INSTITUTO AGRONÔMICO
CURSO DE PÓS-GRADUAÇÃO EM AGRICULTURA
TROPICAL E SUBTROPICAL
QUALIDADE E PRODUTIVIDADE DA CANA-DEAÇÚCAR INOCULADA COM GLUCONACETOBACTER
DIAZOTROPHICUS NO E ADUBADA COM NITROGÊNIO
MINERAL E ORGÂNICO
JOSÉ PAULO QUEIROZ PRADO JÚNIOR
Orientador: Raffaella Rossetto
Co-orientador: Lee Tseng Sheng Gerald
Dissertação submetida como requisito
parcial para a obtenção do grau de
Mestre em Agricultura Tropical e
Subtropical Área de Concentração em
Tecnologia de Produção Agrícola
Campinas, SP
Janeiro 2008
i
Ficha elaborada pela bibliotecária do Núcleo de Informação e Documentação
do Instituto Agronômico
P896q Prado Junior, José Paulo Queiroz
Qualidade e produtividade da cana-de-açúcar inoculada com
Gluconacetobacter diazotrophicus e adubada com nitrogênio mineral
e orgânico./ José Paulo Queiroz Prado Junior. Campinas, 2008.
49 fls
Orientador: Raffaella Rossetto
Dissertação (Mestrado) Agricultura Tropical e Subtropical
Instituto Agronômico
1.
Cana-de-açúcar
2.
Saccharum
officinarum
L.
3
Gluconacetobacter
Diazotrophicus. 4. Inoculação 5. Bactérias fixadoras de nitrogênio,
6. Adubação nitrogenada. I. Rossetto, Raffaella II. Campinas. Instituto
Agronômico III. Título
CDD. 633.61
ii
A minha família
José Paulo, Vaima e Erick
DEDICO
A Raffaella Rossetto e Lee Tseng
Sheng Gerald
OFEREÇO
iii
AGRADECIMENTOS
-
A Deus que nos da força para a batalha do dia a dia;
-
Ao Dr. Victor Branco de Araújo pela utilização do Laboratório de Biotecnologia
Vegetal da Coordenadoria de Assistência Técnica Integral-CATI, localizado em
Tiete-SP;
-
Aos funcionários do IAC-APTA Pólo Centro-Sul, do Laboratório de
Biotecnologia Vegetal da CATI e da UFSCar-Araras;
-
A Coordenadoria de Aperfeiçoamento Pessoal de Nível Superior (CAPES), pela
concessão da bolsa de estudos;
-
A minha família José Paulo Queiroz Prado, Vaima Maria Alberto Queiroz Prado
e Erick Alberto Queiroz Prado pois sem eles nada disso seria possível;
-
Ao Prof. Dr. Lee Tseng Sheng Gerald que me ensinou muito sobre pesquisa e
que se tornou um grande amigo;
-
A Dra. Raffaella Rossetto, uma pessoa espetacular que encontrou tempo para me
orientar neste trabalho;
-
Ao Engenheiro Agrônomo e amigo Diogo Miranda pelo auxílio fundamental na
quantificação das bactérias;
-
Aos funcionários da PG-IAC pelo auxílio e amizade durante o curso;
-
A todos que colaboraram para a realização deste trabalho.
iv
SUMÁRIO
ÍNDICE DE TABELAS ............................................................................................... vi
ÍNDICE DE FIGURAS............................................................................................... vii
RESUMO .................................................................................................................. viii
ABSTRACT ................................................................................................................ ix
1 INTRODUÇÃO ......................................................................................................... 1
2 REVISÃO BIBLIOGRÁFICA ................................................................................... 4
2.1 Importância da Fixação Biológica de Nitrogênio na Agricultura .............................. 4
2.2 Importância do Nitrogênio na Nutrição e Fisiologia da Cana-de-Açúcar .................. 5
2.3 Adubação Nitrogenada e Cana-Planta ..................................................................... 6
2.4 Fixação Biológica de Nitrogênio na Cana-de-Açúcar .............................................. 7
2.4.1 Bactérias Fixadoras de Nitrogênio ...................................................................... 10
2.4.2 Interação Variedades e Fixação Biológica de Nitrogênio .................................... 12
2.4.3 Inoculação de Bactérias Diazotróficas ................................................................ 14
2.4.4 Interação Adubação e Fixação Biológica de Nitrogênio ...................................... 16
3 MATERIAL E MÉTODOS...................................................................................... 18
3.1 Quantificação Prévia das Bactérias Diazotróficas em 6 Variedades de Cana .......... 18
3.2 Local do Experimento ........................................................................................... 19
3.3 Tratamentos e o Delineamento Experimental......................................................... 20
3.4 Preparo do Inóculo e a Inoculação ......................................................................... 21
3.5 Análise Foliar........................................................................................................ 23
3.6 Quantificação das Bactérias Diazotróficas Endofíticas .......................................... 23
3.7 Produtividade e Análise Tecnológica..................................................................... 25
3.8 Análise Estatística ................................................................................................. 25
4 RESULTADOS E DISCUSSÃO .............................................................................. 26
4.1 Quantificação Prévia das Bactérias Diazotróficas .................................................. 26
4.2 Produtividade ........................................................................................................ 26
4.3 Análise Tecnológica .............................................................................................. 30
4.4 Quantificação das Bactérias Fixadoras de Nitrogênio ............................................ 31
4.5 Análise Foliar........................................................................................................ 33
5 CONCLUSÕES ....................................................................................................... 34
6 REFERÊNCIAS BIBLIOGRÁFICAS...................................................................... 35
v
ÍNDICE DE TABELAS
Tabela 1 -
Aspectos relacionados com o processo de fixação do nitrogênio
atmosférico por via industrial e biológica..........................................
4
Tabela 2 -
Porcentagem de nitrogênio proveniente da FBN na variedade
RB72454 em diversas regiões produtoras no Brasil.......................... 14
Tabela 3 -
Metodologia de aplicação reportados para algumas bactérias
diazotróficas....................................................................................... 15
Tabela 4 -
Características químicas (macronutrientes) do solo onde foi
instalado o experimento*................................................................... 19
Tabela 5 -
Características químicas (micronutrientes) do solo onde foi
instalado o experimento..................................................................... 19
Tabela 6 -
Características varietal de RB72454 e IACSP93-6006...................... 20
Tabela 7 -
Tratamentos estudados no experimento.............................................
Tabela 8 -
Quantificação prévia das variedades disponíveis na época de
plantio (% de amostras com presença das bactérias)......................... 26
Tabela 9 -
Produtividade expressa em toneladas de colmos por hectare............. 27
Tabela 10 -
Contrastes ortogonais dos dados relativos a produtividade das duas
variedades estudadas.......................................................................... 28
Tabela 11 -
Aplicação do Teste de Dunnett para a produtividade......................... 29
Tabela 12 -
Parâmetros tecnológicos medidos aos 18 meses após a instalação
do experimento, variedade RB72454................................................. 30
Tabela 13 -
Parâmetros tecnológicos medidos aos 18 meses após a instalação
do experimento, variedade IACSP93-6006........................................ 31
Tabela 14 -
Bactérias diazotróficas (logaritmo do número de células por grama
de matéria fresca) em duas variedades de cana-de-açúcar aos 16
meses.................................................................................................. 32
Tabela 15 -
Nitrogênio total na folha +1 aos 8 meses após a instalação do
experimento........................................................................................ 33
21
vi
ÍNDICE DE FIGURAS
Figura 1 -
Possíveis locais de infecção e colonização das bactérias
diazotróficas nas raízes ...................................................................
9
Células do colmo da cana-de-açúcar colonizadas por
Gluconacetobacter diazotrophicus..................................................
10
Colmos de cana mergulhados em meio sólido. A coloração azul
indica o crescimento de Herbaspirillum spp. e a coloração
amarela
indica
crescimento
de
Gluconacetobacter
diazotrophicus..................................................................................
18
Garrafa PET 2 litros com Gluconacetobacter diazotrophicus
pronto para ser colocado no pulverizador costal..............................
22
Figura 5 -
Pulverização da bactéria no sulco de plantio....................................
22
Figura 6 -
Detalhe da inoculação. A pulverização é dirigida ao corte recém
feito no colmo pois a entrada da bactéria é facilitada em
ferimentos.........................................................................................
23
Metodologia do Número Mais Provável, utilizando frascos de
penicilina de 10 ml com meio semi-sólido......................................
24
Meio de cultura semi-sólido LGI-P. Os dois frascos da direita
estão positivos quanto ao crescimento de Gluconacetobacter
diazotrophicus...................................................................................
24
Meio de cultura semi-sólido JNFb. Os dois frascos da direita estão
positivos quanto ao crescimento de Herbaspirillum spp..................
25
Produtividade entre os tratamentos adubados com nitrogênio
orgânico, nitrogênio mineral e testemunha sem nitrogênio,
independente da inoculação..............................................................
29
Figura 2 Figura 3 -
Figura 4 -
Figura 7 Figura 8 -
Figura 9 Figura 10 -
vii
PRADO JUNIOR, José Paulo Queiroz. Qualidade e produtividade da cana-deaçúcar inoculada com Gluconacetobacter diazotrophicus e adubada com nitrogênio
mineral e orgânico. 2008. 49f. Dissertação (Mestrado em Tecnologia da Produção
Agrícola)-Pós-Graduação-IAC.
RESUMO
O presente trabalho teve por objetivo avaliar o comportamento de duas variedades de
cana-de-açúcar frente à inoculação de Gluconacetobacter diazotrophicus no sulco de
plantio e a aplicação de fontes nitrogenadas. Para isso foi instalado um experimento de
campo em delineamento de blocos ao acaso com 12 tratamentos e 4 repetições, sendo os
tratamentos os seguintes: duas variedades de cana-de-açúcar RB72454 e IACSP936006; inoculação ou não com Gluconacetobacter diazotrophicus estirpe BR 11281;
ausência e presença de adubação nitrogenada como torta de filtro e uréia. As variáveis
analisadas foram: análise foliar (8 meses), quantificação das bactérias fixadoras de
nitrogênio (16 meses), produtividade (18 meses) e características tecnológicas da cana.
As variedades IACSP93-6006 e RB72454 tiveram produtividade favorecida tanto pelo
nitrogênio como pela inoculação. As análises tecnológicas realizadas não indicaram
diferenças entre os tratamentos em nenhuma das variedades estudadas. A inoculação e
as adubações nitrogenadas não interferiram no acúmulo de sacarose e nos açúcares
totais produzidos pela cana. A contagem de bactérias foi estatisticamente diferente
apenas na variedade RB72454 para Herbaspirillum spp. e Gluconacetobacter
diazotrophicus. Entretanto, o tratamento com maior contagem de bactérias não
corresponde exatamente ao tratamento com maior ganho de produtividade, indicando
que apenas a contagem de bactérias não reflete o potencial de fixação da associação. O
número de bactérias presentes não foi inibido pelo de nitrogênio mineral e nem pelo
nitrogênio orgânico. Os tratamentos estudados não aumentaram o teor de nitrogênio nas
folhas. Conclui-se que ambas as variedades estavam colonizadas pelas bactérias,
apresentaram ganhos na produtividade quando se procedeu a inoculação no sulco de
plantio ou a adição de fonte nitrogenada e as bactérias não foram inibidas pela presença
de nitrogênio mineral.
Palavras-chave: Saccharum officinarum L., Gluconacetobacter diazotrophicus,
inoculação, bactérias fixadoras de nitrogênio, adubação nitrogenada.
viii
PRADO JUNIOR, José Paulo Queiroz. Sugarcane quality and yield inoculated with
Gluconacetobacter diazotrophicus and fertilized with mineral and organic nitrogen.
2008. 49f. Dissertação (Mestrado em Tecnologia da Produção Agrícola)-PósGraduação-IAC.
ABSTRACT
The objective of this research was to evaluate the Gluconacetobacter diazotrophicus
inoculation efficiency in two sugarcane varieties and nitrogen sources application. A
field experiment was carried out using a randomized block statistical design with 12
treatments and 4 replications. The treatments were: 2 sugarcane varieties RB72454 and
IACSP93-6006; inoculation or not with Gluconacetobacter diazotrophicus lineage BR
11281 and nitrogen application with urea and filter cake. The parameters measured was:
foliar analysis (8 months); yield at 18 months; technological characteristics of the
sugarcane and quantification of the nitrogen fixing bacteria (16 months). The results
showed that the sugarcane variety RB72454 and IACSP93-6006 yield was higher due to
inoculation or nitrogen application. It was not observed differences between treatments
for technological analyses. Also no differences were observed for inoculation or the
nitrogen application in the sacaroses and the total sugar contents. The number of
nitrogen fixing bacteria evaluation showed differences between treatments but the
higher number of bacteria don’t correspond exactly with the higher yield, showing that
number of bacteria evaluation does not reflect the potential of the nitrogen fixation
association. The number of bacteria was not inhibited by the nitrogen application. It was
concluded that both varieties were colonized by fixing nitrogen bacteria. The both
variety yield was increased by the nitrogen fixing bacteria inoculation and in the furrow
and with the nitrogen fertilizer addition. The nitrogen fixing bacteria had not been
inhibited by the nitrogen mineral presence.
Key words: Saccharum officinarum L., Gluconacetobacter diazotrophicus, inoculation,
nitrogen fixing bacteria, nitrogen fertilization.
ix
1 INTRODUÇÃO
As perspectivas para o setor sucroalcooleiro são as mais otimistas. Desde a
implantação do Proálcool, em 1975, nunca se registrou um horizonte tão promissor
como o que se vislumbra para os próximos anos. O Brasil é o maior produtor e
exportador mundial de açúcar, detém 40% do comércio internacional e deve ampliar
essa participação com a reforma do regime açucareiro na União Européia, determinada
pela OMC - Organização Mundial do Comércio. O novo regime prevê a retirada dos
subsídios para a produção de açúcar (ORTOLAN, 2006).
No entanto, será o álcool o principal propulsor do crescimento do setor por dois
motivos: principalmente pelo aumento da demanda interna face ao crescimento da frota
de veículos leves biocombustíveis e também pela valorização do combustível brasileiro
no cenário internacional. O último motivo influenciado principalmente pelo declínio das
reservas mundiais de petróleo e pela busca por um combustível renovável que
represente menores emissões de CO2 na atmosfera.
A safra brasileira de cana-de-açúcar atingiu no ano agrícola de 2006/2007 427
milhões de toneladas, sendo a região Centro-Sul responsável por 86% do total
produzido. O crescimento da área colhida foi de 6,4% sobre a safra anterior, atingindo
5,812 milhões de hectares contra os 5,462 milhões estimados para a safra anterior. A
produção de açúcar cresceu 7,8%, atingindo 29,99 milhões de toneladas e produção de
álcool foi de 17,9 bilhões de litros (FNP CONSULTORIA & COMÉRCIO, 2007).
As projeções atuais apontam para um crescimento de aproximadamente 83% na
área plantada com cana nos próximos 10 anos. No mesmo período prevê-se um
crescimento de 100% na produção, atingindo em torno de 1 bilhão de toneladas na safra
2017/2018. Atualmente existem 320 usinas em funcionamento e 90 em construção. Nos
próximos 10 anos, pelo menos uma centena de outras novas usinas deverão ser
instaladas (FNP CONSULTORIA & COMÉRCIO, 2007).
Os primeiros programas de melhoramento genético da cana-de-açúcar no Brasil
tinham por base solos de baixa fertilidade com baixa aplicação de nitrogênio,
selecionando materiais adaptados a essas condições. A baixa aplicação de nitrogênio
selecionou naturalmente variedades que se associam naturalmente a bactérias
diazotróficas endofíticas podendo obter parte de seu nitrogênio através da fixação
biológica. Ainda não está claro qual o mecanismo que está envolvido no
1
estabelecimento da interação planta-bactéria, faltam detalhes no processo metabólico e
pouco se sabe sobre o papel da planta nesta associação.
Uma das práticas aplicadas na cana-de-açúcar é o uso de torta de filtro
proveniente do processo de produção de açúcar. Este subproduto industrial é
considerado um excelente fertilizante, além de propiciar melhores condições para
brotação da cana. A adição de materiais orgânicos ao solo estimula a decomposição da
matéria orgânica do próprio solo. Nesse caso, o nitrogênio do solo também é
mineralizado e tornado disponível às plantas, processo que pode ser acentuado pela
adição de fertilizantes nitrogenados.
Para aumentar a produção agricultores aplicam mais de 85 milhões de toneladas
de nitrogênio por ano no mundo todo (ASSOCIAÇÃO NACIONAL PARA DIFUSÃO
DOS ABUDOS, 2004). Sua produção e aplicação somam mais da metade da energia
consumida na agricultura. Grande parte do nitrogênio aplicado como fertilizante não é
utilizado pelas culturas, permanecendo no solo ou perdendo-se por lixiviação para águas
subterrâneas ou por volatilização para a atmosfera. Obviamente, aumentar a eficiência
com que as plantas obtêm nitrogênio é de suma importância (EPSTEIN & BLOOM,
2006).
A fixação natural de nitrogênio (atmosférica mais biológica) se dá a uma taxa de
190 x 1012 gramas de nitrogênio ao ano. Desse total, a emissão de relâmpagos é
responsável por 8%. Um adicional de 2% do nitrogênio fixado naturalmente deriva da
reação fotoquímica entre o óxido nítrico gasoso e o ozônio resultando em ácido nítrico.
Os 90% restantes resultam da fixação biológica de nitrogênio, em que bactérias fixam o
nitrogênio molecular em amônia (EPSTEIN & BLOOM, 2006).
O nitrogênio é constituinte de aminoácidos, proteínas e ácidos nucléicos,
participando direta e indiretamente de diversos processos bioquímicos das plantas,
podendo ser encontrado também na forma inorgânica (NO3- e NH4+). O nitrato é
prontamente absorvido, convertido por enzimas e em contato com carboidratos forma as
amidas e aminoácidos, passando então da forma inorgânica para a orgânica
(FERNANDES & ROSSIELLO, 1995).
No começo da década de 90 e principalmente após a reunião da ONU conhecida
como ECO 92, a agricultura passou a incorporar o conceito de sustentabilidade. Os
preceitos de rendimento e lucratividade que sempre nortearam as atividades agrícolas
até então, tiveram que conviver com os conceitos de sustentabilidade, questões
ambientais e qualidade de vida. A agricultura altamente impactante terá que ser
2
substituída por técnicas alternativas que visem minimizar problemas ambientais. Neste
sentido, a fixação biológica de nitrogênio atende os anseios da agricultura de baixos
insumos, perfeitamente de acordo com os modernos conceitos de Mecanismos de
Desenvolvimento Limpo (MDL).
Estudar as bactérias naturalmente presentes e inocular em condições de campo
promove um passo para o entendimento das diversas interações que possam ocorrer e
para o desenvolvimento da tecnologia de inoculantes, como ocorre no caso da soja e do
arroz (NGUYEN et al., 2003).
Os objetivos deste trabalho foram estudar a eficiência da inoculação de bactérias
fixadoras de nitrogênio no sulco de plantio, o desenvolvimento das bactérias na
presença de nitrogênio orgânico e mineral e avaliar a produtividade da cana-de-açúcar
frente à inoculação e adubação com nitrogênio mineral e orgânico.
3
2 REVISÃO BIBLIOGRÁFICA
2.1 Importância da Fixação Biológica de Nitrogênio na Agricultura
O gás nitrogênio presente em grande abundância na atmosfera apresenta uma
tripla ligação entre os átomos que o torna extremamente estável a temperatura ambiente
(SPRENT & SPRENT, 1990). Apesar da abundância de N2 na atmosfera, os organismos
que pertencem ao grupo dos eucariotos não conseguem utilizar este elemento
diretamente.
Apenas uma parte dos organismos do grupo dos procariotos consegue reduzir
enzimaticamente o nitrogênio da atmosfera em amônia, a qual pode ser incorporada
para o crescimento e manutenção das células. Estes organismos são denominados
diazotróficos e o mecanismo responsável pela incorporação de nitrogênio à biomassa é
chamado de fixação biológica de nitrogênio (FBN). A FBN é o processo responsável
pela incorporação do nitrogênio atmosférico à matéria viva, ao longo da evolução do
nosso planeta. A Tabela 1 mostra alguns aspectos relacionados com a fixação industrial
e biológica.
Tabela 1-Aspectos relacionados com o processo de fixação do nitrogênio atmosférico
por via industrial e biológica
Fixação industrial
Fixação biológica de nitrogênio
(fertilizantes)
(FBN)
Nutriente mais caro, utiliza energia fóssil Segundo
para produção e distribuição, aumentando importante
processo
do
biológico
planeta
mais
depois
da
o CO2 atmosférico e o aquecimento global fotossíntese. Não é poluente, utiliza energia
solar, recicla CO2
Tem baixo aproveitamento agronômico e Mecanismo responsável por 65% do N2
são poluentes de solo, água e atmosfera
incorporado nos seres vivos do planeta
Representam de 5 a 20% do custo de Consome em torno de 2,5% da energia da
produção das culturas
fotossíntese do planeta
Representam apenas 2% da absorção total A vida no planeta terminaria em 30 anos se
de nitrogênio pelas plantas
a fixação biológica do nitrogênio parasse
Fonte: Franco e Balieiro (1999).
4
Um dos objetivos da agricultura sustentável é o aproveitamento eficiente do
nitrogênio atmosférico (GRAHAN & VANCE, 2000). Dentre os sistemas biológicos
capazes de aproveitar o nitrogênio da atmosfera, a simbiose rizóbio–leguminosa tornouse o sistema mais especializado responsável por 22% de todo o nitrogênio incorporado
ao ecossistema terrestre (PEOPLES et al., 1995). No Brasil salienta-se a importância
econômica e ecológica da fixação biológica do nitrogênio na cultura da soja, que
dispensa totalmente a adubação nitrogenada sem causar perda de produtividade.
2.2 Importância do Nitrogênio na Nutrição e Fisiologia da Cana-de-Açúcar
Apesar do nitrogênio contribuir com 1 a 1,5% na matéria seca da cana-deaçúcar, seu papel é tão importante quanto o do carbono, hidrogênio e oxigênio que
constituem, juntos, mais de 90% da matéria seca. No solo, o nitrogênio disponível às
plantas é suprido pela mineralização da matéria orgânica, fixação biológica do
nitrogênio e adição de fertilizantes nitrogenados. A água de irrigação ou de chuva
contém quantidades variáveis de nitrogênio mineral, que pode ser absorvido por raízes e
folhas. Outra fonte de nitrogênio para a cana é o nutriente contido no tolete de plantio
(TRIVELIN, 2000).
Apesar dos solos das regiões tropicais apresentarem baixos níveis de nitrogênio,
no Brasil aplica-se apenas 60 kg ha-1 de nitrogênio na cana-planta, porém as soqueiras
respondem a doses mais altas deste nutriente, entre 100 e 150 kg ha-1 (URQUIAGA et
al, 1992; PENATTI, 1994).
Alguns levantamentos apontam que a cultura da cana acumula entre 100 e 200
-1
kg ha de nitrogênio por estação de crescimento. Parte do nitrogênio incorporado pela
planta é exportada na colheita e parte é perdida pela queima das folhas. Dessa forma, as
extrações contínuas causariam uma exaustão deste nutriente no solo causando acentuada
queda de produtividade caso não houvesse a reposição do nitrogênio. No sistema de
cultivo da cana sem despalha a fogo grande quantidade deste nutriente retorna ao
sistema através da manutenção e incorporação da palhada.
O nitrogênio é importante na nutrição e na fisiologia da cana-de-açúcar pois é
constituinte dos aminoácidos, proteínas, enzimas e ácidos nucléicos (MALAVOLTA et
al., 1989) e juntamente com o potássio são exigidos em maiores quantidades pela
cultura (ORLANDO FILHO et al., 1980; SILVEIRA, 1980).
5
A assimilação de nitrogênio é estritamente relacionada com o metabolismo de
carboidratos e provoca acentuada diminuição nos teores de açúcares do colmo, com
menor expressão na bainha (SILVEIRA, 1985).
A cana por ser do ciclo fotossintético C-4, responde às maiores temperaturas
com aumento na taxa de fotossíntese, desde que a disponibilidade de água, nutrientes e
concentração de O2 e CO2 na atmosfera não sejam limitantes.
O estresse de nitrogênio, mesmo que não se manifeste por sintomas visuais,
reduzirá a atividade da enzima Rubisco, responsável pela assimilação do carbono no
Ciclo de Calvin e Benson localizados nas células do feixe da bainha, muito mais que a
atividade da enzima PEPC, situada nas células do mesofilo foliar e responsável pela
fixação do CO2 no ciclo C-4. O CO2 descarboxilado e não fixado no ciclo do carbono é
refixado no mesofilo pela PEPC, representando um gasto adicional de energia para a
planta, uma vez que ATP é requerida para a regeneração da PEPC (TRIVELIN, 2000).
O acúmulo de nitrogênio pela cana-de-açúcar varia de acordo com o cultivar,
com a idade da cultura, disponibilidade do nitrogênio e de outros elementos na solução
do solo e de fatores edafoclimáticos (CLEMENTS, 1980; MUCHOW & ROBERTSON,
1994; WOOD et al., 1996).
2.3 Adubação Nitrogenada e Cana-Planta
O nitrogênio que é fornecido para a cana-planta raramente ultrapassa dosagens
de 60 kg ha-1. Se a cultura acumula entre 150 a 200 kg ha-1 (ORLANDO FILHO et al.,
1980) e supondo que nada dos 60 kg ha-1 seja perdido, deve existir uma outra fonte para
suprir o restante necessário para a planta.
Em estudos sobre o efeito da adubação nitrogenada em cana-planta, Azeredo et
al. (1986) verificaram que somente em 20% de 135 experimentos de campo conduzidos
no Brasil foram observados efeitos positivos que incrementaram a produção.
A análise de 81 experimentos conduzidos no Estado de São Paulo também
revelou baixa resposta da cana-planta à adubação nitrogenada pois em apenas 26
experimentos houve efeito da aplicação do fertilizante (CANTARELLA & RAIJ, 1986).
Rossiello (1987) também agrupou os resultados de 105 experimentos conduzidos
no Brasil no período de 1960 a 1986 sob a forma de rendimento relativo e verificou que
somente em 22 % houve resposta da cana-planta à adubação nitrogenada.
6
A falta de resposta da cana-planta tem sido atribuída à fixação biológica de
nitrogênio atmosférico e a maior mineralização do nitrogênio devido ao preparo do solo
que ocorre em condições climáticas favoráveis (clima quente e úmido). O sistema
radicular da cana-planta sendo mais vigoroso pode também justificar a maior eficiência
de utilização de nitrogênio do solo (TRIVELIN, 2000).
A produção de palha de um canavial sob colheita mecanizada, que inclui as
folhas, as bainhas e o ponteiro, além de quantidade variável de pedaços de colmo, pode
acrescentar cerca de 10 a 30 toneladas ha-1 de matéria seca. Esse material contém em
torno de 40 a 80 kg ha-1 de nitrogênio, que estará disponível para a cultura após a
mineralização (TRIVELIN, 2000).
As respostas da cana-planta à adubação nitrogenada ocorrem geralmente quando
se cultiva cana pela primeira vez, em cultivo mínimo ou quando há condições de
eutrofismo do solo. Para as soqueiras ocorre o inverso, cerca de 70% dos trabalhos
indicam respostas à adubação com nitrogênio (CÂMARA, 1993).
Uma das práticas aplicadas na cana-de-açúcar é o uso de torta de filtro
proveniente do processo de produção de açúcar. Este subproduto industrial é
considerado um excelente fertilizante, além de propiciar melhores condições para
brotação da cana. A adição de materiais orgânicos ao solo estimula a decomposição da
matéria orgânica do próprio solo. Nesse caso, o nitrogênio do solo também é
mineralizado e tornado disponível às plantas, processo que pode ser acentuado pela
adição de fertilizantes nitrogenados.
A composição química da torta de filtro é variável em função do processo de
produção do açúcar. Na média a torta de filtro possui a seguinte composição em função
da matéria seca: N = 1,4%; P2O5 = 1,9%; K2O = 0,4%; CaO = 2,1%; MgO = 0,5%; C =
7,4% e umidade = 75%. (COPERSUCAR, 1986).
2.4 Fixação Biológica de Nitrogênio na Cana-de-Açúcar
Atualmente são conhecidas mais de 140 espécies de bactérias diazotróficas,
incluindo cianobactérias e actinomicetos (YOUNG, 1992). Neste universo, apenas
quatro espécies são consideradas fitopatogênicas: Agrobacterium tumefaciens,
Burkholderia vietnamiensis, Herbaspirillum rubrisubalbicans e Pantoae herbicola.
Os diazotróficos endofíticos têm uma vantagem sobre os diazotróficos
associativos de raízes, uma vez que ocupa espaços mais intimamente ligados ao
7
hospedeiro com maior acesso às fontes de carbono. Além disso, eles colonizam nichos
protegidos do oxigênio, o qual é necessário para a expressão e atividade da nitrogenase
(DOBBELAERE et al., 2003).
A promoção do crescimento das plantas por bactérias endofíticas pode ser
resultante de ações indiretas, como por exemplo, a supressão de doenças ou de ações
diretas como a produção de fitohormônios, fixação do nitrogênio atmosférico,
solubilização de fosfatos minerais ou outros nutrientes do solo, oxidação do enxofre,
aumento da permeabilidade das raízes e produção de sideróforos (TAN & ZOU, 2001).
A associação entre bactérias diazotróficas e a cana-de-açúcar envolve diversos
gêneros bacterianos e mecanismos singulares ainda pouco compreendidos (JAMES,
2000). Entre as bactérias associadas à cana-de-açúcar destacam-se as espécies
Gluconacetobacter diazotrophicus (CAVALCANTE & DÖBEREINER, 1988),
Azoarcus spp. (REINHOLD-HUREK et al., 1993), Herbaspirillum seropedicae
(BALDANI et al., 1986), Herbaspirillum rubrisubalbicans (GILLIS et al., 1991;
BALDANI et al., 1996) e Burkholderia spp. (YABUUCHI et al., 1992; BALDANI et
al., 1997b).
Pesquisas com bactérias fixadoras de nitrogênio em gramíneas foram iniciadas
pelos pesquisadores Johanna Döbereiner e Alaíde Ruschel na década de 50, onde
isolaram a bactéria fixadora de nitrogênio Beijerinckia fluminensis da rizosfera da canade-açúcar (DÖBEREINER & RUSCHEL, 1958). Nas décadas subseqüentes até hoje
foram feitas grandes descobertas na área de fixação biológica em plantas não
leguminosas, tornando o Brasil referência mundial neste assunto (BODDEY et al.,
1991).
A população das bactérias são influenciadas pela condição hídrica e nutricional
da planta, incluindo o nível de nitrogênio aplicado (REIS JUNIOR et al., 2000), tecidos
utilizados para isolamento (raízes, folhas e colmos), idade da planta (FUENTESRAMIREZ et al., 1993) e inoculações realizadas em condições de laboratório ou de
campo (SEVILLA et al., 2001).
A transmissão de bactérias patogênicas (Leifisonia xyli subsp. xyli e
Xanthomonas albilineans) de uma planta para outra dentro do canavial se dá por
ferramentas de corte como facão, máquinas de plantio e colheita, entre outras. Estas
ferramentas ficam impregnadas com o caldo da planta infectada e contaminam plantas
sadias que são cortadas na seqüência. Também podem ser disseminadas por meio de
plantio de mudas (toletes) vindos de canaviais contaminados (MACCHERONI &
8
MATSUOKA, 2006), o mesmo ocorrendo para as bactérias endofíticas diazotróficas
(DONG et al., 1994).
Outro modo de disseminação local é pela cochonilha Saccharococus sacchari
(ASHBOLT & INKERMAN, 1990). Existe a disseminação por esporos de micorrizas
sendo que no seu interior foram encontradas as bactérias Azospirillum, Klebsiella,
Burkholderia, Gluconacetobacter diazotrophicus entre outras (PAULA et al., 1991).
Restos de cultura também são fontes de disseminação (REIS et al., 1994).
Bastian et al. (1998) mostrou que ambas bactérias, Herbaspirillum seropedicae e
Gluconacetobacter diazotrophicus, produzem giberelinas e ácido indol acético (AIA).
Este resultado pode explicar, em parte, os efeitos benéficos destas bactérias dentro da
planta.
Figura 1 - Possíveis locais de infecção e colonização das bactérias diazotróficas nas
raízes.
Fonte: Reinhold-Hurek e Hurek (1998)
Estes microrganismos possuem a habilidade de colonizar os espaços
intercelulares e o interior das células epidérmicas das raízes. Uma vez dentro da planta,
se locomovem sistematicamente e alcançam os tecidos aéreos através do xilema. São
suportadas pelo transporte de nutrientes do apoplasto, os espaços do tecido
aerenquimático garante um fluxo de oxigênio para a bactéria e estão geralmente em
grupos de 3 ou 4 (JAMES et al., 1994; REINHOLD-HUREK & HUREK 1998).
9
Figura 2 - Células do colmo da cana-de-açúcar colonizadas por Gluconacetobacter
diazotrophicus.
Corte de uma planta com 5 meses crescendo em meio pobre em nitrogênio. A. Corte das
células parenquimatosas armazenadoras de sacarose localizadas próximas do córtex. B.
Corte do tecido vascular, elemento traqueídeo (ponto vermelho), rodeado pelas células
do parênquima (pontos amarelos). O circulo branco mostra as bactérias em ambos
cortes (A e B).
Fonte: Fuentes-Ramírez et al. (1999).
2.4.1 Bactérias Fixadoras de Nitrogênio
Gluconacetobacter diazotrophicus está inserida no grupo das bactérias do ácido
acético e foi isolada de raízes e parte aérea de cana-de-açúcar por Cavalcante e
Döbereiner (1988). É uma bactéria microaeróbia, que cresce em pH baixo (5,0 ou
menos), produz ácido acético a partir de etanol, tolera altas concentrações de sacarose
10
(10%) e inibe parcialmente a fixação de NH4, especialmente em altas concentrações de
sacarose (TEIXEIRA et al., 1997; BODDEY et al., 1991). É encontrada principalmente
em plantas como cana-de-açúcar, Pennisetum purpureum (BALDANI et al., 1997a),
Ipomea batatas (PAULA et al., 1991), Coffea arábica (JIMENEZ-SALGADO et al.,
1997), Ananas comosus (TAPIA-HERNÁNDEZ et al., 2000), arroz selvagem
(LOGANATHAN & NAIR, 2003) e em diversas plantas na Índia (MADHAIYAN et
al., 2004).
A ocorrência restrita deste diazotrófico foi confirmada pela sua completa
ausência no solo e em tecidos de plantas daninhas crescidas entre e dentro das fileiras de
experimentos de campo com cana (DÖBEREINER et al., 1988; SANTOS et al., 2006).
Estes resultados foram confirmados usando primers espécie-específico desenhados para
discriminar Gluconacetobacter diazotrophicus de outros diazotrofos (REIS et al., 1995).
A influência de fatores climáticos, principalmente estress hídrico e estações do
ano no processo de fixação biológica de nitrogênio foi demonstrada por Costa e Ruschel
(1981) e Bellone et al. (1996). Eles sugerem que as atividades das bactérias são
irregulares na primavera, relativamente uniforme no verão e fraca no outono. A
população de Herbaspirillum spp. é menos afetada por fatores climáticos, sugerindo que
são mais bem adaptadas a diferentes condições ambientais.
Gluconacetobacter diazotrophicus é muito sensível a condições secas, mas tem
alta tolerância a tratamentos de calor e concentrações de sais no meio de cultura
(TEJERA et al., 2003). O tratamento térmico (50ºC por 2 a 3 horas), usualmente usado
para controle de patógenos causadores de doenças na cana-de-açúcar não a destroem
(ORTEGA et al., 2001).
Hesbaspirillum seropedicae tem sido isolada de milho, sorgo, arroz, cana-deaçúcar (BALDANI et al., 1986), dendê e banana (DÖBEREINER et al., 1994;
FERREIRA et al., 1995; WEBER, 2003), mas nunca foi isolada de folhas de cana-deaçúcar, apesar de isolada de raízes e colmos (BODDEY et al., 2003).
A disseminação natural de Herbaspirillum seropedicae ainda não está clara,
podendo ocorrer através da propagação vegetativa ou por sementes. A disseminação por
propagação vegetativa foi confirmada pela presença da bactéria em plantas de cana-deaçúcar originadas por processos de micropropagação, nos quais o meristema apical não
foi cuidadosamente extraído (OLIVARES et al., 1996; STURZ, 1995). Por outro lado,
existe a evidência de que sua disseminação pode ocorrer através de sementes
(BALDANI & DOBERËINER, 1995) e de solo (McINROY & KLOEPER, 1995).
11
A bactéria não sobrevive bem no solo natural como outros endófitos, sendo sua
sobrevivência menos afetada em solo estéril, indicando que fatores biológicos
interferem na sobrevivência. Todavia, a taxa de sobrevivência de Herbaspirillum
seropedicae em ambos os solos foi mais alta do que a observada para
Gluconacetobacter diazotrophicus (CARUSO & BALDANI, 1995).
A outra espécie do gênero, Herbaspirillum rubrisubalbicans foi derivada da
reclassificação
de
Pseudomonas
rubrisubalbicans,
considerada
um
agente
fitopatogênico causador da doença conhecida como estria mosqueada em algumas
variedades susceptíveis de cana-de-açúcar cultivadas no Brasil (PIMENTEL et al.,
1991). Estudos de caracterização ecológica demonstraram a ocorrência de
Herbaspirillum rubrisubalbicans em raízes, caules e folhas de cana-de-açúcar em todas
as partes do mundo, sendo também encontrada em arroz e palmeira (FERREIRA et al.,
1995; BALDANI et al., 1997a; OLIVARES et al., 1996).
A população de Herbaspirillum spp. mantém-se estável ao longo do ciclo da
cultura ao contrário da Gluconacetobacter diazotrophicus que decresce ao final do ciclo
(REIS JUNIOR et al., 2000; MUÑOZ-ROJAS & CABALLERO-MELLADO, 2003).
2.4.2 Interação Variedades e Fixação Biológica de Nitrogênio
Existe atualmente quatro programas de melhoramento de cana-de-açúcar no
Brasil. As variedades plantadas comercialmente apresentam diferentes características
quanto a adaptabilidade às regiões de cultivo. A cana-de-açúcar que se tem hoje em dia
é o resultado da hibridação de várias espécies do gênero Saccharum e de outros gêneros.
É um híbrido interespecífico com número variável de cromossomos, entre 100 e 130.
Existem 162 genes ligados ao metabolismo de açúcares em geral, indicando a
complexidade do genoma dessa cultura (FIORAVANTI, 2000).
A manipulação de genes que estão envolvidos no metabolismo do nitrogênio,
carbono e fotossíntese podem melhorar a eficiência do processo de fixação de
nitrogênio, maximizando o potencial da associação planta-bactéria. Nogueira et al.
(2001), estudaram os perfis de expressão gênica de plantas colonizadas pelas bactérias,
usando o banco de dados do SUCEST (Sugarcane Expressed Sequence Tag). Um
catálogo com os genes de cana que são candidatos a se expressar exclusivamente ou
preferencialmente durante a associação foi gerado. Esses dados sugerem que a planta
12
deve ter uma participação ativa na interação, respondendo a diversos processos
metabólicos durante a associação.
Existe uma grande discrepância na estimativa da fixação biológica de nitrogênio
na variedade SP80-1842 entre os Estados de São Paulo e Minas Gerais indicando que o
manejo cultural como a adubação nitrogenada e irrigação podem afetar de forma
expressiva o processo. As variedades RB72454 e SP80-1842 apresentaram elevado
potencial para a fixação biológica de nitrogênio nas lavouras amostradas, no entanto, o
manejo da fertilidade do solo e a nutrição das plantas apresentaram tendência de
influenciar na magnitude da contribuição sendo necessário o monitoramento do estado
nutricional das plantas (POLIDORO et al., 2001).
A fixação biológica contribuiu com 170 kg ha-1 de nitrogênio para a variedade
CB45-3, 150 kg ha-1 para a CB47-89 e 210 kg ha-1 para a SP70-1143, variando entre 60
a 70% do fornecimento de nitrogênio necessário para o desenvolvimento da planta
(URQUIAGA et al., 1992).
A
quantificação
das
bactérias
diazotróficas
Herbaspirillum
spp.
e
Gluconacetobacter diazotrophicus em diversos genótipos mostrou não haver diferença
entre os genótipos e ambas estão presentes em maiores quantidades nas raízes (REIS
JUNIOR et al., 2000). Fisiologia e mudanças metabólicas podem modificar o
estabelecimento e a permanência de Gluconacetobacter diazotrophicus, promovendo o
crescimento da planta quando houve interação variedade e bactéria (MUÑOZ-ROJAS &
CABALLERO-MELLADO, 2003).
A quantificação da fixação biológica de nitrogênio na variedade RB72454 foi
realizada em diversas regiões do Brasil por Polidoro et al. (2001) e Yoneyama et al.
(1997). Devido à metodologia utilizada que avalia a fixação do nitrogênio pelo valor de
abundância natural do isótopo nitrogênio-15, é necessário a utilização de uma planta
testemunha não fixadora. Uma das limitações desta metodologia é exatamente a escolha
da planta testemunha. Na Tabela 2 verifica-se a estimativa de porcentagem de
nitrogênio proveniente da fixação na variedade RB72454 varia conforme o local e a
escolha da testemunha.
13
Tabela 2 - Porcentagem de nitrogênio proveniente da FBN na variedade RB72454 em
diversas regiões produtoras no Brasil
Usina/Município/Estado
Ciclo
Espécie Testemunha
% FBN
São Jose/Macatuba/SP
Planta
Eragrotis pilosa
31,0
Seda rhobifolia
33,0
Emilia sonchifolia
41,2
Panicum maximim
32,9
Acanthopurpreum australe
32,9
Bidens pilosa
33,8
Croton lobatus
45,6
Commelina benghalensis
22,6
Sida rhombifolia
33,4
Euclipta alba
28,2
Nd
26,6
São Jose/Macatuba/SP
UFRRJ/Campos/RJ
Barcelas/Campos/RJ
2º soca
1º soca
nd
Três Pontes/Mg
nd
Cynodon dactylon
49,0
Nova América/Assis/SP
nd
Amaranthus viridis
13,0
Fonte: Adaptado de Polidoro et al. (2001) e Yoneyama et al. (1997).
Nota: nd - Não divulgado.
A eficiência fotossintética, as exigências nutricionais e a resistência às condições
adversas são características ligadas ao genótipo da planta que podem apresentar
influência na eficiência da fixação de N2 pelas bactérias (REIS JUNIOR et al., 2000).
2.4.3 Inoculação de Bactérias Diazotróficas
A Tabela 3 mostra os métodos até agora usados para a aplicação de bactérias
diazotróficas em diversas culturas. A tabela indica que uma grande variedade de
técnicas são usadas, mas pouco tem sido realizado em otimizar o processo de
inoculação. A inoculação sempre foi realizada com um grande número de bactérias
(>108 células ml-1).
14
Tabela 3 - Metodologia de aplicação reportados para algumas bactérias diazotróficas
Bactérias
A. choococcum
Método de Aplicação
Aplicação no solo, sementes ou raízes imersas em suspensão e
aplicação foliar
Azospirillum spp.
Inoculação das sementes antes do plantio, raízes de plântulas
imersas em suspensão de bactérias, aplicação no solo
H. seropedicae
Inoculação em sementes
B. vietnamiensis
Inoculação das sementes antes do plantio, raízes de plântulas
imersas em suspensão de bactérias
Burkholderia sp.
Inoculação das sementes antes do plantio e em plântulas
R. leguminosarum bv. Aplicação da suspensão de bactérias no solo, sementes imersas
Trifolii
na suspensão, sementes revestidas com o inóculo
R. etli bv. Phaseoli
Inoculação em plântulas
A. caulinodans
Inoculação das sementes antes do plantio
G. diazotrophicus
Inoculação em plântulas e colmos de cana-de-açúcar
Fonte: Adaptado de Kennedy et al. (2004)
Trabalhos realizados recentemente em diversos países estudaram a inoculação de
bactérias diazotróficas em plantas micropropagadas de cana-de-açúcar, algumas tendo
respostas positivas em diversos parâmetros (SUMAN et al., 2005) e outras negativas
(OLIVEIRA et al., 2003). Estas plantas estão isentas de diazotróficas, mas não refletem
as condições de plantas coletadas no Brasil (PERIN et al., 2004), Índia
(MUTHUKUMARASAMY et al., 1999), Austrália (BIGGS et al., 2002), África do Sul
(HOEFSLOOT et al., 2005), Japão (YONEYAMA et al., 1997), México (FUENTESRAMÍREZ et al., 1993) e Guatemala (PÉREZ et al., 2003) onde mostram que as plantas
no campo já estão infectadas com algum tipo de bactéria diazotrófica.
Dados mostraram que a inoculação com misturas de bactérias é a melhor
estratégia para melhorar a associação planta-bactéria (OLIVEIRA et al., 2000).
Resultados semelhantes foram demonstrados em 4 variedades de cana na Índia que
foram inoculadas com uma mistura de bactérias diazotróficas e fungos micorrizicos,
produzindo uma resposta equivalente a metade da dose de nitrogênio recomendado para
a cultura (MUTHUKUMARASAMY et al., 1999).
15
A inoculação na variedade SP81-3250 apresentou decréscimo de produtividade
nos três tipos de solos (Ultisol, Oxisol e Alfisol) testados e a resposta à inoculação
ocorreu com maior sucesso em solos de baixa fertilidade (OLIVEIRA et al., 2003).
A contribuição da fixação biológica em plantas de cana (18 meses) inoculadas
em laboratório com uma mistura de estirpes de Gluconacetobacter diazotrophicus foi de
20 a 30% do total de nitrogênio acumulado na planta (OLIVEIRA et al., 2003). Em
outro
trabalho
diferentes
estirpes
foram
inoculadas
na
cana
aumentando
significativamente a brotação e o peso da planta em relação ao controle. A população
ficou mais concentrada nas raízes e foi maior na aplicação de 75 kg ha-1 de nitrogênio
comparado com 0 e 150 kg ha-1, indicando que uma dose inicial é necessária (MORAES
& TORNISIELO, 1997; SUMAN et al., 2005).
O potencial de resposta de cereais à inoculação com estirpes de Herbaspirillum
seropedicae foi avaliado por muitos experimentos nos últimos 15 anos. Resultados
negativos foram obtidos em sorgo, milho e respostas positivas em arroz. A interação de
estirpes de Herbaspirillum seropedicae e Herbaspirillum rubrisulbabicans foram
estudadas em condições de laboratório através da inoculação em cana e arroz. No caso
da cana foi observado que a bactéria entra na planta perto da região meristemática da
raiz, invadindo o tecido vascular e colonizando as células do parênquima e vasos do
xilema. No caso das folhas, formam microcolônias no lúmen do xilema e no apoplasto
(POLIDORO et al., 2001).
2.4.4 Interação Adubação e Fixação Biológica de Nitrogênio
Entre os fatores que podem influenciar o processo da fixação de nitrogênio
destaca-se a presença do nitrogênio mineral. Altas aplicações de fertilizantes
nitrogenados já foram sugeridas como responsáveis pela diminuição do número
populacional de Gluconacetobacter diazotrophicus em variedades de cana-de-açúcar
cultivadas
no
México
(FUENTES-RAMÍREZ
et
al.,
1999),
Índia
(MUTHUKUMARASAMY et al., 1999), Filipinas (GONZALES; BARRAQUIO,
2000) e Brasil (REIS JUNIOR et al., 2000). Também foi observado que íons amônio
inibem o desenvolvimento deste organismo dentro dos tecidos da cana-de-açúcar
(MUTHUKUMARASAMY et al., 2002b). É mais provável que o estado fisiológico da
planta é alterado pelas altas doses de nitrogênio afetando a associação com as bactérias
(FUENTES-RAMÍREZ et al., 1999).
16
Caballero-Mellado e Martinez-Romero (1994) e Caballero-Mellado et al. (1995),
consideram que a fertilização química pode limitar a diversidade genética desta
população, pois no México a diversidade genética é menor do que no Brasil
provavelmente pelo maior requerimento de nitrogênio pelas variedades mexicanas.
Todavia, em experimentos de campo ou em casa de vegetação foram obtidas
respostas positivas a inoculação, mesmo quando foram utilizados altos níveis de
fertilizantes nitrogenados (REYNDERS & VLASSAK, 1982; MILLET & FELDMAN,
1984).
A bactéria Herbaspirillum spp. foi considerada mais resistente a doses maiores
de
nitrogênio
(MUTHUKUMARASAMY
et
al.,
1999).
O
número
de
Gluconacetobacter diazotrophicus é mínimo comparado com outras bactérias em altas
aplicações de nitrogênio (MUTHUKUMARASAMY et al., 2002a; PERIN et al., 2004).
Diferentes níveis de nitrogênio em meio de cultura indicaram que o nitrato e cloreto de
amônio reduziram a colonização de Gluconacetobacter diazotrophicus em relação ao
nitrato de potássio e uréia em plantas micropropagadas. Resultado similar foi obtido em
experimento
de
campo
com
grandes
quantidades
de
nitrogênio
aplicado
(MUTHUKUMARASAMY et al., 2002a).
Na Índia Gluconacetobacter diazotrophicus foi encontrada em canas que não
foram expostas ao nitrogênio industrial, mas somente em aqueles cultivados com
matéria orgânica (MUTHUKUMARASAMY et al., 1999).
Dentre os micronutrientes a limitação na nutrição molíbdica pode ser a mais
importante pelo seu papel nos processos envolvidos na nutrição nitrogenada das plantas
de cana-de-açúcar (POLIDORO et al., 2001). O molibdênio faz parte tanto da redutase
do nitrato, pela qual o nitrato (NO3-) é reduzido a nitrito (NO2-), quanto da nitrogenase,
pela qual bactérias que fixam nitrogênio convertem o gás dinitrogenio (N2) em amônia
(NH3) (EPSTEIN & BLOOM, 2006).
17
3 MATERIAL E MÉTODOS
3.1 Quantificação Prévia das Bactérias Diazotróficas em 6 Variedades de Cana
Para a escolha das variedades do experimento foi realizada uma análise
preliminar para verificar a existência de colonização por parte das bactérias
diazotróficas, em diferentes variedades de cana. Foram estudadas as seguintes
variedades: IACSP93-6006; IAC87-3396; IACSP94-4004; IACSP92-1099; RB72454;
SP81-3250. O teste consistiu em colocar pequenos pedaços de colmos no meio sólido
LGI-P e JNFb para Gluconacetobacter diazotrophicus e Herbaspirillum spp.,
respectivamente, com 10 repetições (DÖBEREINER et al., 1995), quantificando o
crescimento bacteriano (%) após uma semana e procedendo a escolha das duas
variedades utilizadas no experimento.
Na figura 3 visualiza-se a diferença entre a coloração dos meios de cultura
quando na presença das bactérias fixadoras de nitrogênio. Com este método tem-se um
“screening” para trabalhos exploratórios da presença da colonização de bactérias
fixadoras em cana.
Figura 3 - Colmos de cana mergulhados em meio sólido. A coloração azul indica o
crescimento de Herbaspirillum spp. e a coloração amarela indica crescimento de
Gluconacetobacter diazotrophicus.
18
3.2 Local do Experimento
O experimento foi conduzido na APTA Pólo Regional do Centro-Sul,
Piracicaba-SP, em Argissolo Vermelho-Amarelo distrófico com características químicas
apresentadas na Tabela 4 e 5, cultivado há muitos anos com cana-de-açúcar. A análise
do solo foi feita pelo Laboratório de Solos da UFSCar e os métodos de extração e
determinação seguem o utilizado pelo IAC (RAIJ et al., 2001).
A localização geográfica do experimento é dada por 22º41’00” de latitude sul e
47º39’00 de latitude oeste, 554 metros acima do nível do mar. O clima segundo a
classificação de Köppen é do tipo Cwa: subtropical com inverno seco, temperatura
média do mês mais frio inferior a 18ºC e dos mês mais quente superior a 22ºC
(SETZER, 1966). Os parâmetros climáticos apresentam as seguintes médias anuais: (a)
precipitação: 1253 mm; (b) temperatura: 21,1ºC; (c) umidade relativa do ar: 74%; (d)
velocidade do vento: 2,2 m s-1.
Tabela 4 - Características químicas (macronutrientes) do solo onde foi instalado o
experimento*
Prof.
P
S
M.O.
pH
K Ca Mg H+Al
Al
SB
CTC V
Resina
-----mg/dm3--- g/dm3 CaCl2
cm
-------------------mmolc/dm3-------------
%
0-20
5
22
24
5,0
3,2 17
9
50
15,2 29,2 79,2 37
20-40
3
22
20
4,5
2,4 12
7
68
21,5 21,4 89,4 24
*Análise realizada no Laboratório de Solos da UFSCar-Araras.
Tabela 5 – Características químicas (micronutrientes) do solo onde foi instalado o
experimento
Prof.
B
Cu
Fe
Mn
Zn
3
cm
-------------------------------mg/dm ---------------------------
0-20
0,4
1,2
73
28
1,3
20-40
0,27
1,1
71
23
1,1
19
3.3 Tratamentos e o Delineamento Experimental
O experimento foi instalado no campo em Março de 2006 em delineamento
experimental de blocos ao acaso com 4 repetições. As parcelas foram constituídas de 5
linhas de 12 metros espaçadas de 1,4 metros, plantadas com os genótipos de cana-deaçúcar RB72454 e IACSP93-6006 (características na Tabela 6). Como fonte de
nitrogênio, utilizou-se a torta de filtro e uréia na dose de 30 kg ha-1. As doses de fósforo
e potássio foram comuns a todos os tratamentos sendo 140 kg ha-1 de P2O5 na forma de
superfosfato simples e 120 kg ha-1 de K2O como cloreto de potássio. A adubação foi
realizada no sulco, antecedendo o plantio e não foi realizada adubação de cobertura.Os
tratamentos estudados estão descritos na Tabela 7.
Tabela 6 - Características varietal de RB72454 e IACSP93-6006
Características da Planta
RB72454
IACSP93-6006
Brotação
Boa
Boa
Perfilhamento
Bom
Médio
Porte
Ereto
Ereto
Produção Agrícola
Alta
Alta
Brotação das socas
Muito Boa
Boa
Florescimento
Ocasional
Ausência
Média
Semi-precoce
Alto
Bom
Intermediária
Resistente
Resistente
Resistente
Sem Restrições
Baixa exigência
Maturação
Teor de açúcar
Carvão
Ferrugem
Solo
Fonte: Câmara, 1993; Landell et al. 2004
Nota: A variedade RB72454 foi liberada com excelentes resultados de produção agroindustrial em todos
os estados produtores de cana-de-açúcar do Brasil. A variedade IACSP93-6006 foi selecionada na região
de Assis-SP, apresenta alta produtividade agrícola em ambiente favorável e média em ambiente
desfavorável.
20
Tabela 7 - Tratamentos estudados no experimento
Tratamento
Variedade
Inoculação
Sim
Fonte N
Não
T1
RB72454
X
T2
RB72454
X
T3
RB72454
X
T4
RB72454
X
T5
RB72454
X
T6
RB72454
X
T7
IACSP93-6006
X
T8
IACSP93-6006
X
T9
IACSP93-6006
X
T10
IACSP93-6006
X
T11
IACSP93-6006
X
T12
IACSP93-6006
X
Ausência
Orgânico
Mineral
X
X
X
X
X
X
X
X
X
X
X
X
3.4 Preparo do Inóculo e a Inoculação
Para a multiplicação da Gluconacetobacter diazotrophicus estirpe BR 11281 e
posterior inoculação em campo, realizou-se da seguinte maneira: cotas de 20 ml foram
colocados em frascos contendo 180 ml do meio LGI-P e após o sétimo dia (período em
que ocorre o pico de multiplicação das bactérias) (DÖBEREINER et al., 1995) o
material foi retirado dos frascos e acondicionados em garrafas PET de dois litros
esterilizadas (figura 4).
Para a inoculação em campo fez-se uma diluição de 10% com água
(concentração final de 107 células viáveis mL-1) aplicando com um pulverizador costal
no momento do corte dos toletes no sulco de plantio, conforme ilustra as figuras 5 e 6.
21
Figura 4 – Garrafa PET 2 litros com Gluconacetobacter diazotrophicus pronto para ser
colocado no pulverizador costal
Figura 5 – Pulverização da bactéria no sulco de plantio
22
Figura 6 – Detalhe da inoculação. A pulverização é dirigida ao corte recém feito no
colmo pois a entrada da bactéria é facilitada em ferimentos
3.5 Análise Foliar
Foi obtido o terço médio das folhas sem a nervura central da 1ª folha totalmente
expandida que apresenta lígula visível (folha +1-First Visible Dewlep), sendo 20 folhas
coletadas em cada parcela (MALAVOLTA, 1982). As coletas foram realizadas no 8º
mês de cultivo. As amostras foram secas em estufa, moídas e enviadas para análise no
Laboratório de Solos e Análise Foliar do Instituto Agronômico de Campinas, localizado
em Campinas, SP.
3.6 Quantificação das Bactérias Diazotróficas Endofíticas
Para quantificação das bactérias foram coletados cinco colmos/parcela aos 16
meses após a instalação dos tratamentos. As gemas foram plantadas em bandejas
contendo areia esterilizada e aos trinta dias após o plantio foi coletada a parte aérea, que
foi lavada em água corrente, seca em papel toalha e esterilizada superficialmente através
do uso de algodão embebido em álcool. Parte aérea na quantidade de 10 gramas foram
trituradas em liquidificador com 90ml de solução salina por dois minutos. Em seguida,
23
foram feitas as diluições seriadas das amostras em solução salina (10-1 a 10-7) (REIS et
al., 1999).
A contagem em meio semi-sólido foi feita através da inoculação de 0,1 ml de
cada diluição no centro do frasco (Figuras 7, 8 e 9) contendo meio LGI-P e JNFb para
Gluconacetobacter diazotrophicus e Herbaspirillum spp., respectivamente. Foram
realizadas três repetições de cada diluição, sendo o método usado para quantificação o
Número Mais Provável (NMP), que é baseado na presença ou ausência de película
formada no meio semi-sólido, conforme apresentado na Tabela de McCrady
(DÖBEREINER et al., 1995).
Figura 7 – Metodologia do Número Mais Provável, utilizando frascos de penicilina de
10 ml com meio semi-sólido.
Figura 8 – Meio de cultura semi-sólido LGI-P. Os dois frascos da direita estão
positivos quanto ao crescimento de Gluconacetobacter diazotrophicus
24
Figura 9 - Meio de cultura semi-sólido JNFb. Os dois frascos da direita estão positivos
quanto ao crescimento de Herbaspirillum spp.
3.7 Produtividade e Análise Tecnológica
A colheita foi realizada em Agosto de 2007. Para a medição da produtividade as
parcelas foram cortadas, despontadas, despalhadas e tiveram a massa de colmos
avaliada através de pesagens pelo dinamômetro.
Para a análise tecnológica foram coletadas 10 colmos/parcela ao acaso aos 18
meses. Foram analisadas pol% cana, pol ha-1, fibra, pureza, açucares redutores e açúcar
total recuperável.
3.8 Análise Estatística
Os
dados
obtidos
foram
analisados,
estatisticamente,
pelo
programa
STATISTICA 6.0, realizando análise de variância seguida da comparação das médias
pelo teste de Tukey. Para a produtividade foram aplicados Contrastes Ortogonais e o
teste de Dunnett a 5% de significância.
25
4 RESULTADOS E DISCUSSÃO
4.1 Quantificação Prévia das Bactérias Diazotróficas
Na Tabela 8 estão contidos os resultados de percentagem de amostras com
presença de bactérias fixadoras em cada variedade de cana estudada. Verifica-se que
todas as variedades tiveram a presença de bactérias fixadoras em seu colmo. A
variedade IACSP93-6006 foi a escolhida por apresentar alta taxa de colonização e por
ser a variedade IACSP mais plantada. A variedade RB72454 embora não apresentando
os maiores índices de colonização foi escolhida para o experimento no campo, por já ter
sido estudada por outros autores em diversos experimentos relacionados a fixação
biológica de nitrogênio e também por ser a variedade encontrada em maiores áreas de
plantio comercial no Brasil.
Tabela 8 - Quantificação prévia das variedades disponíveis na época de plantio (% de
amostras com presença das bactérias)
Variedades
Gluconacetobacter diazotrophicus
-------------------%------------------
IACSP93-6006
100
IAC87-3396
44
IACSP94-4004
100
IACSP92-1099
100
RB72454
77
SP81-3250
55
4.2 Produtividade
A produtividade de colmos de cana-de-açúcar obtida pelos diversos tratamentos
encontra-se na Tabela 9. Verifica-se que as variedades responderam diferentemente aos
tratamentos aplicados. A variedade RB72454 teve produtividade favorecida tanto pela
presença de nitrogênio como pela inoculação com as bactérias fixadoras ao nível de 5%
de significância. Já a variedade IACSP93-6006 teve diferenças entre os tratamentos
26
somente com 15% de significância. Aplicando esta significância e o teste de Tukey
podemos observar que a produtividade foi favorecida pela presença do nitrogênio
mineral em presença da inoculação. Na média, a variedade IACSP93-6006 foi 9,97%
menos produtiva que a variedade RB72454.
Tabela 9 - Produtividade expressa em toneladas de colmos por hectare
Tratamento
RB72454
IACSP93-6006
---------------------- t ha-1--------------------Inoculado + N-Mineral
143,82ab
148,30a
Inoculado + N-Orgânico
160,82a
136,12ab
Inoculado + ausência N
155,38ab
130,07ab
Não Inoculado + N-Mineral
149,88ab
137,64ab
Não Inoculado + N-Orgânico
166,30a
144,87ab
Não Inoculado + ausência N
137,57b
127,84b
Média
152,69
137,47
F
4,58
2,15
CV (%)
6,55
7,98
5
15
Desvio Padrão
10,00
10,97
D.M.S
15,07
12,00
Nível de Significância (%)
Obs: Médias seguidas de mesma letra nas colunas não diferem entre si pelo teste de Tukey.
Respostas positivas da inoculação com bactérias fixadoras de nitrogênio foram
relatadas na literatura, a exemplo de Shankariah e Hunsigi (2001) que obtiveram
aumento de 9 toneladas ha-1 quando inocularam Azospirillum brasiliense e de 5
toneladas ha-1 quando inocularam Gluconacetobacter diazotrophicus. Moraes e
Tornisielo (1997) obtiveram aumento de produtividade nas plantas inoculadas
cultivadas em solos onde os níveis de fertilidade foram mais elevados.
Por outro lado, a falta de resposta à inoculação também tem sido relatada, a
exemplo de Reis et al. (2000) que comentam que em muitos casos a ausência de
resposta à inoculação pode ser atribuída ao uso de linhagens inadequadas no inóculo.
Este fato não deve ter ocorrido uma vez que o inóculo promoveu aumentos de
produtividade para a variedade RB72454 e IACSP93-6006. Os resultados deste
27
experimento evidenciam um consenso existente na literatura onde a interação genética
entre a variedade e a bactéria, mostra-se como fator chave para a obtenção dos
benefícios provindos da fixação do N2.
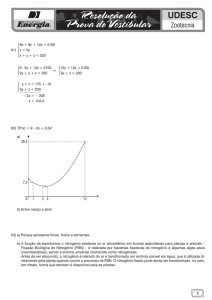
A Tabela 10 indica os diversos contrastes realizados para a produtividade. A
adubação orgânica aparentemente promoveu maiores acréscimos na produtividade para
a RB72454 em relação a testemunha e a adubação mineral promoveu maiores
acréscimos para a IACSP93-6006 em relação a testemunha, independente da
inoculação, conforme pode ser visualizado na figura 10. A torta de filtro, além do
nitrogênio apresenta todas as vantagens da adição de uma fonte de matéria orgânica,
entre elas, a presença de micronutrientes, água, ácidos húmicos e fúlvicos, fatores que
podem incentivar a brotação da cana e a produtividade.
Tabela 10-Contrastes ortogonais dos dados relativos a produtividade das duas
variedades estudadas
Contraste Ortogonal
RB72454
IACSP936006
F**
P***
0,64
F
P
Inoculados x Não inoculados
0,22
0,09
0,75
Testemunha* x Adubação Orgânica
15,15 0,0004 3,59
0,06
Testemunha x Adubação Mineral
1,93
0,17
5,16
0,02
Testemunha x Restante dos tratamentos
8,75
0,005
3,76
0,06
Adubação Orgânica x Mineral independente da
-----
0,07
3,39
----
variedade
* testemunha = ausência de adubação nitrogenada e ausência de inoculação. **F = Teste F. ***P = nível
de significância.
Diversos autores mostraram haver pouca resposta da cana-planta em relação a
adubação nitrogenada (ROSSIELLO, 1987; CANTARELLA & RAIJ, 1986). Esta falta
de resposta pode estar ligada a diversos fatores inerentes ou não a planta como
condições climáticas (COSTA & RUSCHEL, 1981; BELLONE et al., 1996), fertilidade
do solo (OLIVEIRA et al., 2003), calagem, adubação, perdas de nitrogênio por
lixiviação entre outros (TRIVELIN, 2000).
28
Existe diferença também entre a média da testemunha e o restante dos
tratamentos na variedade RB72454 mostrando que quaisquer uns dos tratamentos
tiveram médias melhores do que a da testemunha.
Foi objetivo deste trabalho verificar se a adubação nitrogenada mineral poderia
interferir negativamente na fixação, e compará-la com a adubação orgânica. Os
resultados indicam que o nitrogênio mineral não interfere na fixação ao menos no
parâmetro produtividade da cultura na dose de 30 kg ha-1.
165
colmos t ha -1
160
155
150
145
140
135
Nitrogênio
Orgânico
Nitrogênio
Mineral
Sem Nitrogênio
Figura 10- Produtividade entre os tratamentos adubados com nitrogênio orgânico,
nitrogênio mineral e testemunha sem nitrogênio, independente da inoculação.
Aplicando o Teste de Dunnett para a variedade IACSP93-6006, conforme
Tabela 11, somente o tratamento inoculação+adubação mineral diferiu do controle
(testemunha) ao nível de 5% de significância.
Tabela 11 - Aplicação do Teste de Dunnett para a produtividade
Tratamento
RB72454
IACSP93-6006
----------Nível de significância (%)--------Inoculado + N-Mineral
47,49
3,42
Inoculado + N-Orgânico
1,00
39,39
Inoculado + ausência N
4,37
73,48
Não Inoculado + N-Mineral
16,29
31,39
Não Inoculado + N-Orgânico
2,16
7,75
Não Inoculado + ausência N
Testemunha
29
Para a variedade RB72454 os tratamentos que diferenciaram da testemunha
foram não inoculado+nitrogênio orgânico, inoculado+nitrogênio orgânico e inoculado+
ausência de nitrogênio.
4.3 Análise Tecnológica
As análises tecnológicas realizadas na época da colheita não indicaram
diferenças entre os tratamentos nos parâmetros Pol, Pol ha-1, fibra, pureza, açúcares
redutores e açúcares totais recuperáveis, em nenhuma das variedades estudadas, como
se verifica nas Tabelas 12 e 13. A inoculação com bactérias fixadoras de nitrogênio não
interferiu no acúmulo de sacarose e nos açúcares totais produzidos pela cana. A
adubação orgânica ou mineral também não alterou nenhum dos parâmetros de qualidade
tecnológica.
Tabela 12 – Parâmetros tecnológicos medidos aos 18 meses após a instalação do
experimento, variedade RB72454
Tratamento
Pol
Pol ha-1
Fibra
Pureza
AR
ATR
---------------------------%--------------------------
kg t-1
Inoculado + N-Mineral
14,42
20,90
17,97
92,08
0,21
135,46
Inoculado + N-Orgânico
14,32
22,99
17,58
88,83
0,46
136,78
Inoculado + ausência N
14,66
22,72
17,76
91,99
0,23
137,90
Não Inoculado + N-Mineral
15,34
22,86
14,38
88,49
0,53
146,80
Não Inoculado + N-Orgânico
15,23
25,29
14,52
89,07
0,48
145,37
Não Inoculado + ausência N
15,01
20,69
13,55
89,37
0,48
143,39
Média
14,83
22,57
15,96
89,97
0,39
140,95
ns
ns
ns
ns
ns
ns
14,3
15,41
48,75
2,63
49,98
13,88
5
5
5
5
5
5
Desvio Padrão
2,12
3,47
7,78
2,37
0,19
19,56
D.M.S
3,19
5,24
11,72
3,57
0,29
29,48
F
CV (%)
Nível de Significância (%)
ns - Não significativo aplicando o teste de Tukey.
30
Tabela 13 – Parâmetros tecnológicos medidos aos 18 meses após a instalação do
experimento, variedade IACSP93-6006
Tratamento
Pol
Pol ha-1
Fibra
Pureza
AR
ATR
---------------------------%--------------------------
kg t-1
Inoculado + N-Mineral
15,80
23,16
11,92
89,78
0,44 150,30
Inoculado + N-Orgânico
15,51
21,21
16,69
91,60
0,24 145,83
Inoculado + ausência N
13,59
17,78
21,03
90,50
0,33 128,85
Não Inoculado + N-Mineral
15,50
21,38
12,63
88,97
0,50 148,06
Não Inoculado + N-Orgânico
14,40
20,80
16,28
88,42
0,51 137,99
Não Inoculado + ausência N
15,15
19,40
13,70
89,38
0,46 144,44
Média
14,99
20,62
15,37
89,77
0,41 142,57
ns
ns
ns
ns
17,31
19,21
52,99
2,61
5
5
5
5
5
5
Desvio Padrão
2,60
3,96
8,16
2,34
0,19
23,86
D.M.S
3,91
5,97
12,29
3,53
0,28
35,95
F
CV (%)
Nível de Significância (%)
ns
ns
45,62 16,73
ns - Não significativo aplicando o teste de Tukey.
4.4 Quantificação das Bactérias Fixadoras de Nitrogênio
A Tabela 14 mostra a contagem de bactérias em toletes pré-germinados de cana
aos 16 meses após a aplicação dos tratamentos. Verifica-se que a contagem de bactérias
foi estatisticamente diferente na variedade RB72454 ao nível de 10 e 5% de
significância
para
Gluconacetobacter
diazotrophicus
e
Herbaspirillum
spp,
respectivamente. Entretanto, o tratamento com maior contagem de bactérias não
corresponde exatamente ao tratamento com maior ganho de produtividade, indicando
que apenas a contagem de bactérias não reflete o potencial de fixação da associação.
Njoloma et al. (2006), trabalhou com a inoculação de Herbaspirillum sp.
B501gfp1 em duas concentrações 102 e 108 células ml-1 obtendo como resultado o
aumento do número de células no interior da planta, porém com ausência de resposta no
peso de matéria seca da parte aérea da cana.
31
Tabela 14 - Bactérias diazotróficas (logaritmo do número de células por grama de
matéria fresca) em duas variedades de cana-de-açúcar aos 16 meses.
Tratamento
RB72454
IACSP93-6006
Glucona Herbaspirillum Glucona
Herbaspirillum
Inoculado + N-Mineral
8,24a
8,12a
6,15
6,55
Inoculado + N-Orgânico
5,72b
5,39b
7,31
7,25
Inoculado + ausência N
6,03ab
5,25b
6,5
5,34
Não Inoculado + N-Mineral
6,59ab
6,55ab
7,12
6,79
Não Inoculado + N-Orgânico
7,34ab
7,24ab
5,83
6,03
Não Inoculado + ausência N
5,7b
5,82ab
7,11
6,83
Média
6,59
6,39
6,67
6,46
F
2,86
3,77
ns
ns
CV (%)
18,14
18,18
18,79
36,62
10
5
5
5
Desvio Padrão
1,19
1,16
1,25
2,37
D.M.S
1,48
1,75
1,89
3,56
Nível de Significância (%)
Obs: Médias seguidas de mesma letra nas colunas não diferem entre si pelo teste de Tukey.
ns - Não significativo.
O número de bactérias presentes não foi inibido pela presença de nitrogênio
mineral e nem incentivado pela presença de nitrogênio orgânico. Moraes e Tornisielo
(1997) e Suman et al. (2005) indicam que uma dose inicial de nitrogênio é necessária
para o desenvolvimento das bactérias, sem inibi-las. Um menor número de bactérias foi
encontrado no tratamento sem nitrogênio e não inoculado, indicando que a presença de
nitrogênio pode favorecer o estabelecimento da associação entre a planta e as bactérias.
Verifica-se também que a inoculação não aumentou o número de bactérias em
relação aos tratamentos não inoculados. Vale ressaltar que os toletes usados para a
propagação da cana carregam uma população nativa de bactérias diazotróficas
(OLIVARES, 1997; Tabela 7) podendo explicar a falta de resposta dos tratamentos a
quantificação das bactérias, principalmente a aqueles onde houve a inoculação. Também
pode ser explicado pelos diversos fatores que interagem nesta complexa simbiose
bactéria-planta como época do ano e estádio fisiológico da cultura (COSTA &
RUSCHEL, 1981), características ligadas ao genótipo da planta como eficiência
32
fotossintética e resistência as condições adversas (REIS JUINOR et al., 2000;
NOGUEIRA et al., 2001; MUÑOZ-ROJAS & CABALLERO-MELLADO, 2003).
4.5 Análise Foliar
Os tratamentos estudados não aumentaram o teor de nitrogênio nas folhas +1 da
cana-de-açúcar aos 8 meses após a instalação do experimento, conforme se verifica na
Tabela 15, estando os valores dentro da faixa considerada adequada de acordo com
Malavolta (1982). Há controvérsias quanto à técnica da análise foliar para avaliar o
estado nutricional da cana-de-açúcar. Nem sempre a análise da folha diagnóstico,
representa o potencial de absorção do nutriente ao longo do ciclo da cultura, conforme
Orlando Filho et al. (2001).
Tabela 15 – Nitrogênio total na folha +1 aos 8 meses após a instalação do experimento
Tratamento
RB72454
IAC93-6006
-1
-----------g kg ----------Inoculado + N-Mineral
20,30
20,28
Inoculado + N-Orgânico
20,25
20,25
Inoculado + ausência N
20,83
20,20
Não Inoculado + N-Mineral
20,63
20,10
Não Inoculado + N-Orgânico
19,93
20,30
Não Inoculado + ausência N
20,13
20,23
Média
20,34
20,23
ns
ns
4,06
4,07
5
5
Desvio Padrão
0,825
0,776
D.M.S
1,24
1,17
F
CV (%)
Nível de Significância (%)
ns - Não significativo aplicando o teste de Tukey.
33
5 CONCLUSÕES
Os resultados obtidos no presente trabalho permitem concluir que:
1. As variedades estudadas apresentaram-se colonizadas por bactérias fixadoras de
nitrogênio independente da aplicação da fonte de nitrogênio.
2. As
variedades
RB72454
e
IACSP93-6006
apresentaram
ganhos
na
produtividade de colmos quando se procedeu a inoculação no sulco de plantio ou
a adição de fonte nitrogenada.
3. As bactérias fixadoras de nitrogênio não foram inibidas pela presença de
nitrogênio mineral.
4. A qualidade da matéria prima não foi alterada em nenhum dos parâmetros
avaliados, nem pela inoculação e nem pela adição de fonte nitrogenada.
34
6 REFERÊNCIAS BIBLIOGRÁFICAS
ASSOCIAÇÃO NACIONAL PARA DIFUSÃO DE ADUBOS. Anuário Estatístico do
Setor de Fertilizantes. São Paulo: Editora Nagy, 2004. 162p.
ASHBOLT, N.J.; INKERMAN, P.F. Acetic acid bacterial biota of the pink sugarcane
mealbug, Saccharococcus sacchari, and its environs. Applied and Envionmental
Microbiology, 56, p.707-712, 1990.
AZEREDO, D.F.; BOLSANELLO, J.; WEBER, H.; VIEIRA, J.R. Nitrogênio na cana
planta: doses e fracionamento. Revista STAB, Piracicaba, v.4, p.32-36, 1986.
BALDANI, J.I.; DOBEREINER, J. Recent advances in BNF with non-legumes plants.
In: INTERNATIONAL SYMPOSIUM ON SUSTAINABLE AGRICULTURE FOR
THE TROPICS – THE ROLE OF BIOLOGICAL NITROGEN FIXATION, 1995,
Angra dos Reis. Abstracts... Angra dos Reis: The National Center for Agrobiology
Research (Embrapa-CNPAB), 1995. p.53-54.
BALDANI, J.I.; BALDANI, V.L.D.; SELDIN, L.; DÖBEREINER, J. Characterization
of Herbaspirillum seropedicae gen. nov., sp. nov., a root associated nitrogen-fixing.
International Journal of Systematic Bacteriology, Baltimore, v.36, p.86-93, 1986.
BALDANI, J.I.; CARUSO, L.; BALDANI, V.L.D.; GOI, R.S; DÖBEREINER, J.
Recent advances in BNF with non-legumes plants. Soil Biology Biochemistry, Oxford,
v.29, p.922-928, 1997a.
BALDANI, V.L.D.; REIS, V.M.; BALDANI, J.I.; KIMURA, O.; DÖBEREINER, J.
Bactérias fitopatogênicas fixadoras de N2 em associação com plantas. Seropédica:
Embrapa-CNPAB, 1997b. 25p.
35
BALDANI, J.I.; PLOT, B.; KIRCHOF, G.; FALSEN, E.; BALDANI, V.L.D.;
OLIVARES, F.L.; HOSTE B.; KERSTERS, K.; HARTMANN, A.; GILLIS, M.;
DÖBEREINER, J. Emended description of Herbaspirillum; inclusion of (Pseudomonas)
rubrisubalbicans, a mid plant pathogen, as Herbaspirillum rubrisulbabicans comb.
nov., and classification of a group of clinical isolates (EF group 1) as Herbaspirillum
species 3. International Journal Systematic Bacteriology, v. 46, p.802-810, 1996.
BALDANI, J.I.; CARUSO, L.; BALDANI, V.L.D.; GOI, R.S; DÖBEREINER, J.
Recent advances in BNF with non-legumes plants. Soil Biology Biochemistry, Oxford,
v.29, p.922-928, 1997a.
BASTIAN, F.; COHEN, A.; PICCOLI, P.; LUNA, V.; BARALDI, R.; BOTTINI, R.
Production of indole-3-acetic and gibberelins A1 and A3 by Acetobacter diazotrophicus
and Herbaspirilum seropedicae in chemically-defined culture media. Plant Growth
Regulation, 24, p. 7-11, 1998.
BELLONE, C.H.; BELLONE, S.C.; PEDRAZA, R.O. Hydric deficiency and acetylene
reduction in sugarcane roots. In: MALIK, K.A.; MIRZA, M.S.; KHALID, A.M. (Ed.).
7th Int. Symp. Nitrogwen Fixation with Non-Legumes. Faisalabad, Pakistan, 1996. p.
125-126.
BIGGS, I.M.; STEWART, G.R.; WILSON, J.R.; CRICHLEY, C. 15N natural
abundance studies in Australian commercial sugarcane. Plant and Soil, v. 238, p. 2130, 2002.
BODDEY, R.M.; URQUIAGA, S.; ALVES, B.J.R.; REIS, V.M. Endophytic nitrogen
fixation in sugarcane: present knowledge and future applications. Plant and Soil, v.
252, p. 139-149, 2003.
BODDEY, R. M; URQUIAGA, S.; DÖBEREINER, J.; REIS, J. Biological nitrogen
fixation associated with sugar cane. Plant and Soil, Dordrecht, n. 137, p. 111-117,
1991.
36
CABALLERO-MELLADO, J.; MARTÍNEZ-ROMERO, E. Limited genetic diversity in
the endophytic sugarcane bacterium Acetobacter diazotrophicus. Applied and
Environmental Microbiology, Washington, v.60, p.1532-1537, 1994.
CABALLERO-MELLADO, J.; FUENTES-RAMÍREZ, L.E.; REIS, V.M.;
MARTÍNEZ-ROMERO, E. Genetic structure of Acetobacter diazotrophicus
populations and identification of a new genetically distant group. Applied and
Environmental Microbiology, Washington, v.61, p.3008-3013, 1995.
CÂMARA, G. M. de S. Ecofisiologia da cultura da cana-de-açúcar. In: _______. (Ed.).
Produção de cana-de-açúcar. Piracicaba: Fealq, 1993. p.31-60.
CANTARELLA, H.; RAIJ, B. van. Adubação nitrogenada no Estado de São Paulo. In:
SIMPÓSIO SOBRE ADUBAÇÃO NITROGÊNADA NO BRASIL. Reunião Brasileira
de Fertilidade do Solo,19., 1984, Ilhéus. Resumos... Ilhéus: CEPLAC, 1986. p.47-79.
CARUSO, L.V.; BALDANI, J.I. Monitoring the survival of endophytic diazotrophic
bacteria in soil using LacZ-fusion. In: INTERNATIONAL SYMPOSIUM ON
SUSTAINABLE AGRICULTURE FOR THE TROPICS: THE ROLE OF
BIOLOGICAL NITROGEN FIXATION, 1995, Angra dos Reis. Abstracts... Angra dos
Reis: The National Center for Agrobiology Research (Embrapa-CNPAB), 1995. p.108.
CAVALCANTE, V.A.; DÖBEREINER, J. A new acid-tolerant nitrogen-fixing
bacterium associated with sugarcane. Plant and Soil, Dordrecht, v.108, p.23-31, 1988.
CLEMENTS, H.F. Sugarcane crop logging and control: principles and practices.
[London]: Pitman Publishing, 1980. 520p.
COPERSUCAR. Uso da torta de filtro no plantio da cana-de-açúcar. Cadernos
Copersucar, série agronômica, n. 001, Piracicaba, 6p, 1986.
COSTA, J.M.T.; RUSCHEL, A.P. Seasonal variation in the microbial populations of
sugarcane plants. In: VOSE, P.B., RUSCHEL, A.P. (Ed.). Associative N2-fixation.
Boca Raton: CRC, v.2, 1981. p. 109-118.
37
DOBBELAERE, S.; VANDERLEYDEN, J.; OKON, Y. Plant growth-promoting
effects of diazotrophs in the rhizosphere. Critical Reviews in Plant Science, v. 22,
p.107-149, 2003.
DÖBEREINER, J. Biological nitrogen fixation in the tropics: social and economic
contributions. In: INTERNATIONAL SYMPOSIUM ON SUSTAINABLE
AGRICULTURE FOR THE TROPICS – THE ROLE OF BIOLOGICAL NITROGEN
FIXATION, 1995, Angra dos Reis. Abstracts... Angra dos Reis: The National Center
for Agrobiology Research (Embrapa-CNPAB), 1995. p.3-4.
DÖBEREINER, J.; RUSCHEL, A.P. Uma nova espécie de Beijerinckia. Revista de
Biologia, v. 1, p. 261-272, 1958.
DÖBEREINER, J.; BALDANI, V.L.D.; BALDANI, J.I. Como isolar e identificar
bactérias diazotróficas de plantas não-leguminosas. Brasília – SPI: Itaguaí, RJ:
EMBRAPA-CNPAB, 1995. 60p.
DÖBEREINER, J.; REIS, V.M.; LAZARINI, A.C. New N2-fixing bacteria in
association with cereals and sugarcane. In: BOTHE, H.; BRUIJIN, F.J. de; NEWTON,
W.E. (Ed.). Nitrogen fixation: hundred years after. Stuttgart: Gustav Fisher, 1988.
p.717- 722.
DÖBEREINER, J.; BALDANI, V.L.D.; OLIVARES, F.L.; REIS, V.M. Endophytic
diazotrophs: The key to graminaceous plants. In: HEGAZI, N.A.; FAYEZ, M.;
MORRIB, M. (Ed.). Nitrogen fixation with non-legumes. Cairo: The American
University, 1994. p.395-408.
DONG, Z.; CANNY, M.J.; McCULLY, M.E.; REBOREDO, M.R.; FERNÁNDEZ, C.;
ORTEGA, E.; RODÉS, R. A nitrogen-fixing endophyte of sugarcane stems. A new role
for the apoplast. Plant Physiology, v.105, p. 1139-1147, 1994.
EPSTEIN, E.; BLOOM, A. J. Nutrição mineral de plantas: princípios e perspectivas.
Londrina: Editora Planta, 2006. 401p.
38
FERNANDES, M.S.; ROSSIELLO, R.O.P. Mineral nitrogen in plant physiology and
plant nutrition. Critical Reviews in Plant Sciences, Boca Raton, v.14, p.111-148,
1995.
FERREIRA, A.C.; COZZOLINO, K.; CARVALHO, A.R.V.; DÖBEREINER, J.
Isolation and characterization of diazotrophic bacteria in oil palm trees. In:
INTERNATIONAL SYMPOSIUM ON SUSTAINABLE AGRICULTURE FOR THE
TROPICS – THE ROLE OF BIOLOGICAL NITROGEN FIXATION, 1995, Angra dos
Reis. Abstracts... Angra dos Reis: The National Center for Agrobiology Research
(Embrapa-CNPAB), 1995. p.210.
FIOVARANTI, C. Os Arquitetos da Nova Cana. Revista Pesquisa Fapesp, São Paulo,
n. 59, p.29-35, 2000.
FNP CONSULTORIA & COMÉRCIO. Cana-de-açúcar. In: _______. Agrianual 2007:
anuário da agricultura brasileira. São Paulo, 2002. p.237-247.
FRANCO, A.A.; BALIEIRO, F.C. Fixação biológica de nitrogênio: alternativas aos
fertilizantes nitrogenados. In: SIQUEIRA, J.O.; MOREIRA, F.M.S; LOPES, A.S.;
GUILERME, L.R.G.; FAQUIM, V.; FURTINI NETO, A.E.; CARVALHO, J.G. (Ed.).
Inter-relação fertilidade, biologia do solo e nutrição de plantas. Viçosa, MG:
Sociedade Brasileira de Ciência do Solo/Lavras, 1999. p. 577-595.
FUENTES-RAMÍREZ, L.E.; CABALLERO-MELLADO, J.; SEPÚLVEDA, J.;
MARTÍNEZ-ROMERO, E. Colonization of sugarcane by Acetobacter diazotrophicus is
inhibited by high N-fertilization. FEMS Microbiology Ecology, Amsterdam, v.29,
p.117-128, 1999.
FUENTES-RAMÍREZ, L.E.; JIMÉNEZ-SALGADO, T.; ABARCA-OCAMPO, I.R.;
CABALLERO-MELLADO, J. Acetobacter diazotrophicus, an indole-acetic acid
producing bacterium isolated from sugar cane cultivars in México. Plant and Soil,
Dordrecht, v.154, p.145-150, 1993.
39
GILLIS, M.; DÖBEREINER, J.; POT, P.; GOOR, M.; FALSEN, E.; HOSTE, B.;
REINHOLD, B.; KERSTERS, K. Taxonomic relationship between (Pseudomonas)
Rubrisubalbicans, some clinical isolates (EF group 1), Herbaspirillum seropedicae and
(Aquaspirillum) Autotrophicum. In: POLSINELLI, M.; MATERASSI, R.; VICENZINI,
M. (Ed.). Nitrogen Fixation. Dordrecht: Kluwer Academic Publishers, 1991, p.293294.
GONZALES, M.S.; BARRAQUIO, W.L. Isolation and characterization of Acetobacter
diazotrophicus in Sacharum officinarum L., Saccharum spontaneum L., and Erianthus
sp. The Philippine Agricultural Scientist, v.83, p.173-181, 2000.
GRAHAM, P.H.; VANCE C.P. Nitrogen fixation in perspective: an overview of
research and extension needs. Field Crops Research, v.65, p.93-106, 2000.
HOEFSLOOT, G.; TERMORSHUIZEN, A.J.; WATT, D.A.; CRAMER, M.D.
Biological nitrogen fixation is not a major contributor to the nitrogen demand of a
commercially grown South Africa sugarcane cultivars. Plant and Soil, v. 227, p. 85-96,
2005.
JAMES, E.K. Nitrogen fixation in endophytic and associative symbiosis. Field Crops
Research, v.65, p.197-209, 2000.
JAMES, E.K.; REIS, V.M.; OLIVARES, F.L; BALDANI, J.I.; DÖBEREINER, J.
Infection of sugarcane by nitrogen fixing bacterium Acetobacter diazotrophicus.
Journal Experimental Botany, v.45, p.757-766, 1994.
JIMENEZ-SALGADO, T.; FUENTES-RAMIREZ, L.E.; MARTINEZ-ROMERO, E.;
CABALLERO-MELLADO, J. Coffea arabica, a new host plant for Acetobacter
diazotrophicus and isolation of others nitrogen fixing acetobacteria. Applied
Environmental Microbiology, v.63, p.3676-3683, 1997.
40
KENNEDY, I.R.; CHOUDHURY, A.T.M.A.; KECSKÉS, M. L. Non-symbiotic
bacterial diazotrophs in crop-farming systems: can their potential for plant growth
promotion be better exploited? Soil Biology and Biochemistry, 36, p. 1229-1244,
2004.
LANDELL, M.G. de A.; CAMPANA, M.P.; FIGUEIREDO, P.; SILVA, M.A.;
VASCONCELOS, A.C.M de; BIDOIA, M.A.P.; XAVIER, M.A.; DINARDOMIRANDA, L.L.; PRADO, H.; ROSSETTO, R.; SANTOS, A.S.; CAVICHIOLI, J.C.;
MARTINS, A.L.M.; KANTHACK, R.A.D.; GALLO, P.B.; VEIGA FILHO, A.A.;
SILVA, D.N. Variedades de cana-de-açúcar para o Centro-Sul do Brasil: 14a
liberação do programa cana IAC (1959-2004). Campinas: Instituto Agronômico, 2004.
33p.
LOGANATHAN, P.; NAIR, S. Crop specific endophytic colonization by a novel, salttolerant, nitrogen fixing and phosphate-solubilizing Gluconacetobacter spp. from wild
rice. Biotechnology Letters, 25, p. 497-501, 2003.
MACCHERONI, W.; MATSUOKA, S. Manejo das principais doenças da cana-deaçúcar. In: SEGATO, S.V.; PINTO, A. de S.; JENDIROBA, E.; NOBREGA, J.C.M. de.
(Ed.) Atualização em produção de cana-de-açúcar. Piracicaba: CP2, 2006. p.239256.
MADHAIYAN, M.; SARAVANAN, V.S.; JOVI, D.B.S.S.; LEE, H.; THENMOZHI,
R.; HARI, K.; SA, T. Occurrence of Gluconacetobacter diazotrophicus in tropical and
subtropical plants of Western Ghats, India. Microbiological Research, v. 159, p. 233243, 2004.
MALAVOLTA, E. Nutrição mineral e adubação da cana-de-açúcar. São Paulo:
Ultrafertil, 1982. 80p.
MALAVOLTA, E.; VITTI, G.C.; OLIVEIRA, S.A. Avaliação do estado nutricional
das plantas. Piracicaba: Associação Brasileira para Pesquisa da Potassa e do Fosfato,
1989. 201p.
41
McINROY, J.A.; KLOEPER, J.W. Survey of indigenous bacterial endophytes from
cotton and sweet corn. Plant Soil, v. 173, p. 337-342, 1995.
MILLET, E.; FELDMAN, M. Yield response of common spring wheat cultivars to
inoculation with Azospirillum brasilense at various levels of nitrogen fertilization. Plant
and Soil, n. 80, p.255-259, 1984.
MORAES, V.A.; TORNISIELO, S.M.T. Aumento na produtividade da variedade SP
70-1143 pela inoculação da bactéria Acetobacter diazptrophicus. In: SEMINÁRIO
COPERSUCAR DE TECNOLOGIA AGRONOMICA, 7., 1997. Anais…Piracicaba:
Editora, 1997. p. 131-139.
MUCHOW, R.C.; ROBERTSON, M.J. Relating crop nitrogen uptake to sugarcane
yield. In: AUSTRALIAN SOCIETY OF SUGARCANE TECHNOLOGISTS, 16, 1994,
Tournsvitte. Proceedings… Tournsvitte: Brisbane, 1994. p. 122-130.
MUÑOZ-ROJAS, J.; CABALLERO-MELLADO. Population dynamics of
Gluconacetobacter diazotrophicus in sugarcane cultivars and its effects in plant growth.
Microbiology Ecology, v.46, p.454-464, 2003.
MUTHUKUMARASAMY, R.; REVATHI, G.; LAKSHMINASIMHAN, C. Influence
of nitrogen fertilization on the isolation of Acetobacter diazotrophicus and
Herbaspirillum spp. from Indian sugarcane varieties. Biology and Fertility Soils,
Berlin, v.29, p.157-164, 1999.
MUTHUKUMARASAMY, R.; REVATHI, G.; LOGANATHAN, P. Effect of
inorganic N on the population, in vitro colonization an morphology of Acetobacter
diazotrophicus (syn. Gluconacetobacter diazotrophicus). Plant and Soil, v. 243, p.91102, 2002a.
MUTHUKUMARASAMY, R.; REVATHI, G.; SESHADRI, S.;
LAKSHMINASIMHAN, C. Gluconacetobacter diazotrophics (syn. Acetobacter
diazotrophics), a promising diazotrophic endophyte in tropics. Current Science, v. 83,
p.137-145, 2002b.
42
NGUYEN, H.T.; DEAKER, R.; KENNEDY, I.R.; ROUGHLEY, R.J. The positive
yield response of field-grown rice to inoculation with a multi-strain biofertiliser in the
Hanoi Area, Vietnam. Symbiosis, v. 35, p. 231-245, 2003.
NJOLOMA, J.; TANAKA, K.; SHIMIZU, T.; NISHIGUCHI, T.; ZAKRIA, M.;
AKASHI, R.; OOTA, M.; AKAO, S. Infection and colonization of aseptically
micropropagated sugarcane seedlings by nitrogen-fixing endophytic bacterium,
Herbaspirillum sp. B501gfp1. Biology Fertility Soils, v.43, p. 137-143, 2006.
NOGUEIRA, E. M.; VINAGRE, F.; MASUDA, H.P.; VARGAS, C.; PADUA, V.L.M.;
SILVA, F.R.; SANTOS, R.V.; BALDANI, J.I.; FERREIRA, P.C.G.; HEMERLY, A.S.
Expression of sugarcane genes induced by inoculation with Gluconacetobacter
diazotrophicus and Herbaspirillum rubrisubalbicans. Genetics and Molecular
Biology, v. 24, p.199-206, 2001.
OLIVARES, F.L. Taxonomia, ecologia e mecanismos envolvidos na infecção e
colonização de cana-de-açúcar (Saccharum sp. híbrido) por bactérias diazotroficas
endofíticas do gênero herbaspirillum. 1997. 328p. Tese (Doutorado). Universidade
Federal Rural do Rio de Janeiro, Rio de Janeiro, 1997.
OLIVARES, F.L.; BALDANI, V.L.D.; REIS, V.M.; BALDANI, J.L.; DÖBEREINER,
J. Occurrence of the endophytic diazotrophs Herbaspirillum spp. in roots, stem and
leaves predominantly of graminaceae. Biology Fertility Soils, v. 2, p.197-200, 1996.
OLIVEIRA, A.L.M.; CANUTO, E.L; REIS, V.M; BALDANI, J.I. Response of
micropropagated sugarcane varieties to inoculation with endophytic diazotrophic
bactéria. Brazilian Journal of Microbiology, São Paulo, v.34, p.59-61, nov. 2003.
OLIVEIRA, A.L.M.; URQUIAGA, S.; DÖBEREINER, J.; BALDANI, J.I. Biological
nitrogen fixation (BFN) in micropropagated sugarcane plants inoculated with different
endophytic diazotrophic bacteria. In: PEDROSA, F.O.; HUNGRIA, M.;YATES, M.G.;
NEWTON, W.E. (Ed). Nitrogen fixation: from molecules to crop productivity. Current
Plant Science and Biotechnology in Agriculture. Netherlands: Kluwer Academic
Publishers, 2000. 425p.
43
ORLANDO FILHO, J.; HAAG, H.P.;ZAMBELLO JUNIOR, E. Crescimento e
absorção de macronutrientes pela cana-de-açúcar variedade CB 41-76 em função
de idade em solos do Estado de São Paulo. Piracicaba: Planalsucar, 1980. 128p.
ORLANDO FILHO, J.; ROSSETTO, R.; CASAGRANDE, A.A. Micronutrientes para a
cana-de-açúcar. In: FERREIRA, M. E. et al. (Ed.). Micronutrientes e elementos
tóxicos na agricultura. São Paulo: Ed. Legis Summa, vol. 1, 2001. p. 355-373.
ORTEGA, E.; RODES, R.; de la FUENTE, E.; FERNÁNDEZ, L. Does the routine
treatment of sugarcane stem pieces for xylem pathogen control affect the nitrogenase
activity and N2-fixing endophyte in the cane. Australian of Plant Physiology, 28,
p.907-912, 2001.
ORTOLANI, M.C. de A. Perspectivas para o setor sucroalcooleiro. In: SEGATO, S.V.;
PINTO, A. de S.; JENDIROBA, E.; NOBREGA, J.C.M. de. (Ed.) Atualização em
produção de cana-de-açúcar. Piracicaba: CP2, 2006 p.406-415.
PAULA, M.A.; REIS, V.M.; DÖBEREINER, J. Interactions of Glomus clarus and
Acetobacter diazotrophicus in infection of sweet potato (Ipomea batatas), sugarcane
(Saccharum spp.), and sweet sorghum (Sorghum vulgare). Biology Fertility Soils, v.11,
p. 111-115, 1991.
PENATTI, C. P. Doses de nitrogênio em áreas de vinhaça durante quatro safras na
Usina Santa Luiza. In: SEMINÁRIO COPERSUCAR DE TECNOLOGIA
AGRONOMICA, 6., 1994, Rio de Janeiro. Anais…Piracicaba: Editora, 1994. p. 105110.
PEOPLES, M.B.; HERRIDGE, D.F. & LADHA, J.K. Biological nitrogen fixation: an
efficient source of nitrogen for sustainable agricultural production. Plant and Soil,
Dordrecht, v.174, p.23-28, 1995.
44
PEREZ, O.; OVALLE, W.; RODAS, A.; LÓPEZ, E.; HERNÁNDEZ, F. Bacterias
fijadoras de nitrógeno en caña de azúcar: Métodos para multiplicación y
comportamiento de variedades promisorias en campo. Guatemala: GENGICAÑA, 2003.
26p.
PERIN, L.; BALDANI, J.I.; REIS, V.M. Diversidade de Gluconacetobacter
diazotrophicus isolada de plantas de cana-de-açúcar cultivadas no Brasil. Pesquisa
Agropecuária Brasileira, v. 39, p.763-770, 2004.
PIMENTEL, J.P.; OLIVARES, F.L.; PITARD, R.M.; URQUIAGA, S.C.; AKIBA, F.;
DÖBEREINER, J. Dinitrogen fixation and infection of grasses leaves by Pseudomonas
rubrisubalbicans and Herbaspirillum seropedicae. Plant and Soil, Dordrecht, v.137,
n.1, p.61-65, 1991.
POLIDORO, J.C. O molibdênio na nutrição nitrogenada e na fixação biológica de
nitrogênio atmosférico associado a cultura da cana-de-açúcar. 2001. 185p. Tese
(Doutorado). Universidade Federal Rural do Rio de Janeiro, Rio de Janeiro, 2001.
POLIDORO, J. C.; RESENDE A. S.; QUESADA, D. M.; XAVIER, R .P.; COELHO,
C. H. M.; ALVES, B. J. R.; BODDEY, R. M.; URQUIAGA, S. Levantamento da
contribuição da fixação biológica de nitrogênio para a cultura da cana-de-açúcar
no Brasil. Seropédica: Embrapa Agrobiologia, dez. 2001. 8p.
RAIJ, B. van; ANDRADE, J.C.; CANTARELLA, H.; GUAGGIO, J.A (Ed.). Análise
química para avaliação da fertilidade de solos tropicais. Campinas: Instituto
Agronômico, 2001. 285p.
REINHOLD-HUREK, B.; HUREK, T. Life in grasses: diazotrophic endophytes.
Trends in Microbiology, v.6, 139-143, 1998.
45
REINHOLD-HUREK, B.; HUREK, T.; GILLIS, M.; HOSTE, B.; VANCANNEYT;
M.; KERSTERS, K.; DE LEY, J. Azoarcus gen. nov., nitrogen-fixing proteobacteria
associated with roots of kallar grass (Leptochloa fusca (L.) Kunth), and description of
two species, Azoarcus indigens sp. nov. and Azoarcus communis sp. nov. International
Journal of Systematic Bacteriology, Baltimore, v.43, n.3, p.574- 584, 1993.
REIS, V.M; OLIVARES, F.L; DÖBEREINER, J. Improved methodology for isolation
and identification of Acetobacter diazotrophicus and confirmation of its entophytic
habitat. World Journal of Microbiology and Biotechnology, Oxford, v.10, p.401-405,
1994.
REIS, V.M.; PERIN, L.; REIS JUNIOR, F.B. Uma nova estratégia para isolar
Gluconacetobacter diazotrophicus de plantas de cana-de-açúcar. Comunicado Técnico
nº 35-Embrapa Agrobiologia, 1999, 5p.
REIS, V.M.; BALDANI, J.I.; BALDANI, V.L.; DÖBEREINER, J. Biological
dinitrogen fixation in gramineae and palm trees. Critical Reviews in Plant Sciences,
v.19, p.227-247, 2000.
REIS, V.M.; BALDANI, J.I.; KIRCHHOF, G.; HARTMANN, A. Development of a
molecular assay to identify Acetobacter diazotrophicus species and detect their
occurrence in sugar cane and other plants grown in the field. In: INTERNATIONAL
SYMPOSIUM ON SUSTAINABLE AGRICULTURE FOR THE TROPICS – THE
ROLE OF BIOLOGICAL NITROGEN FIXATION, 1995, Angra dos Reis. Abstracts...
Angra dos Reis: The National Center for Agrobiology Research (Embrapa-CNPAB),
1995. p.130.
REIS JUNIOR, F.B.; SILVA, L.G.; REIS, V.M.; DÖBEREINER, J. Ocorrência de
bactérias diazotroficas em diferentes genótipos de cana-de-açúcar. Pesquisa
Agropecuária Brasileira, Brasília, v.35, p.985-994, 2000.
REYNDERS, L.; VLASSAK, K. Use of Azospirillum brasiliense as biofertilizer in
intensive wheat cropping. Plant and Soil, Dordrecht, n.66, p. 217-273, 1982.
46
ROSSIELO, R.O.P. Bases fisiológicas na acumulação de nitrogênio e potássio em
cana-de-açúcar (Saccharum spp. cv. NA 56-79) em resposta à adubação
nitrogenada em cambissolo. 1987. 172p. Tese (Doutorado) – Escola Superior de
Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba, 1987.
SANTOS, C.C.R.; PERIN, L.; BALDANI, J.I., REIS, V.M. Isolamento de
Gluconacetobacter spp. em diferentes tipos de solos. Pesquisa Agropecuária
Brasileira, v.41, nº1, p.85-92, 2006.
SETZER, J. Atlas climático e ecológico do Estado de São Paulo. São Paulo:
Comissão Interestadual da Bacia Paraná-Uruguai, 1966. 61p.
SEVILLA, M.; BURRIS, R.H.; GUNAPALA, N.; KENNEDY, C. Comparison of
benefit to sugarcane plant growth and 15N in corporation following inoculation of sterile
plants with Acetobacter diazotrophicus wild-type and Nif mutants strains. Molecular
Plant-Microbe Interactions, 4, p. 358-366, 2001.
SHANKARIAH, C.; HUNSIGI, G. Field response of sugarcane to associative N2 fixers
and P solibilizers. In: HOARTH, D.M. (Ed.) Proceeding of the 24th International
Society of Sugar Cane Technologists Congress, 17-21 September 2001. The
Australian Society of Sugarcane Technologists, Brisbane p.40-45.
SHEARE, G.; JOHL, D.H. N2-fixation in field settings: estimations based on natural
15
N abundance. Australian Journal Plant Physiology, v. 13, p. 699-756, 1986.
SILVEIRA, J.A.G. Aspectos bioquímicos e fisiológicos da relação K/N em cana-deaçúcar (Saccharum spp.) cultivada em solução nutritiva. 1980. 127p. Tese
(Mestrado) – Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São
Paulo, Piracicaba, 1980.
SILVEIRA, J.A.G. Interações entre assimilação de nitrogênio e o crescimento de
cana-de-açúcar (Saccharum spp.) cultivada em condições de campo. 1985. 152p.
Tese (Doutorado) - Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de
São Paulo, Piracicaba, 1985.
47
SPRENT, J.I.; SPRENT, P. Nitrogen fixing organisms. 2ed., London: Chapman and
Hall, 1990. 256p.
STURZ, A.V. The role of endophytic bacteria during seed piece decay and potato
tuberization. Plant and Soil, v. 175, p. 257-263, 1995.
SUMAN, A.; GAUR, A.; SHRIVASTAVA, A.K.; YADAV, R.L. Improving sugarcane
growth and nutrient uptake by inoculating Gluconacetobacter diazotrophicus. Plant
Growth Regulation, v. 47, p. 155-162, 2005.
TAN, R. X.; ZOU, W. X. Endophytes: a rich source of functional metabolites. Natural
Product Reports, v.18, p.448-459, 2001.
TAPIA-HERNÁNDEZ, A.; BUSTILLOS-CRISTALES, M.R.; JIMENES-SALGADO,
T.; CABALLERO-MELLADO, J.; FUENTES-RAMÍREZ, L.E. Natural endophytic
occurrence of Acetobacter diazotrophicus in pineapple plants. Microbial Ecology, v.39,
p. 49-55, 2000.
TEIXEIRA, K.R.S. Acetobacter diazotrophicus, endófito diazotrófico associado à
cana-de-açúcar: presença de plasmídeos e sequenciamento do gene nifA,
responsável pela regulação da fixação biológica de nitrogênio. 1997. 152p. Tese
(Doutorado). Fundação Oswaldo Cruz, Rio de Janeiro, 1997.
TEJERA, N.; ORTEGA, E.; GONZALES-LOPES, J.; LLUCH, C. Effect of some
abiotic factors on the biological activity of Gluconacetobacter diazotrophicus. Journal
of Applied Microbiology, v. 95, p. 528-535, 2003.
TRIVELIN, P.C.O. Utilização do nitrogênio pela cana-de-açúcar: três casos
estudados com uso do traçador 15N. 2000. 143p. Livre docência, Escola Superior de
Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba, 2000.
URQUIAGA, S.; CRUZ, K.H.S.; BODDEY, R.M. Contribution of nitrogen fixation to
sugarcane: nitrogen-15 and nitrogen balance estimate. Soil Science Society of America
Journal, Madison, v.56, p.105-114, 1992.
48
WEBER, O. B. Contribuição de bactérias diazotróficas na cultura da bananeira:
perspectivas de utilização na produção integrada. Boletim de Pesquisa e
Desenvolvimento nº15, Jaguariúna: Embrapa Meio Ambiente, 2003.15p.
WOOD, A.W.; MUCHOW, R.C.; ROBERTSON, M.J. Growth of sugarcane under high
input conditions in tropical Australia. Fields Crops Research, v. 48, 1996, p.223-233.
YABUUCHI, E.; KOSAKO, Y.; OYAIZU, H.; YANO, I.; HOTTA, H.; HASHIMOTO,
Y.; EZAKI, T.; ARAKAWA, M. Proposal of Burkholderia gen. nov. and transfer of
seven species of the genus Pseudomonas homology group II to the new genus, with the
type species Burkholderia cepacia (Palleroni and Holmes, 1981) comb. nov.
Microbiology and Immunology, Tokyo, v.36, p.1251-1275, 1992.
YONEYAMA, T.; MURAOKA, T.; KIM, T.H.; DACANAY, E.V.; NAKANISHI, Y.
The natural 15N abundance of sugarcane and neighboring plants in Brazil, the
Philippines and Miyako (Japan). Plant and Soil, Drodrecht, v.189, p.239-244, 1997.
YOUNG, J.P.W. Phylogenetic classification of nitrogen-fixing organisms. In:
STANCEY, G.; BURRIS, R.H.; EVANS, J., (Ed.). Biological Nitrogen Fixation. New
York: Chapman, p.43-86, 1992.
49