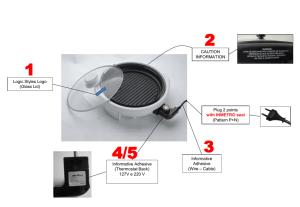
Arquitetura das
Redes de Neurônios
Tiposdearquitetura
• Ar/ficial
• Determinadapordados
experimentais(data-driven)
ArquiteturasAr/ficiais
•
•
•
•
GrafodeErdős–Rényi
Mundopequeno
Livredeescala
…
Solé&Valverde,2004
TeoriadeGrafos:algumasdefinições
Grafo:direcionadoou
não-direcionado
Arestas:bináriasou
ponderadas
Network Architectures and Metrics
Graphs: Visualization
Graphs can be displayed in matrix form or by embedding them in (usually
2D) space. Graph embedding and visualization is an extremely active
area of research in its own right@.
The image cannot be displayed. Your computer may not have enough memory
to open the image, or the image may have been corrupted. Restart your
computer, and then open the file again. If the red x still appears, you may
have to delete the image and then insert it again.
matrizdeadjacências
Visualizing a graph goes a long way towards understanding its structure.
www.brain-connec/vity-toolbox.net
Redescomplexas
Regular
Lgrande,Cgrande
MundoPequeno
Lpequeno,Cgrande
Erdős–Rényi
Lpequeno,Cpequeno
Aumentonaaleatoriedadedasconexões
!
WaZs&Strogatz(1998)Nature393,440
of random links (Figure Ib). Combining elements of order and
randomness, such networks were characterized by high degrees of
local clustering as well as short path lengths, properties shared by
genetic, metabolic, ecological and information networks [1–3].
The nodes in random graphs have approximately the same DEGREE
(number of connections). This homogeneous architecture generates a
normal (or Poisson) degree distribution. However, the degree
distributions of most natural and technological networks follow a
random networks but with significantly higher values for the
clustering coefficient. At the structural level, cortical networks
either do not appear to be scale-free [35] or exhibit scale-free
architectures with low maximum degrees [44], owing to saturation
effects in the number of synaptic connections, which prevent the
emergence of highly connected hubs. Instead, functional brain
networks exhibit power law degree distributions as well as smallworld attributes [52,62].
Tiposderedescomplexas
(b) L=1.79 (0.04)
C=0.52 (0.04)
(a) L=1.68 (0.01)
C=0.35 (0.03)
rk Architectures and Metrics
(c) L=1.73 (0.06)
C=0.52 (0.05)
Spornsetal.(2004)
TrendsCognSci8,418
ll-World Networks
lustering Figure
andI. Structure
path oflength
aloneandare
insufficient
to have
fully
characterize
random, small-world
scale-free
networks. All networks
24 nodes
and 86 connections with nodes arranged on a circle. The characteristic
path length L and the clustering coefficient C are shown (mean and standard deviation for 100 examples in each case; only one example network is drawn). (a) Random
Erdős–Rényi
Mundopequeno
Livredeescala
etwork topology.
There
are
several
types
of
small-world
architectures.
network. (b) Small-world network. Most connections are among neighboring nodes on the circle (dark blue), but some connections (light blue) go to distant nodes,
creating short-cuts across the network. (c) Scale-free network. Most of the 24 nodes have few connections to other nodes (red), but some nodes (black connections) are
linked to more than 12 other nodes. For comparison, an ideal lattice with 24 nodes and 86 connections has LZ1.96 and CZ0.64.
Modular
and wiring [19], our focus is on the large-scale and
dynamics of neurons and neuronal populations result in
intermediate-scale networks of the cerebral cortex, allowpatterns of statistical dependencies (functional connecing us to examine links between neural organization and
tivity) and causal interactions (effective connectivity),
cognition arising at the ‘systems’ level. We divide this
defining three major modalities of complex brain networks
review into three parts, devoted in turn to the organiz(Box 2). Human cognition is associated with rapidly
ation (structure), development (growth) and function
changing and widely distributed neural activation pat(dynamics) of brain networks.
terns, which involve numerous cortical and sub-cortical
4
DEUTSCHE PHYSIKALISCHE GESELLSCHAFT
regions activated in different combinations and contexts
[12–15]. Two major organizational principles of the
Structural organization of cortical networks
cerebral cortex are functional segregation and functional
Most structural analyses of brain networks have been
gular graph
with shortcuts
C
integration
[16–18], enabling
the rapid extraction of
carried out on datasets describing the large-scale conneccommunities
information and the generation of coherent brain states.
tion patterns of the cerebral cortex of rat [20], cat [21,22],
Which structural and functional principles of complex
and monkey [23] – structural connection data for the
networks promote functional segregation and functional
human brain is largely missing [24]. These analyses have
integration, or, in general, support the broad range and
revealed several organizational principles expressed
flexibility
of cognitive
processes?
within structural brain networks. All studies confirmed
Graph
with
hierarchically
arranged modules C
In this review we examine recent insights gained about
that cerebral cortical areas in mammalian brains are
fractal
(self-similar)
adjacency
patterns
of brain connectivity
from thematrix
application of novel
neither completely connected with each other nor ranquantitative computational tools and theoretical models to
domly linked; instead, their interconnections
show
Figure 1. Schematic
view a
of a hierarchical cluster network with five clusters
containing five sub-clusters each.
Kaiseretal.(2007)NewJPhys9,110
empirical datasets. Whereas many studies of single
specific and intricate organization. Methodologically,
Strogatz (1998)
Nature
393, 440.
Kaiser etmorphology
al. (2007) Newinvestigations
J Phys 9, 110. have used either graph(a) theoretical(b)
(c)
neuron
networks
have revealed their complex
Livrede
Escala
Hierárquica
emodular
k Architectures and Metrics
p of Network Architectures
Redescomplexassãocomuns
al-world networks occupy distinct locations in a continuous space of
sible network architectures.
Internet
Solé&Valverde(2004)LectNotesPhys650,189
lverde (2004)
Lect Notes Phys 650, 189.
Tráfegoaéreo
15
Rede
metabólica
Redes
sociais
Review
the intuitive, common sense notion of complexity by emphasizing the idea
that complex systems are neither completely regular nor completely random.
For example, neither a random string nor a periodically repeating string of letters is complex, while a string of English text certainly is. More generally, any
system of elements arranged at random (e.g. gas molecules) or in a completely
regular or homogeneous way (molecules in a crystal lattice) is not complex.
By contrast, the arrangement and interactions of neurons in a brain or of
molecules in a cell is obviously extremely complex (see Fig.).
Redescomplexasnocérebro
TRENDS in Cognitive Sciences
(a)
Vol.8 No.9 September 2004
421
(c)
Lattice
Clustering coefficient
0.6
ar
fr
w
re
iz
v
fo
q
(h
Macaque visual cortex
0.5
0.4
Random
0.3
(b)
1.7
1.8
Path length
co
n
tr
n
th
b
n
d
o
tu
st
y
1.9
R
(d)
Tononietal.(1998)TrendsCognSci2,474
104
Counts (k)
500
Counts (k)
103
0
102
700
800
Degree k
101
rc = 0.6
rc = 0.7
100
100
101
102
103
Degree k
Figure 1. Small-world and scale-free structural and functional brain networks. (a) Characteristic path length and clustering coefficient for the large-scale connection matrix
(see Glossary) of the macaque visual cortex (red) (connection data from [23], results modified from [35]). For comparison, 10 000 examples of equivalent random and lattice
networks are also shown (blue). Note that the cortical matrix has a path length similar to that for random networks, but a much greater clustering coefficient. (b) Cluster
structure of cat corticocortical connectivity, based on [32] and visualized using Pajek (http://vlado.fmf.uni-lj.si/pub/networks/pajek/). Bars indicate borders between nodes in
separate clusters. Cortical areas were arranged around a circle by evolutionary optimization, so that highly inter-linked areas were placed close to each other. The ordering
agrees with the functional and anatomical similarity of visual, auditory, somatosensory-motor and frontolimbic cortices. (c) A typical functional brain network extracted from
human fMRI data (from [52]). Nodes are colored according to degree (yellowZ1, greenZ2, redZ3, blueZ4, other coloursO4). (d) Degree distribution for two correlation
thresholds. The inset depicts the degree distribution for an equivalent random network (data from [52]).
A number of complexity measures have been proposed, but only a few satisfy
the requirement of attaining small values for both completely random and
completely regular systems. In neurobiology, for example, one often encounters the term ‘dimensional complexity’ or just ‘complexity’ referring to the socalled correlation dimension of EEG signalsa. Its value appears to increase, for
instance, from sleep to waking states, or with brain maturationb,c. The correlation dimension is a measure developed in the context of nonlinear dynamics, which should be proportional, roughly speaking, to the number of independent neuronal populations giving rise to an EEG signald. But because the
correlation dimension would be higher for complete independence than for
the mixture of functional segregation and integration that characterizes brain
dynamics, it violates the criterion for complexity mentioned above.
Complexity measures have been proposed in the context of algorithmic
information theory, which deals with the information necessary to generate
individual bit strings. For example, the well-known algorithmic (or
Asorganizaçõesestruturaise
funcionaisdocérebrotêm
caracterís/casderedescomplexas–
comotopologiademundopequeno,
hubsaltamenteconectadose
modularidade–,tantonaescalado
cérebrointeiro(reveladaportécnicas
deneuroimagememhumanos)como
the system, summed over all subset sizes. Thus, com
naescalacelular(reveladapor
provides a measure for the amount of information
integrated within a neural system (for a discuss
estudosemanimais).
complexity measures, see Box 2).
CONNECTEDNESS of neural structures can affect the funcSpornsetal.(2004)TrendsCognSci8,418
A schematic illustration of the notion of comp
tional impact of local and remote network lesions [43], and
Bullmore&Sporns(2009)NatRevNeurosci10,186
this property might also be an important factor for
based on the results of large-scale computer simulati
as well as primate prefrontal cortex [41]. The algorithm
could be steered to identify clusters that no longer
contained any known absent connections, and thus
Modelosderedescomplexas
paraasredescerebrais
(anatômicasefuncionais)
Redesdemundopequenosãoummodeloatraentepara
aorganizaçãodasredesanatômicasefuncionaisdo
cérebroporqueatopologiademundopequenopermite
conciliarduasformasdis/ntasdeprocessamentode
informação:segregada(especializada)edistribuída
(integrada).
Basset&Bullmore(2006)TheNeuroscien/st12,512
Determinadapordados
• Incorporainformaçãoquan/ta/vasobreaarquitetura
deredescerebraisreais
Sirosh&Miikkulainen,19942004
The image cannot be displayed. Your computer may not have enough memory to open the image, or the image may have been corrupted. Restart your computer, and then open the file again.
If the red x still appears, you may have to delete the image and then insert it again.
Mazzaetal.,2004
OlfactoryEpithelium
OlfactoryBulb
Olfactorycortex
Simões-de-Souza&Roque,2004
Escalaespacial:macrooumicroscópica
Global:connec/vitybetweencor/calareas
Local:connec/vitybetweenneuronsincor/callayers/
• A figura abaixo ilustra o padrão geral de conexões excitatórias
Short-scale cortical connectivity: layers
columns
corticais.
Connec/vitybetweenneuronsinnon-cor/calstructures(e.g.hippocampusdentategyrus)
1568
DYHRFJELD-JOHNSEN ET AL.
NETWORK REORGANIZATION IN EPILEPSY
TABLE
1.
1569
Connectivity matrix for the neuronal network of the control dentate gyrus
Granule Cells
Mossy Cells
Basket Cells
Axo-axonic
Cells
MOPP
Cells
HIPP Cells
HICAP Cells
IS Cells
Downloaded from http://jn.physiology.org/ a
Downloaded from http://jn.physiology.org/ at CAPE
Granule cells
X
9.5
15
3
X
110
40
20
(1,000,000)
X
7–12
10–20
1–5
X
100–120
30–50
10–30
ref. [1–5]
ref. [6]of
ref. [7]
ref. [6–9]
ref. [6,7,9]
ref. [6]
ref. [4,10,11] ref. [4,7,10,11]
ref. [7]
FIG. 1. Schematic
of the basic circuitry
the dentate gyrus
andcells
the changes to 32,500
the netMossy
350
7.5
7.5
5
600
200
X
work during sclerosis.
A: relational30,000–35,000
representa(30,000)
200–500
5–10
5–10
5
600
200
X
tion of the healthy
the
ref.dentate
[11] gyrus illustrating
ref. [4,11–13]
ref. [12,13]
ref. [13]
ref. [13]
ref. [14]
ref. [12,13]
ref. [12,13]
ref. [15]
network connections between the 8 major cell
Basketcell;
cellsBC, basket cell;
1,250
75
35
X
X
0.5
X
X
types: GC, granule
MC,
(10,000)
1,000–1,500
50–100
20–50
X
X
0–1
X
X
mossy cell; AAC, axo-axonic cells; MOPP,
[16,17] with axons
ref. [4,16–19]
ref. [11,16,17,19]
ref. [16,17,20,21]
ref. [18]
ref. [18]
ref. [18]
ref. [18]
ref. [10,20]
molecular layer ref.
interneurons
in perforant-path termination
zone;
interAxo-axonic
cellsHIPP, hilar
3,000
150
X
X
X
X
X
X
neurons with axons
in perforant-path
termina(2,000)
2,000–4,000
100–200
X
X
X
X
X
X
tion zone; HICAP,
axref. hilar
[4,22]interneurons
ref.with
[4,18,22]
ref. [4,5,11,14,23]
ref. [5,18]
ref. [5,18]
ref. [5,18]
ref. [5,18]
ref. [5,18]
ref. [5,18,19]
ons in the commissural/associational pathway
MOPP
7,500
X
40
1.5
7.5
X
7.5
X
termination zone;
andcells
IS, interneuron selective
5,000–10,000
X
30–50
1–2
5–10
X
5–10
X
cells. Schematic(4,000)
shows the characteristic
locaref.cell
[11,14]
[14]
ref. [14,24]
ref. [14,25]
ref. [14,26] ref. [14,25] ref. [14,20,25]
ref. [14,25]
ref. [14,15]
tion of the various
types within theref.
3 layers
of the dentateHIPP
gyrus.
Note, however, that
this
cells
1,550
35
450
30
15
X
15
X
diagram does not
indicate the topography
of
(12,000)
1,500–1,600
20–50
400–500
20–40
10–20
X
10–20
X
axonal connectivity
(present in both
strucref. [11]
ref.the[4,11,20]
ref. [4,11,12,27,28]
ref. [4,11,20]
ref. [20,25]
ref. [25]
ref. [14,20,25]
ref. [25]
ref. [15,20]
tural and functional dentate models) or the soHICAP cells
700
35
175
X
15
50
50
X
matodendritic location of the synapses (incor(3,000)
700
30–40
150–200
X
10–20
50
50
X
porated in the functional network models). B1:
[5,29,30]
ref. [4,11,20]
ref. [20]
ref. [4,11,20]
ref. [20]
ref. [14,20]
ref. [20]
ref. [20]
schematic of theref.
excitatory
connectivity
of the
IS cells
X
7.5
X
X
7.5
7.5
450
healthy dentate
gyrus is illustrated (onlyX cell
X
X
5–10
X
X
5–10
5–10
100–800
types in the hilus(3,000)
and granule cells are shown).
Note that the granule
cell axons (the
ref. [15,29,30]
ref.mossy
[15]
ref. [15]
ref. [15,19]
ref. [15]
ref. [19]
ref. [19]
ref. [15]
fibers) do not contact other granule cells in the
healthy network.Cell
B2: numbers
schematicand
of connectivity
the dentate values were estimated from published data for granule cells, Mossy cells, basket cells, axo-axonic cells, molecular layer
gyrus at 50% sclerosis
shows
theaxons
loss (indicated
interneurons
with
in perforant-path termination zone (MOPP), hilar interneurons with axons in perforant-path termination zone (HIPP), hilar interneurons
by the large ✕with
symbols)
the population
axons of
in half
the commissural/associational
pathway termination zone (HICAP), and interneuron-selective cells (IS). Connectivity is given as the number of
Arquiteturamicroscópica(córtex)
• Subdivisãodoneocórtexem6camadas
• Ascamadasdiferememtermosde
densidadese/posdecélulas
Abeles,1991
Tuckwell(2006)
Arquiteturamicroscópica(córtex)
Arquiteturamicroscópica(córtex)
conexõesaferenteseeferentes
• Aferentes
– Externas(não-locais).Origem:
• Tálamo(aferentestálamo-cor/cais)
principalmenteparaacamadaIV
• Outrasáreascor/cais(aferentescór/cocor/cais)viamatériabranca
principalmenteparaascamadas
superficiais
– Entradasvindasdeneurônioscor/caisna
vizinhançalocal
• Eferentes
– Eferentescór/co-cor/caisdosneurônios
piramidaisdascamadasII/III
– Eferentescór/co-talâmicosdos
neurôniospiramidaisdacamadaVI
– Axôniosdeneurôniospiramidaisgrandes
dacamadaVparaotroncoencefálicoea
medulaespinhal
Abeles,1991
Arquiteturamicroscópica:conexõesver/cais
Cadafiguramostraaproporçãodototaldesinapsesnocórtexvisualprimáriodogatoque
existeentreos/posdeneurôniosindicados.Osnúmerostotaisdesinapsesdacada/po
tambémestãoindicados.
EàE
13.6x1010
EàI
2.1x1010
IàE
IàI
2.4x1010
0.4x1010
Binzeggeretal.,2004
Arquiteturamicroscópica(córtex):
Conexõeshorizontais
• Sinapseslocais:feitasporcolateraisdosaxôniosemumraiode
cercade0,5mm(todosos/posdeneurônios).
• Conexõesintrínsecasdelongoalcance:feitasporneurônios
piramidais,alcançamdistânciasdeváriosmilímetrospassando
pelamatériacinzenta.
• Conexõesextrínsicasdelongoalcance:feitasporneurônios
piramidaisatravésdamatériabranca.
Vogesetal.,2010
Arquiteturamicroscópica(córtex):
Conexõeslocais
• Aprobabilidadedeumaconexãosináp/caentredois
neurônioscor/caisadjacentescaiparazeroauma
distânciahorizontaldecercade0,5mm
Hellwig,2000
Boucseinetal.,2011,2000
Arquiteturamicroscópica(córtex):
Conexõesdelongoalcance
• Asconexõesintrínsecasdelongoalcanceformamum
padrãode“retalhos”:osneuroniospiramidaisprojetamseus
axôniosparaagrupamentoscelulares
Vogesetal.,2007
Lundetal.,2003
Arquiteturamacroscópica
Felleman&VanEssen,1991
Arquiteturamacroscópica(córtex)
§ Asáreascerebraissãotratadascomonósdarede
§ Asconexõesentreosnóssãoreveladaspordiferentes
técnicas:neuroimagem,traçadoresradioa/vos,etc
Bressler&Menon,2010
ImagemporTensordeDifusão
(diffusiontensorimaging–DTI)
• Cadanócorrespondeauma
áreadocérebro
• Nãopermiteadeterminação
dasconexõesdentrodecada
área
• Conexõesbinárias(há/não
há,semponderação)
• Nãopermitedeterminaros
atrasosnasconexões
• Nãopermitedis/nguirentre
conexõesausentesou
desconhecidas
Honeyetal.,2007
Arquiteturamacroscópica(córtex)
• Estruturahierárquicaemodular
• “Clubedericos”
Meunieretal.,2010
VandenHeuvel&Sporns,2011
Juntandotudo:modelosde
neurôniosesinapsesemumarede
comumadadaarquitetura
(algunsmodelosselecionados)
RedesdeErdős–Rényi:modelodeBrunel
• NeurôniosLIF:80%excitatórios,20%inibitórios
• Conec/vidadeesparsa(#conexõesk<<#neurôniosN)
NE=10.000
NI=2.500
p=0,1
CE=conexões
recebidaspelas
célulasE=1.000
CI=250
Tiposdea/vidadeemum
modeloderedeneuronal
(a) Assíncronaregular:aa/vidadeda
populaçãoéaproximadamente
constanteeosneurônios
individuaisdisparamdeforma
regular;
(b) Síncronaregular:Tantoa
a/vidadepopulacionalcomoados
neurôniosindividuaissão
oscilatórias;
(c) Síncronairregular:aa/vidade
populacionaloscilaeosneurônios
individuaisdisparamdeforma
irregular;
(d) Assíncronairregular:aa/vidade
populacionaléaproximadamente
constanteeosneurônios
individuaisdisparamdeforma
irregular.
Vogelsetal.(2005)
Estadobalanceado
• Ocórtexoperaemumestado
balanceadoemqueosvalores
médiosdascorrentesdeentrada
excitatóriaeinibitóriaemum
neurôniosecancelam
mutuamente.
• Osdisparosdeumneurôniosão
causadosporflutuaçõesemtorno
daentradamédialíquida.
• Issoexplicaosdisparosirregulares
dosneurônios(parecendoruído)no
estadoassíncronoirregular(AI).
RedesdeErdős–Rényi:modelodeVogelseAbboZ
N=10.000neurôniosLIF;p=0,02;#excitatórios/#inhibitórios=4:1
Plas/cidadeemummodelodesistemasensorial:
Estudossobrelesões
Modelocomarquiteturacor/calmicroscópica
Potjans & Diesmann (2014)
Modelocomarquiteturacor/calmicroscópica
(modelosdeneurôniosindividuaisbiofisicamentedetalhados)
hZp://bluebrain.epfl.ch/
• ProjetoBlueBrain:modelodeumacolunacor/caldorato
jovemcom10.000neurôniosreconstruídosmorfologicamente
interconectadospor3x107sinapses
• RodanosupercomputadorparaleloBlueGene:8912
processadores.Umasimulaçãodeumdadotempobiológico
leva2xmaisqueessetempoparasersimulada.
Modeloquecombinainformaçãomacroemicroscópica:
dadosdeDTI,estruturamicroscópicaeneurôniosque
reproduzemdiferentespadrõesdedisparo
Esquerda.Propagaçãodeondasno
modelo.Disparosdosneurônios
excitatórios(inibitórios)indicados
porpontosvermelhos(pretos).
Direita.Sensibilidadeàadiçãodeum
únicodisparo.
Izhikevich&Edelman,2008
Modelosparaestruturasnãocor/cais:
girodenteadodohipocampo
•
•
•
•
•
Modeloestruturalemescala1:1dogirodenteado(GD)dorato(~1milhãodeneurônios)
Modelofuncionalemescala20:1doGDcommaisde50.000modelosdeneurônios
compar/mentaisreduzidos
Usadoparaavaliaroefeitodediferentesníveisdeescleroseneuralsobreaexcitabilidadedo
GD
GDnormaltemestruturademundopequeno(baixoLealtoC)
Aescleroseaumentaascaracterís/casdemundopequenodarede(LdiminuieCaumenta)
J Neurophysiol 97: 1566 –1587, 2007.
First published November 8, 2006; doi:10.1152/jn.00950.2006.
Topological Determinants of Epileptogenesis in Large-Scale Structural and
Functional Models of the Dentate Gyrus Derived From Experimental Data
Jonas Dyhrfjeld-Johnsen,1,* Vijayalakshmi Santhakumar,1,* Robert J. Morgan,1 Ramon Huerta,2 Lev Tsimring,2
and Ivan Soltesz1
1
Department of Anatomy and Neurobiology, University of California, Irvine; and 2Institute for Nonlinear Science, University of California,
San Diego, California
Colunas:neurôniopré-sináp/cos
Linhas:neurôniospós-sináp/cos
Dyhr•eldetal.(2007)
www.sisne.org/lascon
Latin American School on Computational Neuroscience
LASCON
NeuroMat
Research,Innova/onandDissemina/on
CenterforNeuromathema/cs
hZp://neuromat.numec.prp.usp.br