Cadeira de Biofísica Molecular
Teórico-Prática 11
Pergunta de desenvolvimento
Paula Tavares, FCUL (2012-2013)
1
Proposta de Trabalho para as Teórico-Práticas
• Trabalho escrito (a discutir na última TP):
Nomeie e caracterize em termos de estrutura e
funcionamento os principais tipos de músculo
cardíaco. Nesse contexto, descreva os fluxos iónicos
que estão na origem das flutuações do potencial
intracelular durante o cíclo cardíaco, e a acção
modulatória do sistema nervoso autónomo sobre os
mesmos.
• Máximo de 1000 palavras. Podem juntar figuras ao vosso
trabalho.
2
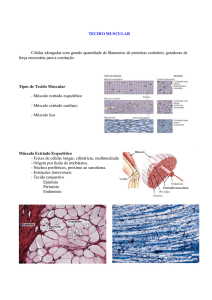
O músculo cardíaco
Três tipos principais de músculo cardíaco:
• Músculo auricular (ou miocárdio auricular);
• Músculo ventricular (ou miocárdio
ventricular);
• Sistema especializado de condução, que inclui:
nó sinoauricular (nó SA); tractos internodais
anterior, médio e posterior; feixe de Bachmann;
nó aurículo-ventricular (nó AV); feixe de His;
ramo esquerdo e direito do feixe de His; divisão
ântero-superior, ântero-medial e póstero-inferior
do ramo esquerdo do feixe de His; rede de
Purkinje.
Os músculos auricular e ventricular contraem da mesma forma que o músculo
esquelético, mas a duração da contracção é muito mais longa no músculo cardíaco.
Os músculos do sistema especializado de condução contraem muito pouco pelo
facto de terem poucas fibras contrácteis. A sua função principal é a de gerar sinais
3
rítmicos, a velocidades variáveis, que controlam o ritmo cardíaco.
1
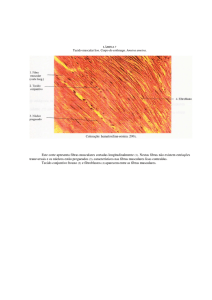
As células do músculo cardíaco
2
As células do músculo cardíaco são
ramificadas, uninucleadas, estriadas
como resultado do arranjo dos
filamentos de actina e miosina de forma
semelhante à do músculo esquelético.
São conhecidas por cardiomiócitos.
As áreas escuras que atravessam as fibras
musculares cardíacas chamam-se discos
intercalares. Os discos intercalares são http://umanitoba.ca/faculties/medicine/units/anatomy/images/cardiaccopy.JPG
membranas celulares que separam duas
células musculares cardíacas adjacentes.
Consequentemente, as fibras cardíacas
Katz, 2006, Physiology of the Heart, Lippincott Williams & Wilkins.
são formadas por muitas células individuais, ligadas entre si em série.
Na zona dos discos intercalares as membranas celulares fundem-se, comunicando entre
si através de junções de hiato que permitem uma difusão quase livre dos iões entre
células adjacentes. Consequentemente os potenciais de acção conseguem propagar-se
sem obstáculos ao longo de toda a fibra, viajando de célula a célula. Por essa razão o
coração é um sincício de muitas células musculares cardíacas.
4
Os sincícios cardíacos
3
O coração é composto por dois sincícios:
• O sincício auricular, que constitui as paredes externas das duas aurículas.
• O sincício ventricular, que constitui as paredes dos dois ventrículos.
Os dois sincícios estão separados por
tecido fibroso. Em condições normais,
os potenciais não são conduzidos do
sincício auricular directamente ao
sincício ventricular.
A comunicação eléctrica entre os dois
sincícios faz-se através dum tecido de
condução especializado designado nó
aurículo-ventricular (nó AV; A-V
bundle).
5
Potenciais de acção no músculo cardíaco
4
O potencial de acção no músculo ventricular tem uma
amplitude média de 105 mV (varia entre -85/-90 mV
e os 20 mV): -85/-90 mV entre batidas e +20 mV
durante a batida.
Despolarização Inicial: Mediada pela abertura de
canais rápidos de Na+, que fecham abruptamente no
pico da despolarização.
Formação do Pico: Abertura temporária de canais
de K+ e de Cl- provoca descida ligeira do potencial.
Segue-se uma Zona de Repolarização Lenta onde o
& Hall, 2005, Textbook of Medical Physiology,
potencial permanece positivo durante cerca de 0.2 a Guyton
Pennsylvania: Elsevier
0.3 segs – esta zona chama-se Plateau (a contracção
ventricular dura cerca de 15× mais do que a esquelética). Mediada pela abertura de
canais lentos de cálcio (tipo L) e pela diminuição da condutividade membranar ao K+
(cerca de 5×) após o início do potencial de acção.
Repolarização ligeira durante o Plateau: Mediada pela saída de iões K+.
Repolarização final: Fecho canais lentos de cálcio (tipo L) e aumento condução da
6
membrana ao K+. Saída rápida de K+ da fibra muscular cardíaca.
O sistema especializado de condução
5
O sistema especializado de condução é constituído por:
Nó ou nódulo sinusal ou sinoauricular (nó
SA) Sinus Node, Sinoatrial Node or S-A
node: é gerado o sinal que provoca o impulso
rítmico normal do coração.
Tractos internodais anterior, médio e
posterior Anterior, middle and posterior
internodal tract: : transportam o sinal do nó
sinusal ao nó aurículo-ventricular.
Nó ou nódulo aurículo-ventricular (nó AV)
Atrioventricular Node or AV node: atrasa a
condução do sinal das aurículas aos
ventrículos.
Feixe de Bachmann ou via inter-auricular
Bachmann´s bundle or Interartrial band:
conduz o sinal do nó sinusal à aurícula
esquerda.
7
O sistema especializado de condução
5
Feixe de His ou Feixe aurículo-ventricular
Bundle of His or A-V bundle: transporta o
sinal do nó AV para os ramos esquerdo e
direito do Feixe de His.
Ramo esquerdo e direito do feixe de His
Left and right bundle branch: conduz o
sinal para a rede de Purkinje.
Rede de Purkinje Purkinje fibers: conduz
o sinal a todas as partes dos ventrículos.
8
Nó sinusal (células pacemaker)
6
Algumas fibras cardíacas tem a capacidade
de se auto-excitarem, um processo que
causa a descarga e a contracção muscular
rítmica e automática. Isto é especialmente
verdade no sistema especializado de
condução do coração. A porção do sistema
que apresenta os maiores índices de autoexcitação é o nó sinusal. Por essa razão, é o
nó sinusal que controla a frequência de
batimento cardíaco.
O nó sinusal é pequeno, achatado e com formato
elíptico (15 mm comprimento, 3 mm de largura e 1 mm
de espessura). Este nó fica localizado na parede
posterolateral superior da aurícula direita, muito
próximo da zona de inserção da veia cava superior. As
fibras não contêm quase nenhum filamento contráctil e
têm cerca de 3 a 5 mm de diâmetro em contraste com o
diâmetro de 10 a 15 mm característico das fibras
musculares cardíacas circundantes.
9
Nó sinusal
6
As terminações das fibras nodais contactam
directamente com as fibras musculares
auriculares circundantes. Consequentemente,
um potencial de acção que tenha origem no
nó sinusal propaga-se ao músculo
circundante, que por sua vez o transmite às
fibras vizinhas, conduzindo à excitação de
todo o músculo auricular.
A velocidade de condução é de cerca de 0.3
m/s no músculo auricular.
Existem umas fibras especiais, as fibras internodais e o feixe
de Bachmann, que ligam o nó sinusal ao nó aurículoventricular (nó AV) e o nó sinusal à aurícula esquerda,
respectivamente, onde a condução é mais rápida, na ordem do
1 m/s. A rápida condução nestas fibras deve-se à presença de
fibras de condução especializadas, similares ao rápido sistema
de condução das fibras de Purkinje.
10
Nó sinusal
Note que o potencial de repouso das células
do nó sinusal é de cerca –55 a –60 mV,
por comparação com os –85 a –90 mV da
célula muscular ventricular.
As células membranares do nó sinusal
apresentam uma maior condutividade ao Na+ e ao Ca2+, que
tendem a entrar na célula aumentando o potencial de repouso.
Guyton & Hall, 2005, Textbook of Medical Physiology, Pennsylvania: Elsevier.
Nesta figura apresentam-se os potenciais de
acção gerados no nó sinusal em três batimentos
cardíacos, e por comparação o potencial de
acção gerado numa fibra muscular ventricular
durante um batimento cardíaco.
7
• As fibras nodais possuem um número moderado de canais de Na+ que estão abertos
durante o estado de repouso. Como consequência entre batimentos cardíacos, o influxo
de iões Na+ (if) provoca um aumento lento do potencial de repouso membranar.
• Canais de cálcio (tipo T, de transient), activos numa gama de potenciais mais
negativa que os tipo L, permitem a entrada de Ca2+ (iCaT) na parte final deste período de
11
subida progressiva do potencial (Katz, 2006, Physiology of the Heart, Lippincott Williams & Wilkins).
Nó sinusal
7
• Quando o potencial atinge os –40 mV, dá-se a
abertura dos canais lentos de cálcio (tipo L, de
lasting), que permitem a entrada de mais cálcio
(iCaL) , gerando o potencial de acção.
• Cerca de 100 a 150 ms após a abertura, os canais
de cálcio (tipo L) ficam inactivos. Ao mesmo tempo
abrem os canais de K+ (iK).
• O influxo através dos canais de cálcio (tipo L)
cessa, enquanto que grandes quantidades de K+
movimentam-se para o exterior da célula, reduzindo
o potencial de repouso para o seu valor mínimo de
–55 a –60 mV (hiperpolarização), e terminando o
potencial de acção. Nesta altura fecham os canais de
K+. E o processo volta ao início.
As concentrações citosólicas de repouso para o
Na+, K+ e Ca2+ são restabelecidas, por todo o
coração, através da bomba1 Na+-K+ ATPase, do
trocador1 Na+-Ca2+e de bombas1,2 Ca2+ ATPase.
1. Sarcolema; 2. Retículo Sarcoplasmático.
3 Na+/Ca2+ (inward current +1)
Katz, 2006, Physiology of the Heart, Lippincott Williams & Wilkins.
12
Nó aurículo-ventricular
8
O atraso da condução do sinal das
aurículas para os ventrículos é feito no nó
aurículo-ventricular e fibras de condução
adjacentes.
O nó aurículo-ventricular está localizado
na parede posterior da aurícula direita,
atrás da válvula tricúspide.
A baixa velocidade de condução das fibras
do nó aurículo-ventricular e das fibras de
condução adjacentes deve-se em parte ao
facto do seu tamanho ser consideravelmente
menor do que o das fibras musculares das
aurículas. Por outro lado, é menor o número
de junções de hiato entre as células
musculares adjacentes. No conjunto, estes
factores provocam uma maior resistência à
passagem do sinal iónico.
13
Nó aurículo-ventricular
8
Na figura estão representadas as diferentes
partes do nó aurículo-ventricular e a sua
ligação às fibras internodais e ao de feixe de
His. A figura também indica o intervalo de
tempo que já decorreu desde o disparo do sinal
pelo nó sinusal.
• O sinal chega ao nó aurículo-ventricular 30
ms após a sua origem no nó sinusal.
• Sofre um atraso de 90 ms no nó aurículoventricular (Tempo total após passagem do
nó é de: 30 ms + 90 ms = 120 ms = 0.12
segs).
• Depois atravessa a porção penetrante do
feixe de His, que penetra o tecido fibroso
Guyton & Hall, 2005, Textbook of Medical Physiology, Pennsylvania: Elsevier.
que separa as aurículas dos ventrículos.
Esta passagem provoca um atraso adicional no sinal de 40 ms. O atraso conjunto do nó
aurículo-ventricular e feixe de His é de 130 ms (90 ms + 40 ms = 130 ms). No total o
sinal demora 160 ms a chegar ao princípio dos ramos esq. e dir. do feixe de His.
14
Fibras de Purkinje
9
As fibras de Purkinje são
muito grandes, maiores do
que as fibras musculares
típicas dos ventrículos, e
transmitem potenciais de
acção com uma velocidade
entre os 1.5 e os 4 m/s.
Esta velocidade é 6× maior
que a do tecido muscular
típico dos ventrículos e
150× maior do que a de
algumas fibras
do nó aurículo-ventricular. Isto permite uma transmissão quase imediata do sinal a
todo o músculo ventricular.
As fibras de Purkinje tem origem no nó aurículo-ventricular, onde constituem o
Feixe de Hiss. Na base do coração os ramos direito e esquerdo do Feixe de Hiss
terminam nos ramos direito e esquerdo, respectivamente, da rede de Purkinje, que
ascendem pelas paredes laterais do coração.
15
Fibras de Purkinje
9
A rede de Purkinje penetra em cerca
de 1/3 da fibra muscular, e no seu
término torna-se contínua com as
fibras musculares cardíacas. A
propagação do sinal desde a entrada
nos ramos do feixe de His até às
extremidades da rede de Purkinje
demora cerca de 30 ms.
A elevada velocidade de transmissão do
sinal nas fibras de Purkinje deve-se à
existência de inúmeras junções de hiato
nos discos intercalares que separam as
células adjacentes, o que facilita a
propagação do sinal iónico entre células.
As células de Purkinje tem muito poucas
miofibrilhas, portanto quase que não
contraem durante a transmissão do
impulso.
16
Fibras de Purkinje
9
Uma das características especiais do feixe de His é a sua incapacidade, excepto em
situações anormais, de conduzir os potenciais de acção em sentido inverso. Isto evita
a reentrada de impulsos cardíacos dos ventrículos para as aurículas.
Os tecidos musculares das aurículas e dos ventrículos estão separados, excepto no
feixe de His, por uma barreira isolante de tecido fibroso. Quando há falhas nesta
barreira, o sinal pode reentrar nas aurículas provocando arritmias graves (alterações do
ritmo cardíaco normal).
O músculo cardíaco enrola-se à volta do coração,
formando uma espiral com duas camadas, separadas entre
si por um septo fibroso (parede divisória). O impulso
cardíaco segue estas espirais de músculo. Devido a isso, a
transmissão do impulso do músculo que se encontra junto
da superfície endocárdica até ao músculo que se encontra
junto da superfície epicárdica requer cerca de 30 ms.
Portanto, a propagação do sinal desde a entrada nos ramos
do feixe de His até à activação de todo o músculo
ventricular demora 60 ms.
17
Controlo da excitação e condução no coração
10
Em condições normais, o impulso cardíaco começa no nó sinusal. Em condições
anormais, outras partes do coração podem exibir excitação rítmica, em especial o nó
aurículo-ventricular e as fibras de Purkinje.
As fibras do nó aurículo-ventricular, quando
não são estimuladas por uma fonte externa,
disparam 40 a 60 vezes por minuto. As fibras de
Purkinje, quando não são estimuladas por uma
fonte externa, disparam 20 a 40 vezes por
minuto. Estas taxas de disparo são
significativamente mais baixas do que as
produzidas pelo nó sinusal: 70 a 80 vezes por
minuto.
De cada vez que o nó sinusal dispara um impulso, o sinal excitatório chega ao nó
aurículo-ventricular e às fibras de Purkinje, onde também dispara impulsos. No fim
desses impulsos, em ambos os nós e fibras de Purkinje, dá-se a repolarização
membranar. O nó sinusal controla o batimento cardíaco porque é sempre ele que chega,
antes de todas as outras fibras, ao limiar de activação. Diz-se por isso que o nó sinusal é
o pacemaker do coração.
18
Controlo do ritmo cardíaco pelo sistema nervoso autónomo
11
A frequência cardíaca é controlada pelos nervos
simpáticos (estimulam) e nervos parassimpáticos
(inibem).
A estimulação simpática pode aumentar a
frequência de batimento cardíaco das 70× por
minuto, para as 180 ou 200× por minuto, e
raramente até 250× por minuto, ou seja, duas a
três vezes mais do que o normal. A estimulação
simpática aumenta também a força de contracção
cardíaca, e consequentemente o volume de
sangue bombeado e a pressão de ejecção.
A estimulação parassimpática pode fazer
Guyton & Hall, 2005, Textbook of Medical Physiology, Pennsylvania:
com que o coração pare durante
Elsevier
alguns segundos. Mas depois o coração retoma os batimentos, batendo cerca de 20
a 40× por minuto. Forte estimulação parassimpática também pode reduzir a força
de contracção cardíaca em 20 a 30%. As fibras parassimpáticas estão
fundamentalmente distribuídas pelas aurículas, mas não pelos ventrículos. Isto
explica porque é que a estimulação parassimpática afecta fundamentalmente a
19
frequência de batimento cardíaco e não a força de contracção.
Estimulação Simpática
12
A estimulação simpática aumenta a velocidade
de disparo do nó sinusal, a velocidade de
condução do sinal no sistema especializado de
condução, a excitabilidade de todo o coração, e
a força de contracção em toda a musculatura
cardíaca. Máxima estimulação pode quase
triplicar a frequência de batimento cardíaco e
duplicar a força da contracção cardíaca.
Gαs
Os nervos simpáticos libertam norepinefrina.
A norepinefrina aumenta a permeabilidade membranar ao Na+ e ao Ca2+ (via cAMP).
No nó sinusal o aumento da permeabilidade ao Na+ e ao Ca2+ origina um potencial de
repouso mais positivo e faz com que se chegue ao potencial limiar mais rapidamente
(dispara mais frequentemente). No restante sistema de condução, o aumento da
permeabilidade ao Na+ e ao Ca2+ origina um potencial de repouso mais positivo (facilitando
a sua excitação). O aumento da força de contracção muscular dever-se-á, em parte, ao
20
aumento da concentração do Ca2+ no interior das fibras musculares.
Matthews (2001), Neurobiology, Wiley-Blackwell.
Os nervos simpáticos encontram-se distribuídos
por todo o coração, com uma forte
representação no músculo ventricular.
Estimulação Vagal
13
Os nervos parassimpáticos estão distribuídos
fundamentalmente nos nós sinusal e aurículoventricular, e em menor extensão nos músculos
das duas aurículas, e quase nada nos músculos dos
dois ventrículos.
A estimulação dos nervos parassimpáticos do
coração (nervo vago) provoca a libertação de
acetilcolina nas terminações nervosas. A
acetilcolina: (1) Diminui o ritmo de disparo do nó
sinusal; (2) Diminui a excitabilidade das fibras de
condução adjacentes ao nó aurículo-ventricular,
atrasando desta forma a transmissão do sinal aos
ventrículos.
Estimulação vagal fraca ou moderada pode diminuir o batimento cardíaco para metade
do seu ritmo normal. Forte estimulação vagal pode bloquear a excitação rítmica do
coração pelo nó sinusal ou a transmissão deste sinal ao nó aurículo-ventricular. Nestas
circunstâncias os ventrículos param de bater até às fibras de Purkinje, normalmente da
zona do septo interventricular, se tornarem o pacemaker. Este fenómeno chama-se
21
escape ventricular.
Estimulação Vagal
13
Matthews (2001), Neurobiology, Wiley-Blackwell.
A acetilcolina provoca o aumento da
permeabilidade das fibras ao K+.
O K+ sai da célula, fazendo com que
esta fique hiperpolarizada, tornando-a
por isso menos excitável. No nó sinusal,
o potencial de repouso passa a estar nos
–65 a –75 mV, em vez dos normais –55
a –60 mV.
Desta forma, a subida do potencial
membranar até ao potencial limiar,
devido à entrada lenta de iões Na+ e de
Ca2+, demora mais tempo.
Gαi – Inibe adenilato
ciclase (cAMP↓)
Isto atrasa consideravelmente a taxa de disparo do nó sinusal.
No nó aurículo-ventricular, o estado de hiperpolarização faz com que o sinal
despolarizante transportado pela fibras de condução adjacentes ao nó não seja
suficiente para o excitar. Broodle et al, 2001, Basic Res Cardiol 96: 528 – 538.
22