Seleção Natural
DNA codifica informações que em conjunto
com ambiente influenciam fenótipo
• Seleção Natural: Sobrevivência e reprodução
diferencial de indivíduos na população
• Valor Adaptativo: progênie gerada que sobrevive
e reproduz na próxima geração
• Seleção natural requer variação herdável para
valor adaptativo (não basta sorte!)
• Define valor adaptativo esperado de um genótipo
em um ambiente específico
Entre características que podem ser influenciadas por
respostas geneticamente determinadas ao
ambiente estão:
1.
A Viabilidade no ambiente
2.
Uma vez vivo, o sucesso reprodutivo no ambiente
3.
Uma vez vivo e tendo reproduzido, a fertilidade
ou fecundidade no ambiente.
Estes são os componentes do valor adaptativo
Seleção Natural
O ambiente influencia a probabilidade de vários
genótipos replicarem o seu DNA.
Seleção Natural
Seleção favorece um caráter que aumente o valor
adaptativo.
Enquanto valor adaptativo for herdável, as diferenças na
contribuição à próxima geração induzidas pela
seleção natural irão alterar o pool gênico.
Indivíduos com maior valor adaptativo tendem a passar
mais genes para a próxima geração.
Seleção natural tende a aumentar o valor adaptativo
médio da população.
Enquanto valor adaptativo for herdável, as diferenças na
contribuição à próxima geração induzidas pela
seleção natural irão alterar o pool gênico.
Este aumento do valor adaptativo médio é a adaptação
ao meio ambiente.
Valor Adaptativo (“Fitness”)
• Embora seja individual, definimos em relação a grupos
de indivíduos:
– De mesmo genótipo
– De mesmo fenótipo
• Contribuição genética coletiva à próxima geração.
• Evitem usar “a sobrevivência do mais apto”.
• Porquê?
– Vários argumentam que é tautológico.
• Quem são os mais aptos?
• Os que sobrevivem!!!
– A maioria das pessoas não compreende os termos
“sobrevivência” e “apto”.
Seleção Natural em um locus único em um deme se reproduzindo ao acaso
Freqüências zigóticas
Meio Ambiente
Viabilidades
Freqüências no adulto ∝
Meio Ambiente
Prob. acasalamento
AA
ZAA
Aa
ZAa
aa
Zaa
lAA
lAa
laa
AA
ZAAlAA /l
Aa
ZAalAa /l
mAA
mAa
AA
Freqüências em adultos que
∝ mAAZAAlAA
acasalaram
/ lm
Meiose
Pool Gênico
1*bAA
maa
aa
Aa
mAa ZAalAa /lm
1/ b
2 Aa
aa
Zaalaa /l
maa Zaalaa /
lm
1/ b
2 Aa
1*bAa
A
a
p’= (bAAmAAZAAlAA +
1/2bAamAaZAalAa) / w
q’= (baamaaZaalaa +
1/2bAamAaZAalAa) / w
1
Façamos wAA = bAAmAAlAA; wAa = bAamAalAa; waa = baamaalaa
Freqüências zigóticas
AA
ZAA
Meio Ambiente
Valor Adaptativo
Aa
ZAa
WAA
WAa
Waa
AA
Freqüências em adultos que
∝ Z W
acasalaram pelo # prole
AA
AA
Aa
ZAaWAa
aa
ZaaWaa
Aa
ZAaWAa / W
aa
ZaaWaa / W
Converter para Freq ao dividir por
μ = W = p2WAA+2pqWAa+q2Waa
Freqüência de adultos
que se acasalaram
Meiose
Pool Gênico
AA
ZAAWAA / W
1
1/
2
1/
2
A
p’= ZAAWAA/W +
½ZAAWAa/W
Pool Gênico
aa
Zaa
1
A
a
p’= p2 WAA/W + pqWAa/W
q’= q2 Waa/W + pqWAa/W
p’= p2 WAA/W + pqWAa/W
=( p2 WAA+ pqWAa)/W
= p(pWAA+ qWAa)/W
a
q’= ZaaWaa/W + ½ZAaWAa/W
Quando ocorrerá evolução?
Pool Gênico
A
a
p’= p2 WAA/W + pqWAa/W
q’= q2 Waa/W + pqWAa/W
Δp = p’ - p
= p(pWAA+ qWAa)/W - p
= p[pWAA+ qWAa)/W - 1]
p’= p2 WAA/W + pqWAa/W
=( p2 WAA+ pqWAa)/W
Δp = p[pWAA+ qWAa- W]/W
p’ = p(pWAA+ qWAa)/W
Quando ocorrerá evolução?
Quando ocorrerá evolução?
Observe que W = W(p+q)=pW+qW
Observe que W = W(p+q)=pW+qW
Δp = p[pWAA+ qWAa- W]/W
Δp = p[pWAA+ qWAa- W]/W
= p[pWAA+ qWAa- pW - qW]/W
= p[pWAA+ qWAa- pW - qW]/W
=p[p(WAA-W)+ q(WAa-W)]/W
=p[p(WAA-W)+ q(WAa-W)]/W
Desde que p e W serão sempre > 0, esta é a única parte da
equação que pode mudar de sinal e portanto determinar
a direção da evolução sob seleção natural.
2
Pool Gênico
A
p’= ZAAWAA/W +
½ZAAWAa/W
Δp
a
q’= ZaaWaa/W + ½ZAaWAa/W
= [ZAA(WAA- W) + ½ZAa(WAa -W)]/W
Quando ocorrerá evolução?
O que é:
p(WAA-W)+ q(WAa-W)?
= p[ZAA(WAA- W) + ½ZAa(WAa -W)]/pW
= p/W[ZAA(WAA- W) + ½ZAa(WAa -W)/p]
Quando ocorrerá evolução?
Quando ocorrerá evolução?
O que é:
O que é:
p(WAA-W)+ q(WAa-W)?
p(WAA-W)+ q(WAa-W)?
Fenótipo médio do valor
adaptativo
Quando ocorrerá evolução?
Desvios genotípicos para o fenótipo
médio do valor adaptativo
Quando ocorrerá evolução?
O que é:
O que é:
p(WAA-W)+ q(WAa-W)?
p(WAA-W)+ q(WAa-W)?
Freqüência com que os desvios no valor
adaptativo serão encontrados na população
Este é o excesso médio (Average Excess) do alelo A
para o fenótipo do valor adaptativo
3
Quando ocorrerá evolução?
Quando ocorrerá evolução?
Δp = paA/W
Δp = paA/W
Seleção Natural será uma força evolutiva
quando:
p ≠ 0 ou p ≠ 1 (ou seja, quando existir
variação genética), e
aA ≠ 0 (ou seja, quando existir variação
herdável para o valor adaptativo)
Para se entender seleção natural
Esta é a equação fundamental da Seleção
Natural para genótipos mensurados.
Apenas o componente herdável do valor
adaptativo pode promover mudanças
evolutivas por seleção natural.
Quando ocorrerá evolução?
Δp = paA/W
PENSE COMO UM GAMETA!
Seleção Natural será uma força evolutiva
quando:
p ≠ 0 ou p ≠ 1 (ou seja, quando existir
variação genética), e
aA ≠ 0 (ou seja, quando existir variação
herdável para o valor adaptativo
Teorema Fundamental de Fisher da
seleção natural
Δp = paA/W
A equação acima descreve a evolução por seleção
natural em um locus único.
O equivalente para loci quantitativos é uma equação a
que Fisher se referiu como FTNS:
descreve como a seleção natural opera sobre o
fenótipo “valor adaptativo”quando este é considerado
como um caráter herdável, mas geneticamente não
mensurado.
FFTNS
Como não temos genótipos sendo medidos, focamos em
fenótipos.
Neste caso não podemos usar definição “genética” de
valor adaptativo, e sim definição “ecológica”, em
que definimos W para uma classe fenotípica.
Desta forma, temos que o fenótipo médio do caráter x
será:
μ = ∫ xf ( x)dx
x
4
FFTNS
FFTNS
w = ∫ w ( x)f ( x)dx
μs =
x
Será o fenótipo médio da população - o mesmo já
definido anteriormente.
Temos então que o fenótipo médio do caráter x será:
μs =
x
w
Considerando que R=h2S, S=(μs - μ) e R = (μo - μ), que
x= w e μ = w e mais algumas coisinhas:
∫ xw ( x)f ( x)dx
x
w
FFTNS
S = μ −μ =
∫ xw ( x)f ( x)dx
σ2 +w
w
2
FFTNS
2
−w=
σ2 +w −w
2
w
=
σ2
w
Acima temos a medida S da intensidade de seleção para
o valor adaptativo.
No caso de x = w, temos que a resposta a seleção R será
Δw e:
R=h2S
⎛ σ 2 ⎞⎛ σ 2 ⎞ σ 2
Δ w = ⎜⎜ a2 ⎟⎟⎜⎜ ⎟⎟ = a
⎝ σ ⎠⎝ w ⎠ w
Implicações do FFTNS
Δw =
σ a2
w
• Seleção natural apenas pode operar quando existe
variação genética para valor adaptativo na população
• Os únicos efeitos no valor adaptativo que influenciam
a resposta à seleção natural são aqueles transmitidos
através do gameta
Δw =
σ a2
w
Acima temos o Teorema Fundamental de Fisher sobre
Seleção Natural, que diz que:
A mudança no valor adaptativo médio da população
será proporcional à variância genética aditiva do valor
adaptativo na população. Equivalente de caracteres
não mensurados à:
Δp = paA/W
Implicações do FFTNS
Δw =
σ a2
w
• A resposta adaptativa representa uma interação do
valor adaptativo com a estrutura populacional
– Resposta à seleção natural depende do contexto
populacional. Mesmo o completo conhecimento do valor
adaptativo de cada indivíduo não permite o conhecimento
da resposta à seleção
5
Implicações do FFTNS
Δw =
Implicações do FFTNS
σ a2
Δw =
w
• Equilíbrio seletivo apenas pode ocorrer quando todos
os excessos e efeitos médios forem iguais a zero; ou
seja, quando os gametas tiverem o mesmo impacto na
média.
– Isso não implica que não exista variação genética, mas sim
que não existe variação genética aditiva para o valor
adaptativo.
– Caracteres relacionados ao valor adaptativo devem ter
menor variação genética aditiva - Merilä & Sheldon (1996)
σa
– O equilíbrio do ponto anterior deve se referir sempre a um
ótimo populacional.
– Uma população não pode reduzir seu valor adaptativo
médio.
Implicações do FFTNS
Δw =
Implicações do FFTNS
2
w
• Seleção natural apenas leva populações a soluções
adaptativas locais e não necessariamente ao estado
adaptativo com maior valor adaptativo. S.N. pode na
verdade impedir que tal estado surja.
σ a2
w
• Seleção natural normalmente não otimiza, mesmo em
nível local, qualquer caráter individual que não o
próprio valor adaptativo, mesmo que tal caráter
contribua ao valor adaptativo de forma positiva.
– Na maioria dos casos, investigamos caracteres relacionados
a valor adaptativo, e não o próprio valor adaptativo. Muitas
vezes, o processo de adaptação nestes é considerado como
sendo igual ao do valor adaptativo.
w
• Seleção natural age aumentando o valor adaptativo
médio da população de uma geração a outra.
Implicações do FFTNS
Δw =
σ a2
Δw =
σ a2
w
• Seleção natural apenas
leva populações a
soluções adaptativas
locais e não
necessariamente ao
estado adaptativo com
maior valor adaptativo.
Implicações do FFTNS
• Seleção natural normalmente não otimiza, mesmo em
nível local, qualquer caráter individual que não o
próprio valor adaptativo, mesmo que tal caráter
contribua ao valor adaptativo de forma positiva.
Crow and Nagylaki mostram que:
weq = w( xeq ) + 1 2 w" ( xeq )σ eq2 ( x)
Portanto, xeq apenas será um valor que maximize w(x):
w" ( xeq )σ eq2 = 0
6
Implicações do FFTNS
w" ( xeq )σ
2
eq
=0
Em 2 situações apenas a seleção natural otimiza o valor de
um caráter que contribua ao valor adaptativo, mas que
não seja o valor adaptativo:
– Não exista variância fenotípica no equilíbrio [σ2eq(x) = 0];
– O caráter esteja relacionado ao valor adaptativo de uma forma
extritamente linear [w”(xeq) = 0]
Se existe não linearidade, e variação fenotípica, o valor do
caráter que aumenta o valor adaptativo não é o mesmo
que evolui por seleção natural. Seleção não otimiza
caracteres individuais, age no todo do indivíduo como
medido pelo valor adaptativo.
Implicações do FFTNS
• As mesmas condições restritivas necessárias para
otimizar um caráter individual tem que ser satisfeitas
para otimizar cada um dos dois caracteres contribuindo
ao valor adaptativo neste caso, e além disso, um outro
termo, que depende da covariância de x e y, também
deve ser 0.
• Pleiotropia pode criar uma covariância entre 2
caracteres, que pode ser inclusive negativa - mesmo
alelo estará associado a caracteres que têm efeito
oposto no valor adaptativo - como o alelo S.
Implicações do FFTNS
w" ( xeq )σ eq2 = 0
• O processo de adaptação pode resultar na evolução de
alguns caracteres aparentemente não adaptativos.
A equação acima apenas considera um caráter contribuindo
ao valor adaptativo, mas em geral vários contribuem.
Quando consideramos 2 caracteres, X e Y, de forma que
w(x,y) seja o valor adaptativo dos indivíduos com valores x
e y temos que o equivalente à equação acima será:
Implicações do FFTNS
• As correlações entre caracteres induzida por pleiotropia
são algumas vezes chamados de restrições do
desenvolvimento (developmental constraints).
– Sob certas circunstâncias, tais restrições são essenciais para a
evolução adaptativa. Wagner (1988) observou que seleção
natural se torna cada vez mais inefetiva à medida que o número
de caracteres envolvidos no valor adaptativo aumenta.
– O pico associado a uma adaptação complexa está presente em
apenas uma pequena porção do espaço genético possível.
– Restrições são necessárias para se manter versatilidade
adaptativa face à interdependências funcionais e complexidade
fenotípica
Implicações do FFTNS
Δw =
σ a2
w
• O curso da evolução adaptativa é fortemente
influenciado pela arquitetura genética.
– A arquitetura genética inclui pleiotropia, epistasia,
dominância e recessividade.
– A forma da paisagem adaptativa é fortemente influenciada
pelas regras de herança, transmissão e encontro de genes na
população, como vimos no exemplo da anemia falciforme
na África.
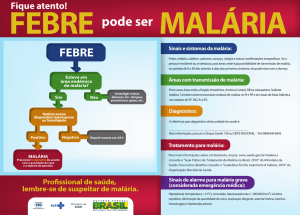
Anemia Falciforme na África
Um exemplo de seleção natural
7
Infecção de uma hemácia pelo
parasita da malária
A mutação
• Células falciformes são filtradas
preferencialmente no baço
• células infectadas com malária são
freqüentemente filtradas pois o processo de
“virar falciforme” ocorre antes do parasita
completar o ciclo de vida
• O alelo S é portanto, um alelo autossômico,
dominante para resistência à malária.
O fenótipo da anemia falciforme
A maioria de mortes devido a anemia falciforme
e malária ocorrem antes da fase adulta.
Lembrem-se que viabilidade é o fenótipo de
viver até a fase adulta!
• Em um ambiente sem malária, o alelo S é um alelo
recessivo para viabilidade uma vez que apenas os
homozigotos desenvolvem a anemia falciforme.
• Em um ambiente com malária, o alelo S é um
alelo sobredominante para viabilidade uma vez
que apenas os heterozigotos são resistentes à
malária e não desenvolvem anemia falciforme.
Distribuição de anemia falciforme e malária na África e no mundo
Duas complicações para esta simples
estória na África:
•
Malária epidêmica é recente na maioria da
África tropical úmida, e o processo de
adaptação à malária na África ainda não
está em equilíbrio.
8
Malária epidêmica na África
Malária epidêmica na África
ICELAND
ICELAND
MADAGASCAR
MADAGASCAR
Há cerca de 2000 anos
atrás, uma colônia
Malásia-Indonésia
estabeleceu-se em
Madagascar
Malária epidêmica na África
Esta colônia
introduziu a
complexa
agricultura Malásia
nesta região
A agricultura Malásia na África
Malária epidêmica na África
Esta agricultura foi
assimilada por povos
que falavam Bantu,
e se seguiu uma
grande expansão dos
Bantu na África há
cerca de 1500 anos
atrás.
Malária epidêmica na África
• É associada a uma agricultura de corte e
queima: Fornece habitat e sítios de criação
para Anopheles gambiae, o principal
mosquito vetor da malária.
• Resulta em alta densidade local de
populações humanas que são necessárias
para estabelecer e manter malária como
uma doença comum.
9
Duas complicações para esta simples
estória na África:
•
•
Malária epidêmica é recente na maioria da
África tropical úmida, e o processo de
adaptação à malária na África ainda não
está em equilíbrio.
Existe um terceiro alelo, Hemoglobina C,
também envolvido na adaptação à malária
na África.
A Mutação Hemoglobina C
Hb-S
Hb-A
GTG
GAG
Valina
Ácido Glutâmico
Hb-C
AAG
6o Códon
Lisina
Hb-A, S e C
Genótipos
AA AS
Anemia
Não Não
SS
Hb-A, S e C
AC
CS
CC
Sim
Sim
Não
NÃO
(Severa)
(Leve)
Resistência à
Não Sim
Malária
Sim
Não
Sim
Sim
Viabilidade
sem Malária
0.2
1
0.71
1
1
1
Genótipos
AA AS
Anemia
Não Não
Anemia
AC
CS
CC
Resistência à
Não Sim
Malária
Sim
Não
Sim
Sim
Viabilidade
sem Malária
0.2
1
0.71
1
1
CC
Sim
Sim
Não
NÃO
(Severa)
(Leve)
Sim
Não
Sim
Sim
Viabilidade
sem Malária
0.2
1
0.71
1
1
1
Hb-A, S e C
Sim
Sim
Não Não
Não
NÃO
(Leve)
(Severa)
1
CS
Resistência à
Não Sim
Malária
Hb-A, S e C
SS
AC
Os alelos A e S definem uma doença genética autossômica
recessiva: A seleção irá garantir que continue rara, mas será difícil
de eliminá-la em uma população acasalando-se ao acaso.
Estes valores podem ser usados como valores adaptativos
relativos
Genótipos AA AS
SS
Os alelos A e C definem um conjunto de alelos neutros em um
ambiente sem malária: Suas freqüências são determinadas pela
deriva genética, migração e mutação.
Genótipos AA AS
Anemia
SS
AC
CS
CC
Sim
Sim
Não Não
Não
NÃO
(Leve)
(Severa)
Resistência à
Não Sim
Malária
Sim
Não
Sim
Sim
Viabilidade
sem Malária
1
0.2
1
0.71
1
1
0.2
0.89
0.7
1.31
1
Viabilidade
0.89
com Malária
Viabilidades relativas observadas na África Tropical
10
Hb-A, S e C
• CC é sem dúvida o melhor genótipo!
• Se seleção natural é a “sobrevivência do
mais apto”, então a freqüência do alelo C e
do genótipo CC deve aumentar.
• Ao contrário do que se diz, a seleção natural
não é a “sobrevivência do mais apto.”
• Seleção natural é variação herdável no
valor adaptativo, portanto, pense como
um gameta: Qual gameta terá o maior
excesso médio (ou seja, terá filhos que
serão em média os melhores)?
Valor adaptativo médio inicial após
transição para a agricultura Malásia
Pool gênico inicial pré-Malária
pS=.005 pC=.005
A
pA = 0.99
Fenótipos iniciais após transição para a
agricultura Malásia
pS=.005 pC=.005
pS=.005 pC=.005
A
pA = 0.99
Com acasalamento ao acaso,
fenótipo médio = W = 0.901
Fenótipos iniciais após transição para a
agricultura Malásia
Genótipos
AA AS SS AC CS CC
Viabilidade com
0.9
1
0.2 0.9 0.7 1.3
Malária
Desvios Genotípicos
-.001 .099 -.701 -.001 -.201 .399
(W = 0.901)
aA = -0.0005
aS = 0.0935
aC = 0.0000
A
pA = 0.99
Genótipos
Viabilidade c/
Malária
Desvios genotípicos
(W = 0.901)
AA
AS
SS
AC
CS
CC
0.9
1
0.2
0.9
0.7
1.3
-.001 .099 -.701 -.001 -.201 .399
Fenótipos iniciais após transição para a
agricultura Malásia
Δpx = pxax/W
aA = -0.0005
aS = 0.0935
aC = 0.0000
A resposta adaptativa
inicial a um ambiente com
malária é:
diminuir A,
aumentar S,
deixar C do mesmo jeito
11
Pool Gênico após várias gerações de
seleção em um ambiente com malária
Pool Gênico após várias gerações de
seleção em um ambiente com malária
pC=.005
A
pA = 0.95
W = 0.907
S
pS = 0.045
pC = 0.005
Pool Gênico após várias gerações de
seleção em um ambiente com malária
Após a resposta adaptativa
inicial a malária, seleção
natural continua a
diminuir A
aumentar S, e
diminuir C
aA = -0.003
aS = 0.055
aC = -0.014
Um equilíbrio por seleção natural irá
ocorrer apenas quando Δp = 0 para todos
os alelos.
A
pA = 1-pS
Genótipos
S
pS
AA AS SS
Viabilidade c/ Malária 0.9
1
A
pA = 0.95
Genótipos
Viabilidade com
Malaria
Desvios genotípicos
(W = 0.907)
S
.045
AA
AS
SS
AC
CS
CC
0.9
1
0.2
0.9
0.7
1.3
-.007 .093 -.707 -.007 -.207 .393
Pool Gênico após várias gerações de
seleção em um ambiente com malária
pC ≈ 0
A
pA ≈1-pS
Genótipos
S
pS
AA AS SS AC CS CC
Viabilidade c/ Malária 0.9
1
0.2 0.9 0.7 1.3
À medida que pS aumenta, W aumenta e os desvios
genotípicos ficam cada vez mais negativos. Portanto, a
seleção natural elimina o alelo C.
Um equilíbrio por seleção natural irá
ocorrer apenas quando Δp = 0 para todos
os alelos.
A
pA = 1-pS
S
pS
aA = (1-pS)(0.9-W)+pS(1-W) = aS = (1-pS)(1-W)+pS(0.2-W)
0.2
aA = (1-pS)(0.9-W) + pS(1-W) = 0 = aS = (1-pS)(1-W) + pS(0.2-W)
Coeficientes de Seleção (s) – mede o valor do valor
adaptativo do genótipo em relação a alguma referência
12
Um equilíbrio por seleção natural irá
ocorrer apenas quando Δp = 0 para todos
os alelos.
A
pA = 1-pS
As freqüências alélicas no equilíbrio são
mantidas por seleção natural em um
polimorfismo balanceado
S
pS
aA = (1-pS)(0.9-W)+pS(1-W) = aS = (1-pS)(1-W)+pS(0.2-W)
(1-pS)(0.9)+pS(1) = (1-pS)(1)+pS(0.2)
0.9+0.1pS = 1-0.8pS
0.9pS = 0.1
pS = 0.1/0.9 = 0.11
A
pA = 0.89
Portanto, em equilíbrio, pS = 0.11 e pA=0.89
As freqüências alélicas no equilíbrio são
mantidas por seleção natural em um
polimorfismo balanceado
A
pA = 0.89
S
pS=0.11
Este balanço ocorre porque quando pS < 0.11, aS > 0
(resistência à malária domina o excesso médio), e
quando pS > 0.11, aS < 0 (a anemia domina o excesso
médio)
S
pS=0.11
O equilíbrio
A
pA = 0.89
S
pS=0.11
AA
0.79
AS
0.20
SS
0.01
WAA = 0.9
WAS = 1
WSS
=0.2
No equilíbrio, existe variação genotípica no
valor adaptativo, mas não existe variação
genética aditiva (Excessos médios = 0).
Um equilíbrio por seleção natural irá
ocorrer quando Δp = 0 para todos os alelos.
aA = p(wAA-w) + (1-p)(wAa-w) = (1-p)(waa-w) + p(wAa- w) = aa
pwAA + (1-p)wAa = pwAa + (1-p)waa
(1-p) (wAa - wAA) = p(wAa -waa)
Se considerarmos WAa = 1, temos que
(1-peq) t = peqs
peq = t / (s+ t)
• Seleção tem dois efeitos:
– Promover mudança adaptativa
– Impedir alteração ao impedir alteração do status
quo
peq = t / (s+ t)
13
Duas respostas possíveis à Malária
Duas respostas possíveis à Malária
A
pA ≈1
A
pA ≈1
pS ≈ 0 pC ≈ 0
A
pA = 0.89
S
pS=.11
pS ≈ 0 pC ≈ 0
A
pA = 0.89
C
pC = 1
S
pS=.11
1. O genótipo mais apto é eliminado
1. O genótipo mais apto é fixado
2. Valor adaptat. médio vai de .9 a .91.
2. Valor adaptat. médio vai de .9 a 1.3.
3. 20% dos indivíduos tem viabilidade
relativa de 1 e 80% ou tem anemia ou
susceptibilidade à malaria.
3. 100% dos indivíduos tem viabilidade
relativa de 1.3 e nenhum tem anemia ou
susceptibilidade à malaria.
Duas respostas possíveis a Malária
Hb-A, S e C
Genótipos
A
pA ≈1
pS ≈ 0 pC ≈ 0
A
pA = 0.89
S
pS=.11
C
pC = 1
Com uma exceção
1. O genótipo mais apto é eliminado
1. O genótipo mais apto é fixado
2. Valor adaptat. médio vai de .9 a .91.
2. Valor adaptat. médio vai de .9 a 1.3.
3. 20% dos indivíduos tem viabilidade
relativa de 1 e 80% ou tem anemia ou
susceptibilidade à malaria.
3. 100% dos indivíduos tem viabilidade
relativa de 1.3 e nenhum tem anemia ou
susceptibilidade à malaria.
Hb-A, S e C
Genótipos AA AS
SS
AC
CS
CC
Viabilidade
sem Malária
0.2
1
0.7
1
1
1
C
pC = 1
C é um alelo neutro em
relação a A, logo,
algumas vezes o alelo
C terá sua freqüência
aumentada por deriva.
Viabilidade
sem Malaria
AA AS
1
1
SS
AC
CS
CC
0.2
1
0.7
1
S é um alelo recessivo,
deletério em relação a
A, portanto, seleção
natural irá mantê-lo
raro em um ambiente
pré-Malária.
Imagine que existisse um deme com este
pool gênico antes da agricultura começar
A
pA = 0.95
C
.045
pS=.005
14
Pool Gênico após várias gerações de
seleção em um ambiente com malária
Fenótipos iniciais após transição para a
agricultura Malásia
Genótipos
AA
AS
SS
AC
CS
CC
Viabilidade com Malaria
0.9
1
0.2
0.9
0.7
1.3
Desvios genotípicos
(W = 0.902)
.05
C
0.17
S
-.002 .098 -.702 -.002 -.202 .398
A resposta adaptativa inicial a
um ambiente com malária é o
aumento da freqüência de S e
de C.
aA = -0.001
aS = 0.081
aC = 0.015
A
pA = 0.78
Genótipos
AA
AS
SS
AC
CS
CC
Viabilidade com Malária
0.9
1
0.2
0.9
0.7
1.3
Desvios Genotípicos
(W = 0.914)
-.01
.09
-.71 -.01 -.21 .39
Pool Gênico após várias gerações de
seleção em um ambiente com malária
Após a resposta adaptativa
inicial à malária, a seleção
natural continua a reduzir
A, aumentar C, e agora
também irá reduzir S.
aA = -0.009
aS = -0.005
aC = 0.044
Existe uma correlação negativa entre as freqüências
de S e C em regiões com malária na África
0.25
Excesso médio de C em ambiente com
malária
0.20
o.15
C
Allele
Frequency
0.10
o.o5
o.o
o.o
o.o5
0.10
o.15
S Allele Frequency in 72 West African Populations
15
Adaptação por seleção natural
Excesso médio de C em ambiente com
malária
Adaptação por seleção natural depende da história:
Quais mutações estão presentes e quais suas freqüências.
Destas condições iniciais, a seleção natural modifica o
pool gênico a partir da perspectiva do gameta até que não
haja mais herdabilidade para o valor adaptativo (ou seja,
não exista mais variação genética aditiva).
Adaptação por seleção natural
Adaptação por seleção natural depende também do
presente:
Quais mutações estão presentes, quais suas freqüências e
como eles se encontram (sistema de acasalamento).
Como vocês esperam que este sistema se adapte à malária
caso não haja acasalamento ao acaso, e sim uma pequena
endogamia?
Como vocês esperam que este sistema se
adapte caso não haja acasalamento ao acaso,
e sim uma pequena endogamia?
• Como vimos, a existência de endogamia aumenta a
freqüência de homozigotos e diminui a de
heterozigotos na população.
• A vantagem do alelo S se expressa em heterozigose e a
desvantagem em homozigotos SS, logo, um aumento
da endogamia aumentará a seleção contra S
• Por outro lado, a vantagem do alelo C se expressa em
homozigose e a desvantagem em heterozigotos, logo,
um aumento da endogamia aumentará a seleção
favorável a C
Como vocês esperam que este sistema se
adapte caso não haja acasalamento ao acaso,
e sim uma pequena endogamia?
Genótipos AA AS
Anemia
SS
AC
CS
CC
Sim
Sim
Não Não
Não
NÃO
(Severa)
(Leve)
Resistência à
Não Sim
Malária
Sim
Não
Sim
Sim
Viabilidade
sem Malária
1
0.2
1
0.7
1
1
0.2
0.9
0.7
1.3
1
Viabilidade
0.9
com Malária
Adaptação como um processo poligênico
• Variação na região de Hbβ foi uma forma das
populações se adaptarem à malária, mas não foi a
única.
• Mais de 300 mutações independentes foram
identificadas para G6PD, que está envolvida em
estresse oxidativo celular. Tais deficiências limitam
capacidade de Plasmodium de sobreviver nas células.
• Mutações associadas à talassemia também foram
associadas à resistência a malária. Mais de 80
mutações induzem à α- talassemia e mais de 200 a β
- talassemia.
16
Adaptação por Seleção Natural
• O curso da adaptação é sempre restrito pela
disponibilidade de variação genética.
• Mesmo pressões de seleção uniformes podem criar
respostas adaptativas diferentes.
• Adaptação em geral envolvem vários loci, com
funções bioquímicas, celular e/ou no
desenvolvimento distintas.
17