MEDIDAS DE DIVERSIDADE BIOLÓGICA *
Ronald S. M. Barros
Programa de Pós-Graduação em Ecologia Aplicada ao Manejo e Conservação de Recursos Naturais –
PGECOL. Universidade Federal de Juiz de Fora – UFJF. Juiz de Fora, MG.
CONCEITOS
O termo biodiversidade tem sido amplamente empregado, tanto na literatura científica
quanto na mídia popular, mas nem sempre sua definição é clara, podendo ser usado com diferentes significados. De uma maneira mais simples o termo biodiversidade é usado como sinônimo de riqueza de espécies. Este, entretanto, refere-se apenas ao número de espécies presente numa determinada área definida. Algumas definições quantitativas de diversidade incluem
não apenas o aspecto variedade como também uma medida de abundância relativa, e têm sido
empregadas de maneira mais restrita ao considerarem apenas um determinado nível. No nível
das espécies pode-se falar, então, da diversidade de espécies, que inclui a variedade e a abundância relativa das espécies.
Como exemplo da diversidade de espécies considere uma amostra de 100 indivíduos retirados de uma comunidade, distribuídos em 10 espécies com 10 indivíduos cada, e outra amostra de uma segunda comunidade, também com 10 espécies, mas com uma das espécies
apresentando 55 indivíduos e todas as outras apenas cinco. Numa amostra ao acaso, as chances de retirar dois indivíduos que sejam de diferentes espécies é maior na primeira comunidade do que na segunda. Assim, embora as duas comunidades apresentem a mesma riqueza de
espécies (10), a primeira possui uma distribuição das abundâncias mais uniforme, sendo, portanto, mais diversa do que a segunda.
Uma definição precisa de biodiversidade, entretanto, deve considerar três níveis: a diversidade de espécies, a diversidade genética e a diversidade de ecossistemas. Assim, biodiversidade refere-se a todas as formas de vida, incluindo suas variações ao nível de genes, e
todos os ecossistemas num espaço definido como, por exemplo, a própria Terra. Uma vez que
grande parte dos estudos de biodiversidade está focada no nível da diversidade de espécies,
esta será enfatizada ao longo do texto.
*
Este texto foi elaborado como parte dos requisitos da disciplina Estágio Docência do Programa de PósGraduação em Ecologia/UFJF. 2007. Uso livre para propostas educativas.
DEFINIÇÃO DE DIVERSIDADE EM ESCALAS DISTINTAS
A diversidade dentro de um habitat não deve ser confundida com a diversidade de
uma região que contém vários habitats. Portanto, de acordo com a escala utilizada, pode-se
distinguir três tipos de diversidade: alfa (α), beta (β) e gama (γ). A diversidade α, ou local,
corresponde à diversidade dentro de um habitat ou comunidade, e é bastante sensível à definição de habitat, e à área e intensidade da amostragem. A diversidade γ, ou regional, corresponde à diversidade de uma grande área, bioma, continente, ilha, etc. A diversidade β corresponde à diversidade entre habitats ou outra variação ambiental qualquer, isto é, mede o quanto
a composição de espécies varia de um lugar para outro.
O QUE DEVEMOS MEDIR
É bastante inviável, ou mesmo impossível, amostrar todas as espécies de um determinado habitat. Assim, as medidas de diversidade se restringem a determinados grupos como,
por exemplo, a diversidade de aves ou de espécies arbóreas de um determinado habitat. Entretanto, alguns problemas de ordem prática podem surgir quando medindo a diversidade de espécies. Para alguns grupos taxonômicos não é possível identificar os indivíduos ao nível de
espécie. Neste caso, é possível medir a diversidade considerando outros níveis de classificação como gênero, família, ordem ou mesmo tipos morfológicos. Outro problema surge quando o pesquisador está interessado em espécies que possuem fases de vida com características
distintas. Por exemplo, fases de larva e adulto, que vivem em diferentes ambientes, exercendo
papéis distintos no ecossistema, devem ser consideradas como um mesmo grupo? Uma solução proposta, neste caso, é medir a diversidade considerando-se tipos ecológicos (guildas)
mais do que espécies. Ainda, o uso do número de indivíduos pode não ser uma medida adequada. Por exemplo, ao medir a diversidade de carnívoros num determinado ambiente pode
ser mais útil considerar uma medida de biomassa de cada espécie ao invés do número de indivíduos. De fato, o que a medida de diversidade requer é uma estimativa da importância da
espécie na comunidade, e esta pode ser números, biomassa ou até medidas de produtividade.
Os objetivos do estudo irão determinar a melhor medida.
AMOSTRAGEM
A riqueza e a diversidade de espécies dependem, além da própria natureza da comunidade, do esforço amostral despendido, uma vez que o número de espécies aumenta com o
2
aumento do número de indivíduos amostrados. As curvas de acumulação de espécies (curvas
do coletor) permitem avaliar o quanto um estudo se aproxima de capturar todas as espécies do
local. Quando a curva estabiliza, ou seja, nenhuma espécie nova é adicionada, significa que a
riqueza total foi obtida. A partir disso, novas amostragens não são necessárias. Um exemplo
Número acumulado de espécies
de curva do coletor é apresentado na figura 1.
60
50
40
30
20
10
0
0
5
10
15
20
25
30
Número de indivíduos
Figura 1. Curva de acumulação de espécies hipotética. O número de espécies
aumenta com o aumento do esforço amostral. Após 30 indivíduos amostrados a
curva não atingiu a assíntota.
Obtendo-se várias curvas a partir da adição aleatória das amostras pode-se calcular
uma curva do coletor média. Em todo caso, a estabilização da curva é bastante difícil, pois
muitas espécies raras costumam ser adicionadas após muitas amostragens, sobretudo em regiões tropicais. Assim, medidas de riqueza de espécies que permitam estimar a riqueza a partir
dos dados obtidos, ou comparar inventários entre diferentes áreas com diferentes unidades
amostrais são bastante úteis nestes casos.
MEDIDAS DE RIQUEZA DE ESPÉCIES
Em comparações de comunidades, o número de espécies por número de indivíduos amostrados é uma medida bastante útil. Entretanto, freqüentemente a comparação de comunidades é baseada em diferentes tamanhos amostrais, que, por sua vez, irão dificultar conclusões. Como comparar, por exemplo, uma comunidade em que foram observadas nove espécies numa amostra de 23 indivíduos com uma segunda onde foram observadas seis espécies
numa amostra de 13 indivíduos? Para lidar com este problema uma solução proposta é a técnica de Rarefação, que consiste em calcular o número esperado de espécies em cada amostra
para um tamanho de amostra padrão. O número esperado de espécies é obtido pela equação:
3
⎡ ⎛ N − Ni ⎞ ⎤
⎟⎟ ⎥
⎢ ⎜⎜
n
⎝
⎠⎥
E ( S ) = ∑ ⎢1 −
⎢
⎛N⎞ ⎥
i =1
⎜⎜ ⎟⎟ ⎥
⎢
⎝n⎠ ⎦
⎣
S
onde E(S) é o número esperado de espécies em uma amostragem aleatória, S é o número total
de espécies registradas, N é o número total de indivíduos registrados, Ni é o número de indivíduos da espécie i, e n é o tamanho padronizado da amostra escolhido.
⎛N⎞
N!
O termo ⎜⎜ ⎟⎟ é calculado como
n!( N − n)!
⎝n⎠
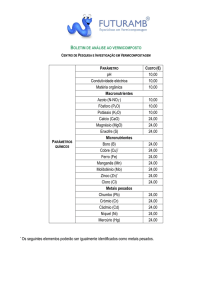
Um exemplo de rarefação é apresentado abaixo para amostras de duas comunidades hipotéticas com 23 e 13 indivíduos cada, e nove e seis espécies, respectivamente (Tab. 1; Fig.
2). É possível determinar qual a comunidade mais rica, comparando o número de espécies
esperado para 13 indivíduos em cada amostra. Neste caso E(S) é igual a 6,56 para a amostra A
e 6 para a amostra B.
A rarefação deve ser usada apenas para amostras obtidas com métodos padronizados, e
em habitats iguais ou similares. Outra restrição é que as curvas não podem ser extrapoladas
para além do número de indivíduos (N) na maior amostra. O método de rarefação permite
ainda o cálculo da variância do número esperado de espécies. A fórmula para calcular a variância é descrita em Krebs (1999).
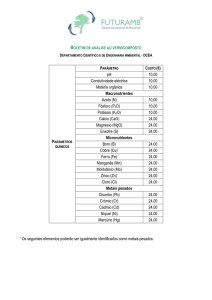
Tabela 1. Amostras de espécies de
duas comunidades hipotéticas A e
B.
Comunidade
Espécies
A
B
Espécie 1
9
1
Espécie 2
3
0
Espécie 3
0
1
Espécie 4
4
0
Espécie 5
2
0
Espécie 6
1
0
Espécie 7
1
1
Espécie 8
0
2
Espécie 9
1
0
Espécie 10
0
5
Espécie 11
1
3
Espécie 12
1
0
Total
23
13
4
Número esperado de espécies
10
9
8
7
6
5
4
3
2
1
0
0
5
10
15
20
Número de indivíduos na amostra
Figura 2. Curva de rarefação para duas comunidades. A amostra da comunidade A
(■) tem 23 indivíduos representando nove espécies, enquanto a amostra da comunidade B (▲) tem apenas 13 indivíduos representando seis espécies. O número de
espécies esperado na comunidade A para uma amostra de 13 indivíduos é 6,56.
Além da rarefação, alguns índices simples são utilizados como medidas de riqueza de
espécies, incluindo o índice de Margalef (DMg) e o de Menhinick (DMn). Estes índices combinam o número de espécies registrado (S) com o número total de indivíduos (N) e são calculados pelas seguintes equações:
DMg = ( S − 1) / ln N
DMn = S / N
Para o exemplo da tabela 1 os resultados são DMg = 2,55 e DMn = 1,88 para a amostra
A, e DMg = 1,95 e DMn = 1,66 para a amostra B.
Outras medidas de riqueza de espécies, que permitem comparar dados obtidos com métodos e esforço de coleta diferentes, são as estimativas de riqueza. Tais medidas possibilitam
estimar o número total de espécies numa determinada comunidade a partir dos dados amostrais. Entre estas estão os estimadores Jackknife (1 e 2), Chao (1 e 2) e Bootstrap. A vantagem
desses estimadores é a disponibilidade de equações para o cálculo de limites de confiança da
estimativa.
O método Jackknife 1 estima a riqueza total utilizando o número de espécies que ocorrem em apenas uma amostra (uniques). A estimativa de riqueza é calculada pela equação:
S j = s + Q1
n −1
n
onde Sj é a riqueza estimada, s é a riqueza observada, Qj é o número de espécies que ocorrem
em exatamente j amostras, e n é o número de amostras. Já o Jackknife 2 utiliza os uniques e o
5
número de indivíduos que ocorrem em duas amostras (duplicates), e é calculado pela equação:
Q (2n − 3) Q2 (n − 2)
Sj = s+ 1
−
n
n(n − 1)
2
O método Chao 1 estima a riqueza total utilizando o número de espécies representadas
por apenas um indivíduo nas amostras (singletons), e o número de espécies com apenas dois
indivíduos nas amostras (doubletons). A estimativa de riqueza é calculada pela equação:
Sc = s +
F12
2F2
onde Sc é a riqueza estimada, s é a riqueza observada, Fi é o número de espécies que têm exatamente i indivíduos em todas as amostras. O Chao 2 é uma adaptação que utiliza os uniques e
os duplicates, e pode ser calculada substituindo Fi por Qj, o qual corresponde ao número de
espécies que ocorrem em exatamente j amostras.
O método Bootstrap estima a riqueza total utilizando dados de todas as espécies, não
se restringindo às espécies raras. Este método, entretanto, requer o uso de simulação em computador, onde uma amostra aleatória de tamanho n é retirada dos dados obtidos, usando amostragem com reposição. A estimativa de riqueza é então calculada pela equação:
S b = s + ∑ (1 − pi )
n
onde Sb é a riqueza estimada, s é a riqueza observada, e pi é a proporção das amostras n que
contêm a espécie i. Estes passos devem ser realizados de 100 a 500 vezes no computador.
MODELOS DE DISTRIBUIÇÃO DE ABUNDÂNCIA
Nenhuma comunidade real apresenta de fato uma diversidade máxima onde todas os
grupos definidos (espécies, guildas) têm importâncias iguais. Em geral, na natureza, para o
número total de grupos, a maior parte dos seus constituintes é rara (poucos indivíduos, pequena biomassa, baixa produtividade, ou outra medida de importância) enquanto poucos são dominantes ou comuns (altos valores de importância). Tal observação levou ao desenvolvimento
de modelos de abundância de espécies, também chamados diagramas de ranks de abundância. Estes modelos utilizam o conjunto total dos valores de importância de cada espécie, plotando tais valores contra um rank de abundância das espécies, ou seja, as espécies são plotadas das mais comuns para as mais raras. A linha que se ajusta aos pontos é o modelo que explica a diversidade. Quatro modelos principais são propostos: a série geométrica, a série loga-
6
rítmica, a distribuição logaritmo-normal, e o modelo broken stick (“pau quebrado”) de MacArthur (Fig. 3).
Figura 3. Curvas hipotéticas para quatro modelos de
ranks de abundância: série geométrica, série logarítmica,
distribuição logaritmo-normal, e broken stick. Adaptado
de Magurran (1988).
Se a espécie mais abundante for duas vezes mais numerosa que a espécie seguinte, a
qual tem duas vezes a abundância da terceira, e assim por diante, o modelo ajustado será o da
série geométrica. Este modelo prediz a distribuição menos uniforme de todos e aparece como
uma reta. A série geométrica ocorreria numa situação em que as espécies chegariam num habitat não saturado em intervalos de tempo regulares, e ocupariam frações do nicho remanescente. Pode-se supor que a primeira espécie ocupa uma fração k do nicho disponível, a segunda espécie ocupa a mesma fração k do nicho remanescente, e assim por diante. Dessa forma,
cada espécie ocupa uma parte do espaço de nicho, sem sobreposição. Assim a série geométrica é chamada de hipótese de apropriação de nicho (niche pre-emption hypothesis). A série
geométrica é encontrada em algumas comunidades vegetais, sobretudo em ambientes extremos, pobres em espécies, ou em estágios iniciais de sucessão.
No extremo oposto, o espaço de nicho é aleatoriamente dividido em segmentos, e as
distribuições das abundâncias são mais uniformes que nos outros modelos. Neste caso, o modelo do “pau quebrado” apresenta o melhor ajuste. De acordo com o modelo, o nicho se divide como um pau que se fragmenta simultaneamente, e de maneira aleatória, em pedaços de
vários tamanhos. Este modelo não é freqüentemente encontrado na natureza, mas pode ser
encontrado, por exemplo, em comunidades de aves que apresentam comportamentos territoriais.
7
Os modelos intermediários (série logarítmica e distribuição logaritmo-normal) indicam padrões mais complexos de sobreposição de nicho e uniformidades intermediárias. No
caso da série logarítmica, a distribuição seria resultante de espécies chegando num habitat a
intervalos de tempo irregulares, ou seja, aleatórios, antes que a espécie anterior ocupe totalmente a fração de nicho potencial. Neste caso, a maioria das espécies é rara. Já a distribuição
logaritmo-normal deriva de espécies coexistindo em condições de competição parcial, em vez
de direta, com adaptações promovendo uma diferenciação de nicho sem uma exclusão competitiva de um habitat. Neste caso, a maioria das espécies possui abundâncias intermediárias
(nem raras, nem comuns). Muitos conjuntos de dados serão descritos igualmente bem por
ambos os modelos, podendo ser bastante difícil decidir qual é o mais apropriado.
Embora o método de plotar os dados num gráfico possa dar uma idéia do modelo apropriado, é necessário testar os ajustes matematicamente. Os cálculos para cada um dos modelos, bem como críticas sobre a utilização de cada um deles, podem ser encontrados em Magurran (1988) e em Martí & García-Álvarez (2002). Krebs (1999) também apresenta os cálculos para a série logarítmica e a distribuição logaritmo-normal.
MEDIDAS DE DIVERSIDADE α
Embora os modelos de abundância de espécies descrevam a diversidade de maneira
mais completa, os índices baseados nas abundâncias proporcionais das espécies são as medidas de diversidade mais utilizadas em ecologia. Estes índices levam em conta, tanto a uniformidade (eqüitabilidade) quanto a riqueza de espécies, sendo chamados também de índices de
heterogeneidade. O aumento do número de espécies ou o aumento da uniformidade das abundâncias aumenta a diversidade.
O índice mais utilizado é o índice de Shannon-Wiener (H’), procedente da teoria da
informação. Este índice dá maior peso para as espécies raras, e é obtido pela equação:
S
H ' = −∑ pi ln pi
i =1
onde S é o número de espécies, pi é a proporção da espécie i, estimada como ni/N, onde ni é a
medida de importância da espécie i (número de indivíduos, biomassa), e N é o número total
de indivíduos. Na prática o valor máximo de H’ é ln S, e o mínimo é ln [N/(N – S)].
Para o exemplo da tabela 1, H’ é 1,831 para a amostra A e 1,586 para a amostra B.
Embora o índice de Shannon-Wiener leve em conta a uniformidade das abundâncias de espécies, é possível calcular esta medida separadamente. A uniformidade (E; eqüitabilidade de
Pielou) pode ser calculada como a razão entre a diversidade obtida e a diversidade máxima
8
(H’max), a qual seria possível em uma situação onde todas as espécies fossem igualmente abundantes. Neste caso H’max = ln S. O valor de E varia entre 0 e 1 com 1 representando uma
situação em que todas as espécies são igualmente abundantes. Para o exemplo anterior, os
valores de E são 0,833 e 0,885 para as amostras A e B, respectivamente.
Um índice semelhante ao índice de Shannon-Wiener é o índice de Brillouin:
HB = (ln N! – ∑ln ni!)/N
Embora diversos autores sugiram que este índice é mais satisfatório do que o de
Shannon-Wiener, a maioria dos pesquisadores continua utilizando o primeiro pela sua simplicidade de cálculo.
Outros índices utilizados pertencem ao grupo das medidas de dominância, uma vez
que dão mais peso para as espécies comuns. O mais utilizado é o índice de Simpson (D). De
acordo com este índice, a probabilidade de dois indivíduos retirados aleatoriamente de uma
comunidade pertencerem à mesma espécie é dada por:
S
D = ∑ pi2
i =1
onde pi é a proporção da espécie i na comunidade, e S é o número de espécies. Para uma população finita, a forma apropriada de calcular essa probabilidade é dada por:
S
⎡ n (n − 1) ⎤
D = ∑⎢ i i
⎥
i =1 ⎣ N ( N − 1) ⎦
onde ni é o número de indivíduos na espécie i, e N é o número total de indivíduos.
Uma vez que quando D aumenta a diversidade diminui, para converter essa probabilidade a uma medida de diversidade, o índice é expresso como 1/D. No exemplo anterior
(Tab. 1) as estimativas D e 1/D são, respectivamente, 0,182 e 5,5 para a amostra A, e 0,179 e
5,571 para a amostra B.
Outras medidas de dominância, incluindo a medida de diversidade de McIntosh e o
índice de Berger-Parker, são descritas em Magurran (1988). Krebs (1999) descreve outras
medidas de uniformidade, incluindo as medidas de Simpson, de Camargo, de Nee e de Smith
e Wilson.
Um método para calcular a estatística t é descrito por Magurran (1988) para testar diferenças significativas entre índices de Shannon-Wiener obtidos para diferentes amostras. De
fato, em todos os casos os índices deveriam ser testados estatisticamente para prover uma informação mais refinada sobre suas diferenças. Obtendo-se medidas de diversidade para diver-
9
sas amostras em cada comunidade, pode-se utilizar métodos paramétricos ou nãoparamétricos para testar as diferenças.
MEDIDAS DE DIVERSIDADE β
As medidas descritas anteriormente permitem comparar diferentes comunidades em
termos de diversidade ou riqueza, mas não descrevem o quanto as comunidades são distintas
(ou similares) em termos de composição de espécies. Para lidar com essa questão, deve-se
utilizar medidas de diversidade β. A diversidade β, ou diversidade diferencial, é uma medida
de como a variedade (e em alguns casos as abundâncias) de espécies difere entre comunidades
ou amostras ao longo de gradientes. Quanto menos espécies as diferentes comunidades compartilham, mais alta é a diversidade β.
A maneira mais simples de medir a diversidade β entre pares de locais é pelo uso dos
coeficientes de similaridade. Estes coeficientes comparam comunidades de forma qualitativa
ou quantitativa. Os índices qualitativos, ou coeficientes binários, mais utilizados são o índice
de Jaccard (Sj) e o índice de Sorensen (Ss):
Sj = a/(a + b + c)
Ss = 2a/(2a + b + c)
onde a é o número de espécies encontrados em ambos os locais, A e B, b é o número de espécies no local B, mas não em A, c é o número de espécies no local A, mas não em B. A
desvantagem destes índices é que eles não levam em conta as abundâncias das espécies. Um
dos índices propostos que considera as abundâncias das espécies é o índice de Morisita-Horn
(CH):
CH =
[(∑ X
2∑ X ij X ik
2
ij
) (∑ X
/N +
2
j
2
ik
)]
/ N k2 N j N k
onde Xij e Xik são, respectivamente, o número de indivíduos da espécie i nas amostras j e k, e
Nj e Nk são, respectivamente, o número total de indivíduos nas amostras j e k.
O valor dos coeficientes de similaridade varia de 0 (nenhuma similaridade) a 1 (total
similaridade). Para o exemplo da tabela 1 os resultados de cada índice são: Sj = 0,25; Ss = 0,4;
CH = 0,189. Comparando o índice de Morisita-Horn (CH) com os dois primeiros, percebe-se
que a similaridade diminuiu bastante quando as abundâncias das espécies foram consideradas.
Os coeficientes de similaridade são medidas bastante úteis quando comparando três
comunidades ou mais, permitindo verificar quais são as mais similares (ou dissimilares). Os
índices são calculados para cada par de comunidades e os resultados comparados.
10
Além dos coeficientes de similaridade, várias medidas de diversidade β são propostas
para avaliar o quanto a composição de espécies muda ao longo de um gradiente ou transecto,
entre as quais a medida de Whittaker (βW), obtida pela equação:
βW = (S / α ) − 1
onde S é o número total de espécies registrado no sistema, e α é a riqueza de espécies média
das amostras. Outras cinco medidas de diversidade β, incluindo as medidas de Cody, de Routledge e de Wilson e Shmida, além da medida de Whittaker, são descritas em Magurran
(1988), assim como uma avaliação destas.
Quando há várias comunidades ou gradientes sendo comparados simultaneamente
uma boa representação da diversidade β pode ser obtida através das análises de agrupamento
(cluster analysis). A análise de agrupamento é obtida através de uma matriz contendo a similaridade (ou dissimilaridade) para cada par de amostras. As duas amostras mais similares são
combinadas para formar um grupo único. As análises procedem até o último grupo ser combinado, originando um dendograma. Um exemplo é dado abaixo para cinco amostras, comparadas pelo coeficiente de similaridade de Jaccard. Os dados são apresentados na tabela 2; uma
matriz de similaridade é apresentada na tabela 3; a figura 4 apresenta o dendograma resultante. A similaridade foi maior entre as amostras 3 e 5, que formaram um primeiro grupo no dendograma.
Tabela 2. Abundâncias de 12 espécies em cinco amostras hipotéticas.
Amostras
Espécies
1
2
3
4
5
Espécie 1
9
1
0
2
0
Espécie 2
3
0
4
0
6
Espécie 3
0
1
2
1
0
Espécie 4
4
0
1
9
6
Espécie 5
2
0
0
5
0
Espécie 6
1
0
0
1
0
Espécie 7
1
1
1
0
2
Espécie 8
0
2
0
1
2
Espécie 9
1
0
3
0
2
Espécie 10
0
5
0
3
0
Espécie 11
1
3
5
0
3
Espécie 12
1
0
0
0
2
Tabela 3. Matriz obtida pelo índice de Jaccard (%).
Amostra 1 Amostra 2 Amostra 3 Amostra 4 Amostra 5
Amostra 1
*
25
50
33,3333
60
Amostra 2
*
*
33,3333
44,4444
30
Amostra 3
*
*
*
18,1818
62,5
Amostra 4
*
*
*
*
16,6667
Amostra 5
*
*
*
*
*
11
Figura 4. Dendograma de similaridade para cinco amostras hipotéticas. A análise de agrupamento
foi obtida pelo coeficiente de similaridade de Jaccard, método de ligação simples.
O VALOR DAS MEDIDAS DE DIVERSIDADE
Ecólogos estão interessados em medir a diversidade por diversas razões, sobretudo por
sua utilidade em biologia da conservação e avaliação ambiental. Medidas de diversidade de
espécies são geralmente úteis para comparar padrões em diferentes locais ou em diferentes
gradientes, ou, ainda, numa mesma área ao longo do tempo, como, por exemplo, ao longo de
uma sucessão, ou após um distúrbio. Além disso, a avaliação de espécies raras é útil para direcionar esforços de conservação.
Ainda, o interesse de ecólogos em explicar porque algumas áreas são mais ricas em espécies do que outras, ou porque uma espécie é abundante em um local, mas rara em outro,
tem estimulado pesquisas sobre diversidade de habitats e largura de nicho. Nestes casos as
mesmas medidas para avaliação da diversidade de espécies podem ser utilizadas.
12
REFERÊNCIAS BIBLIOGRÁFICAS
BEGON, M.; TOWNSEND, C.R. & HARPER, J.L 2005. Ecology: from individuals to ecosystems. 4. ed. Malden: Blackwell Publishing, 2006. 738 p.
MARTÍ, J.J.I. & GARCÍA-ÁLVAREZ, A. 2002. Diversidad: biodiversidad edáfica e
geodiversidad. Edafología 9 (3): 329-385.
KREBS, C.J. 1999. Ecological Methodology. 2. ed. New York: Benjamin/ Cummings, 620
p.
MAGURRAN, A.E. 1988. Ecological diversity and its measurement. New Jersey: Princeton University Press, 179 p.
ODUM, E.P. 1988. Ecologia. Rio de Janeiro: Guanabara Koogan, 434 p.
RICKLEFS, R.E. 1996. A economia da natureza. 3. ed. Rio de Janeiro: Guanabara Koogan.
470 p.
SANTOS, A.J. 2003. Estimativas de riqueza em espécies. In: L. Cullen Jr., C. ValladaresPádua, Rudy Rudran (orgs.). Métodos de estudos em biologia da conservação e manejo da
vida silvestre. Curitiba: UFPR/ Fundação O Boticário de Proteção à Natureza, 665 p. p.1941.
PROGRAMAS ÚTEIS
EstimateS - http://viceroy.eeb.uconn.edu/EstimateS
Biodiversity Pro - http://www.sams.ac.uk/research/software
Past - http://folk.uio.no/ohammer/past/
13