. Organismos estressados
em função de exposição prévia a outras substâncias tóxicas também podem ser
mais susceptíveis aos compostos químicos, cenário comum na realidade dos
8 ecossistemas, pois normalmente há a presença simultânea de diferentes produtos
(Barak e Mason, 1990).
1.5. PEIXES NOS ESTUDOS DE TOXICOLOGIA
Os peixes constituem um grupo de grande importância nas avaliações de
toxicidade ambiental, pois refletem a saúde do ecossistema aquático, ambiente
fundamental para a sobrevivência dos seres vivos. Além disso, são vertebrados de
ampla distribuição e participam de diferentes níveis tróficos da cadeia alimentar,
sendo considerados excelentes modelos biológicos de estudo (Mela, 2004).
Os teleósteos são considerados excelentes organismos para o monitoramento
e estudo da disponibilidade dos metais pesados para as populações humanas, já
que os peixes podem acumular esses elementos em seus tecidos (Bastos et al.,
2006; Lacerda, 1997; Pfeiffer et al.,1992). Porém, deve ser considerado que alguns
fatores influenciam na acumulação de metais pesados pelos organismos, como o
tamanho, idade, ecologia do animal e hábitos alimentares (Haines et al., 1992).
A principal via de acesso à contaminação por metais pesados ao homem é
através do consumo de pescado. Os peixes constituem uma importante fonte de
proteínas, em especial para as populações que vivem nas proximidades dos corpos
d’água (Pfeiffer et al.,1992 ; Silva Filho et al., 2000).
9 1.5.1. ESPÉCIE UTILIZADA: Gymnotus carapo.
A tuvira (Gymnotus carapo) (Linnaeus, 1758) (Figura 5) é um peixe da classe
dos Actinopterígios de água doce e pertence à ordem dos Gymnotiformes, família
dos Gymnotidae (FishBase, 2010). Essa espécie é também conhecida sarapoá,
peixe espada, sarapó, carapó e ituí, sendo muito utilizada como isca viva na pesca
esportiva por ser a mais preferida de espécies consideradas nobres, como os
piscívoros de grande porte.
Figura 3. Foto da espécie Gymnotus carapo (Tuvira).
Os Gymnotiformes apresentam corpo anguiformes, abertura branquial muito
estreita e são desprovidos das nadadeiras dorsal e ventral; possuem nadadeira anal
muito longa, estendendo-se por quase toda a face ventral (Theodoro, 2003).
Também possuem órgãos elétricos que são utilizados para produzir choques (p. ex.:
poraquê) ou para a eletrocominucação (p. ex.: tuvira) promovida pela emissão de
uma pequena descarga elétrica. A tuvira utiliza o campo elétrico gerado por esta
descarga para sua orientação, que compensa sua falta de visão na hora de predar
espécies menores, como também para suas interações sociais, principalmente na
época de reprodução (Westby, 1975).
10 Figura 4. Localização do órgão elétrico em Gymnotiformes (A). Para localizar um
objeto, o cérebro do peixe elétrico emite um sinal de comando excitando os
eletrócitos (células contidas nos órgãos elétricos), essas células se tornam
polarizadas agindo como baterias interconectadas, resultando em descargas dos
órgãos elétricos (EODs) que são emitidas na água, gerando um campo elétrico (B)
(adaptado de Markham et al., 2009).
As tuviras habitam em locais rasos, com profundidades inferiores a um metro,
em áreas marginais de baías e vazantes, com bastante vegetação aquática
(Resende, 1999). Podem ainda sobreviver em valas, canais e pequenos lagos secos
(FishBase, 2010). Caracterizam-se por respiração aérea facultativa, sendo uma
adaptação comum em peixes que vivem em ambientes hipóxicos e pela bexiga
natatória altamente vascularizada (Moraes et al., 2002). Estudos feitos por Resende
(1999) e Resende e Pereira (2000) sobre os aspectos biológicos e ecológicos da
tuvira demonstram que esses peixes vivem em ambientes lênticos, com plantas
aquáticas, onde se abrigam e encontram alimento. Possuem ainda hábito noturno,
saindo no crepúsculo para águas mais abertas (Menin, 1989; Barbieri & Barbieri,
1984).
Silva & Oliveira (1997) concluíram que a tuvira está adaptada a uma dieta
predominantemente carnívora, com o bolo alimentar sofrendo uma rápida absorção
ao chegar ao intestino. Dessa forma, o intestino deste peixe está adaptado para uma
dieta constituída principalmente de pequenos organismos de fácil digestão. Nesta
espécie, a presença do estômago químico desenvolvido sugere uma adaptação à
digestão de uma dieta rica em proteínas.
11 Quando em ambiente natural, a tuvira alimenta-se principalmente à noite de
vermes,
insetos
(por
exemplo,
larvas
de
libélulas),
camarão,
peixe
(preferencialmente caracídeos de pequeno porte, por exemplo, Ctenobrycon e
curimatã) e matéria vegetal (FishBase, 2010).
A tuvira apresenta crescimento rápido em comprimento no seu primeiro ano
de vida, antes de alcançar a maturidade sexual, quando os indivíduos atingem, em
média, cerca de 20 cm de comprimento. Já quanto ao peso, o comportamento é o
inverso, uma vez que esses peixes começam a ganhar mais peso após o segundo
ano de vida (Barbieri & Barbieri, 1984).
Barbieri & Barbieri (1983), estudando um ambiente lêntico-lótico (represa) em
São Paulo observaram que houve uma relação direta entre o período reprodutivo de
Gymnotus sp. e a variação dos fatores abióticos, como temperatura da água,
concentração de oxigênio dissolvido, precipitação pluviométrica e fotoperíodo, não
evidenciando relação com a variação do pH e do nível da água. Entretanto, Resende
& Pereira, (2000), estudando ambientes sazonalmente inundados no Pantanal do
Mato Grosso do Sul, afirmam que a chegada da inundação é o fator desencadeador
da atividade reprodutiva.
Para Barbieri & Barbieri (1982, 1983, 1984) na Represa do Lobo (Estado de
São Paulo) a reprodução da tuvira ocorre de setembro a janeiro, com pico entre
outubro e dezembro, coincidindo com o início da época mais quente do ano. Os
mesmos autores observaram que para as fêmeas, o pico ocorre no mês de outubro
e, para os machos, nos meses de outubro a dezembro.
A reprodução da tuvira corresponde a um período de repouso intercalado por
períodos de atividade sexual os quais finalizam com o surgimento de nova prole.
Segundo Westby (1975) não é possível reconhecer macroscopicamente
diferenças sexuais na tuvira. Entretanto, alguns isqueiros da região do Pantanal
acreditam que tuvira fêmeas sejam mais amarronzadas e tuvira machos sejam mais
escuras, quase pretas.
A disposição das gônadas masculinas e femininas acha-se em um arranjo
semelhante ao tubo digestivo, pois os ovidutos e os dutos espermáticos dirigem-se
para a região cranial, terminando na papila genital que se encontra próximo ao
istmo, sob a cabeça, imediatamente posterior ao ânus (Figura 5) (González et al.,
2001).
12 Os machos, diferentemente das fêmeas, não apresentam alterações
morfológicas perceptíveis nos testículos que possibilitem identificar diferentes
estádios de maturação (Resende, 1999). A atividade espermatogênica dos machos
ocorre durante o ano todo, variando, porém, de intensidade, que é maior de
setembro a janeiro, com pico entre outubro e dezembro (Barbieri & Barbieri, 1982,
1983, 1984). A morfologia externa dos testículos da tuvira difere da descrita para a
maioria dos peixes teleósteos. Apresentam-se como órgãos pares, ovais,
consistentes (Figura 6) e com dois longos dutos espermáticos, ao contrário dos
testículos alongados que se estendem por todo o comprimento da cavidade
abdominal observado na maioria das espécies de teleósteos (Barbieri & Barbieri,
1984). Eles permanecem, independentemente de seu estádio de maturação,
próximos à extremidade caudal do conjunto visceral (Menin, 1989).
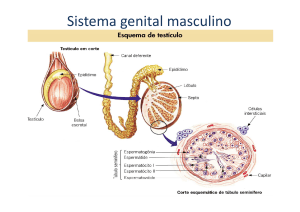
Figura 5. Localização dos testículos em Gymnotus carapo. Em A: seta indica o ânus
e a papila genital, em B: detalhe do duto deferente partindo do testículo até a papila
genital.
13 Figura 6. Testículos de Gymnotus carapo. Barra: 1cm
1.6. TESTÍCULOS E ESPERMATOGÊNESE EM PEIXES
Os testículos da maioria dos peixes são órgãos pares, assimétricos, podendo
estar parcial ou totalmente fundidos entre si e são frequentemente alongados,
embora existam outras formas como lobulados e foliáceos (Le Gac; Loir, 1999). Os
lóbulos testiculares são preenchidos por túbulos seminíferos os quais apresentam
paredes formadas por cistos que se apóiam na lâmina basal e que são revestidos
por prolongamentos citoplasmáticos das células de Sertoli (Matta, 2000).
A organização básica do testículo é comum a todos os peixes ósseos e
anfíbios. Este órgão tem as funções espermatogênica e androgênica, possuindo dois
compartimentos principais: intersticial e tubular. No compartimento intersticial ou
interlobular estão situados vasos sanguíneos, fibras nervosas, células e fibras do
conjuntivo, além das células de Leydig que possuem função esteroidogênica. A
produção de andrógenos é importante para diferenciação sexual, e para regulação
da espermatogênese (Weltizien et al., 2004). O compartimento tubular contém
células somáticas (células de Sertoli) e as células germinativas que irão formar os
espermatozóides, após passarem por um processo bastante complexo e altamente
organizado, a espermatogênese (Billard, 1990; Koulish et al., 2002) (Figura 7).
14 Figura 7. Espermatogênese em peixes. O epitélio germinal contém células de Sertoli
(SE) e células germinativas envolvidas por uma lâmina basal (LB) e células mióides
peritubulares (MI). No espaço intersticial são encontrados as células intersticiais ou
de Leydig (LE) e vasos sanguíneos (VS). A(Ind*): Espermatogônia tipo A
indiferenciada (célula-tronco); A(Ind): Espermatogônia tipo A indiferenciada; B (tardia):
Espermatogônia B tardia; Z/L: Espermatócitos primários em leptóteno e zigóteno; P:
Espermatócito primário em paquíteno; D/MI: Espermatócito primário em diplóteno /
metáfase;
S/MII:
Espermatócito
secundário/
metáfase
secundária
II;
E1:
Espermátide inicial; E2: Espermátide intermediária; E3: Espermátide final. (Adaptado
de Schulz et al., 2010).
Unidas entre si por complexos juncionais especializados, as células de Sertoli
dos teleósteos delimitam física e funcionalmente um clone de células germinativas
no mesmo estágio de desenvolvimento que tem origem a partir de uma única
espermatogônia
primária,
formando,
assim
os
espermatocistos
ou
cisto
espermatogênico. Desta forma, as células de Sertoli fornecem as células
germinativas suporte físico e fatores importantes para sobrevivência, proliferação e
diferenciação das mesmas, estando ainda envolvidas na intermediação hormonal e
na fagocitose de restos celulares originados da espermiogênese (Weltizien et al.,
2004). As células de Sertoli que delimitam os cistos espermatogênicos se apóiam na
15 túnica própria, que é constituída pela membrana basal (camada acelular) e pelas
células tubulares mióides que possuem capacidade contrátil (Pudney, 1995; Le Gac;
Loir, 1999). Portanto, diferentes dos mamíferos, em peixes as espermatogônias
diferenciadas (a partir da espermatogônia primária) não estão em contato com a
membrana basal (Billard, 1984).
Exceto pelo arranjo cístico, no qual as células germinativas se desenvolvem
de forma sincronizada, provavelmente devido à presença de pontes intercelulares
(Russell et al., 1990; Koulish et al., 2002), o processo espermatogênico de
teleósteos assemelha-se muito ao de mamíferos. Durante a fase proliferativa, a
espermatogônia tipo A se divide e origina a espermatogônia secundárias ou do tipo
B que, depois de um número de divisões mitóticas que varia entre as espécies,
originam os espermatócitos primários, iniciando a fase meiótica. Após a primeira
divisão meiótica formam-se os espermatócitos secundários haplóides que, num curto
intervalo de tempo, originam as espermátides através da segunda divisão meiótica.
Em seguida ocorre a fase de diferenciação, no qual as espermátides se transformam
em espermatozóides (Silva, 1987). As espermatogônias primárias são as maiores
células germinativas na maioria das espécies de teleósteos, apresentando um
núcleo grande e claro, com pouca heterocromatina, contendo um ou dois nucléolos
bastante evidentes (Miura, 1999). Após sucessivas divisões, a partir da
espermatogônia primária, o número de espermatogônias secundárias por cistos
aumenta geograficamente, enquanto o diâmetro nuclear das mesmas sofre gradual
redução (Vilela et al., 2006).
Considerando-se o número de espécies de peixes, existem poucos relatos na
literatura a respeito da duração dos eventos espermatogênicos nestes vertebrados.
No entanto, pode ser evidenciado que, de maneira geral, a duração da
espermatogênese em peixes é bem mais rápida do que o observado para mamíferos
(Silva; Godinho, 1983; Koulish et al., 2002; França et al., 2002; Vilela et al., 2003).
16 1.7. HISTOPATOLOGIA
As lesões detectadas em células, tecidos ou órgãos representam uma
integração dos efeitos cumulativos dos estressores em nível fisiológico e bioquímico
e potencialmente em níveis mais altos de organização biológica (Myers & Fournie,
2002). Além disso, em função de possibilitar a visualização dos locais dos danos, as
respostas histopatológicas dos peixes a várias classes de xenobióticos precisam ser
determinadas e caracterizadas, sendo um indicativo dos sítios de ação de tal
xenobiótico (Hinton et al., 1976).
A histopatologia é reconhecida como uma importante ferramenta para
avaliação do impacto ambiental de poluentes em peixes (Oliveira Ribeiro et al;
2002). É um método capaz de detectar efeitos adversos agudos e crônicos em
vários tecidos e órgãos de peixes mediante exposição a diversos compostos
químicos. Além disso, a histopatologia pode fornecer uma indicação da saúde dos
peixes, determinando lesão precoce de células (Pieterse, 2004).
Recentemente, vem ocorrendo o enriquecimento e a implementação da
literatura relacionada aos efeitos histopatológicos de estresse ambiental. Isto tem
aumentado os argumentos quanto ao uso destes dados como indicador de
adversidades, efeitos crônicos subletais e injúrias para vertebrados aquáticos
(especialmente peixes) em ambientes biomonitorados ou mesmo em experimentos
em laboratórios (Myers & Fornie, 2002).
17 1.8. EFEITO DOS METAIS PESADOS NOS TESTÍCULOS
O mercúrio e o cádmio são dois metais capazes de provocar diversas
alterações testiculares devido à sua toxicidade, essas alterações são observadas em
uma variedade de modelos animais (camundongos, coelhos e peixes) em diferentes
fases de crescimento e maturidade (Thompson & Bannigan, 2008).
Segundo Lockhart e Uthe, 1972; McKim et al., 1976; Hodson et al., 1994;
Pelletier e Audet, 1995) em concentrações baixas o mercúrio tem a capacidade de
acumulação nas gônadas de peixes das espécies (Esox lucius, Salvelinus fontinalis,
Anguilla rostrata, Myoxocephalus aenaus), onde inibe o desenvolvimento gonadal
(Dey e Bhattacharya, 1989; Kirubagaran e Joy, 1992), reduz o crescimento
(Panigrahi e Misra, 1978; Rodgers e Beamish, 1982; Snarski e Olson, 1982; Weis e
Khan, 1990; Niimi e Kissoon, 1994) e função imunológica (Roales e Perlmutter,
1977) que pode afetar a sobrevivência da prole.
Estudos têm mostrado que exposição aguda in vivo a cádmio causa o
rompimento da barreira hematotesticular, perda de células germinativas, edema
testicular, hemorragia, necrose e esterilidade em várias espécies de mamíferos
(roedores, coelhos, cães, bezerros e cavalos).
O cádmio também é capaz de provocar diversas lesões testiculares em
peixes de várias espécies. Embora os danos testiculares induzidos pelo cádmio
sejam reconhecidos por décadas atrás (Parizek, 1957), os mecanismos que levam a
esses danos ainda permanecem obscuros (Siu et al., 2009). A tabela 1 mostra um
resumo das principais alterações testiculares causadas por metais em diversas
espécies de peixes.
18 Tabela 1. Resumo das alterações histológicas nos testículos de peixes causados
por metais pesados (adaptado de Pieterse, 2004).
Metais pesados
Rota de
exposição
Espécies
Tecidos
afetados
CdCl2
Água
Brook trout
Testículo
CdCl2
Água
Salvelinus
tontinalis
Testículo
1ppb
Cádmio
Injeção
Intraperitoneal
Goldfish
Testículo
Chumbo e
Cádmio
Água
Clarias
batrachus
Testículo
HgCl2
Água
Clarias
batrachus
Testículo
Lesões no tecido
Hemorragia
Vasodilatação
Congestão
Necrose das
células de Leydig
Referência
Sangalang
and
O'Halloran,
1972
Sangalang
and Freeman,
1972
Baixo índice de
desenvolvimento
espermatogênico
Aumento do
número de
macrófagos
Tafanelli and
Necrose das
Summerfelt,
células
1975
germinativas com
atrofia dos túbulos
seminíferos
Infiltração de
células
inflamatórias
Fibrose
Estereidogênese
comprometida (Cd)
Diminuição da
Kalti and
reprodutibilidade
Sathyanesan,
(Pb)
1985
Inibição da
espermatogênese e
Danos testiculares
Inibição da
atividade gonadal
Picnose das células Kirubagaran
and Joy,
de Leydig
1992
Túbulos
seminíferos
menores
Obs: as setas indicam as alterações testiculares encontradas no presente estudo.
19 2. OBJETIVOS
2.1. OBJETIVO GERAL
Este trabalho tem como objetivo verificar se os metais pesados (Cloreto de
Mercúrio e Cloreto de Cádmio) provocam alterações nos testículos de Gymnotus
carapo (tuvira).
2.2. OBJETIVOS ESPECÍFICOS
- Descrever a estrutura do testículo e dos espermatozóides de Gymnotus carapo
(tuvira) através da microscopia óptica e eletrônica.
- Empregar a metodologia de contaminação in vitro e de contaminação aguda para
visualização de danos nos testículos.
- Descrever as alterações teciduais e celulares dos testículos e espermatozóides de
Gymnotus carapo (tuvira) expostos à ação do HgCl2 e CdCl2.
20 3. MATERIAL E MÉTODOS
3.1. COLETA DOS EXEMPLARES DE Gymnotus carapo.
Os exemplares utilizados neste trabalho foram coletados na Lagoa de Cima
(Figura 7), localizada na região norte do Estado do Rio de Janeiro (21º 46’ S e 41º
31’W), obtidos diretamente de pescadores locais. As amostras foram coletadas
durante o período de março de 2009 a junho de 2010. No presente estudo foram
utilizados 112 exemplares machos de Gymnotus carapo. Durante a coleta foram
considerados todos os peixes, e somente através da dissecação, realizada no
Laboratório de Ciências Ambientais (LCA) foi possível identificar machos e fêmeas.
Figura 8. Localização da Lagoa de Cima.
21 Os peixes coletados foram transportados vivos em caixas de isopor com água
obtida de seu habitat natural, e encaminhados para o Laboratório de Ciências
Ambientais (LCA), na UENF, onde permaneceram acondicionados num sistema de
aquários de fluxo contínuo (livre de cloro, por evaporação), com circulação constante
da água por um sistema de bombas com filtros internos. A qualidade da água foi
monitorada, medindo-se amônia tóxica, nitrito, pH através de kits colorimétricos
(Labcon Teste – Alcon). Os valores de pH estão de acordo com o banco de dados
publicados pelo FishBase para a espécie (FishBase, 2010). Durante este período, os
peixes foram mantidos a uma temperatura média de 25ºC.
3.2. PREPARAÇÃO DE SOLUÇÕES
Para o preparo das soluções de HgCl2 e CdCl2 utilizadas nesse estudo, uma
solução estoque com o sal desses metais foi preparada, diluindo-se o sal em água
destilada na concentração de 0,1M. Diluições da solução estoque foram em H2O
destilada nas concentrações finais de 5μM, 10μM, 20μM, 30μM e 40μM (Vergílio,
2009).
3.3. CONTAMINAÇÃO DOS EXEMPLARES DE Gymnotus carapo POR HgCl2 E
CdCl2.
Para cada experimento de contaminação por HgCl2 e CdCl2 foram
selecionados 6 exemplares machos, sendo que quatro foram expostos ao HgCl2 e
CdCl2 (um para cada tempo de exposição 24h/ 48h/ 72h/ 96h) e 2 restantes foram
estabelecidos como controle (dissecados após 96h). O procedimento descrito
anteriormente foi repetido duas vezes consecutivas para HgCl2 nas concentrações
5μM, 10μM, 20μM e 30μM, em cada tempo de exposição, totalizando 8 exemplares
utilizados para cada concentração testada e 16 controles.
O experimento de CdCl2 também foi repetido por duas vezes consecutivas
nas concentrações 10μM, 20μM, 30μM e 40 μM, em cada tempo de exposição,
totalizando 8 exemplares utilizados para cada concentração testada e 16 controles
22 (Tabela 2 e 3). A concentração testada para os dois metais (HgCl2 e CdCl2) foi
diferente, pois durante a fase de teste observamos que os peixes expostos a
concentrações
de
5μM
de
CdCl2 não
apresentaram
nenhuma
alteração
anatomopatológica, justificando assim a utilização de concentrações mais altas
desse metal no decorrer do experimento.
A contaminação foi realizada através de injeção intraperitonial da solução de
HgCl2 e CdCl2 , nos tempos de exposição de 24, 48, 72 e 96 horas. No grupo
controle, por sua vez, foi injetado solução de PBS (Phosphate Buffer Solucion). A
contaminação aguda foi documentada após uma única dose nas concentrações
5μM, 10μM, 20μM e 30μM e 40μM, sendo o volume injetado de acordo com o peso
do peixe (Vergílio, 2009).
Após cada tempo de exposição, os exemplares de Gymnotus carapo foram
anestesiados para obtenção dos seguintes dados: peso total (PT) e comprimento
total (CT), logo foram seccionados para a retirada dos testículos, que foram então
pesados para obtenção do índice gonadossomático (IGS), através da expressão:
IGS= Pg/PCX100. Os testículos retirados foram utilizados para a análise histológica
e ultraestrutural.
23 Tabela 2. Tabela de conversão das concentrações de HgCl2 em uM para ug/g.
HgCl2
ug
ug/g
5 μM:
135,7
1,04846
10 μM:
271,5
2,0884615
20 μM:
543
4,1769231
30 μM:
814,5
6,2653846
Tabela 3. Tabela de conversão das concentrações de CdCl2 em uM para ug/g.
CdCl2
ug
ug/g
10 μM:
201,3
1,54846
20 μM:
402,6
3,09692
30 μM:
604
4,64615
40 μM:
805
6,1923
24 Tabela 4. Número amostral, comprimento padrão e peso dos exemplares de G.
carapo utilizados para o experimento de HgCl2.
Número de exemplares
Comprimento (cm)
Peso (g)
controle
20
31
125
5 µM
8
33
100
10 µM
8
31
110
20 µM
12
33
153
30 µM
8
32
161
32
130
56
Total
Obs: No grupo controle foram utilizados 20 exemplares, 16 exemplares foram utilizados para
Histologia e 4 foram utilizados para Microscopia de Varredura. Na concentração de 20µM, 8
exemplares foram utilizados para Histologia e 4 exemplares foram utilizados para Microscopia de
Varredura, totalizando 12 exemplares.
Tabela 5. Número amostral, comprimento padrão e peso dos exemplares de G.
carapo utilizados para o experimento de CdCl2.
Número de exemplares
Comprimento (cm)
Peso (g)
controle
20
31
120
10 µM
8
30
116
20 µM
12
31
130
30 µM
8
35
135
40 µM
8
34
128
32
126
Total
56
Obs: No grupo controle foram utilizados 20 exemplares, 16 exemplares foram utilizados para
Histologia e 4 foram utilizados para Microscopia de Varredura. Na concentração de 20µM, 8
exemplares foram utilizados para Histologia e 4 exemplares foram utilizados para Microscopia de
Varredura, totalizando 12 exemplares.
25 3.4. PREPARAÇÃO PARA MICROSCOPIA ÓPTICA (MO)
Pequenas porções do testículo (aproximadamente 5 x 5 x 5 mm3) foram
fixados por meio da técnica de imersão com formol tamponado 10% por 24 horas.
As amostras foram então desidratadas em séries crescentes de álcool (70% / 80%/
90% / 95 % por uma hora e 100% por 30 minutos, três vezes consecutivas),
sofrendo diafanização em (1:1) xilol:álcool e xilol puro, em processador automático
(Leica TP 1020). Por fim, as amostras foram incluídas em parafina a 60° C e
cortadas com o auxílio de um micrótomo tipo Minot (Leica RM 2145) para obtenção
de cortes de 5 μm. Os cortes foram corados com hematoxilina-eosina (HE) e
observados
ao
microscópio
óptico
de
campo
claro
(Axioplan
–
Zeiss).
Microfotografias digitais foram obtidas através do microscópio Axioplan acoplado á
câmera Cannon Power Shot A610/620.
Adicionalmente, pequenas porções do testículo (aproximadamente 1,0 mm3)
foram fixados com tampão cacodilato de sódio 0,1M; formaldeído 4%; glutaraldeído
2,5%, sacarore 5%, cloreto de cálcio 5 mM e pós-fixadas com tetróxido de ósmio 1%
e ferricianeto de potássio 0,8%. Após sucessivas desidratações com acetona (50% /
70%/ 90%/ 100% comum / 100% comum / 100% super seca), as amostras foram
incluídas em resina Epóxi, para serem cortadas no ultra-micrótomo REICHERCUTS
LEICA em seções semi-finas de 0,4 μm e coradas com azul de toluidina 1% para
posterior observação em microscópio óptico de campo claro (ZEISS - AXIOPLAN).
26 3.5. PREPARAÇÃO PARA MICROSCOPIA ELETRÔNICA DE TRANSMISSÃO
(MET)
Pequenas porções do testículo (aproximadamente 1,0 mm3) fixados com
tampão cacodilato de sódio 0,1M; formaldeído 4%; glutaraldeído 2,5%, sacarose 5%,
cloreto de cálcio 5 mM e pós fixadas com tetróxido de ósmio 1% e ferricianeto de
potássio 0,8% seguiram o mesmo processo realizado para microscopia óptica citado
anteriormente.
Com
o
auxilio
de
um
ultramicrótomo
(Reichert
Ultracur
ultramicrotome) e faca de diamante foram obtidos cortes ultrafinos (70nm) que foram
contrastados com acetato de uranila 5% (30min) e citrato de chumbo 0,2% em
0.01mol/L NaOH (5 min) (Reynolds, 1963). Depois de lavadas, as grades secas
foram guardadas para posterior observação em um Microscópio Eletrônico de
Transmissão (ZEISS TEM 900), com uma aceleração de voltagem de 80 Kv.
3.6. PREPARAÇÃO PARA MICROSCOPIA ELETRÔNICA DE VARREDURA
(MEV).
Para o experimento de Microscopia de Varredura utilizamos espermatozóides
de peixes contaminados com a concentração de 20µM de HgCl2 e CdCl2 por 96h.
Essa concentração foi utilizada por ser considerada tóxica e por provocar alterações
nos testículos, visualizados através da histologia.
O fluido com espermatózoides dos peixes do grupo controle e do grupo
contaminado foram fixados com tampão cacodilato 0,1 M, formaldeído 4%,
glutaraldeído 2,5%, sacarose 5% e cloreto de cálcio. A amostra foi separada em
duas partes: uma foi centrifugada a 800 rpm por 15 min a 4 °C, o pellet foi lavado
duas vezes em tampão fosfato 0,1 M (pH 7,2) por 30 min. A outra amostra não foi
centrifugada. Testadas as duas amostras, utilizamos a amostra não centrifugada
para análise no microscópio óptico e no varredura.
Três gotas da amostra foram aderidas com poly L lisina em lamínulas, pós fixadas com tetróxido de ósmio 1% e tampão cacodilato de sódio 0,1M, desidratadas
em séries crescentes de etanol e secas em C02 pelo método de ponto crítico (BALTEC CPD 030 Critical Point Dryer), as amostras secas foram posicionados em um
suporte e cobertas com uma camada fina de ouro (BAL-TEC SCD 050 Sputter
Coater). A observação foi através de um Microscópio Eletrônico de Varredura (Zeiss
27 EVO 40 a 15Kv), utilizando elétrons secundários. Adicionalmente, o fluido seminal foi
colocado sobre a lâmina histológica, realizando assim um esfregaço celular. As
lâminas foram secas ao ar. Após esse processo, as células foram coradas com
solução de Giemsa (azul de metileno), por 30 minutos. As lâminas coradas foram
lavadas em H20 destilada e, após secas ao ar, a lamínula então foi aderida com
Bálsamo
do
Canadá
ao
esfregaço
celular.
Foram
obtidas
imagens
dos
espermatozóides devidamente fixados e corados para contagem e análise
morfológica das mesmas, através do microscópio Axioplan acoplado á câmera
Cannon Power Shot A610/620.
28 4. RESULTADOS
4.1.
EFEITO TÓXICO DO HgCl2 E CdCl2 NOS EXEMPLARES DE TUVIRA
A morte dos exemplares de Gymnotus carapo foi observada durante a
execução dos experimentos de contaminação in vitro, principalmente nas
concentrações mais elevadas de HgCl2 e CdCl2 (Tabela 6 e 7).
É importante
ressaltar que na ocorrência de morte de um dos espécimes de Gymnotus carapo,
outro exemplar era contaminado em seu lugar, respeitando a concentração e o
tempo de exposição da amostra, para não haver a redução do número amostral.
Tabela 6. Número de exemplares de G. carapo que morreram durante a exposição
de HgCl2.
5µM
10µM
20µM
30µM
Até 24h
0
0
3
1
Entre 24 e 48h
1
1
1
2
Entre 48h e 72h
0
2
2
2
Entre 72h e 96h
1
1
2
2
Total
2
4
8
7
Tabela 7. Número de exemplares de G. carapo que morreram durante a exposição
de CdCl2.
5µM
10µM
20µM
30µM
Até 24h
0
0
0
1
Entre 24 e 48h
0
1
1
1
Entre 48h e 72h
0
1
1
1
Entre 72h e 96h
0
1
2
2
Total
0
3
4
5
29 4.2. ASPECTOS ANATÔMICOS DO TESTÍCULO DE Gymnotus carapo
Uma investigação externa foi realizada em cada peixe antes e após a execução
dos experimentos. Os peixes do grupo controle demonstraram estar saudáveis, de
acordo com as condições macroscópicas de suas brânquias, olhos e escamas. Os
órgãos internos também foram investigados.
Nos peixes do grupo controle, os órgãos internos como fígado, rim e testículos
demonstraram estar normais. Os testículos apresentavam-se consistentes e com
uma coloração amarelada (Figura 9A e B). Já os peixes que foram expostos a
concentrações de 30µM e 40µM de CdCl2 nos tempos de exposição de 24 a 96
horas, foram observados alterações no fígado e no rim, esses órgãos apresentavam
um aspecto liquefeito, já os testículos apresentavam um aspecto amarelado (com a
presença de sangue) e flácidos (Figura 9C e D). Nas concentrações abaixo de 30µM
e 40µM de CdCl2 e em todas as concentrações de HgCl2 não foi visualizada
nenhuma alteração significativa nos órgãos internos.
A fim de avaliar a mudança testicular em resposta à exposição ao HgCl2 e
CdCl2, o índice gonadossomático foi calculado (Tabela 8 e 9), através disso
observamos que houve uma diminuição no GSI do grupo que foi exposto a 30µM e
40µM de CdCl2 (0,05), quando comparado ao GSI do grupo controle (0,09) (Tabela
8). Esses resultados confirmam as alterações anatômicas visualizadas no grupo
exposto a 30µM e 40µM de CdCl2, concluindo que a redução do GSI pode ser uma
resposta do testículo ao CdCl2.
O índice gonadossomatico também foi calculado no grupo exposto ao HgCl2 e
observamos que não houve uma diminuição significativa do GSI do grupo exposto a
concentrações de HgCl2 (0.07), quando comparado ao GSI do grupo controle (0,07 e
0,06) (Tabela 9).
30 Figura 9. Aspectos anatômicos do testículo de Gymnotus carapo. Em A-B:
testículos (espécime controle), C-D: testículos (espécime exposto à 30µM de CdCl2
por 96 horas).
Tabela 8. Número amostral, peso do testículo e o índice gonadossomático dos
exemplares de G. carapo expostos ao HgCl2
N° de
exemplares
Comprimento
(cm)
Peso do
testículo
GSI
Controle
20
33
100
0,070
0,07
5µM
8
31
110
0,081
0,07
10µM
8
33
153
0,110
0,07
20µM
12
32
161
0,120
0,07
30µM
8
32
131
0,080
0,06
Peso (g)
31 Tabela 9. Número amostral, peso do testículo e o índice gonadossomático dos
exemplares de G. carapo expostos ao CdCl2
N° de
exemplares
Comprimento
(cm)
Peso (g)
Peso do
testículo
Controle
20
30
116
0,109
0,09
10µM
8
31
130
0,080
0,06
20µM
12
35
135
0,089
0,06
30µM
8
34
128
0,069
0,05
40µM
8
32
127
0,071
0,05
GSI
4.3. ASPECTOS HISTOLÓGICOS
Para análise histológica dos testículos do grupo controle e do grupo exposto
ao HgCl2 e CdCl2 foram realizados cortes transversais (Figura 10). O testículo de G.
carapo mostrou-se arredondado, envolvido por uma camada de epitélio peritoneal
(Figura 11A).
Figura 10. Região do testículo de Gymnotus carapo (seta) onde foram realizados
cortes transversais. Barra:1cm
32 Os testículos de G. carapo são formados por um grande número de túbulos
seminíferos com diferentes diâmetros (Figura 11B), dentro desses túbulos foram
visualizadas as células de Sertoli (S) e células germinativas (espermatogônias,
espermatócitos, espermátides e espermatozóides) (Figura 11B e 11C). As células de
Sertoli (S) são pequenas e em maior aumento possuem o formato triangular, muitas
vezes localizada na periferia dos túbulos associadas às células germinativas,
principalmente as espermatogônias (SPG) (Figura 11E).
Entre os túbulos seminíferos identificamos o tecido intersticial onde
encontramos as células de Leydig ou células intersticiais e vasos sanguíneos (Figura
11B, 11D e 11F). As células de Leydig são ovóides e geralmente aparecem em
grupos (Figura 11D). No peixe teleósteo esta célula é responsável pela síntese de
11
cetotestosterona
(11KT),
um
hormônio
que
induz
a
progressão
da
espermatogênese (Billard, 1990)
Assim, verificamos que a estrutura interna dos testículos do grupo controle
mostrava-se compacta, sem espaçamento. Os túbulos seminíferos tinham um
arranjo normal e entre estes túbulos o tecido intersticial não apresentava nenhuma
alteração. As células germinativas aparecem normais e os espermatozóides livres na
luz do túbulo sem aparente agregação (Figura 11A - 11E, 15A e 15B).
Durante a análise histológica dos testículos do grupo controle e do grupo
exposto ao HgCl2 e CdCl2, observamos o estágio de maturação gonadal seguindo a
escala de maturidade de Barbieri para G. carapo (1981), com isso, verificamos que a
maioria dos peixes encontravam-se no estágio II (Adultos em reprodução -maduros),
neste estágio os testículos possuem túbulos seminíferos que exibem células
germinativas em várias fases de desenvolvimento, principalmente espermátides e
espermatozóides.
Várias alterações histológicas foram identificadas nos exemplares expostos
ao HgCl2 e CdCl2, porém no grupo tratado com HgCl2 os efeitos foram visualizados
mais rapidamente. Após o tratamento com 5µM de HgCl2 alterações histopatológicas
foram observadas a partir de 72 horas, enquanto que para 10μM, 20µM e 30μM
essas alterações foram visualizadas em todos os tempos de exposição analisados.
As lesões mais significativas identificadas no testículo dos espécimes expostos ao
HgCl2, incluem aumento do tecido intersticial caracterizado por fibrose (Figura 12A,
12C, 15C, 15D), congestão dos vasos sanguíneos ou hiperemia passiva (Figura
12B), desorganização dos túbulos seminíferos (Figura 12D e 13A) desintegração
33 intersticial e tubular caracterizado por áreas com espaçamento (Figura 13B e 13D) e
variações acentuadas no tamanho dos túbulos seminíferos (Figura 13C).
Com relação ao grupo exposto ao CdCl2, às alterações histopatológicas foram
observadas a partir de 72h de exposição na concentração de 10µM. É importante
ressaltar que as lesões encontradas no tecido reprodutor do grupo exposto ao HgCl2
e CdCl2 foram semelhantes em todas as concentrações administradas, sendo os
efeitos progressivos observados com o aumento da concentração ao metal exposto.
As lesões mais significativas identificadas no testículo dos espécimes expostos ao
CdCl2, incluem proliferação do tecido intersticial entre os túbulos seminíferos (Figura
14A), fibrose (Figura 14B, 14F e 15E), congestão dos vasos sanguíneos ou
hiperemia passiva (Figura 14C), desorganização dos túbulos (Figura 14D),
degeneração do tecido, caracterizado por um processo de vacuolização (Figura 14E,
14F e 15F), agregação espermática na luz dos túbulos (Figura 14E).
34 Figura 11. Organização estrutural do testículo de Gymnotus carapo (grupo controle).
Em A: destaca-se a túnica albugínea envolvendo o testículo (seta) e o lúmen do
túbulo, B: as setas indicam os túbulos seminíferos. Inset: túbulos seminíferos, C:
destacam-se
as
células
germinativas:
espermatogônias,
espermatócitos,
espermátides, espermatozóides e células de Sertoli (seta). Inset: Célula de Sertoli.
Em D as setas indicam células sanguíneas e células de Leydig. Inset: célula de
Leydig, F: destaca-se o tecido intersticial entre túbulos seminíferos. Inset: tecido
intersticial. TA: Túnica Albugínea, LI: Lúmen do túbulo, it: tecido intersticial, L: célula
35 de Leydig, S: célula de Sertoli, SZ: espermatozóides, SCI: espermatócitos primários,
SD: espermátides, SGI: espermatogônias primárias. Coloração: Hematoxilina e
Eosina. Barra de Escala: A: 200µm, B: 100µm, C-E: 50µm.
Figura 12. Organização estrutural do testículo de Gymnotus carapo (grupo exposto
a 5μM e 10μM de HgCl2). Em A e C: as setas indicam fibrose. Insets: regiões do
testículo com fibrose, B: a seta indica hiperemia passiva, D: desorganização dos
túbulos seminíferos. Inset: região do testículo com desorganização dos túbulos.
Coloração: Hematoxilina e Eosina. Barra de escala: A-D: 100µm.
36 Figura 13. Organização estrutural do testículo de Gymnotus carapo (grupo exposto
a 20μM e 30μM de HgCl2). Em A: desorganização dos túbulos seminíferos, B e D:
desintegração intersticial e tubular. Insets: região do testículo com o tecido
intersticial e tubular desintegrado. Em C: variações acentuadas no tamanho dos
túbulos seminíferos (setas). Coloração: Hematoxilina e Eosina. Barra de escala: A-D:
100µm.
37 Figura 14. Organização estrutural do testículo de Gymnotus carapo (grupo exposto
a 10µM, 30µM e 40µM CdCl2). Em A: as marcações indicam uma proliferação
intersticial entre os túbulos seminíferos. Inset: região entre túbulos com proliferação,
B: fibrose (seta). Inset: região do tecido com fibrose, C: hiperemia passiva (seta), D:
desorganização dos túbulos, E: agregação espermática (seta) e degeneração do
tecido. Inset: detalhe dos espermatozóides em agregação no lúmen do túbulo, F:
degeneração do tecido acompanhada de fibrose. Inset: região do tecido degenerada
e com fibrose. Coloração: Hematoxilina e Eosina. Barra de escala: A-F: 100µm.
38 Figura 15. Corte transversal dos testículos de Gymnotus carapo. Em A e B, detalhe
do tecido intersticial e células germinativas (grupo controle), em C e D as marcações
indicam o aumento de tecido intersticial entre túbulos seminíferos (grupo exposto a
HgCl2). Em E destaca-se o aumento do tecido intersticial e desorganização nos
túbulos, em F as setas mostram vacúolos entre as células germinativas indicando
uma degeneração do tecido (grupo exposto a CdCl2). Coloração: Azul de Toluidina.
Barra de escala: A-F: 20µm.
39 4.4. ASPECTOS ULTRAESTRUTURAIS
As observações ultraestruturais revelam que as espermatogônias primárias
(SPGI), que são as maiores células da linhagem germinativa, possuem o formato
arredondado, um núcleo esférico e central com cromatina granular e uniformemente
distribuída. O nucléolo proeminente é esférico e eletrodenso (Figura 16A). As
espermatogônias primárias se dividem mitoticamente para dar origem as
espermatogônias secundárias. As espermatogônias secundárias (SPCII) são
caracterizadas por uma diminuição no tamanho da célula, o núcleo é central e
agregação da cromatina difere segundo a fase do ciclo celular observada (Figura
16A).
Nos
túbulos
seminíferos,
também
podemos
observar
espermatócitos
primários (SPCI), que se originam de espermatogônias secundárias tardias e são
vistas em várias fases de desenvolvimento meiótico. Essas células possuem um
núcleo central esférico, com um formato regular, ocupando uma grande parte da
célula, o nucléolo não é visível e a presença de diferentes graus de condensação da
cromatina no núcleo revela que essas células estão em diferentes estágios da
prófase meiótica (Figura 16B). Os espermatócitos primários se transformam em
espermatócitos secundários, que são células menores que os espermatócitos
primários. Cistos de espermatócitos secundários são raramente observados.
Diversas espermátides (SPD) foram visualizadas na luz dos túbulos
seminíferos em diferentes estágios de espermiogênese (Figura 16B e C). Com o
avanço do processo de espermiogênese, as espermátides tornam-se células
menores e o núcleo ocupa grande parte da célula (Figura 16B e C). Ao mesmo
tempo, uma migração dos centríolos ocorre em direção à espermátide. Dependendo
do estágio da espermiogênese, o volume de citoplasma diminui consideravelmente e
ocorre um aumento no volume de mitocôndrias (Figura 16C).
A análise ultraestrutural também revelou alterações em células da linhagem
espermatogênica e essas alterações foram mais evidentes com o aumento da
concentração e tempo de exposição do organismo ao HgCl2 e CdCl2.
No grupo tratado com 20µM de HgCl2 por 72h, observamos na luz dos túbulos
seminíferos várias espermátides, que não possuíam formato arredondado e
apresentavam-se deformadas, com núcleo irregular e mitocôndrias maiores (Figura
17A). No grupo tratado com 30µM de HgCl2 por 96h, foram observadas alterações
40 mais severas, já que em algumas células germinativas não foi possível identificar o
seu estágio de desenvolvimento espermatogênico pois estavam totalmente
danificadas e algumas já se encontravam em processo apoptótico (Figura 17B).
Nos testículos dos espécimes tratados com 30µM e 40µM de CdCl2 por 96h e
24h respectivamente, observamos várias células germinativas em processo de
vacuolização (Figura 17C-17D).
Figura 16. Organização ultraestrutural do testículo de Gymnotus carapo (grupo
controle). Em A destacam-se as espermatogônias, B: espermatócitos primários e
espermátides C: lúmen do túbulo com espermátides, D: espermatozóide. SPGI:
espermatogônia primária, SPGII: espermatogônia secundária, SPCI: espermatócitos
primários SPD: espermátides, SZ: espermatozóide N: núcleo, n: nucléolo, c:
centríolo, Ll: Lúmen do lóbulo, m: mitocôndria, f: flagelo.
41 Figura 17. Organização ultraestrutural do testículo de Gymnotus carapo (grupo
exposto a HgCl2 e CdCl2). Em A, a seta indica uma espermátide com várias
mitocôndrias com o seu tamanho anormal, B: células germinativas em apoptose
(cabeça da seta) e restos celulares (seta), C: célula germinativa vacuolizada (seta),
D: célula germinativa danificada com pequenos vacúolos. n: núcleo, m: mitocôndria,
v: vacúolos.
42 4.4. ASPECTOS MORFOLÓGICOS DOS ESPERMATOZÓIDES DE Gymnotus
carapo.
Os espermatozóides de G. carapo são as menores células da linhagem
espermatogênica e são constituídos de uma cabeça arredondada (com ausência de
acrossoma), um flagelo alongado e uma peça intermediária (que não é evidente
nesta espécie de peixe) (Figura 16D, 18A e 19A e B).
Durante a análise morfológica dos espermatozóides do grupo controle foi
observado um núcleo arredondado e uma pequena parte do citoplasma evidente na
região ântero-posterior da cabeça (Figura 18A).
Para análise da morfologia dos espermatozóides do grupo exposto ao HgCl2 e
CdCl2 foi testado apenas uma concentração (20µM) nos tempos (24, 48, 72 e 96h).
No grupo exposto a 20µM de HgCl2 e CdCl2 por 24h e 48h não foi observado
nenhuma alteração significativa nos espermatozóides, já após 72h foram
observados espermatozóides com a cabeça totalmente corada indicando uma
máxima condensação do DNA (Figura 18B), além disso, verificamos por meio da
microscopia de varredura, espermatozóides apresentando cabeças deformadas e
com ausência de flagelo (Figura 19C-19F).
Através da análise quantitativa para verificação da presença de flagelo,
observamos que no grupo controle aproximadamente 90% dos espermatozóides
apresentavam flagelo. No grupo exposto a 20µM de HgCl2 (24h/72h) houve uma
diminuição
do
número
de
espermatozóides
com
flagelo
(54%
e
62%
respectivamente, Figura 20A). No grupo exposto a 20µM de CdCl2 (24h/ 96h)
também foi observado a diminuição do número de espermatozóides com flagelo (63
e 62% respectivamente, Figura 20B).
O cloreto de mercúrio e o cloreto de cádmio apresentaram efeitos similares na
morfologia dos espermatozóides expostos a 20µM, entretanto, o HgCl2 aparentou
ser mais tóxico do que o CdCl2. De acordo com a Figura 20, pode-se observar que o
percentual de espermatozóides com flagelo não variou muito em função do tempo
de exposição ao CdCl2 e HgCl2, porém, realizado um comparativo entre os metais,
foi revelado que a porcentagem do número de espermatozóides com flagelo foi o
mesmo (62%) para o HgCl2 e o CdCl2 nos tempos (72h e 96h respectivamente).
43 Figura 18. Fotomicrografia dos espermatozóides de Gymnotus carapo corados com
Giemsa. A: espermatozóides do grupo controle, B: espermatozóides do grupo
exposto a 20μM de HgCl2 por 72h, C: espermatozóides do grupo exposto a 20µM de
CdCl2 por 96h c: cabeça, f: flagelo. Barra de escala: A-C: 20µm.
44 Figura 19. Fotomicrografia (MEV) dos espermatozóides de Gymnotus carapo. Em A
e B: espermatozóide do grupo controle, C: espermatozóide do grupo exposto a
HgCl2 por 72h D: espermatozóides do grupo exposto a CdCl2 por 96h. c: cabeça e f:
flagelo. Barra de escala: A: 2μm, B, D e E: 1μm.
45 Figura 20. Porcentagem de espermatozóides do grupo controle e do grupo exposto
ao HgCl2 (A) e ao CdCl2 (B) com flagelo e sem flagelo.
46 5. DISCUSSÃO
Os bioensaios em laboratório têm como objetivo analisar a toxicidade dos metais
pesados em concentrações específicas e com isso as mudanças induzidas por
esses elementos podem ser empregadas para observação da relação dose/resposta
dos organismos expostos (Vergílio, 2009). Além disso, a análise histopatológica vêm
como uma ferramenta essencial para diagnosticar e fornecer uma descrição exata
das lesões causadas por metais em tecidos e células. Assim, torna-se importante
detectar estas alterações histopatológicas antes dos efeitos prejudiciais tornarem-se
visíveis nos níveis mais altos, como o nível de organismo ou população.
Os organismos, incluindo os peixes, podem responder de diferentes formas a
exposição ao Hg e Cd. Essa resposta irá depender tanto da dose quanto do tempo
de exposição a esses compostos químicos. Com isso, em alguns casos, o
organismo pode sofrer adaptação quando for exposto continuamente a um agente
químico. Porém, em outros casos, o composto pode induzir respostas, sendo
observadas principalmente através de danos celulares e teciduais. Finalmente, se a
exposição for quantitativamente além do limite de detoxificação dos organismos, ou
ocorrer por um período prolongado, se estabelece uma resposta tóxica que pode
ocasionar a mortalidade dos organismos susceptíveis (Boelsterli, 2007).
O testículo de peixe é um órgão pouco estudado, pois não é considerado alvo
direto do mercúrio e cádmio, porém nos últimos anos, estudos têm demonstrado que
o testículo vem sofrendo diversas alterações devido à exposição a esses poluentes
e com isso tem se mostrado importante nas avaliações de toxicidade ambiental
(Thompson & Bannigan, 2008; Siu et al., 2009).
Com base em estudos anteriores do grupo, onde foram utilizados em cultura de
células (hepatócitos HUH-7) e no tecido hepático de G. carapo várias concentrações
de HgCl2 em diferentes tempos de exposição, decidimos ajustar algumas dessas
concentrações para visualização de efeitos tóxicos nos testículos. No presente
estudo, a investigação das respostas celulares e sub-celulares por microscopia de
luz e microscopia eletrônica foram fundamentais para descrever as alterações
celulares precoces e danos testiculares nos peixes expostos a diferentes
concentrações de cloreto de mercúrio e cádmio.
Kirubagaran e Joy (1988) e Pelgrom et al. (1995) observaram que o mercúrio
e o cádmio têm a capacidade de inibir a gametogênese, induzindo lesões
47 testiculares (desorganização dos túbulos e desintegração das células de Sertoli). No
presente estudo, a desorganização dos túbulos seminíferos foi claramente
observada após 96h de exposição a 10μM de HgCl2. Além disso, foi observado
também outro tipo de lesão no tecido testicular como a desintegração dos túbulos e
do tecido intersticial após 72h de exposição a 20µM de HgCl2. Alterações similares
também foram observadas nos peixes expostos ao cádmio.
Adicionalmente, em estudo de contaminação subletal por cobre em O.
mossambicus (0.16; 0.40 e 2.00 mg Cu/L) por 96 horas, Pieterse (2004), também
observou
no
tecido
testicular
desorganização
dos
túbulos
seminíferos
e
desintegração do tecido intersticial e das células de Sertoli. Para o autor, essas
alterações degenerativas que ocorrem nos túbulos podem comprometer o processo
espermatogênico, fazendo com que haja uma mudança ou até mesmo uma
interrupção desse processo.
Segundo Wester e Canton (1992), testículos maduros de P. reticulata
expostos
a
5.6µg
MeHg/L
durante
três
meses
demonstraram
alterações
histopatológicas incluindo a ocorrência de fibrose e inflamação no interstício. A
fibrose ocorre quando o tecido conjuntivo fibroso em excesso se desenvolve no
tecido testicular como um processo reparativo. Em nosso estudo a fibrose foi
observada nos testículos dos espécimes expostos a 5µM - 30µM de HgCl2 e a 10µM
- 40µM de CdCl2 nos maiores tempos de exposição, indicando que esse processo
ocorre como um tipo de defesa do tecido testicular contra os danos induzidos pelos
metais.
Além disso, verificamos que, nos testículos de espécimes expostos a 10µM
HgCl2 e a 30µM de CdCl2 houve um aumento das células sanguíneas nos vasos,
esse fenômeno é conhecido por congestão ou hiperemia. A hiperemia é causada
pelo aumento no volume de sangue em uma região pela intensificação do
abastecimento de sangue ou de uma diminuição do fluxo venoso. É um indício de
que está havendo uma inflamação aguda local. No presente estudo, a ocorrência de
hiperemia pode estar relacionada com o tempo de exposição ao HgCl2 e ao CdCl2, já
que esse evento foi documentado a partir de 24 horas nas concentrações descritas
acima.
Durante o andamento dos experimentos foi observada a morte de alguns
exemplares de G. carapo (Tabela 4 e 5), principalmente quando ambos estavam
expostos as maiores concentrações de HgCl2 e CdCl2. Isso indica que o tratamento
48 foi estabelecido desde concentrações sub-letais (5µM e 10µM) onde puderam ser
observados os efeitos progressivos da contaminação por HgCl2 e CdCl2, até
concentrações letais (20µM, 30µM e 40µM) onde a concentração foi num nível tal
que proporcionou a morte dos peixes analisados.
Em um estudo realizado por Predes et al. (2010), onde foram injetados
intraperitonealmente baixas doses de CdCl2 em ratos (1 e 1.2 mg/kg) foi observado
através da análise morfométrica uma diminuição no diâmetro dos túbulos
seminíferos após 7 dias de exposição, além disso, observações microscópicas
mostraram danos progressivos no tecido testicular de animais expostos ao CdCl2.
Após sete dias de exposição, o lúmen dos túbulos seminíferos foram preenchidos
com células germinativas degeneradas e agregado de espermátides com o núcleo
apoptótico. Vacuolização do epitélio seminífero também foi observado. Após 56 dias,
houve um aumento dos danos, resultando em túbulos seminíferos vacuolizados. De
acordo com Creasy (2001), a vacuolização é uma resposta morfológica comum do
epitélio testicular a vários tipos de agressões causadas por metais.
No presente estudo, os túbulos seminíferos apresentaram variações
acentuadas no tamanho após 72h de exposição a 30µM de HgCl2. Alterações como
degeneração, agregado de espermátides e espermatozóides e vacuolização do
epitélio e do túbulo seminífero também foram observados nos espécimes expostos a
40µM de CdCl2. Assim como o estudo citado acima, o presente estudo mostra que os
efeitos tóxicos do cádmio no aparelho reprodutor masculino foram dose-dependente,
as alterações mais drásticas foram visualizadas no tecido testicular de G. carapo na
concentração de 40µM de CdCl2.
Cameron e Foster (1963) demonstraram em um estudo realizado com
coelhos, que através de uma única injeção subcutânea de cloreto de cádmio
(18mg/kg) após 72h de exposição, foi possível observar nos testículos vários efeitos
histopatológicos, dentre eles destacam-se: desorganização do epitélio seminífero,
interstício distendido com extravasamento de células sanguíneas e aumento da área
intersticial. Para esses autores, as principais linhas de evidência para explicar o
efeito destrutivo do cádmio no tecido testicular são: (a) que inibe as enzimas sulfidril,
formando mercaptides com seus grupos SH, (b) que se combina com grupos
similares SH no aparato mitótico (c) que compete com o zinco. A última explicação é
a mais provável porque as injeções de zinco dão proteção contra danos induzidos
pelo cádmio, embora apenas por um período limitado.
49 No presente estudo nós observamos no tecido testicular de G. carapo o
interstício distendido após 72h de exposição a 10µM de CdCl2, além disso,
verificamos um aumento da área intersticial no tecido testicular dos exemplares
expostos a 20µM de HgCl2 (24 e 72h) e 30µM de CdCl2 (96h).
Homma - Takeda et al. (2001) mostraram a indução de apoptose em células
germinativas de camundongos machos que haviam sido expostos ao cloreto de
metilmercúrio (10 mg/kg por 8 dias). Após a contaminação, eles observaram o
fenômeno apoptótico principalmente em espermatócitos e espermátides. Apoptose é
um processo que está geralmente envolvido no desenvolvimento e homeostase
tecidual, mas também parece estar envolvido numa ampla faixa de condições
patológicas (Thompson, 1995). Muitos autores mostraram a formação de ROS na
toxicidade de metais pesados dentro das células, sendo proposto que estariam
envolvidos na citotoxicidade e apoptose (Bower, 2005; Lemarie et al., 2004).
As células que sofrem apoptose encolhem morfologicamente devido à perda
de volume citoplasmático, além disso, ocorre condensação da cromatina e restos
celulares de organismos apoptóticos ficam evidentes. Assim, a apoptose, é um
indicador precoce de estresse a xenobióticos e fornece dados sobre o estado de
saúde do organismo (Sweet et al., 1999).
Através de uma análise ultraestrutural, podemos observar em nosso estudo
que o testículo do grupo contaminado com 20µM de HgCl2 por 72h apresentou a
formação de espermátides deformadas com a presença de núcleos irregulares e
mitocôndrias maiores. No grupo tratado com 30µM de HgCl2 por 96h foram
observados alterações mais severas, já que em algumas células germinativas não
foi possível identificar o seu estágio de desenvolvimento espermatogênico, pois
estavam totalmente danificadas e algumas espermátides já se encontravam em
processo apoptótico.
A morfologia dos espermatozóides de peixes tem sido pouco investigada,
porém estudos com mamíferos sugerem que baixas concentrações de mercúrio
podem causar danos no flagelo (Mohamed et al., 1986; Rao, 1989)
Em estudos realizados por Popek, et al. (2006), Dietrich et al. (2004 e 2005),
Rurangwa, et al. (1998) onde foram testados em peixes várias concentrações de
mercúrio e cádmio (1, 10, 100 mg/l) em um curto período de tempo (0-24h), concluiuse que esses metais tem a capacidade de diminuir a qualidade dos gametas,
50 afetando
a
motilidade
do
espermatozóide,
integridade
do
plasmalema
e
fragmentação do DNA.
Kime, (1999) em uma revisão sobre os efeitos dos xenobióticos na
reprodução de peixes revela que a maioria das substâncias químicas presentes no
ecossistema aquático exerce efeitos nos testículos durante a espermatogênese o
que resulta em espermatozóides anormais, malformados e com motilidade reduzida.
Segundo este autor, a morfologia espermática pode, portanto fornecer uma boa
medida quantitativa do desenvolvimento dos efeitos aos xenobióticos.
No presente estudo, observamos através da microscopia óptica e eletrônica
alterações nos espermatozóides de G. carapo expostos ao HgCl2 e CdCl2. Essas
alterações incluem anormalidade na cabeça, ausência de flagelo e máxima
condensação do DNA.
É importante ressaltar, que as lesões encontradas no tecido reprodutor do
grupo exposto ao HgCl2 e CdCl2 foram semelhantes em todas as concentrações
administradas, sendo os efeitos progressivos observados com o aumento da
concentração ao metal exposto, porém no grupo tratado com HgCl2 os efeitos foram
considerados mais severos. Após 72h de exposição a 5µM de HgCl2 foi observado
alterações no tecido reprodutivo. Já o CdCl2 as alterações foram visualizadas após
72 horas de exposição a 10µM. Isso indica que o HgCl2 foi mais tóxico no tecido
reprodutor que o CdCl2.
A ocorrência de alterações histopatológicas documentadas no presente
estudo
sugerem
uma
disfunção
reprodutiva
e
uma
falha
no
processo
espermatogênico no testículo de Gymnotus carapo após exposição ao HgCl2 e
CdCl2. Dessa forma, os resultados obtidos através da análise dos espermatozóides
e da histologia do tecido reprodutivo elucidam as alterações a nível celular e tecidual
induzidas pela exposição aos metais pesados, podendo ser empregadas como
ferramentas sensíveis no monitoramento ambiental e programas de avaliação de
áreas impactadas, apresentado assim a habilidade de complementar as análises
químicas.
51 6. CONCLUSÕES
Através de análise microscópica o testículo de Gymnotus carapo mostrou-se
arredondado e envolvido por uma camada de epitélio peritoneal. Internamente os
testículos são formados por um grande número de túbulos seminíferos com
diferentes diâmetros, dentro desses túbulos foram visualizados as células de Sertoli
(S) e células germinativas (espermatogônias, espermatócitos, espermátides e
espermatozóides). As células de Sertoli (S) são pequenas e em maior aumento
possuem o formato triangular, muitas vezes localizadas na periferia dos túbulos
associadas a células germinativas, principalmente as espermatogônias (SPG). Entre
os túbulos seminíferos identificamos o tecido intersticial onde encontramos as
células de Leydig ou células intersticiais e vasos sanguíneos. As células de Leydig
são ovóides e geralmente aparecem em grupos.
Os espermatozóides de G. carapo são constituídos de uma cabeça
arredondada (com ausência de acrossoma), um flagelo alongado e uma peça
intermediária (que não é evidente nesta espécie de peixe).
A metodologia de contaminação in vitro e de contaminação aguda empregada
no presente estudo foi realizada com sucesso. A espécie de peixe utilizada
(Gymnotus carapo) se adaptou bem as condições do laboratório.
Após testadas várias concentrações (5µM, 10µM, 20µM, 30µM, 40µM) de
HgCl2 e CdCl2 em diferentes tempos de exposição (24h, 48h, 72h, 96h) conclui-se
que tanto o HgCl2 e CdCl2 afetam o testículo de Gymnotus carapo, provocando
diversas alterações, tanto teciduais quanto celulares, porém neste estudo o HgCl2 foi
mais tóxico que o CdCl2. Os efeitos nos testículos foram visíveis a partir de 5µM para
HgCl2 e 10µM para CdCl2. As alterações nos testículos e espermatozóides foram
semelhantes para ambos os metais e progressivas com o aumento da concentração
e do tempo de exposição.
52
As lesões mais significativas identificadas no testículo dos espécimes
expostos ao HgCl2, incluem aumento do tecido intersticial caracterizado por fibrose,
hiperemia, desorganização dos túbulos seminíferos, desintegração intersticial e
tubular caracterizado por áreas com espaçamento e variações acentuadas no
tamanho dos túbulos seminíferos, já as lesões identificadas no testículo dos
espécimes expostos ao CdCl2, incluem proliferação do tecido intersticial entre os
túbulos seminíferos, fibrose, hiperemia, desorganização dos túbulos seminíferos,
degeneração do tecido caracterizado por um processo de vacuolização, agregação
espermática na luz dos túbulos.
Para análise da morfologia dos espermatozóides do grupo exposto ao HgCl2 e
CdCl2 foi testado apenas uma concentração (20µM) nos tempos (24, 48, 72 e 96h).
No grupo exposto a 20µM de HgCl2
e CdCl2 após 72h foram observados
espermatozóides com a cabeça totalmente corada indicando uma máxima
condensação do DNA, além disso, verificamos por meio da microscopia de
varredura, espermatozóides apresentando cabeças deformadas e com ausência de
flagelo.
53 7. REFERÊNCIAS BIBLIOGRAFICAS
ABEL, P. D. (1989) Water pollution biology. Chichester: Ellis Howood. 328p.
AZEVEDO, F. A. (2003) Toxicologia do mercúrio. Editora Rima, São Carlos, 272p.
BAATRUP, E. (1991) Structural and functional effects of heavy metals on the
nervous system, including sense organs, of fish. Comparative Biochemical
Physiology; 100: 253-257.
BARBIERI, G.; BARBIERI, C. B. (1982) Fecundidade e tipo de desova de Gymnotus
carapo (Linnaeus, 1758) na represa do Lobo, Estado de São Paulo (Pisces,
Gymnotidae). Spectrum (J. Bras. Ci.). 2:25-29.
BARBIERI, G.; BARBIERI, C. B. (1983) Dinâmica da reprodução de Gymnotus
carapo na represa do Lobo, estado de São Paulo. Influência de fatores abióticos.
(Pisces, Gymnotidae). Tropical Ecology. 24:244-259.
BARBIERI, G.; BARBIERI, C. B. (1984) Note on nutritional dynamics of Gymnotus
carapo (L.) from the Lobo reservoir, São Paulo State, Brazil. Jornal of Fecha Bióloga.
24:351-355.
BARAK, N. A. E; MASON, C.F. (1990) Mercury, cadmium and lead concentrations in
five species of freshwater fish from eastern England. The Science of the Total
Environment. 92: 257-263.
BASTOS, W. R.; GOMES, J. P. O.; OLIVEIRA, R. C.; ALMEIDA, R.; NASCIMENTO,
E. L.; BERNARDI, J. V. E.; LACERDA, L. D.; SILVEIRA, E. G.; PFEIFFER, W. C.
(2006) Mercury in the environment and riverside population in the Madeira River
Basin, Amazon, Brazil. Science of the Total Environment. 368:344-351.
BEYERSMANN, D; HECHTENBERG, S (1997) Cadmium, gene regulation, and
cellular signalling in mammalian cells. Toxicology and applied pharmacology.
144(2):247-61.
54 BEYERSMANN, D., HARTWIG, A. (2008) Carcinogenic metal compounds: recent
insight into molecular and cellular mechanisms. Arch. Toxicol. 82:493-512.
BILLARD,
R.
(1984)
Ultrastructural
changes
in
the
spermatogonia
and
spermatocytes of Poecilia reticulata during spermatogenesis. Cell Tiss. Res.
237:219-226.
BILLARD, R. (1990) Spermatogenesis in teleost fish. In: LAMMING, G.E. (ed)
Marshall´s physiology of reproduction, v.2. Reproduction in males. Edinburgh,
Churchill Livingston. 183-212p.
BOELSTERLI, U. A. (2007) Mechanistic toxicology: the molecular basis of how
chemicals disrupt biological targets, Taylor and Francis, London, 338p.
BOWER, J. J.; LEONARD, S. S.; SHI, X. (2005) Conference overview: Molecular
mechanisms of metal toxicity and carcinogenesis. Molecular and Cellular
Biochemistry. 279:3-15.
BUCIO, L.; SOUZA, V.; ALBORES, A.; SIERRA, A.; CHÁVEZ, E.; CÁRABEZ, A.;
GUTIERREZ-RUIZ, M. C. (1995) Cadmium and mercury toxicity in a human fetal
hepatic cell line (WRL-68 cells). Toxicology. 102:285-299.
BUCIO, L.; GARCIA, C.; SOUZA, V.; HERNANDEZ, E.; GONZALEZ, C.;
BETANCOURT, M.; GUTIERREZ-RUIZ, M. C. (1999) Uptake, cellular distribution
and DNA damage produced by mercuric chloride in a human fetal hepatic cell line.
Mutation Research. 423:65-72.
BURGER, J. (2008) Assessment and management of risk to wildlife from cadmium.
Science of the Total Environment. 389:37-45.
BURGER, J.; GOCHFELD, M. (2001) On developing bioindicators for human and
ecological health. Environmetal Moniring Assess. 66:23-46.
55 CAMERON, E.; FOSTER, L.C. (1963) Observations on the histological effects of sublethal doses of cadmium chloride in the rabbit I. The effect on the testis. J. Anat.
Lond. 97:269-280.
CANELA, M. C. (1995) “Determinação de Mercúrio”, UNICAMP.
CARIGNAN, V.; VILLARD, M. A. (2001) Selecting indicator species to monitor
ecological integrity: a review. Environ. Monit. Assess. 78:45-61.
CONELL, D. W.; MILLER, G. J. (1984) Chemistry and ecotoxicology of pollution.
John Willey & Sons. 444p.
CREASY, D. M. (2001) Pathogenesis of male reproductive toxicity. Toxicol. Pathol.
29:64-76.
DEY, S.; BHATTACHARYA, S. (1989) Ovarian damage to Channa punctatus after
chronic exposure to low concentrations of Elsan, mercury, and ammonia. Ecotoxicol.
Environ. Safety. 17(2):247-257.
DIETRICH, G.; KOWALSKI, R.; WOJTCZAK, M.; DOBOSZ, S.; GORYCZKO, K.;
DEMIANOWICZ,
W.;
CIERESZKO,
A.;
GLOGOWSKI,
J.
(2004)
Motility
characteristics and fertilizing capacity of rainbow trout sperm (Oncorhynchus mykiss
Walbaum) after short-term exposure of milt to mercury and cadmium. (in Polish). In:
Trout culture. Law, health and quality issues. pp 57-65. Ed K Goryczko. Inland
Fisheries Institute, Poland.
DIETRICH, G.; DEMIANOWICZ, W., KOWALSKI, R.; WOJTCZAK, M.; CIERESZKO,
A.; GLOGOWSKI, J. (2005) Fertilizing capacity of cryopreserved sperm of rainbow
trout after short-time exposure of milt to mercury and cadmium. The 4th Symposium
of the Society for Biology of Reproduction and Joint Polish-Japanese. Poland, 251p.
ERCAL, N.; GURER-ORHAN, H.; AYKIN-BURNS, N. (2001) Toxic metals and
oxidative stress part I: Mechanisms involved in metal-induced oxidative damage.
Med Chem. 1:529-39.
56 FIELDER, R. D.; DALE, E. A. (1983) Cadmium and its compounds. Toxicity Rev, No.
7. Health and safety executive, Her Majesty's Stationery Office, London.
FISHBASE.
Gymnotus
carapo.
Disponível
http://www.fishbase.org/Summary/SpeciesSummary.php?id=10915.
Acessado
em:
em
junho de 2010.
FÖRSTNER, U. E.; WITTMAN, G. T. W. (1983) Metal pollution in the aquatic
environment. 2ed. Berlin, Spriger-Verlag, 475-486p.
FRANÇA, L. R.; ROCHA, D. C. M.; MIRIANDA, J. R.; DEBELJUK, L. (2002)
Proliferación de células de Sertoli y función testicular. Boletin Informativo da La
Sociedad Argentina de Andrologia. 11:52-57.
FRIBERG, L. (1983) Cadmium. Annual Reviews in Public Health; 4:367-367.
GONZÁLEZ, A. O.; ROUX, J. P.; SÁNCHEZ, S. (2001) Evaluación de algunos
aspectos biológicos de morena (Gymnotus carapo, Linnaeus 1758). Morfología e
histología de ovário. Corrientes: UNIVERSIDAD NACIONAL DEL NORDESTE.
Comunicaciones
Científicas
y
Tecnológicas.
Disponível
em:
<www.unne.edu.ar/cyt/2001/4-Veterinarias/V-038.pdf>. Acesso em: 30 junho de
2010.
HAINES, T. A.; KOMOV, V. T.; JAGOE, C. H. (1992) Lake acidity and mercury
content of fish in Darwin natural reserve, Russia. Environmental Pollution. 78:107112.
HINTON, D. E.; POOL, C. R. (1976) Ultrastructure of the liver in channel catfish
Ictalurus punctatus (Rafinesque). Journal of Fish Biology. 8:209-219.
HODSON, P.V.; CASTONGUAY, M.; COUILLARD, C.M.; DESJARDINS, C.;
PELLETIER, E.; MCLEOD, R. (1994) Spatial and temporal variations in chemical
contamination of American eels, Anguilla rostrata, cap-tured in the estuary of the St.
Lawrence River. Can. J. Fish. Aquat. Sci. 2:464-478.
57 HOFFMAN, D.J.; RATTNER, B.A.; BURTON, G.A. & CAIRNS, J. (1995) Handbook of
ecotoxicology. London, Lewis publisher.
HOMMA - TAKEDA, S.; KUGENUMA, Y.; IWAMURO, T.; KUMAGAI, Y.; SHIMOJO,
N. (2001) Impairment of spermatogenesis in rats by methylmercury: involvement of
stage- and cell-specific germ cell apoptosis. Toxicology. 169:25-35.
HSDB - HAZARDOUSSUBSTANCES DATA BANK. Mercury. In: TOMES CPS
SYSTEM. Toxicology, occupational medicine and environmetal series. Englewwod:
Micromedex, 2000, CD-ROM.
HUTTON, M. (1987) Cadmium. In: Hutchinson TC, Meema KM, editors. Lead,
mercury, cadmium and arsenic in the environment. Chichester, UK: John Wiley &
Sons; 35-42p.
JARUP, L.; AKESSON, A. (2009) Current status of cadmium as an environmental
health problem. Toxicology and Applied Pharmacology. 238:201-108.
JOBLING, M. (1995) Simple indices for the assessment of the influences of social
environment on growth performace, exemplified by studies on Arctic charr.
Aquaculture International. 3:60-65.
KIME, E. D. (1999) A strategy for assessing the effects of xenobiotics on fish
reproduction. The Science of the Total Environment. 225:3-11.
KIRUBAGARAN, R.; JOY, K.P. (1988) Inhibition of testicular 3ß -hydroxy-? 5-steroid
dehydrogenase (3ß-HSD) activity in catfish, Clarias batrachus (L.) by mercurials.
Indian Journal of Experimental Biology. 26:907-908.
KIRUBAGARAN, R.; JOY, K. P. (1992) Toxic effects of mercury on testicular activity
in freshwater teleost, Clarias batrachus (L.). Journal. Fish Biol. 41:305-315.
58 KOULISH, S.; KRAMER, C. R.; GRIER, H. J. (2002) Organization of the male gonad
in a protogunous fish, Thalassoma bibasciatum (Teleostei: Labridae). Journal of
Morphology. 254:292-331.
LACERDA, L. D. (1997) Global emissions of Hg from gold and silver mining. Water,
Air Soil Pollut. 97:209-221.
LE GAC, F.; LOIR, M. (1999) Male reproductive system fish. In: KNOBIL, E.; NEILL,
J.D. (ed). Encyclopedia of Reproduction. San Diego: Academia Press. 3:20-30.
LEMARIE, A.; LAGADIC-GOSSMANN, D.; MORZADEC, C.; ALLAIN, N.; FARDEL,
O.; VERNHET, L. (2004) Cadmium induces caspase-independent apoptosis in liver
Hep3B cells: role for calcium in signaling oxidative stress-related impairment of
mitochondria and relocation of endonuclease G and apoptosis-inducing factor. Free
Radic Biol Med. 36:1517-1531.
LOCKHART, W. L.; UTHE, J. F. (1972) Methylmercury in northern pike (Esox lucius):
Distribution, elimination and some biochemical characteristics of contaminated fish. J.
Fish. Res. Board.Can. 29(11): 1519-1523.
MALAVOLTA, E. (1994) Fertilizantes e seu impacto ambiental: micronutrientes e
metais pesados: mitos, mistificação e fatos. São Paulo. Petroquímica 153p.
MALM, O.; GUIMARAES, J. R. D.; CASTRO, M. B.; BASTOS, W. R.; PFEIFFER, W.
C.; VIANA, J. P.; SILVEIRA, E. G. (1997) Mercúrio na Amazônia: Evolução da
contaminação ambiental e humana. Ciência Hoje. 22:16-23.
MARETTOVÁ, E.; MARETTA, M.; LEGATH, J. (2010) Changes in the peritubular
tissue of rat testis after cadmium treatment. Biol. Trace. Elem. Res. 134:288-295.
MARKHAM, R. M.; MCANELLY L. M.; STODDARD, K. P.; ZAKON, H.H. (2009)
Circadian and Social Cues Regulate Ion Channel Trafficking. PLoS Biology.
7(9):100-203.
59 MATTA, M. B.; LINSE, J.; CAIRNCROSS, C.; FRANCENDESE, L.; KOCAN, R. M.
(2000) Reproductive and transgenerational effects of methylmercury or aroclor 1268
on Fundulus heteroclitus. Environ. Toxicol. Chem. 20:327-335.
MAURO, J. B. N.; GUIMARÃES, J. R & MELAMED, R. (1999) Aguapé agrava
contaminação por mercúrio. Ciência hoje, 25:68-72.
MCKIM, J. M.; BENOIT, D. A. (1976) Effect of long term exposure to copper on
survival, growth and reproduction of brook trout (Salvelinus fontinalis). J. Fish. Res.
Bd. Can. 28:655-622.
MELA, M. (2004) Uso de Biomarcadores na avaliação dos efeitos do metilmercúrio
em Hoplias malabaricus (BLOCK, 1794) (Traíra). Dissertação de Mestrado.
Universidade Federal do Paraná, 138p.
MENIN, E. (1989) Anatomia funcional do tubo digestivo de Gymnotus carapo
Linnaeus, 1758 (Siluriformes, Gymnotoidei, Gymnotidae). Revista Ceres. 36:435457.
MIURA, T. (1999) Spermatogenic cycle in fish. In: KNOBIL, E; NEILL, J.D (ed).
Encyclopedia of Reproduction. 4:571-578.
MOHAMED, M. K.; Lee, W. I.; MOTTET, N. K.; BURBACHER, T.M. (1986) Laser
light-scattering study of the toxic effects of methylmercury on sperm motility. J Androl.
7:11-15.
MORAES, G.; AVILEZ, I. M.; ALTRAN, A. E. (2002) Respostas bioquímicas e
hematológicas de tuvira Gymnotus carapo (Linnaeus, 1758) exposta à hipóxia
ambiental. Brazilian Journal of Biology. 62:633-640.
MYERS, M. S.; FOURNIE, J. W. (2002) Histopathological biomarkers as integrators
of anthropogenic and environmental stressors. In: biological indicators of aquatic
ecosystem stress (Adams S.M.) American Fisheries Society. 24:221-287.
60 NIIMI, A. J.; KISSOON, G. P. (1994) Evaluation of the critical body burden concept
based on inorganic and organic mercury toxicity to rainbow trout (Oncorhynchus
mykiss). Arch. Environ. Contam. Toxicol. 26(2):169-178.
OLIVEIRA RIBEIRO C. A.; BELGER, L.; PELLETIER, E.; ROULEAU, C. (2002)
Histopathological evidence of inorganic mercury and methyl-mercury toxicity in the
arctic charr (Salvelinus alpinus). Environmental Research. 90:217-225.
OSA, R. H. (1994) "Mercury Atmospheric Processes: A Synthesis Report", Workshop
Proceedings, Tampa, Flórida, E.U.A.
PALENZUELA…
PANIGRADHI, A. K.; MISRA, B. N. (1978) Toxicological effects of mercury on a
freshwater fish, Anahas scandens, Cuv. and Val, and their ecological implications.
Environ. Pollut. 16:31-39.
PALENZUELA, B.; MANGANIELLO, L.; RIOS, A.; VALCARCEL, M. (2004)
Monitoring inorganic mercury and methylmercury species with liquid chromatography
piezo electric detection. Environmental research. 511: 289-294.
PARIZEK, J. (1957) O efeito destrutivo dos íons de cádmio no tecido testicular e sua
prevenção por zinco. J. Endocrinol. 15: 56-63.
PELGROM, S. M. G. J.; LAMERS, L. P. M.; LOCK, R. A. C.; BALM, P. H. M.;
WENDELAAR BONGA, S. E. (1995) Interactions between copper and cadmium
modify metal organ distribution in mature Tilapia, Oreochromis mossambicus.
Environmental Pollution. 90(3):415-423.
PELLETIER, E.; AUDET, C. (1995) Tissue distribution and histopathological effects
of dietary methymercury in benthic grubby Myoxocephalus aenaeus. Bull. Environ.
Contam. Toxicol. 54:724-730.
61 PFEIFFER, W. C.; MALM, O.; LACERDA, L. D.; KAREZ, C. S. (1992) Contaminação
ambiental e humana por metais pesados: Uma revisão. In Maciel T B. O ambiente
inteiro: A contribuição crítica da universidade na questão ambiental, RJ. Editora
UFRJ.180-194p.
PIETERSE, G. M.; VAN VUREN, J. H. J. (2004) Histopathologic changes in the
testes of the freshwater fish Oreochromis mossambicus (Cichlidae) as biomarker of
heavy metal pollution. SETAC Europe, 12th Annual Meeting, 12-16 May, 2002,
Vienna, Austria.
POPEK, W.; DIETRICH, G.; GLOGOWSKI, J.; DEMSKA-ZAKES, K.; DRAG-KOZAK,
E.; SIONKOWSKI, J.; LUSZCZEK-TROJAN, E.; EPLER, P.;DEMIANOWICZ, W.;
SAROSIEK, B.; KOWAESKI, R.; JAKUN, M.; ZAKES, Z.; KROL, J.; CZERNIAK, S.;
SZCZEPKOWSKI, M. (2006) Influence of heavy metals and 4-nonylphenol on
reproductive function in fish. Reproductive Biology.1:175-188.
PREDES, DE SOUZA F.; DIAMANTE, S. A. M.; DOLDER, H. (2010) Testis response
to low doses of cadmium in Wistar rats. Int.J.Exp.Path. 91:125-131.
PUDNEY, J. (1995) Spermatogenesis in nonmmamalian vertebrates. Microscopy
research and technique. 6:459-497.
RAND, G. M; PETROCELLI, S. R. Introduction. In: RAND, G.M. & PETROCELLI,
S.R., (Ed.). Fundamentals of aquatic toxicology: methods and applications, New
York: Hemisphere, p.1-28.
RAO, M. V (1989) Histophysiological changes of sex organs in methylmercury
intoxicated mice. Endocrinol. 23:55-62.
RESENDE, E. K. (1999) Estudos biológicos da Gymnotus carapo e Rhamphychthys
cf. marmoratus no Pantanal. Relatório Final do Projeto nº 01.0.94.572.03. Corumbá:
Embrapa Pantanal,17p (não publicado).
62 RESENDE, E. K. DE.; PEREIRA, R. A. C. (2000) Dieta alimentar de Gymnotus cf.
carapo, (Ostariophysi, Gymnotiformes), na planície inundável do Rio Negro, Mato
Grosso do Sul, Brasil. In: SIMPOSIO SOBRE RECURSOS NATURAIS E
SOCIOECONOMICOS DO PANTANAL, 3, Corumbá. Os desafios do novo milênio.
Resumos... Corumbá: Embrapa Pantanal, 285p.
ROALES, R. R.; PERLMUTTER, A. (1977) The effects of sub-lethal doses of
methylmercury and copper applied singly and jointly, on the immune response of the
blue gourami (Trichogaster trichopferus) to viral and bacterial antigens. Arch.
Environ. Contam. Toxicol. 6:325-331.
RODGERS, D. W.; BEAMISH, F. W. H. (1982) Dynamics of dietary methylmercury in
rainbow trout, Salmo gairdneri. Aquat. Toxicol. 2:271-290.
RURANGWA, E.; ROELANTS I.; HUYSKENS G.; EBRAHIMI, M.; KIME, D. E (1998)
The minimum effective spermatozoa to egg ratio for artificial insemination and the
effects of mercury on sperm motility and fertilization ability in Clarias gariepinus.
Journal of Fish Biology. 53:402-413.
RUSSELL, L. D.; ETTLIN, R. A.; SINHA-HIKIM, A. P.; CLEGG, E. D.(1990)
Histological and histophatological evaluation of the testis. 286p.
SCHULZ, W. R.; DE FRANÇA, R. L.; LAREYRE, J. J.; LEGAC, F.; CHIARINIGARCIA, H.; NOBREGA, H. R.; MIURA, T. (2010) Spermatogenesis in fish. General
and Comparative Endocrinology. 165:390-411.
SILVA FILHO, M. V.; OLIVEIRA, M. M.; CUNHA, B. V. L .F.; ALVES, M. V.; CUNHA,
B. J. (2000) Validação de espécies sentinelas para biomarcação com colinesterases
de peixes. In: Ecotoxicologia: perspectivas para o século XXI (ESPÍNDOLA, E. L. G.;
PASNCHOAL, C. M. R. B.; ROCHA, O.; BOHRER, M. B. C. & OLIVEIRA NETO, A.
L.) São Carlos, Rima.
63 SILVA, J. M.; OLIVEIRA, J. I. J. de. (1997) Morfologia do intestino do “tuvira”
Gymnotus carapo L., 1758 (Pices - Gymnotidae). Revista Científica, UFMS, Campo
Grande. 4:18-22.
SILVA, M.; GODINHO, H. P. (1983) Timing of some events of the gametogenesis in
the male Nile tilapia, Sarotherodon niloticus. Archives of Anatomy and Microscopy.
72: 231-237.
SILVA,
M.
(1987)
Morfologia
ultraestrutural
do
testículo,
cinética
da
espermatogênese e barreira hematotesticular da tilápia do Nilo, Oreochromis
niloticus. Tese (Doutorado em Biologia Celular). Universidade Federal de Minas
Gerais, Belo Horizonte.
SIU, M. K.; MRUK, D .D.; PORTO, S. C.; CHENG, C. Y (2009) Cadmium-induced
testicular injury. Toxicology and Applied Pharmacology. 238:240-249.
SNARSKI, V. M.; OLSON, G. F. (1982) Chronic toxicity and bioaccumulation of
mercuric chloride in the fathead minnow (Pimephales promelas). Aquat. Toxicol.
2:143-156.
STANSLEY, W.; ROSCOE, D. E.; HAZEN, R. E. (1991) Cadmium contamination in
deer livers in New Jersey; human health risk assessment. Science of Total
Environment. 107:71-82.
STOHS, S. J.; BAGCHI, D. (1995) Oxidative mechanisms in the toxicity of metal
ions. Free Radic. Biol. Med. 18:321-336.
SWEET, L. I.; PASSINO-READER, D. R.; MEIER, P. G.; OMANN, G. M. (1999)
Xenobiotic - induced apoptosis: Signicance and potential application as a general
biomarker of response. Biomarkers. 4:237-253.
THEODORO, E. (2003) Caracterização sócio-econômica da atividade de coleta e
comercialização de isca viva na BAP-MT (2003) In: Projeto implementação de
práticas de gerenciamento integrado de bacia hidrográfica para o Pantanal e bacia
64 do
alto
Paraguai.
Resumo
Executivo
do
Relatório
Final.
Cuiabá:
ANA/GEF/PNUMA/OEA - FEMA/MT, 27p.
THOMPSON, C. B. (1995) Apoptosis in the pathogenesis and treatment of disease.
Science. 267:1456-1462.
THOMPSON, J.; BANNIGAN, J. (2008) Cadmium: toxic effects on the reproductive
system and the embryo. Reprod. Toxicol. 25:304-315.
VALKO, M.; MORRIS, H.; CRONIN, M.T. (2005) Metals, toxicity and oxidative stress.
Curr. Med.Chem. 12:1161-1208.
VALLEE, B. L.; ULMER, D. D. (1972) Biochemical effects of mercury, cadmium an
lead. Reviews in Biochemical Toxicology. 41:91-128.
VAN DER OOST, R.; BEYER, J.; VERMEULEN, N. P. E. (2003) Fish
bioaccumulation and biomarkers in environmental risk assessment. Environmental
Toxicology and Pharmacology.13:57-149.
VERGILIO, DOS SANTOS C (2009) Efeitos morfológicos e ultraestruturais da
contaminação por cloreto de mercúrio e cádmio in vivo e in vitro. Dissertação de
mestrado. Universidade Estadual do Norte Fluminense. Centro de Biociências e
Biotecnologia, 45p.
VIARENGO, A. (1989) Heavy metals in marine invertebrates: mechanisms of
regulation and toxicity at the cellular level. Reviews in Aquatic Sciences. 1:295p.
VILELA, D. A. R.; SILVA, S. G. B.; PEIXOTO, M. T. D.; GODINHO, H. P.; FRANÇA,
L. R. (2003) Spermatogenesis in teleost: insights from the Nile tilapia (Oreochromis
niloticus) model. Fish Physiology and Biochemistry. 28:187-190.
WORLD HEALTH ORGANIZATION (WHO) Inorganic mercury. Environment Health
Criteria 171: Geneva, 1991, 140pp.
65 WORLD HEALTH ORGANIZATION (WHO) Cadmium. Environmental Health Criteria
134: Geneva, 1992, 280pp.
WEIS, J. S.; KHAN, A. A. (1990) Effects of mercury on the feeding behavior of the
mummichog, Fundulus hereroclitus from a polluted habitat. Marine Environ. Res. 30:
243-249.
WELTZIEN, F.A.; ANDERSON, E.; SHALCHIAN-TABRIZI, K.; NORBERG, B. (2004).
The brain pituitary gonad axis in male teleosts with special emphasis on flatfish
(Pleuronectiformes). Comparative Biochemistry and Phisiology. 3:447-477.
WESTBY, G. W. M. (1975) Comparative studies of the aggressive behaviour of two
Gymnotid eletric fish (Gymnotus carapo and Hypopomus artedi). Animal Behaviour.
23:192-213.
WESTER, P. W.; CANTON, J. H (1992) Histopathological effects in Poecilia
reticulate (guppy) exposed to methyl mercury chloride. Toxicol. Pathol. 20:81-92.
66