Aula: 30
Temática: Metabolismo dos lipídeos – parte II
Hoje continuaremos a estudar o metabolismo dos lipídeos. Boa aula!
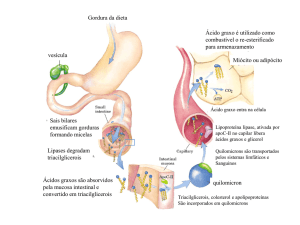
Metabolismo das Gorduras no Fígado:
O primeiro estágio do metabolismo das gorduras ocorre no fígado e consiste
na decomposição dos triglicerídeos em glicerol e ácidos graxos por meio das
lipases. Em seguida o glicerol é transformado em hidratos de carbono,
enquanto os ácidos graxos sofrem oxidação (degradação) começando no
grupo carboxílico e levando o CO2 e água. No fígado podem ocorrer diversas
vias metabólicas:
O glicerol pode seguir para a glicólise depois de oxidado a dihidroxiacetona
fosfatada na face externa da membrana interna da mitocôndria. Os dois
elétrons libertados nesta oxidação são recebidos pela ubiquinona (Q), que os
transfere para a cadeia transportadora de elétrons. Os ácidos graxos terão um
destino diferente: a ß-oxidação que ocorre nas mitocôndrias das células.
Oxidação dos ácidos graxos (ß-oxidação): Ácidos graxos, em animais
superiores, são degradados por uma série de reações, algumas das quais
oxidativas, que resultam na quebra dos ácidos em fragmentos de dois
carbonos.
A ß-oxidação dos ácidos graxos ocorre na matriz mitocondrial (parte interna da
mitocôndria), onde todas as enzimas associadas com o sistema da ß-oxidação
estão localizadas (nas membranas internas e na matriz da mitocôndria). Antes
de entrarem na mitocôndria, os ácidos graxos são ativados. A reação de
ativação ocorre no citoplasma celular e consiste na sua transformação em acilCoA (figura 6).
BIOQUÍMICA
Fig. 6 – Mecanismo de transporte dos ácidos graxos do citoplasma para o local da ß-oxidação
na mitocôndria. Carnitina: acil-CoA-transferase - I (face externa) e carnitina: acil-CoAtransferase - II (face interna); duas enzimas distintas que catalisam a mesma reação.
O esquema da figura 3 mostra a translocação do acil-CoA de fora da
mitocôndria para o local interno do sistema de ß-oxidação. A enzima-chave
nesse processo é a carnitina -aci1- CoA-transferase.
Observar-se, no esquema acima (fig. 3), que apenas uma molécula de ATP é
necessária para ativar um ácido graxo à sua total degradação em acetil-CoA,
qualquer que seja o número de átomos de carbono, da cadeia de
hidrocarboneto, o que significa uma grande economia.
Quatro enzimas catalisam a ß-oxidação, além da acil-CoA-sintetase, que está
envolvida no passo inicial de ativação. Então, quando acetil-CoA é produzida
na quebra de ácidos graxos, ela pode ser subseqüentemente oxidada a CO2 e
H2O através das enzimas do ciclo do ácido tricarboxílico que estão localizadas
como enzimas solúveis na matriz mitocondrial.
A ß-oxidação dos ácidos graxos consiste num ciclo de 3 reações sucessivas
(figura 7).
Fig. 7 - Oxidação do ácidos graxas (ß-oxidação). Cada ciclo (1) a (4) e (1’) a (4’) resulta na
libertação de um fragmento de dois carbonos ligado a CoA. As enzimas implicadas são: (1)
acil-CoA desidrogenase (2) enoil hidrase (3) f3-hidroxilacil-desidroge-nase e (4) f3cetoaciltiolase (retirado de BENNET & FREDEN, 1987).
Essas reações de ß-oxidação, são idênticas: desidrogenação, hidratação da
ligação dupla formada e oxidação do álcool a uma cetona, pela ação da enzima
tiolase, liberta-se acetil-CoA, e um acil-CoA com menos dois carbonos que o
acil-CoA original.
A repetição do ciclo permite a degradação total de um ácido graxo de cadeia
por um acetil-CoA, que pode entrar no ciclo de Krebs, onde é completamente
oxidado a CO2.
BIOQUÍMICA
Degradação peroxissomal de ácidos graxos: Os peroxissomas são pequenas
organelas citoplasmáticas onde ocorre a ß-oxidação de ácidos graxos de
cadeia longa, de forma a facilitar a sua degradação subseqüente pela
mitocôndria. Esses ácidos graxos de cadeia longa se difundem livremente para
dentro do peroxissoma, não precisando ser transportados pela carnitina. Porém
essa degradação não é completa e tem que ser terminada na mitocôndria.
A β-oxidação é a principal, ou pelo menos a mais conhecida via de oxidação
dos ácidos graxos, embora haja outras vias como:
α-oxidação: Esse sistema, observado inicialmente em sementes e tecidos
verdes de plantas, é também encontrado em células do cérebro e do fígado.
Neste caso, apenas ácidos graxos livres funcionam como substratos (oxigênio
molecular está envolvido indiretamente). O produto pode ser um ácido graxo
com um átomo de carbono a menos.
ω-oxidação: Essa reação envolve uma hidroxilação inicial de um grupo de metil
terminal a álcool primário e subseqüente oxidação a ácido carboxílico. Dessa
maneira, hidrocarbonetos de cadeias retas são oxidados em ácidos graxos, e
estes, por sua vez, são β-oxidados a acetil-CoA. Essas séries de reações, que
à primeira vista pareciam de pouco interesse, assumiram agora um papel
extremamente importante, de limpeza, pela biodegradação bacteriana, de
detergentes derivados de ácidos graxo e mais importante ainda, das grandes
quantidades de óleo expelida nas superfícies dos oceanos. O mecanismo de
oxidação dos óleos é feito principalmente pelo sistema da ω-oxidação.
Para ácidos graxos insaturados, são necessárias duas importantes enzimas
adicionais
(∆3cis
e
∆2trans-enoil-CoA-isomerase
e
D-3-OH-acil-CoA-
epimerase) que torna possível a β-oxidação desses ácidos.
Assim, os ácidos graxos, em animais superiores, são degradados por uma
série de reações, as mais conhecidas, são as oxidativas, que resultam na
quebra dos ácidos em fragmentos de dois carbonos, acetil CoA.
BIOQUÍMICA
FORMAÇÃO DE CORPOS CETÔNICOS (CETOGÊNESE):
Sob certas condições metabólicas em que existe aumento da oxidação dos
ácidos graxos, o fígado produz acetoacetato e 3 hidroxibutirato. O acetoacetato
espontaneamente é descarboxilado até acetona. Estas três substâncias são
conhecidas coletivamente como corpos cetônicos. A concentração de corpos
cetônicos total no organismo bem alimentado não excede 0,2 mol/l. Para
formação dos corpos cetônicos, inicialmente ocorre a condensação de 2
unidades de 2 carbonos formadas na α-oxidação pela enzima tiolase, formando
o acetoacetil-CoA, que vai dar origem aos corpos cetônicos. Outra via de
formação deste composto é direta, ao final da α-oxidação. Em seguida ocorre a
formação do acetoacetato, pela condensação do acetoacetil-CoA com outra
molécula de acetil-CoA, formando hidroximetilglutaril-Coa (HMG-CoA). Depois,
ocorre a clivagem da acetil-CoA da molécula de HMG-CoA, liberando
acetoacetato (as enzimas envolvidas estão na mitocôndria).
Quantidades aumentadas de corpos cetônicos no sangue e na urina são
denominadas de cetonemia e cetonúria, respectivamente, e a condição geral é
denominada de cetose. A forma mais simples de cetose ocorre durante o jejum
prolongado e envolve depleção de carboidratos disponíveis com mobilização
de ácidos graxos livres. Outra condição associada é a diabetes mellitus.
Não se esqueça de enviar suas dúvidas para nossa tutoria. Até a
próxima aula!
BIOQUÍMICA