PRODUÇÃO DE ENERGIA
-GLICÓLISE:
Quebra do açúcar. Ocorre no citoplasma das células,Segunda etapa da
degradação dos alimentos, a primeira é a quebra na digestão dos alimentos em
suas unidades fundamentais, ex. Proteínas em aminoácidos .Produz ATP em
ausência de O2 . Para cada molécula de glicose (6 c) são gastos dois ATP e
produzidos 2 piruvatos (3 C) e 4 ATP ( saldo dois ). São 10 reações catalisadas
por diferentes enzimas (terminação ase) em cada uma é produzido um açúcar
intermediário diferente.
O 1º passo na Glicólise é fosforilar a Glicose transformando-a em
Glicose 6-Fosfato, pois assim ela não consegue deixar a membrana
plasmática, ficando aprisionadas dentro das células. A Glicólise utiliza,
também, como estratégia a formação de moléculas intermediárias com 3
carbonos que facilmente transfiram os grupamentos fosfato para o ADP,
facilitando a síntese de ATP.Para poder ser utilizada na produção de energia, a
glucose-6-fosfato é primeiro isomerizada a frutose-6-fosfato. A frutose-6-fosfato
é depois fosforilada a frutose-1,6-difosfato numa reação catalisada pela
fosfofrutoquinase. Este é o ponto de não-retorno desta via metabólica: a
partir do momento em que a glicose é transformada em frutose-1,6-difosfato já
não pode ser usada em nenhuma outra via. Dai ela é quebrada em dois
gliceraldeido 3-P (3C cada).
CICLO DE KREBS:
O piruvato é transformado pela piruvato desidrogenase em acetil-coa
ainda no citoplasma. Daí a enzima citrato sintetase é quem transporta acetilcoa para dentro da mitocôndria onde se combina com oxaloacetato ( produzido
num ciclo anterior). A quantidade de ATP produzidos regula o ciclo. Por ciclo
são consumidos dois O2 e produzidos dois CO2. Por sua vez , a combinação de
acetil coa com oxaloacetato gera um composto de cinco carbonos, o alfacetoglutarato com libertação de NADH, e de CO2. O alfa-cetoglutarato vai dar
origem a outros compostos de quatro carbonos com formação de GTP, FADH 2
e NADH e oxaloacetato. Após o ciclo de krebs ocorre outro processo
denominado fosforilação oxidativa. A fosforilação oxidativa é uma via
metabólica que utiliza energia libertada pela oxidação de nutrientes de forma a
fosforilar ADP à ATP, apartir da transferência de eletróns, ocorre na
mitocôndria.
REGULAÇÃO:
Quando há excesso de acetil coa, a enzima piruvato desidrogenase é
inibida, assim como quando há excesso de ATP o ciclo de Krebs também. A
degradação de ácidos graxos leva à produção de acetil côa por um processo
chamado beta oxidação dos ácidos graxos, esta ocorre na mitocôndria.
Glicólise enzimas regulatórias, irreversíveis: Hexoquinase (fosforila Glc a Glc6P, gasta ATP) fosfrutoquinase (produz frutose 1,6 DP, gastando ATP) piruvato
quinase ( produz piruvato de fosfoenolpiruvato, gera ATP).
GLICONEOGÊNESE
Gliconeogênese é a biossíntese de glicose a partir de substâncias como
lactato, glicerol, oxaloacetato, aminoácidos e a partir de alguns carboidratos
(ex. Quando célula já esta lotada de ATP ou de piruvato). Este conjunto de
reações ocorre no citossol e utiliza muitas enzimas da via glicolítica, mas na
direção inversa. Enquanto a glicólise gera 2 ATPs por molécula de glicose
oxidada, a gliconeogênese consome 6 ATPs. A neoglicogênese é de
importância equivalente à glicólise, sendo portanto a via anabólica central. É
no entanto, uma via diferente daquela, apesar do número de enzimas em
comum. Os pontos de diferença (ditos os três desvios da neoglicogênese)
estão justamente nas enzimas regulatórias (que são pontos irreversíveis da
glicólise), e permitem uma regulação coordenada e recíproca. Como o piruvato
pode ser proveniente também da fermentação, a entrada também se dá via
lactato. Além do piruvato e lactato os pontos de entrada são: intermediários do
Ciclo do Ácido Cítrico (pois o acréscimo de um deles proporciona maior síntese
de oxaloacetato), aminoácidos glicogênicos (por produzirem diretamente
piruvato ou qualquer intermediário do Ciclo do Ácido Cítrico) e o glicerol
(pulando o primeiro desvio). Lembre que o Acetil-CoA jamais pode ser
convertido a glicose, contribuindo apenas no fornecimento de energia e NADH.
Daí a importância desta via: fornecer glicose para as células que não são
capazes de utilizar lipídios como fonte de energia (eritrócitos, neurônios, etc)
Em animais superiores, ocorre largamente no fígado e em pequena proporção,
no córtex renal.
Piruvato não pode ser transformado em fosfoenolpiruvato (PEP) por
ação da piruvato cinase, por isso, na mitocôndria, ele sofre a ação da piruvato
carboxilase na presença de dióxido de carbono e é transformado em
oxaloacetato. Este composto não atravessa a membrana interna da
mitocôndria, mas pode ser transformado em malato (produto da redução do
oxaloacetato), que migra para o citosol e aí é oxidado transformando-se em
oxaloacetato. A enzima fosfoenolpiruvato carboxicinase, presente tanto na
mitocôndria como no citosol, catalisa a transformação de oxalacetato em
fosfoenolpiruvato
(PEP).
As etapas de PEP até frutose-1,6-bisfosfato são etapas da via glicolítica,
invertidas. Na seqüência, frutose-1,6-bisfosfatase gera F6P, que é, em
seguida, transformada em G6P. A última etapa é catalisada pela glicose-6fosfatase, com a liberação de glicose.
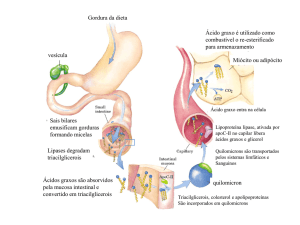
BETA OXIDAÇÃO DOS ÁCIDOS GRAXOS;
A maior parte da reserva energética do organismo encontra-se
armazenada sob a forma de triacilgliceróis. Estes podem ser hidrolisados por
lipases a glicerol e ácidos graxos. O glicerol pode ser convertido em DHAP
(dihidroxiacetona) seguindo a via glicolítica, os ácidos graxos na mitocondria
sofrerão beta oxidação.
A oxidação mitocondrial de ácidos graxos é um processo catabólico de
ácidos graxos. Eles sofrem remoção, por oxidação, de sucessivas unidades de
dois átomos de carbono na forma de acetil-CoA. Como exemplo pode ser
citado o ácido palmítico, que é um ácido graxo de 16 carbonos, ele vai sofrer
sete reações oxidativas perdendo em cada uma delas dois átomos de carbono
na forma de acetil-coa. Ao final desse processo os dois carbonos restantes
estarão na forma de acetil-coa (cada volta no ciclo 18 ATP são produzidos).
Mas quando a cadeia de ácidos graxos for ímpar, o produto final da β-oxidação
será o propionil-CoA (3 CH3), esse composto, através da incorporação de CO2
e gasto energético através de quebras de ligações do ATP, se transforma em
succinil-CoA, que é um composto do Ciclo de Krebs.Após a β-oxidação, os
resíduos acetil do acetil-CoA são oxidados até chegarem a CO2, o que ocorre
no ciclo de Krebs. Em acidos graxos insaturados há enzimas apropriadas para
permitirem a beta oxidação.
Portanto a energia gerada na oxidação de ácidos graxos vai ser
conservada na forma de ATP. Sendo que a oxidação de ácidos graxos produz
muito mais energia que a oxidação de carboidratos. Uma molécula de
palmitato, por exemplo, produz mais de 100 ATPs, enquanto uma molécula de
glicose produz apenas 38. Para entrarem na mitocôndria os ácidos graxos
necessitam do aminoácido carnitina, pois a membrana da mitocôndria é
impermeável aos acetil-CoA sem este transportador.
Biossíntese de ácidos graxos:
Em situações de abundância de acetil-CoA, o fígado e o tecido adiposo
sintetizam ácidos graxos. O processo de síntese apresenta bastantes
semelhanças com o inverso da b-oxidação, mas também tem diferenças
importantes:
ocorre no citoplasma, e não na mitocôndria.
usa NADPH como fonte de elétrons.
o transportador de grupos acetil é a ACP (Enzima transportadora de
grupos acetil), e não a coenzima A.
Síntese de corpos cetónicos (Cetogênese) :
Uma grande quantidade do acetil-CoA produzido pela b-oxidação dos
ácidos graxos nas mitocôndrias do fígado é convertida em acetoacetato e bhidroxibutirato (também denominados corpos cetônicos). Estes compostos
podem ser usados pelo coração e pelos músculos esqueléticos para produzir
energia. O cérebro, que normalmente depende da glicose como fonte de
energia, pode também utilizar corpos cetônicos durante um jejum prolongado
(maior do que 2-3-dias). A síntese de corpos cetônicos começa pela
condensação de duas moléculas de acetil-CoA, para formar acetoacetil-CoA. O
acetoacetato assim produzido passa para a corrente sanguínea e é distribuído
pelos tecidos. Uma vez absorvido, reage na mitocôndria com o succinil-CoA,
produzindo succinato e acetoacetil-CoA, que pode ser clivado em duas
moléculas de acetil-CoA.