Anatomia, Histologia e Fisiologia Humanas I
Apontamentos de Apoio ao Estudo
O presente texto constitui apenas um auxiliar de estudo, não devendo substituir a consulta da
bibliografia recomendada e das matérias leccionadas nas aulas.
Atualizado em Setembro de 2015
Histologia Básica
- alguns conceitos -
Tecidos e histologia
Um tecido é constituído por um grupo de células com função e estrutura semelhantes, inseridas numa
matriz extra-celular com características sobreponíveis.
O estudo microscópico da estrutura dos tecidos é designada por histologia.
Os tecidos organizam-se para formar órgãos, sistemas orgânicos e estes encontram-se dispostos e
interrelacionados no organismo.
Os quatro tipos fundamentais de tecidos são: tecido epitelial, tecido conjuntivo, tecido muscular e
tecido nervoso.
Histologia Básica - Serviço de Patologia Experimental . FMUC!
1/15
Anatomia, Histologia e Fisiologia Humanas I
Preparação de tecidos para microscopia óptica
1. Colheita
2. Fixação. Os mecanismos envolvidos na fixação compreendem: oxidação, desnaturação das
proteínas e formação de novas ligações químicas com as moléculas do fixador, que o tornam inerte.
O fixador mais utilizado é o formaldeído. Este agente químico reage com as proteínas e forma pontes
entre grupos funcionais próximos, inativando-as. Ao mesmo tempo endurece os tecidos, facilitando o
seu posterior corte.
3. Desidratação com banhos de álcoois progressivamente mais concentrados
4. Diafanização com xilol
5. Impregnação com parafina
6. Inclusão em parafina
7. Microtomia
8. Estendimento
9. Coloração. Um corante é uma substância capaz de se ligar a constituintes do tecido e conferir-lhe
cor fixa.
10. Montagem da lamela
Histologia Básica - Serviço de Patologia Experimental . FMUC!
2/15
Anatomia, Histologia e Fisiologia Humanas I
Tecidos epiteliais
O tecido epitelial encontra-se no corpo humano disposto de forma a revestir as superfícies internas e
externas. Também as glândulas são constituídas por células epiteliais especializadas. Os epitélios
são constituídos por células, com pouca matriz extracelular.
Genericamente os epitélios são constituídos por uma superfície livre, constituída por uma camada de
células epiteliais com uma das faces sem relação com nenhuma célula e, uma membrana basal que
constitui a interface do tecido epitelial com os tecidos subjacentes. O tecido epitelial não possui vasos
sanguíneos, pelo que os gases e nutrientes difundem através da membrana basal para chegar a
todas as células que constituem o epitélio. Também os produtos do metabolismo excretados pelas
células epiteliais sofrem o processo de difusão através da membrana basal, atingindo os vasos
sanguíneos existentes no tecido conjuntivo subjacente.
Funções dos epitélios:
1. Protecção das estruturas adjacentes
2. Função de barreira física e química
3. Controlo da passagem e absorção de substâncias
4. Excreção de substâncias
Epitélio pavimentoso simples
Epitélio constituído por uma fina camada de células com forma poligonal e núcleo central.
Encontra-se nos alvéolos pulmonares, ansa de Henle, camada parietal da cápsula de Bowman,
endotélio dos vasos sanguíneos e linfáticos e mesotélio das cavidades pleural e peritoneal.
Epitélio cúbico simples
As células apresentam secção quadrada com núcleo redondo central.
Encontra-se nos ductos das glândulas corporais e como revestimento do ovário e alguns túbulos
renais.
Histologia Básica - Serviço de Patologia Experimental . FMUC!
3/15
Anatomia, Histologia e Fisiologia Humanas I
Epitélio colunar simples
Constituído por uma camada de células que vistas superficialmente são cúbicas, mas que em secção
longitudinal são rectangulares com núcleo ovóide central ou polarizado.
Encontra-se como revestimento do tubo digestivo, bexiga e grandes ductos glandulares, útero,
oviducto, ducto eferente e pequenos brônquios.
Epitélio pavimentoso estratificado não queratinizado
Tecido constituído por várias camadas celulares. Na zona mais basal as células apresentam forma
cúbica e as mais superficiais são achatadas.
Encontra-se como revestimento da boca, faringe e esófago.
Epitélio pavimentoso estratificado queratinizado
Tecido semelhante ao anterior, em que as células superficiais se encontram queratinizadas. Encontrase na epiderme cutânea.
Epitélio cúbico estratificado
Constituído por mais de duas camadas de células cúbicas, como o epitélio dos ductos das glândulas
sudoríparas.
Epitélio colunar estratificado
As camadas mais profundas são constituídas por células cúbicas, apresentando as células das
camadas superficiais forma colunar. Encontra-se em algumas regiões da uretra e em alguns ductos
excretores.
Histologia Básica - Serviço de Patologia Experimental . FMUC!
4/15
Anatomia, Histologia e Fisiologia Humanas I
Epitélio de transição
As células basais apresentam forma colunar e as células superficiais possuem forma variável de
acordo com o órgão e a sua distensão, podendo adoptar uma forma cúbica ou achatada. Encontra-se
no sistema urinário, nomeadamente na bexiga.
Epitélio pseudoestratificado
Epitélio constituído por uma só camada de células, com núcleos dispostos em níveis diferentes, que
lhe confere o aspecto aparente de epitélio estratificado.
Encontra-se no epididimo, traqueia, brônquios principais, ou saco lacrimal.
Histologia Básica - Serviço de Patologia Experimental . FMUC!
5/15
Anatomia, Histologia e Fisiologia Humanas I
Glândulas
As glândulas são formadas por células epiteliais que invadem o tecido conjuntivo. São constituídas
por parênquima que constitui o ducto e a unidade secretora, disposto num estroma de tecido
conjuntivo.
Quanto à forma de distribuição dos produtos secretados, as glândulas podem ser exócrinas (secreta
os produtos directamente para ductos, localmente); endócrinas (liberta os seus produtos para os
vasos sanguíneos ou linfáticos) ou mistas (alguns produtos de secreção são lançados no sangue e
outros conduzidos através de canais).
As glândulas exócrinas podem ser classificadas quanto ao número de células como unicelulares ou
pluricelulares. As glândulas pluricelulares apresentam uma porção secretora e um ducto. De acordo
com a estrutura dos ductos podem ser simples ou compostas. De acordo com a estrutura da porção
secretora podem ser tubulares, acinosas ou tubuloacinosas.
Nas glândulas exócrinas autócrinas a célula alvo é a célula sinal. Nas glândulas exócrinas parácrinas
a célula alvo é uma célula vizinha da célula sinal.
Quando à forma como os produtos de secreção são libertados pelas glândulas exócrinas, podemos
classificá-las em: glândulas merócrinas que libertam grânulos citoplasmáticos, sem que haja
interferência com a integridade da célula; apócrinas, cuja secreção é libertada conjuntamente com
porções do citoplasma da célula, para o lúmen glandular; holócrinas, cuja secreção leva à
desintegração da célula.
Quanto ao tipo de secreção, as glândulas exócrinas podem ser serosas (as células apresentam
aspecto granuloso que segregam fluido rico em enzimas. Possuem citoplasma acidófilo) ou mucosas
(o seu produto de secreção é viscoso, com função de lubrificação e protecção. O núcleo das células
glandulares é achatado e comprimido contra a base das células).
Histologia Básica - Serviço de Patologia Experimental . FMUC!
6/15
Apontamentos de Apoio ao Estudo
O presente texto constitui apenas um auxiliar de estudo, não devendo substituir a consulta da
bibliografia recomendada e das matérias leccionadas nas aulas.
Tecidos conjuntivos
Os tecidos conjuntivos constituem um meio de trocas de nutrientes e de produtos do metabolismo.
Contribuem para a defesa e protecção do organismo e constituem uma fonte extracelular de
nutrientes.
Alguns tecidos conjuntivos possuem características gerais, enquanto outros são especializados.
Os tecidos conjuntivos são constituídos por uma abundante matriz extracelular rica em fibras, onde se
encontram dispersas células com funções específicas.
A matriz extracelular é rica em glicosaminaglicanos, proteoglicanos, glicoproteínas de adesividade
(laminina, condronectina, osteonectina e fibronectina), fibras de colagénio e fibras elásticas.
As células dos tecidos conjuntivos derivam de uma célula mesenquimatosa indiferenciada.
As células fixas do tecido conjuntivo são: fibroblastos (células alongadas e fusiformes com função de
síntese de fibras), adipócitos (células com capacidade de armazenamento de gorduras neutras),
macrófagos (células com capacidade fagocitária), mastócitos (células esféricas a ovóides com
grânulos finos basófilos, com função de síntese de heparina e histamina).
Tecido conjuntivo laxo
Tecido constituído por abundante matriz extracelular, fibroblastos e adipócitos dispersos. Constitui o
suporte do tubo digestivo e do tecido epitelial respiratório.
Tecido conjuntivo denso
Possui fibras de colagénio mais espessas e abundantes, com menos células e menos matriz
extracelular. Pode ser irregular, se as fibras não se apresentam orientadas, ou regular, se as fibras se
encontram dispostas paralelamente.
Tecido conjuntivo reticular
Constitui suporte de órgãos parenquimatosos e possui matriz com fibras reticulares (fibras de
colagénio delgadas e finas dispostas em rede).
Histologia Básica - Serviço de Patologia Experimental . FMUC!
7/15
Apontamentos de Apoio ao Estudo
O presente texto constitui apenas um auxiliar de estudo, não devendo substituir a consulta da
bibliografia recomendada e das matérias leccionadas nas aulas.
Tecido cartilagíneo
As principais funções deste tecido são a absorção dos choques e a redução do atrito das superfícies
articulares.
Possui células especializadas: os condrócitos, localizados em lacunas.
Cartilagem hialina
Possui pericôndrio de revestimento que permite a sua regeneração. A matriz interterritorial encontrase adjacente ao pericôndrio e nela podemos observar lacunas de menores dimensões. Estas
aumentam de dimensão à medida que nos afastamos do pericôndrio, constituindo a matriz territorial,
onde os condrócitos se encontram mais maduros.
Rica sobretudo em colagénio tipo II.
Cartilagem elástica
Semelhante à anterior, apresenta matriz extracelular com fibras elásticas.
A matriz tem um aspecto mais uniforme, onde é notória a estriação.
Fibrocartilagem
Pobre em condrócitos, que se dispoem paralelamente entre as numerosas fibras de colagénio tipo I.
Não possui pericôndrio.
Condroblastos
Células de forma oval, basófilas com organitos de síntese proteica.
Condrócitos
Células de forma ovóide ou esferóide, com núcleo grande e nucléolo evidente.
Histologia Básica - Serviço de Patologia Experimental . FMUC!
8/15
Apontamentos de Apoio ao Estudo
O presente texto constitui apenas um auxiliar de estudo, não devendo substituir a consulta da
bibliografia recomendada e das matérias leccionadas nas aulas.
Tecido ósseo
É um tecido conjuntivo especializado cuja matriz extracelular está calficicada. Tem como função o
suporte físico e protecção.
A matriz é constituída por uma componente orgânica (colagénio tipo I, glicosaminaglicanos, agrecano,
osteocalcina, osteopontina e osteonectina) e uma componente inorgânica (hidroxiapatite).
Osteoblastos
Derivam de células percursoras (células fusiformes, com núcleo ovóide e claro) do mesênquima
embrionário que se encontram no periósteo (porção externa do osso) e no endósteo (porção interna
do osso).
Sintetizam componentes orgânicos da matriz óssea: o osteóide. São células cúbicas ou cilíndricas de
citoplasma basófilo.
Osteócitos
São células pequenas, encerradas em lacunas, com núcleo achatado.
Osteoclastos
São agregados gigantes multicelulares com citoplasma acidófilo, que possuem numerosas vesículas,
vacúolos e lisossomas, responsáveis pela reabsorção da substância óssea. Derivam de células
percursoras específicas presentes na medula óssea.
Não se encontram encerrados em lacunas, antes, dispoem-se na superfície do osso, onde formam
pequenas depressões (lacunas de Howship).
Histologia Básica - Serviço de Patologia Experimental . FMUC!
9/15
Apontamentos de Apoio ao Estudo
O presente texto constitui apenas um auxiliar de estudo, não devendo substituir a consulta da
bibliografia recomendada e das matérias leccionadas nas aulas.
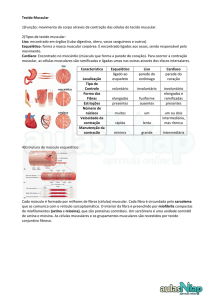
Tecido Muscular
O tecido muscular divide-se em dois tipos histológicos fundamentais, o tecido muscular liso e o tecido
muscular estriado. O tecido muscular estriado apresenta ainda duas variedades histológicas, o tecido
muscular cardíaco e o tecido muscular esquelético.
Músculo cardíaco
No corte longitudinal, as células musculares cardíacas apresentam um ou dois núcleos e um
citoplasma extenso com ramificações que lhe conferem a aparência de uma rede tridimensional
contínua.
O tecido muscular cardíaco encontra-se apenas no coração e tem capacidade de contracção rítmica.
As células que o constituem possuem um só núcleo central, cada uma com cerca de µm de
comprimento e 15µm de diâmetro. As células encontram-se em contacto umas com as outras, através
dos vários ramos em que cada uma delas se divide e anastomosa com as vizinhas, formando os
sincicios. O endomísio é altamente vascularizado e inervado. Entre as extremidades das células
musculares cardíacas adjacentes há junções intercelulares especializadas, os discos intercalares,
que permitem a disseminação rápida dos estímulos contrácteis entre as células, o que permite uma
contracção quase instantânea e simultânea das fibras adjacentes. Algumas destas células
encontram-se modificadas e constituem as zonas de controlo da contracção muscular, constituindo o
sistema de Purkinje.
Músculo esquelético
À observação em microscopia de luz, o músculo esquelético em corte transversal, apresenta-se com
as fibras musculares com forma poliédrica com os núcleos periféricos. Os espaços endomisiais
podem-se encontrar aumentados como artefacto da contracção do tecido. Nestes espaços podem-se
observar inúmeros capilares.
As fibras dos tipos I e II podem ser identificadas pela actividade da ATPase da miosina, que
apresenta uma diferente estrutura proteica nos dois casos. As fibras do tipo I numa coloração de
imunohistoquímica para ATPase apresentam-se escuras e as fibras do tipo II claras.
O tecido muscular esquelético compreendendo cerca de 40% da massa muscular corporal, é
responsável pela contracção muscular voluntária, assim designado pela sua inserção
maioritariamente osteo-esquelética. Este está constituído por feixes musculares, que são o conjunto
das fibras musculares (células musculares), que contêm miofibrilhas compostas por miofilamentos. O
Histologia Básica - Serviço de Patologia Experimental . FMUC!
10/15
Apontamentos de Apoio ao Estudo
O presente texto constitui apenas um auxiliar de estudo, não devendo substituir a consulta da
bibliografia recomendada e das matérias leccionadas nas aulas.
tecido conjuntivo envolvente é um tecido conjuntivo elástico que rodeia o músculo (epimísio), os
feixes musculares (perimísio) e as fibras musculares (endomísio).
O músculo esquelético é composto por várias fibras, com 10 a 80 mm de diâmetro. Cada uma destas
fibras é constituída por várias subunidades menores. Em quase todos os músculos esqueléticos,
cada fibra estende-se ao longo de todo o comprimento do músculo. Cada fibra muscular é invervada
por uma só terminação nervosa, localizada no centro da fibra. A membrana citoplasmática da fibra
muscular é designada por sarcolema, constituída por uma membrana citoplasmática verdadeira,
rodeada de uma fina camada de polissacarídeos e numerosas fibrilhas de colagénio. Na terminação
de cada fibra muscular, a camada superficial do sarcolema funde-se com uma fibra tendinosa, para
constituir as fibras tendinosas nas inserções ósseas.
Cada miofibrilha, é constituída por cerca de 1500 filamentos de miosina e 3000 filamentos de actina,
polímeros proteicos responsáveis pela contracção muscular. A observação de microfotográfica
electrónica, permite a observação de bandas mais claras que contêm apenas filamentos de actina
(bandas I). As bandas escuras contêm filamentos de miosina e terminações de filamentos de actina
que se sobrepõem à miosina, sendo designadas por bandas A.
As pontes cruzadas prolongam-se da superfície dos filamentos de miosina ao longo de toda a
extensão do filamento, excepto na zona mais central. A interacção entre estas pontes cruzadas é
responsável pela contracção muscular.
No final dos filamentos de actina, encontram-se ligados discos Z, constituídos por proteínas
filamentosas diferentes da actina e da miosina, cruza-se com as miofibrilhas, interligando-as através
de toda a fibra muscular. Destes discos, estes filamentos estendem-se em ambas as direcções para
se relacionarem com os filamentos de miosina. A porção de fibra muscular que está compreendida
entre dois discos Z sucessivos é designada por sarcómero, que mede cerca de dois micrómetros
quando o músculo se encontra relaxado.
As moléculas de tropomiosina, são responsáveis pelo alinhamento dos filamentos de miosina e actina
e permite a contractibilidade do sarcómero, através da manutenção da relação entre a actina e a
miosina.
As miofibrilhas, encontram-se suspensas no interior da fibra muscular, no sarcoplasma. Este, é rico
em potássio, magnésio, fosfato e proteínas enzimáticas.
Histologia Básica - Serviço de Patologia Experimental . FMUC!
11/15
Apontamentos de Apoio ao Estudo
O presente texto constitui apenas um auxiliar de estudo, não devendo substituir a consulta da
bibliografia recomendada e das matérias leccionadas nas aulas.
Mecanismo geral da contracção muscular
Quando há a libertação de um estímulo através da fibra nervosa motora, com a libertação de
acetilcolina, ocorre a entrada de grandes quantidades de iões sódio no interior da fibra muscular,
através de canais controlados pela acetilcolina. Com a entrada dos iões sódio, há a formação de um
potencial de acção na fibra muscular, que viaja ao longo da sua membrana, com despolarização
desta. Este potencial de acção viaja então até ao interior da célula e provoca a libertação de grandes
quantidades de cálcio do interior do retículo sarcoplasmático, que é particularmente desenvolvido nas
fibras de contracção rápida. Os iões cálcio são responsáveis pela formação de forças atractivas entre
os filamentos de actina e de miosina e provocam o seu deslizamento um sobre o outro. Quando os
iões de cálcio são bombeados novamente para o interior do retículo sarcoplasmático, cessa a
contracção muscular.
Filamento de Miosina
Cada filamento de miosina é formado por cerca de 200 moléculas de miosina, cada uma constituída
por seis cadeias polipeptídicas: duas cadeias pesadas que formam uma dupla hélice e quatro cadeias
leves. A terminação de cada uma destas cadeias está ligada a um polipéptido globular que constituí a
chamada “cabeça de miosina”.
As cabeças de miosina formam as pontes cruzadas, que se estendem em todas as direcções à volta
do filamento e funcionam como as ATPases.
Filamento de Actina
Os filamentos de actina possuem duas cadeias enroladas em hélice, constituída por moléculas de
actina G polimerizadas. Em cada volta da hélice, existem cerca de 13 moléculas de actina G, sendo
que a cada uma delas se encontra ligada uma molécula de ADP, constituindo o local activo onde
actuam as pontes cruzadas da miosina. As bases dos filamentos de actina estão fortemente inseridas
no disco Z.
As moléculas de Tropomiosina
As moléculas de tropomiosina encontram-se enroladas em espiral à volta da hélice de actina. No
estado de repouso, ligam-se aos locais activos para evitar a atracção entre os filamentos de actina e
de miosina. Para que ocorra esta interacção, a tropomiosina tem que ser mobilizada e deixar a
Histologia Básica - Serviço de Patologia Experimental . FMUC!
12/15
Apontamentos de Apoio ao Estudo
O presente texto constitui apenas um auxiliar de estudo, não devendo substituir a consulta da
bibliografia recomendada e das matérias leccionadas nas aulas.
descoberto os locais activos da actina. Este acontecimento ocorre quando o cálcio liga a troponina C,
que puxa a tropomiosina e deixa a descoberto os locais de ligação da miosina, para que ocorra
contracção muscular.
A Troponina
A troponina é formada por complexos de três unidades proteicas, cada uma delas com um papel
importante no controlo da contracção muscular: troponina I, com função inibitória; troponina C, capaz
de ligar cálcio; troponina T, que liga a tropomiosina.
!
Interacção da Miosina, Filamentos de Actina e Cálcio: O potencial de membrana da fibra muscular é
de -90mV, sendo por isso uma célula dificilmente estimulável. A contracção surge apenas de forma
dependente da vontade.
Despolarização da célula: Como a célula muscular possui uma grande dimensão, e o potencial de
membrana tem que viajar até ao retículo sarcoplasmático, é transmitido por túbulos transversos
(invaginações de membrana celular até ao interior da célula). O cálcio é libertado do retículo
sarcoplasmático por difusão simples, através de canais de cálcio.
Um filamento de actina puro, sem a presença do complexo de troponina-tropomiosina, liga de forma
forte e instantânea às cabeças das moléculas de miosina na presença de iões magnésio e de ATP. Os
locais activos do filamento normal de atina de um músculo relaxado, encontram-se inibidos, cobertos
fisicamente pelo complexo troponina-tropomiosina, não permitindo a ligação das cabeças de miosina
nos locais devidos para haver contracção.
Na presença de grandes quantidades de iões cálcio, o efeito inibitório do complexo troponinatropomiosina do filamento de actina é inibido. Quando os iões cálcio combinam com a troponina C,
provocam uma alteração conformacional no complexo de troponina, que induz a tropomiosina a entrar
nas duas cadeias de actina. Este movimento, descobre os locais activos para a actina, atraindo as
cabeças de miosina e permitindo a contracção muscular. Quando uma cabeça de miosina liga no
local activo correspondente, na actina, provoca alterações profundas nas forças intermoleculares
entre a cabeça e braço da ponte cruzada. O novo alinhamento de forças obriga a cabeça a oscilar
sobre o braço, arrastando consigo o filamento de actina. Logo de seguida, a cabeça desliga do local
activo, voltando à sua posição natural. A cabeça de miosina desliga-se da actina, quando se liga uma
molécula de ATP. Combina então com um novo local activo ao longo do filamento de actina.
Histologia Básica - Serviço de Patologia Experimental . FMUC!
13/15
Apontamentos de Apoio ao Estudo
O presente texto constitui apenas um auxiliar de estudo, não devendo substituir a consulta da
bibliografia recomendada e das matérias leccionadas nas aulas.
Depois, para haver relaxamento, a troponina que ligou quatro iões cálcio, tem que os libertar através
de uma bomba de cálcio. Todo este processo mediado pelo cálcio que permite a ligação da cabeça
de miosina é conhecido como teoria da cremalheira ou deslizamento.
Músculo Liso
O tecido muscular liso é responsável pela contracção involuntária dos órgãos internos (vísceras – no
tubo digestivo, nas vias respiratórias e urogenitais, vasos sanguíneos, derme – músculos erectores
dos pelos, músculos intrínsecos do olho, músculo dartos do escroto). Em alguns casos particulares,
encontra-se algum tecido muscular liso intercalado em músculo esquelético: músculos esfíncteres do
ânus e da bexiga urinária, músculos tarsais superior e inferior das pálpebras e o músculo orbital, o
ligamento suspensor do duodeno, regiões intermédias do esófago e fáscias e ligamentos na face
pélvica do diafragma pélvico. Não apresenta estriação, já que a actina e a miosina encontram-se
menos ordenadas e a sua contracção, embora possa ser de maior duração, é mais lenta do que no
músculo estriado. A sua inervação provém do sistema nervoso autónomo. É constituído por miócitos
fusiformes mononucleados, dispostos em longos eixos paralelos à direcção de contracção,
organizados em pequenos fascículos separados por tecido conjuntivo.
De órgão para órgão as características do músculo liso são diferentes, consoante a função
fundamental a desempenhar: dimensões, organização das fibras, resposta a diferentes tipos de
estímulo, inervação e função. Assim, encontramos tecido muscular liso unitário (do tipo visceral) e
tecido muscular liso tipo unidades múltiplas. No tecido muscular liso tipo unidades múltiplas, cada
fibra opera independentemente das outras, inervada por uma só terminação nervosa, cujos sinais
nervosos são a principal fonte de controlo, mas permitindo que ocorram alguns fenómenos de
contracção espontânea. Um exemplo desta situação é o músculo constrictor da íris.
O tecido muscular liso tipo visceral é constituído por inúmeras fibras que contactam entre si,
contraindo-se simultaneamente. As células encontram-se ligadas por junções, através das quais os
iões podem passar de forma livre de uma célula para a seguinte. A inervação não é individualizada
para cada fibra, e a contracção pode ocorrer quando há a libertação e difusão de um
neurotransmissor à distância, que é lenta e ocorre em uníssono.
As fibras musculares lisas são células alongadas, fusiformes, com extremidades afiladas que podem
por vezes ser bifurcadas. São habitualmente fibras mais curtas do que as fibras musculares
esqueléticas e apresentam um núcleo alongado e localizado centralmente. Dependendo do estado de
contracção no momento da fixação, os núcleos podem por vezes surgir com uma forma espiralada.
As fibras musculares lisas encontram-se paralelas umas às outras, agrupadas em fascículos com
ramificações irregulares, que constituem unidades funcionais contrácteis.
Histologia Básica - Serviço de Patologia Experimental . FMUC!
14/15
Apontamentos de Apoio ao Estudo
O presente texto constitui apenas um auxiliar de estudo, não devendo substituir a consulta da
bibliografia recomendada e das matérias leccionadas nas aulas.
No corte longitudinal (microscopia de luz), as fibras encontram-se muitas vezes dispostas de um
modo tão regular e denso que pode ser impossível identificar os limites das individuais de cada
célula. No corte transversal, porque este pode seccionar as células em diferentes níveis, dá a ilusão
de cada uma delas possuir diâmetros diferentes. Os núcleos apresentam uma localização central e
forma arredondada.
O músculo liso também contém actina e miosina, com características semelhantes às do músculo
estriado, mas com um mecanismo de controlo diferente, já que não possui troponina. O processo de
contracção e as interacções entre as proteínas filamentosas é semelhante, estando dependente da
activação pelos iões de cálcio e utilizando ATP para que ocorra.
Os filamentos de actina encontram-se ligados a corpos densos, ligados à membrana celular ou
dispersos no interior da célula, mantidos na posição correcta por proteínas estruturais que interligam
os diferentes corpos densos, podendo mesmo ocorrer a interligação de corpos densos pertencentes a
células diferentes e que permite a transmissão intercelular da força de contracção. Os filamentos de
actina são os mais numerosos, encontrando-se entre eles alguns filamentos de miosina, que
apresentam pontes cruzadas com uma orientação polar, o que permite à miosina puxar o filamento de
actina numa direcção e na outra extremidade da fibra, traccionar o filamento de actina na direcção
oposta.
A contracção do músculo liso é mais prolongada, sendo que o ciclo de pontes cruzadas da miosina é
mais lento, o que motiva uma maior lentidão no início da contracção, que começa alguns
milissegundos depois da estimulação. O músculo liso necessita de menos energia para manter a
contracção, que é capaz de se perpetuar durante mais tempo.
O músculo liso não tem troponina, pelo que a combinação dos iões de cálcio com a calmodulina
permite a activação da miosina e fosforilação da cabeça de miosina e ganha capacidade de se ligar
ao filamento de actina. Quando a concentração iónica de cálcio cai abaixo de um nível critico, todo o
processo regride, excepto para a fosforilação da cabeça de miosina que, mediante a activação da
miosina fosfatase permite a ligação de fosfato à cadeia leve reguladora
A via aeróbia é a via energética utilizada por excelência.
Histologia Básica - Serviço de Patologia Experimental . FMUC!
15/15