UNIVERSIDADE ESTADUAL DE SANTA CRUZ
GEDEON ALMEIDA GOMES JÚNIOR
INFLUÊNCIA DE FUNGOS MICORRIZICOS ARBUSCULARES NO
CRESCIMENTO E NUTRIÇÃO DE MUDAS DE GRAVIOLEIRA (Annona muricata
L.) SUBMETIDAS A ADUBAÇÃO ORGÂNICA
ILHÉUS – BAHIA
2015
GEDEON ALMEIDA GOMES JÚNIOR
INFLUÊNCIA DE FUNGOS MICORRIZICOS ARBUSCULARES NO
CRESCIMENTO E NUTRIÇÃO DE MUDAS DE GRAVIOLEIRA (Annona muricata
L.) SUBMETIDAS A ADUBAÇÃO ORGÂNICA
Dissertação apresentada ao Programa de Pósgraduação em Produção Vegetal da Universidade
Estadual de Santa Cruz como parte dos requisitos
para a obtenção do título de Mestre em Produção
Vegetal.
Área de concentração: Cultivos em ambiente
tropical úmido.
Orientador: Prof. Dr. Célio Kersul do Sacramento
Coorientador: Prof. Dr. Eduardo Gross
ILHÉUS – BAHIA
2015
GEDEON ALMEIDA GOMES JÚNIOR
INFLUÊNCIA DE FUNGOS MICORRIZICOS ARBUSCULARES NO
CRESCIMENTO E NUTRIÇÃO DE MUDAS DE GRAVIOLEIRA (Annona muricata
L.) SUBMETIDAS A ADUBAÇÃO ORGÂNICA
_______________________________________
Célio Kersul do Sacramento
UESC (Orientador)
_______________________________________
José Basílio Vieira Leite
CEPLAC (Avaliador)
______________________________________
Paulo Cesar Lima Marrocos
UESC (Avaliador)
DEDICO
Aos meus pais!!!
AGRADECIMENTOS
A Deus pela vida, e por todas as oportunidades e bênçãos que tem me proporcionado.
Ao Programa de Pós-Graduação em Produção Vegetal – PPGPV que proporcionou a
aquisição de novos conhecimentos e o desenvolvimento deste trabalho
A Biofábrica do Cacau pela ajuda.
Ao meu orientador Célio Kersul do Sacramento.
Ao meu co-orientador e amigo, Prof. Dr. Eduardo Gross, pela oportunidade de
aprendizado, pelas valiosas sugestões, pelo incentivo, pelas injeções de ânimo, apoio
nos momentos de maiores dificuldades e pela confiança que sempre depositou em
mim.
Ao professor George Sodré pelo auxilio sempre que foi preciso.
A coordenação do Laboratório de Química e Fertilidade do solo, por ter disponibilizado
toda estrutura durante o planejamento e execução do estudo.
A meus tios Mitermayer Galvão e Eliana Reis pela ajuda, conselhos e confiança
sempre depositada em mim.
À Coordenação do Aperfeiçoamento de Pessoal de Nível Superior (CAPES), pela
concessão da bolsa durante a realização do curso de mestrado.
À CEPLAC, (Laboratório de Análises de tecidos vegetais) em especial aos técnicos
Maurino, José e Vilma Mororó pelo auxilio nas análises.
Aos colegas do Pós-Graduação em Produção Vegetal, em especial a amiga Rafaela
Alves por toda a ajuda.
Aos colegas de minha turma de mestrado, em especial ao colega Mateus Caliman.
A todos os amigos que não foram aqui citados, mas que sabem do meu
reconhecimento e apreço.
LISTA DE FIGURAS
1. Vista parcial do experimento com mudas de gravioleira em solo com diferentes
doses de composto orgânico e inoculadas com E. colombiana, A. scrobiculata e a
mistura
desses
dois
fungos
micorrízicos,
e
não
inoculadas
(controle)....................................................................................................................16
2. Gráfico da análise de regressão para a média da massa seca total das plantas de
gravioleira submetido a diferentes doses de composto orgânico e inoculado com
Entrophospora colombiana, Acaulospora scrobiculata e a mistura desses dois fungos
micorrízicos
(MIX),
e
não
inoculados
(controle)....................................................................................................................22
3. Gráfico da análise de regressão para o grau de colonização micorrízica da
gravioleira submetido a diferentes doses de composto orgânico e inoculado com
Entrophospora colombiana, Acaulospora scrobiculata e a mistura desses dois fungos
micorrízicos (MIX), e não inoculados (controle)..........................................................26
4. Raízes de gravioleira dos tratamentos controle e inoculado com A. scrobiculata,
Entrophospora colombiana e a mistura desses fungos micorrízicos arbusculares
(FMAs) submetida a diferentes doses de composto orgânico.....................................28
5. Análise de regressão para o número de esporos no solo após o plantio com
gravioleira inoculados com E. colombiana, A. scrobiculata e a mistura desses dois
fungos micorrízicos (Mix), e não inoculados (controle) submetidos a diferentes doses
de composto orgânico.................................................................................................30
6. Análise de regressão para a concentração de glomalina no solo após o plantio com
gravioleira inoculados com E. colombiana, A. scrobiculata e a mistura desses dois
fungos micorrízicos (Mix), e não inoculados (controle) submetidos a diferentes doses
de composto orgânico.................................................................................................32
7. Análise de regressão para o teor de K nas folhas da gravioleira inoculadas com E.
colombiana, A. scrobiculata e a mistura desses dois fungos micorrízicos (Mix), e não
inoculados
(controle)
submetidos
a
diferentes
doses
de
composto
orgânico......................................................................................................................35
8. Análise de regressão para o teor de Mg nas folhas da gravioleira inoculadas com
E. colombiana, A. scrobiculata e a mistura desses dois fungos micorrízicos (Mix), e
não inoculados (controle) submetidos a diferentes doses de composto
orgânico......................................................................................................................36
9. Análise de regressão para o teor de P nas folhas da gravioleira inoculadas com E.
colombiana, A. scrobiculata e a mistura desses dois fungos micorrízicos (Mix), e não
inoculados
(controle)
submetidos
a
diferentes
doses
de
composto
orgânico......................................................................................................................37
10. Análise de regressão para o teor de Ca nas folhas da gravioleira submetidas a
diferentes doses de composto orgânico......................................................................38
11. Análise de regressão para o teor de Fe nas folhas da gravioleira submetidas a
diferentes doses de composto orgânico......................................................................41
12. Análise de regressão para o teor de Zn nas folhas da gravioleira inoculadas com
E. colombiana, A. scrobiculata e a mistura desses dois fungos micorrízicos (Mix), e
não inoculados (controle) submetidos a diferentes doses de composto
orgânico......................................................................................................................42
LISTA DE TABELAS
1. Resumo da Análise de Variância para altura e diâmetro do coleto das
plantas........................................................................................................................20
2. Valores médios de altura e diâmetro do coleto de mudas de gravioleira cultivadas
em solo com diferentes doses de composto orgânico e inoculados com E. colombiana,
A. scrobiculata e a mistura desses dois fungos micorrízicos (MIX), e não inoculados
(controle)....................................................................................................................20
3. Resumo da Análise de Variância para matéria seca total das
plantas....................................................................................................................... 21
4. Valores médios de matéria seca total (MST) de mudas de gravioleira cultivadas em
solo com diferentes doses de composto orgânico e inoculados com E. colombiana, A.
scrobiculata e a mistura desses dois fungos micorrízicos (MIX), e não inoculados
(controle)....................................................................................................................22
5. Resumo da Análise de Variância para dependência micorrizica da
gravioleira...................................................................................................................23
6. Valores médios de dependência micorrizica das plantas submetidas diferentes
doses de composto orgânico e inoculado com inoculados com E. colombiana, A.
scrobiculata
e
a
mistura
desses
dois
fungos
micorrízicos
(MIX)...........................................................................................................................24
7. Níveis categóricos (Habte e Manjunath, 1991) de dependência micorrizica da
gravioleira submetidas diferentes doses de composto orgânico e inoculado com
inoculados com E. colombiana, A. scrobiculata e a mistura desses dois fungos
micorrízicos (MIX).......................................................................................................24
8. Resumo da Análise de Variância para porcentagem de colonização micorrízica
gravioleira...................................................................................................................25
9. Valores médios do grau de colonização micorrizica de mudas de gravioleira
cultivadas em solo sob diferentes doses de composto orgânico e inoculado com
inoculados com E. colombiana, A. scrobiculata e a mistura desses dois fungos
micorrízicos (MIX), e não inoculados (controle)..........................................................25
10. Resumo da Análise de Variância para densidade de esporos em 50g de solo
rizosférico ao final do experimento..............................................................................29
11. Valores médios do número de esporos no solo após o plantio com gravioleira
inoculados com E. colombiana, A. scrobiculata e a mistura desses dois fungos
micorrízicos (Mix), e não inoculados (controle) submetidos a diferentes doses de
composto orgânico.....................................................................................................29
12. Resumo da Análise de Variância para teor de glomalina.......................................31
13. Valores médios para a concentração de glomalina no solo após o plantio com
gravioleira inoculados com E. colombiana, A. scrobiculata e a mistura desses dois
fungos micorrízicos (Mix), e não inoculados (controle) e submetidos a diferentes doses
de composto orgânico.................................................................................................31
14. Resumo da análise de variância para o macronutrientes na folha da gravioleira
submetida a diferentes doses de composto orgânico e inoculada com Entrophospora
colombiana, Acaulospora scrobiculata e a mistura desses dois fungos micorrízicos
(Mix), e não inoculado (controle).................................................................................33
15. Média do teor de macronutrientes nas folhas da gravioleira submetida a diferentes
doses de composto orgânico e inoculada com Entrophospora colombiana,
Acaulospora scrobiculata e a mistura desses dois fungos micorrízicos (Mix), e não
inoculado (controle)....................................................................................................34
16. Resumo da análise de variância para as micronutrientes na folha da gravioleira
submetida a diferentes doses de composto orgânico e inoculada com Entrophospora
colombiana, Acaulospora scrobiculata e a mistura desses dois fungos micorrízicos
(Mix), e não inoculado (controle).................................................................................39
17. Média do teor de macronutrientes nas folhas da gravioleira submetida a diferentes
doses de composto orgânico e inoculada com Entrophospora colombiana,
Acaulospora scrobiculata e a mistura desses dois fungos micorrízicos (Mix), e não
inoculado (controle)....................................................................................................40
SUMÁRIO
RESUMO................................................................................................................... xii
ABSTRACT .............................................................................................................. xiii
1. INTRODUÇÃO ...................................................................................................... 1
2. OBJETIVOS.......................................................................................................... 3
3. REVISÃO DE LITERATURA ................................................................................. 4
3.1
Aspectos gerais sobre a cultura da gravioleira (Annona muricata L.) ............ 4
3.2
Adubação orgânica ........................................................................................ 5
3.3
Fungos micorrízicos arbusculares .................................................................. 7
3.4
Dependência micorrizica ................................................................................ 9
3.5
Produção de glomalina por fungos micorrízicos arbusculares ..................... 10
4. MATERIAL E MÉTODOS ................................................................................... 13
4.1
Preparo do solo ............................................................................................ 13
4.2
Avaliação da capacidade de campo ............................................................. 13
4.3
Curva de incubação do calcário ................................................................... 14
4.4
Preparo do composto orgânico .................................................................... 14
4.5
Obtenção de inóculos de fungos micorrízicos arbusculares ........................ 15
4.6
Estabelecimento das doses do composto orgânico ..................................... 15
4.7
Condução do experimento ........................................................................... 15
4.8
Avaliação da massa seca............................................................................. 16
4.9
Dependência micorrizica .............................................................................. 16
4.10 Teor de nutrientes na folha .......................................................................... 17
4.11 Coloração de raízes micorrizadas ................................................................ 17
4.12 Quantificação de raízes micorrizadas .......................................................... 18
4.13 Teor de Gomalina facilmente extraível ......................................................... 18
4.14 Extração e contagem dos esporos de fungos micorrízicos arbusculares ..... 19
4.15 Analise estatística ........................................................................................ 19
5. RESULTADOS ................................................................................................... 20
5.1
Altura e diâmetro do coleto das plantas ....................................................... 20
5.2
Produção de matéria seca ........................................................................... 21
5.3
Dependência micorrízica .............................................................................. 23
5.4
Grau de colonização micorrizica .................................................................. 25
5.5
Densidade de esporos ................................................................................. 29
5.6
Glomalina no solo ........................................................................................ 31
5.7
Nutrientes na folha ....................................................................................... 33
5.7.1
Macronutrientes ..................................................................................... 33
5.7.2
Micronutrientes ...................................................................................... 39
6. DISCUSSÃO....................................................................................................... 44
6.1
Altura e diâmetro do coleto .......................................................................... 44
6.2
Produção de massa seca ............................................................................. 45
6.3
Dependência micorrízica .............................................................................. 45
6.4
Grau de colonização micorrízica .................................................................. 46
6.5
Densidade de esporos ................................................................................. 47
6.6
Glomalina no solo ........................................................................................ 48
6.7
Nutrientes na folha ....................................................................................... 49
7. CONCLUSÃO ..................................................................................................... 51
8. REFERÊNCIAS .................................................................................................. 52
xii
RESUMO
Os fungos micorrízicos arbusculares (FMAs), em solos de baixa fertilidade natural,
geralmente, promovem aumento no crescimento beneficiando a produção de massa
seca nas plantas hospedeiras, entretanto a depender da espécie vegetal em questão
os efeitos desses fungos podem ser diferenciados. Dessa forma o presente trabalho
foi proposto com o objetivo de avaliar a influência da adição de composto orgânico à
base de casca de cacau, no crescimento inicial de mudas de gravioleira inoculadas
com FMAs. O experimento foi conduzido em casa de vegetação utilizando solo
esterilizado no qual foram aplicadas cinco doses do composto orgânico (0, 5, 10, 20 e
30 g dm-3) e quatro tratamentos micorrizicos (controle não inoculado, inoculação com
Acaulospora scrobiculata, com Entrophospora colombiana e a mistura desses
inoculos) com um delineamento em blocos casualizados, constando de quatro blocos.
Aos 150 dias após a montagem do experimento foram avaliados altura das plantas,
diâmetro do coleto, avaliação da massa seca das plantas, dependência micorrizica da
gravioleira, grau de colonização micorrizica, densidade de esporos, teor de glomalina
facilmente disponível e teores de macro e micronutrientes na folha. A massa seca da
gravioleira, bem como altura e diâmetro do coleto das plantas foram influenciados pela
interação dos fatores composto orgânico e FMAs, apresentando geralmente, ajuste
de modelos de natureza quadrática. A gravioleira foi considerada uma espécie
micotrófica obrigatória, sendo classificada como altamente dependente a simbiose
micorrizica, independente da dose do composto orgânico aplicada. O grau de
colonização micorrizica e a densidade de esporos de FMA foram influenciados pelas
doses do composto orgânico aplicado ao solo, e o modelo que melhor explicou os
resultados obtidos foi o quadrático. O teor de glomalina facilmente disponível no solo
pela inoculação de FMAs, assim como pela adição do composto orgânico ao solo. Os
teores de P, K, Mg e Zn foi influenciado pela interação dos fatores composto orgânico
e FMAs, a análise de regressão possibilitou ajuste linear para K e Mg e quadrático
para P e Zn. Os teores de Ca e Fe foram influenciados pela incorporação de composto
orgânico ao solo, com o teor de Ca elevando-se a medida em que aumentou-se a
dose do composto e no caso do Fe, reduções em seu teor foram observadas nas
doses mais elevadas. O conjunto de resultados permite concluir que a aplicação de
compostos orgânico concomitantemente à inoculação influenciaram positivamente o
crescimento e nutrição da gravioleira.
Palavras-chave: compostagem, nutrição de plantas, glomalina, anonáceas
xiii
ABSTRACT
In low fertility soils Arbuscular Mycorrhizal Fungi (AMF) generally promote increased
growth benefiting the dry mass of the host plants, however depending on the plant
species effects of AMF can be differentiated. Thus, the present study was proposed
with the aim of evaluating the influence of organic compost from cocoa shell base on
initial growth of soursop (Annona muricata L.) inoculated with AMF. The experiment
was conducted in a greenhouse using sterilized soil in which were applied five doses
of organic compost (0, 5, 10, 20 and 30 g dm-3) and four mycorrhizal treatments (noninoculated control, inoculation with Acaulospora scrobiculata, with Entrophospora
colombiana and the mixture of these inoculants) with a randomized block design,
consisting of four blocks. After 150 days of growth plants were evaluated on height, lap
diameter, dry mass, mycorrhizal dependence, degree of mycorrhizal colonization,
spore density, shoot macro and micronutrients and glomalin content from rhizospheric
soil. The dry mass of the soursop, as well as height and plant lap diameter were
influenced by the interaction of organic compost and AMF factors, showing usually
quadratic adjustment of models. Soursop is considered a mandatory micotrophic
species, being classified as highly dependent on the mycorrhizal symbiosis, regardless
of the dose of organic compost applied. The degree of mycorrhizal colonization and
density of AMF spores were influenced by doses of organic compost applied to the
soil, and the model that best explained the results was the quadratic. The readily
available glomalin content in soil was influenced by AMF inoculation as well as by
organic compound fertilization. The P, K, Mg and Zn was influenced by the interaction
of factors organic compost and AMF, the regression analysis allowed linear fit for K
and Mg and quadratic for P and Zn. Ca and Fe contents were influenced by the
incorporation of organic compost to the soil, with the Ca content rising the extent to
which increased the dose of the compost and in the case of Fe, reductions in their
content were observed in highest doses. The set of result showed that the application
of organic compost concurrently with inoculation positively influenced the growth and
nutrition of soursop.
Keywords: composting, plant nutrition, glomalin, Annonaceae
1
1. INTRODUÇÃO
O nordeste brasileiro apresenta condições favoráveis ao cultivo de várias
espécies frutíferas. Dentre essas, muitas têm importância econômica real ou
potencial, a exemplo da gravioleira (Annona muricata L.). A crescente demanda por
seus frutos, para consumo in natura e, principalmente, para o setor da agroindústria,
tem impulsionado a expansão da área cultivada, colocando a cultura como uma das
principais alternativas de investimento do setor frutícola nordestino (COSTA et al.,
2005).
O crescimento na produção de graviola vem sendo evidente nos estados da
Paraíba, Ceará, Pernambuco e Bahia (LIMA, 2003), sendo seus frutos utilizados na
fabricação de suco, sorvetes, compotas, geléias e doces.
A utilização dos recursos encontrados no próprio imóvel ou em suas
proximidades para a fabricação do fertilizante no local vem tornando-se uma pratica
cada vez mais comum. Em sistemas orgânicos, isso pode ser feito usando diferentes
práticas de adubação (BORGES et al., 2003). A fertilização é componente chave para
a manutenção da qualidade dos solos e, como consequência, para a sustentabilidade
dos sistemas produtivos em médio e longo prazo (SILVA e MENDONÇA, 2007).
Entretanto, a melhoria da capacidade produtiva do solo é um processo gradual onde
a matéria orgânica tem influência direta (BONILLA, 1992). Dessa forma, aproveitando
o potencial natural do ambiente e com o desenvolvimento de práticas agroecológicas,
a fertilização pode ser alcançada com uso de compostos orgânicos ou biofertilizantes,
promovendo assim sustentabilidade ao agroecossistema.
O sul da Bahia, também chamado de região cacaueira, devido à grande área
plantada de cacaueiros tem um grande potencial em gerar resíduos, estes são
advindos no período de colheita, pois o resíduo gerado em maior quantidade é a casca
do fruto. A quantidade de resíduos gerada por essa cultura é bastante significativa,
podendo chegar a acumular 7 toneladas de casca fresca para cada tonelada de
amêndoas. Essa matéria prima, que quase nunca é aproveitada, formando
amontoados no interior da lavoura, onde a quebra dos frutos é realizada. O acúmulo
desse material quando deixado ao solo sem nenhum tratamento acaba por constituir
fonte de inóculo para fungos patogênicos a cultura do cacau (MORORÓ, 2007).
Diversos destinos podem ser dados a esta matéria prima: alimentação animal
(ABIOLA,1991), extração de pectinas (BERBERT,1972) e gomas, (FIGUEIRA et al.,
2
1993) e por fim a compostagem, sendo esta uma pratica muito viável ao produtor,
otimizando a dinâmica da propriedade, pelo fato dos elevados teores de potássio
presente na casca do fruto do cacaueiro.
Nessa região, o cultivo da gravioleira, assim como de outras fruteiras tropicais,
tem possibilitado um incremento de renda aos produtores, sobretudo após o declínio
da lavoura cacaueira, tornando-se uma alternativa de diversificação agrícola para os
produtores rurais.
A micorriza, associação mutualística entre plantas e fungos, pode ser utilizada
de maneira proveitosa para a agricultura, considerando que a simbiose aumenta a
área de absorção de nutrientes pela planta, graças as hifas radiculares fungo, que
captam nutrientes, tendo uma maior importante em relação aqueles de mobilidade
reduzida no solo.
A aplicação de matéria orgânica em solos tropicais em forma de adubo de pátio
de fazenda, composto, adição de resíduos agrícolas ou adubo verde estimula a
proliferação de fungos micorrízicos arbusculares (FMA) (HARINIKUMAR e
BAGYARAJ, 1989). Assim como fontes orgânicas de nutrientes como os compostos
orgânicos e ainda fertilizantes naturais de baixa solubilidade, como o fosfato de rocha,
não parecem suprimir o crescimento e desenvolvimento dos fungos micorrizicos
arbusculares, podendo até estimular seu desenvolvimento (SILVA JUNIOR,2008).
Na literatura há poucos trabalhos que avaliem os efeitos do uso concomitante
de composto orgânico e fungos micorrizicos sobre o desenvolvimento de mudas de
gravioleira. A maioria dos trabalhos envolvendo simbiose micorrizica em gravioleira,
tem avaliado o comportamento desses microrganismos e da espécie vegetal em
questão, com adubação inorgânica. Corroborando assim, para a justificativa do
presente estudo, que tem como objetivo avaliar o desenvolvimento da gravioleira
submetida a adubação orgânica e inoculação com FMAs.
3
2. OBJETIVOS
Geral:
Avaliar a influência do composto orgânico e dos FMAs sobre o crescimento
inicial de mudas de gravioleira.
Específicos:
Analisar o efeito das diferentes doses do composto orgânico sobre os fungos
micorrízicos inoculados; avaliar o grau de dependência da gravioleira em relação aos
FMAs; e avaliar a influência das diferentes espécies de fungos micorrízicos e das
doses do composto orgânico na nutrição de mudas de graviola.
4
3. REVISÃO DE LITERATURA
3.1
Aspectos gerais sobre a cultura da gravioleira (Annona muricata L.)
A gravioleira (Annona muricata L.), família Annonaceae, tem a América Central
e vales peruanos como centro de origem, sendo considerada por muitos a mais
tropical das anonáceas (RAMOS et al., 2001). Plantas de A. muricata podem ser
encontradas na forma silvestre, assim como cultivada em uma ampla faixa territorial,
abrangendo regiões desde o nível do mar até altitudes superiores a 1100 m
(MORTON, 1966).
Annona muricata L. faz parte de um grupo de espécies frutíferas de importância
econômica em inúmeros países, dentre eles: Venezuela, Colômbia, Porto Rico, Costa
Rica, México, Panamá, Jamaica, Cuba, Espanha, Índia, Honduras, Guiana, Suriname,
Brasil, Peru, Senegal e Cingapura (RAMOS et al., 2001). No Brasil, a graviola é
amplamente cultivada nas regiões Norte, Nordeste, Centro-Oeste e Sudeste
(JUNQUEIRA et al., 1996), destacando-se os Estados de Alagoas, Bahia, Ceará,
Distrito Federal, Minas Gerais, Pará, Paraíba e Pernambuco como grandes produtores
(SACRAMENTO et al., 2009).
O Estado da Bahia, especialmente as regiões Sul e Extremo Sul, devido às
suas condições edafoclimáticas favoráveis ao cultivo dessa frutífera, ocupa o primeiro
lugar a nível nacional em produção e área plantada, de acordo com levantamentos
realizados pela Agência de Defesa Agropecuária da Bahia (ADAB, 2010). Nesses
levantamentos, a área plantada georreferenciada é de cerca de 1.300 ha e estima-se
que a produção estadual em 2010 foi de aproximadamente 8 mil toneladas.
A gravioleira apresenta crescimento ereto, com altura média em sua fase adulta
de 4 a 8 metros, apresentando um caule único e ramificação assimétrica. Inicia a sua
fase reprodutiva entre o terceiro e o quinto ano e alcança o seu pleno desenvolvimento
entre o sexto e o oitavo ano (FREITAS, 1997). As folhas apresentam pecíolo curto,
são oblongo-lanceoladas ou elípticas, medem 14 a 16 cm de comprimento e 5 a 7 cm
na maior largura; as nervuras são pouco perceptíveis (MANICA, 1997).
Suas flores são hermafroditas, de cor verde-escura, tornando-se verde-clara
quando aproxima-se do estádio de antese. São distribuídas em pedúnculos curtos
axilares, podendo estar diretamente no tronco, agrupadas de 2 a 4 flores (MANICA,
1997). Apresentam cálice com 3 sépalas pequenas e sua corola formada por 6 pétalas
5
carnosas (PINTO e GENÚ, 1984). Apresentam androceu com diversos estames, com
2 anteras cada; o gineceu é característico por apresentar numerosos pistilos, com
ovário supero, uninucleolar e uniovulado (PINTO e GENÚ, 1984; PINTO E SILVA,
1994; PINTO et al., 2001).
O fruto em si trata-se de uma baga composta, fruto múltiplo, carnoso, com peso
variando de 0,4 a 10 kg (MANICA, 1997). Possui formato variável, casca em cor verdeescura quando os frutos ainda estão imaturos e verde-clara quando estão apropriados
para colheita, possuindo ainda espículas carnosas moles (PINTO e SILVA, 1994). A
polpa, que se trata da parte de interesse econômico apresenta coloração branca,
sendo ainda ligeiramente ácida e muito rica em vitamina A e ácido ascórbico.
(MANICA, 1997). Possui até 490 sementes, as quais são obovoides, aplainadas,
medem de 17 a 20 mm de comprimento e pesam de 0,57 a 0,61 g, com a testa dura
e de cor marrom-escura-brilhante (SACRAMENTO et al., 2009).
A gravioleira, adapta-se em diferentes tipos de solo, tendo preferência por solos
profundos, com drenagem e retenção de umidade satisfatória, e com pH ligeiramente
ácido (MELO et al., 1983). A incorporação de fontes de adubo orgânico é fundamental
a um bom desenvolvimento da cultura, por esta apresentar raízes superficiais, o que
possibilita um aproveitamento mais eficiente dos nutrientes (SÃO JOSÉ, 2003).
3.2
Adubação orgânica
A utilização de compostos orgânicos no solo vem sendo uma pratica bastante
utilizada no cultivo de inúmeras culturas, sendo essa pratica de grande importância
ao sistema solo-planta pois além de melhorar a fertilidade dos solos onde estes vem
sendo utilizados, acabam por melhorar características como formação de agregados,
aumento da porosidade, maior retenção de agua, aumento da capacidade de troca
catiônica e aumento da diversidade microbiana do solo (MIYASAKA et al., 1997).
Em regiões tropicais e subtropicais, onde a matéria orgânica decompõe-se com
grande rapidez (MALAVOLTA et al., 2002) a adubação orgânica apresenta-se como
uma pratica bastante interessante, tornando-se uma componente chave na
manutenção e melhora de solos tropicais e dando sustentabilidade para a manutenção
de sistemas produtivos tanto a médio como a longo prazo (SILVA e MENDONÇA,
2007).
6
No passado, o uso de adubo orgânico aplicado ao solo pelos agricultores era
praticado em conjunto com fertilizantes minerais, tais como NPK, sulfato de amônio,
nitrato de potássio e superfosfatos, sendo que seu uso quase foi abandonado devido
utilização dos adubos fabricados pelo homem (OLAWUYI et al., 2012). Nos últimos
anos, a utilização de produtos naturais como substitutos para os fertilizantes sintéticos
convencionais assume uma grande importância (THIRUMARAN et al., 2007). A
produção de adubos orgânicos pode ser feita com a utilização de fontes diversas de
matéria orgânica, podendo até contribuir para a proteção ambiental.
O despejo indiscriminado de lodo de esgoto, por exemplo, em áreas abertas e
rios é um grave problema em muitos países por ocasionar a degradação dos
ecossistemas (KAKULU e OSIBANJO, 1992), problema que pode ser resolvido por
meio de um planejamento social e ambiental, pensando na utilização na agricultura
do composto originado no final do processo de tratamento do lodo de esgoto. Esse
fato é mais agravante em países onde pouco ou nenhum tratamento é realizado nos
materiais antes de serem descartados.
Evidencias experimentais revelam que substancias húmicas participam de
importantes reações e acabam por influenciar as condições química, físicas e
biológicas do solo (CANELLAS et al. 1999). Efeitos diretos dessas substâncias sobre
o metabolismo das plantas podem ser considerados como resultado de uma influência
positiva sobre o transporte de íons, aumento da respiração e da velocidade das
reações enzimáticas do ciclo de Krebs, aumento no conteúdo de clorofila, aumento na
síntese de ácidos nucleicos (NANNIPIERI et al.1993). Entretanto estudos ainda
precisam ser desenvolvidos, para elucidar o funcionamento da bioatividade dessas
substâncias.
Fertilizantes de natureza orgânica são constituídos por substancias húmicas
que correspondem 65-75% da matéria orgânica do solo, sendo tema de estudos em
diversas áreas da agricultura, com destaque para química do solo, fisiologia vegetal,
química do solo, bem como ciências ambientais, o que pode ser corroborado pelas
múltiplas funções que esses materiais de origem orgânica podem desempenhar no
desenvolvimento de plantas (CACCO e AGNOLLA, 1984).
O emprego dos compostos orgânicos como base central de sistemas orgânicos
de produção é uma tecnologia adotada no mundo inteiro e é uma alternativa viável
para o aproveitamento racional de resíduos, sendo o método mais antigo de
reciclagem. (ALVES, 2012).
7
Levando isso em conta o Ministério da Agricultura e do Abastecimento possui
uma instrução normativa de Nº 007, datada em 17 de maio de 1999, onde se torna
permitido o uso de adubos e condicionadores obtidos de restos orgânicos sólidos ou
líquidos, de origem animal ou vegetal, oriundos ou não da unidade de produção desde
que a certificadora responsável pela fiscalização de uma produção orgânica aprove a
utilização. Uma outra Instrução Normativa SDA/MAPA nº 25 de 2009, estabeleceu que
fertilizantes orgânicos simples, mistos ou compostos para aplicação no solo deverão
seguir alguns requisitos. Dentre esses: N total (mín.) 0,5%, carbono orgânico (mín.)
15 %, pH (mín.) de 6,0 e relação C/N (máx.) de 20.
3.3
Fungos micorrízicos arbusculares
Existem relatos do surgimento de fungos micorrízicos há mais de 400 milhões
de anos, quase que coincidindo com o surgimento das espécies vegetais terrestres
(BERBARA et al., 2006). Entre 70 e 90% das plantas apresentam associação com os
fungos micorrízicos arbusculares, incluindo algumas espécies de pteridófitas e
briófitas, indicando que a simbiose mutualística entre raiz e fungo é mais regra do que
exceção na natureza (MOREIRA e SIQUEIRA, 2006).
Os dois principais tipos de micorrizas e de maior ocorrência nos ecossistemas
são as ectomicorrizas e as micorrizas arbusculares. As ectomicorrizas são formadas,
na maioria, por fungos septados em geral pertencentes aos Basidiomicetos, que só
penetram intercelularmente no córtex das raízes, com formação de estrutura
anatômica característica, a rede de Hartig, substituindo a lamela média e também
ocorrendo a formação do manto fúngico ao redor das raízes. Esse tipo de micorriza
se caracteriza ainda pelas intensas modificações morfológicas das raízes colonizadas,
sendo típico de árvores de clima temperado, como as coníferas (ex.: Pinus). Ocorrem
também em plantas tropicais das famílias Caesalpiniaceae, Dipterocarpaceae e
Myrtaceae.
Aprofundando o conhecimento em FMA, esta designação é a mais recente
para as então denominadas micorrizas vesículo-arbusculares. Elas são formadas por
fungos classificados como Glomeromycota (ordem Glomales e classe Zygomycotina),
que são asseptados e colonizam as raízes de plantas de quase todos os gêneros das
Gimnospermas e Angiospermas, além de alguns representantes das Pteridófitas e dos
gametófitos das Briófitas (PETERSON et al., 1981; ALLEN, 1996). São as mais
8
abundantes e geralmente as menos específicas no que diz respeito ao fitossimbionte
(MOLINA, MASSICOTTE e TRAPPE, 1992).
As indicações apontam que 80% das espécies vegetais formam esse tipo de
micorriza, onde o fungo coloniza as células do córtex inter e intracelularmente, de
modo muito característico, formando os arbúsculos, estruturas intra-radiculares
altamente ramificadas e típicas das MA’s (MOREIRA e SIQUEIRA, 2006). Além disso,
é provável que eles sejam os fungos de solo mais abundantes na maioria dos
ecossistemas tropicais, principalmente nos sistemas agrícolas, onde eles podem
representar quase 50% da biomassa microbiana (OLSSON et al., 1999).
A simbiose se resume basicamente em uma troca onde as plantas acabam
sendo beneficiadas com o aumento da absorção de agua e nutrientes, sendo o fosforo
o nutriente que apresenta uma maior dependência das FMA. Enquanto os fungos
envolvidos nesse processo recebem das plantas os fotoassimilados necessários para
que ciclo reprodutivo desses fungos seja completado (CARNEIRO et al., 1998).
O beneficio da simbiose micorrizica sobre o crescimento na maioria das plantas
é bem mais significativo quando se trata de nutrientes de baixa mobilidade no solo,
como fosforo e zinco, que movem-se em direção ao sitema radicular das plantas
atraves de mecanismos de difusão (MARSCHNER, 1995).
As hifas dos FMA aumentam de forma considerável a área de absorção das
raízes explorando lugares onde as raízes dificilmente alcançariam, e por isso
geralmente acumulam maiores quantidades de nutrientes. Dentre os nutrientes,
fósforo e zinco são os nutrientes que tem sua absorção mais facilitada pela planta,
entretanto a simbiose também beneficia a absorção dos demais nutrientes como
nitrogênio, o potássio, enxofre, cálcio e magnésio, transferindo-os para as plantas
através de estruturas fúngicas denominadas arbúsculos (BERBARA et al., 2006).
Em pesquisas envolvendo a inoculação de fungos micorrízicos arbusculares
(FMA’s), a utilização de espécies eficientes configura-se como um fator extremamente
essencial ao desenvolvimento de pesquisas de qualidade. Um bom exemplo a ser
dado é a relação entre a colonização desses microrganismos com o teor de P no solo.
Para viabilizar o uso das micorrizas em pesquisas, é essencial possuir
isolados fúngicos eficientes. O desempenho de isolados selecionados para inoculação
deve ser avaliado sob diferentes níveis de fertilidade de P, por exemplo, pois esse
fator influencia o grau de colonização e o benefício da planta hospedeira. Quanto mais
ampla a faixa de P no solo em que o fungo mantém a simbiose mutualista, maior é
9
sua eficiência e potencial de uso. Essa eficiência pode ser estimada relacionando-se
o benefício em crescimento que o isolado promove à planta (benefício micorrízico)
com aquele que a planta sem o fungo obtém da adição de fósforo (benefício de P)
(MOREIRA e SIQUEIRA, 2006). E é baseado nessas informações que pesquisas
como esta devem ser planejadas, com o máximo de conhecimento sobre os fatores
que irão provocar efeitos sobre as variáveis analisadas, adquirindo informações e
materiais confiáveis para obtenção de bons resultados.
3.4
Dependência micorrizica
Uma simbiose será bem sucedida sempre que houver combinações
satisfatórias entre fungo-planta-ambiente, dessa forma fatores ambientais (nível de
fósforo, conteúdo de água, pH, salinidade, temperatura, qualidade e intensidade
luminosa) acabam por exercer importante influência sob associações micorrizicas,
muitas vezes sendo a condição necessária para a compatibilidade da simbiose
(MEHROTRA, 1998).
A dependência micorrizica pode ser definida como sendo o grau no qual uma
determinada espécie vegetal depende da ação dos fungos micorrízicos para atingir o
seu máximo crescimento em um dado nível de fertilidade do solo (GERDEMANN,
1975). Declerck et al. (1995) afirmaram que este fenômeno é influenciado por fatores
genéticos relacionados a planta e ao fungo, entretanto pode ser influenciado por
fatores ambientais, principalmente pela concentração de fosforo presente no solo.
Diferentes espécies vegetais podem comportar-se de forma diferenciada em
relação aos fungos micorrizicos arbusculares. Dessa forma elas podem ser agrupadas
em micotróficas obrigatórias quando não sobrevivem sem micorriza até a idade
reprodutiva); micotróficas facultativas quando são beneficiadas pela micorrização em
condições de baixa fertilidade, e estando não micorrizadas conseguem atingir a idade
reprodutiva; e não micotróficas quando sobrevivem sem micorriza até a maturidade
reprodutiva (JANOS, 1980).
A obtenção dos valores de dependência micorrizica das plantas foi sugerido
por Menge et al. (1978), onde esses valores corresponderiam a massa da matéria
seca de plantas micorrizadas e não micorrizadas expressa em % (DM = massa seca
da planta micorrizada/massa seca da planta não micorrizada x 100). Porém, essa
metodologia acarretou em porcentagens muito elevadas, como em limoeiro (923%) e
10
laranjeira (723%). Plenchette et al. (1983), propôs alterações na metodologia
preconizada por Menge et al. (1978), que foi denominada dependência micorrizica
relativa. Esta pode ser obtida pela diferença entre a massa seca da matéria seca de
plantas micorrizadas e não micorrizadas em relação à massa da matéria seca de
plantas micorrizadas (DMR = massa da matéria seca das plantas micorrizadas massa da matéria seca das plantas não micorrizadas/massa da matéria seca das
plantas micorrizadas x 100).
Habte e Manjunath (1991) afirmaram que a dependência micorrizica pode ser
melhor compreendida quando se estabelece um gradiente de fertilidade no solo.
Dessa forma, baseando-se no que foi proposto por Plenchette et al. (1983)
estabeleceram níveis categóricos, podendo uma planta ser classificada como:
extremamente dependentes quando as espécies apresentam DM ≥ 75%; altamente
dependentes para espécies com DM entre 50% a 75%; moderadamente dependentes
para espécies com DM variando de 25 a 50%; marginalmente dependentes - espécies
com valores de DM inferiores a 25%; e plantas independentes -espécies que não são
colonizadas por FMA ou que não respondem à inoculação.
Brandão (2003) avaliou o grau de dependência micorrizica da gravioleira,
observando que a DM foi influenciada pelas concentrações de fósforo no solo e pelas
espécies de FMA associadas a planta. A gravioleira apresentou redução nos valores
de DM com o aumento nas doses de P no solo, entretanto as mudas foram
consideradas altamente dependentes, mesmo na dose mais elevada de P. Esse
comportamento pode significar uma determinada obrigatoriedade na presença de
FMAs no desenvolvimento da espécie.
3.5
Produção de glomalina por fungos micorrízicos arbusculares
A agregação de um solo é definida como sendo um dinâmico e complexo
processo que é influenciado por inúmeros fatores. Dentre esses fatores podem-se
incluir os componentes ambientais, manejo ao qual o solo é submetido, efeitos da
comunidade vegetal e de forma mais significativa pelas propriedades intrínsecas de
cada solo (BORIE et al., 2006).
Os microrganismos do solo desempenham importante funções na estabilidade
e estruturação destes. Um bom exemplo a citar são as hifas de fungos, em especial
os fungos micorrízicos, que tem como característica a liberação de polissacarídeos e
11
outros compostos orgânicos cimentantes, unindo a partículas presentes no solo,
auxiliando de maneira significativa no processo de formação de agregados
(GONZÁLES-CHAVEZ et al., 2004; BORIE et al., 2006).
Dois
importantes
mecanismos
relacionados
aos
fungos
micorrízicos
influenciam na formação de agregados, um físico, que é possível graças a ação de
hifas extra-radiculares que envolvem partículas minerais e orgânicas no solo e um
outro mecanismo de característica quelante, que ocorre devido a ação de uma
proteína produzida pelas micorrizas denominada glomalina (RILLIG, 2004; BERBARA
et al., 2006; PURIN e RILLIG, 2007).
A glomalina é uma glicoproteína que tem como importante característica ser
imunorreativa e recalcitrante. É parte constituinte da parede celular das hifas de
FMA’s, acumulando-se no solo após a decomposição por microrganismos do solo
(RAMOS e MARTINS, 2010).
Estudos anteriores revelaram uma estreita relação desta glicoproteína com a
estabilidade de agregados assim como com o estoque de carbono no solo, sendo isso
relacionado à sua produção abundante, elevada capacidade de aderir às partículas
do solo, por ser recalcitrante e por apresentar características hidrofóbicas (BÁEZPERÉZ et al., 2010; SOUSA et al., 2011).
Wright e Upadhyaya (1996), baseando-se na correlação observada de
concentrações de GPRS (glomalin-related soil proteins – Proteina do solo relacionada
à glomalina) com a estabilidade de agregados em água afirmaram que esta é
produzida por FMA’s no solo, atuando na estruturação do solo. O simples fato dos
agregados serem cobertos pela glomalina produzida pelos FMA’s acarreta uma
diminuição no movimento da água no interior desses agregados, o que os torna mais
estáveis (MILLER e JASTROW, 2000).
Essa estabilidade também pode ser explicada pela redução do potencial de
pressão exercido pela água nos esporos internos, evitando assim a expansão das
argilas e consequentemente o rompimento desses agregados ocorrerão em menor
frequência (GONZÁLEZ-CHÁVES et al., 2004).
Outra importante função que vem sendo atribuída a glomalina é a
descontaminação de solos com elevadas concentrações de metais pesados
(GONZÁLEZ-CHÁVES et al., 2004). Cornejo et al. (2008) avaliaram que o sequestro
desses metais pode ocorre como um mecanismo altamente eficiente dos FMA na
busca de melhores condições para seu desenvolvimento, estabilizando esses
12
elementos danosos as plantas, microrganismos e ao homem. Amostras de solo
contendo elevadas concentrações de Pb e Zn foram avaliadas e apresentaram 15,5%
e 20,5% do total desses metais estavam ligados a molécula de glomalina (VODNIK et
al., 2008). Cornejo et al. (2008) também analisaram os teores de Cu e Zn em solos
contaminados e observaram que 27% e 5,8% estavam associados a alguma fração
da glomalina, respectivamente.
13
4. MATERIAL E MÉTODOS
O experimento foi conduzido em casa de vegetação na Universidade Estadual
de Santa Cruz, Ilhéus – BA, durante o período de maio a setembro de 2014.
4.1
Preparo do solo
O solo utilizado foi retirado do horizonte superficial de um latossolo amarelo
distrófico típico, do município de lhéus, em uma área de pastagem. O solo foi seco
ao ar e peneirado em malha de 5 mm de espessura e posteriormente amostras foram
enviadas para análise química e granulométrica. O solo apresentou as seguintes
características químicas: pH em H2O = 4,7; matéria orgânica (MO) = 2,6 dag kg-1; P
Mehlich 1 = 7 mg dm-3; K = 36 mg dm-3; Ca = 0,6 cmolc dm-3 Mg = 0,3 cmolc dm-3; Al
= 1,2 cmolc dm-3; SB = 1 cmolc dm-3; CTC (t) = 2,2 cmolc dm-3; CTC (T) = 9,4 cmolc
dm-3; Cu, Fe, Zn, Mn e B (mg dm-3) = 0,8; 183; 1,6; 46; 0,6; V e m (%) = 10,6; 55. Os
resultados da análise granulométrica do solo foram: areia grossa = 12,26; areia média
= 233,66 g kg-1; areia fina = 379,8 g kg-1; areia mais fina = 112,81 g kg-1; areia total =
740,53 g kg-1; silte = 126,14 g kg-1, argila = 133,33 g kg-1 com textura franca - arenosa.
4.2
Avaliação da capacidade de campo
O teste foi realizado em um período de oito dias, utilizando-se 10 vasos com
furos no fundo, que receberam 3,5 kg de solo seco cada um. Os vasos foram imersos
em um vaso de maior capacidade para que a saturação do solo ocorresse de baixo
para cima para evitar a formação de bolsões de ar e garantir 100% de saturação. Em
seguida foram drenados e cobertos com papel isofilme para evitar a perda de água
por evaporação, e com isso afetar a determinação da capacidade de campo do solo.
Os vasos foram pesados diariamente até o momento em que os mesmos
apresentaram estabilidade em seu peso. O peso inicial foi de 3,5 Kg de solo seco, que
passou a apresentar no último dia uma média de 4,4 kg, ou seja, 0,9 kg a mais, que
corresponde a 100% de capacidade de campo. Durante o experimento, foi utilizada
60% dessa capacidade.
14
4.3
Curva de incubação do calcário
Foram utilizadas sete doses de calcário (0; 1; 2; 3; 4; 5; 6 t/ha) constando de
cinco repetições cada. Depois da aplicação do calcário dolomítico (PRNT=83%), o
solo foi umedecido de acordo com 60% da capacidade de campo. Os sacos contendo
0,5 dm3 de solo devidamente identificados, segundo as doses de calcário, foram
colocados em uma bancada na casa de vegetação. De oito em oito dias foram
agitados para proporcionar melhor mistura e ação do calcário no solo. Ao final de 45
dias, os sacos foram levados ao laboratório de Química e Fertilidade do solo da
Universidade Estadual de Santa Cruz para determinação de pH em água, segundo
metodologia da Embrapa (2011).
Foram utilizados 10 cm³ de solo em becker com capacidade de 50 mL,
devidamente identificados. Em seguida foram adicionados 25 mL de água destilada e,
com auxílio de um bastão de vidro as amostras foram homogeneizadas e deixadas
em repouso por uma hora. Passado esse tempo as amostras foram mais uma vez
homogeneizadas e procedeu-se a leitura utilizando pHmetro. De posse dos dados
ajustou-se uma curva por meio de análise de regressão linear, sendo esta
posteriormente derivada para obtenção da quantidade de calcário necessário para
obtenção do pH 6. A dose de calcário obtida foi de 1,3 g dm-3 de solo.
4.4
Preparo do composto orgânico
O composto orgânico utilizado no experimento foi produzido na fazenda
Sucupira, localizada no município de Ilhéus e que adota um sistema de cultivo
baseada na agricultura orgânica biodinâmica. O composto foi produzido a partir da
casca de frutos de cacau triturada e enriquecida com fosfato natural (fonte de P). Aos
120 dias após ter sido montada a compostagem o material foi coletado e levado a
Universidade Estadual de Santa Cruz, onde posteriormente foi seco a temperatura
ambiente e em seguida peneirado. A fim de avaliar a composição química do
composto orgânico uma amostra foi enviada ao Laboratório de Análise Agronômica
FULLIN, localizado em Linhares (ES). Os resultados foram: pH em CaCl2 = 7,41; P,
K, Ca, Mg e S (g kg-1) = 52,1; 24,1; 72,4; 6,4 e 2,7; Cu, Mn, Zn, Fe e B (mg kg-1) =
57; 185; 195; 7656 e 16 respectivamente.
15
4.5
Obtenção de inóculos de fungos micorrízicos arbusculares
Os inóculos de duas espécies de fungos micorrízicos arbusculares foram
obtidos junto a EMBRAPA Agrobiologia, localizada no município de Seropédica, Rio
de Janeiro. Informações referentes à densidade de esporos presente no solo inoculo
de cada espécie de FMA foram documentadas. A espécie Entrophospora colombiana
contendo 6 esporos/g de solo; e a espécie Acaulospora scrobiculata com uma
densidade de 16 esporos/g de solo inóculo. Ressalta-se que esses valores foram
considerados para determinar a quantidade necessária de cada inóculo para a
inoculação de 30 esporos, bem como a proporção de cada inóculo para os tratamentos
que receberam inóculos das duas espécies de fungos micorrízicos.
4.6
Estabelecimento das doses do composto orgânico
As doses do composto orgânico utilizado no experimento foram calculadas
levando em consideração a análise química do composto previamente realizada. As
doses foram decididas levando em consideração o nutriente K, por este ser dentre os
nutrientes o mais extraído pela gravioleira e está presente em grande quantidade no
composto. A dose central de K adotada foi a de 200 mg dm -3 de solo, o que
correspondeu a uma dose do composto orgânico de 10 g dm -3 de solo. As cinco doses
determinadas para construção da curva padrão, para identificar a melhor dose como
resultado final, foram de 0, 5, 10, 20 e 30 g dm-3 do composto orgânico.
4.7
Condução do experimento
O experimento foi constituído por um fatorial 5 x 4 com 5 doses do composto
orgânico a base de casca de cacau (0; 5; 10 ;20; 30 g dm-3) e 4 tratamentos
micorrizicos (controle não inoculado, inoculação com Acaulospora scrobiculata, com
Entrophospora colombiana e a mistura desses inóculos) com um delineamento em
blocos casualizados, constando de quatro blocos. Cada unidade experimental foi
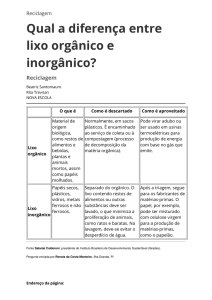
constituída por um vaso contendo uma única planta (Figura 1). Foi aplicado calcário
dolomítico 45 dias antes do plantio das mudas, com o intuito de permitir a correção do
pH do solo para valores desejados para a gravioleira, conforme método de incubação
que foi realizado anteriormente. Sementes de graviola tipo Morada foram coletadas
de frutos de uma mesma planta para obtenção de mudas, que 30 dias após
germinarem foram transplantadas para vasos com capacidade de 3,5 dm3.
16
As plantas foram irrigadas posteriormente com água destilada autoclavada,
sendo ainda realizado mensurações mensais do diâmetro do coleto e altura das
plantas.
Figura 1. Vista parcial do experimento com mudas de gravioleira em solo com diferentes
doses de composto orgânico e inoculadas com E. colombiana, A. scrobiculata e a mistura
desses dois fungos micorrízicos, e não inoculadas (controle).
4.8
Avaliação da massa seca
Ao final do experimento, as plantas foram coletadas e separadas em parte
aérea (folha e caule) e sistema radicular, embaladas em sacos de papel e secas a
70°C, em estufa de circulação forçada até peso constante.
4.9
Dependência micorrizica
A dependência micorrízica (DM) em cada dose do composto orgânico foi
calculada seguindo metodologia proposta por Plenchette et al. (1983), baseando-se
nos valores de biomassa seca das plantas micorrizadas. Utilizando-se das categorias
descritas por Habte e Manjunath (1991), que classifica as plantas como
excessivamente dependente (DM > 75%), altamente dependente (DM entre 50 a
75%), moderadamente dependente (DM entre 25 e 50%), marginalmente
dependentes (DM< 25%) ou independentes (sem resposta a micorrização).
17
4.10 Teor de nutrientes na folha
Após a secagem das folhas a 70°C até o peso constante, as amostras foram
moídas em moinho do tipo Wiley, para serem analisadas com relação aos teores de
macronutrientes (N, K, P, Ca e Mg) e micronutrientes (Fe, Zn, Cu e Mn) nas folhas de
mudas de gravioleira. Para determinação do teor foram utilizadas como folhas
diagnóstico aquelas presentes na parte mediana da planta. As concentrações dos
nutrientes foram determinadas por via úmida segundo metodologia EMBRAPA (2009).
Os teores de P foram determinados pelo método do molibidato em espectrofotômetro
de absorção molecular, K por fotometria de chama; Ca, Mg, Cu, Fe, Mn e Zn por
espectrometria de absorção atômica, e N por destilação e titulação pelo método micro
Kjeldahl (EMBRAPA, 2009).
4.11 Coloração de raízes micorrizadas
Para avaliar a colonização micorrízica foram selecionadas as raízes mais finas
de mudas de gravioleira, que geralmente são as mais densamente colonizadas pelos
FMA. As raízes foram lavadas em peneira com água de torneira para remover todas
as partículas do substrato. Após a lavagem, as raízes foram armazenadas em
recipientes de vidro contendo etanol 50% para sua conservação. Em seguida as
raízes foram clarificadas com KOH a 10 % e coradas em lactoglicerol com azul de
tripano.
Para estimativa da porcentagem de colonização micorrízica nas amostras das
raízes das plantas foi utilizada a técnica de clareamento em KOH a 10% e coloração
com azul de tripano, conforme metodologia descrita por Phillips e Hayman (1970).
As raízes foram segmentadas em 2 cm e mergulhadas em solução de KOH
10% em tubos de ensaio devidamente identificados e em seguida, colocadas em
banho-maria (60oC) por aproximadamente 5 min. Após o término do tempo as raízes
passaram por três lavagens e adicionou-se aos tubos de ensaio contendo as raízes,
solução de H2O2 10% por 5 min. Após esta etapa, as raízes passaram por mais 3
lavagens e foi adicionado aos mesmos tubos a solução ácida (HCl 5%) ficando em
banho-maria durante 3 minutos. Depois, foi retirado o HCl 5% e adicionado à solução
de azul de tripano (0,05%) aos tubos, os quais foram colocados em banho-maria a
60°c de temperatura por um período de 5 minutos e em seguida submetidas a uma
tríplice lavagem. Finalmente as raízes foram conservadas em solução de lactoglicerol
18
(ácido lático, glicerol e água destilada na proporção 1:1) e armazenadas em tubos
identificados para posterior análise.
4.12 Quantificação de raízes micorrizadas
A quantificação da colonização foi realizada pelo método de intersecção de
Mcgonigle et al. (1990). Os fragmentos de raízes foram montados em lâminas riscadas
com 10 linhas transversais (com 0,5 mm de distância de uma linha para outra), e
observados em microscópio óptico em lente de aumento de 40 X. A percentagem de
colonização dos segmentos foi baseada na razão do número intersecções de raízes
colonizadas pelo número total intersecções de segmentos observados, sendo o
resultado multiplicado por 100.
4.13 Teor de Gomalina facilmente extraível
A extração da glomalina facilmente extraível (GFE) foi realizada conforme
metodologia de Wright e Upadhyaya (1996) e quantificada pela técnica de
microtitulação de Bradford (1976).
Para quantificação da GFE foram utilizadas amostras de 2 g de agregados do
solo em erlenmeyers, onde foram adicionados 8 mL do tampão citrato de sódio 20 mM
(pH 7). Posteriormente as amostras foram submetidas a autoclavagem (121°C) por 30
minutos e em seguida foram transferidas para tubos específicos para ultra centrífuga
e centrifugadas a 10.000 RPM por 5 minutos. O sobrenadante foi pipetado e
armazenado em Eppendorfs e conservados à 4 °C até o processo de microtitulação e
leitura.
Para a microtitulação da proteína, utilizou-se placas tipo ELISA com 96 poços,
onde primeiramente foi estabelecida a curva padrão com o uso de soro albumina
bovina (BSA) como proteína purificada (PURIN, 2005), solução tampão citrato de
sódio 20 mM e reagente Bradford (BRADFORD, 1976).
Para a microtitulação da quantidade de glomalina no solo foram adicionados
em cada poço 5 uL dos extratos obtidos das amostras; 45 uL de água destilada e 50
uL do reagente de Bradford. As placas foram levadas a Espectrofotômetro
Spectramax Paradigm (Molecular devices) para efetuar a leitura a uma diluição de 0,2
e uma absorbância de 595 nm. Os resultados foram expressos em mg mL-1 e
posteriormente transformados em mg g-1 de solo.
19
4.14
Extração e contagem
arbusculares
dos
esporos
de
fungos
micorrízicos
Para quantificar o número de esporos de fungos micorrízicos arbusculares foi
utilizada a metodologia de peneiramento úmido de Gerdemann e Nicolson (1963)
modificado, onde amostras de solos foram coletadas ao final do experimento e
armazenadas em refrigerador para evitar a germinação dos esporos de FMA.
Posteriormente foram pesados 50 g de cada amostra que após terem sido peneiradas
foram centrifugadas por duas vezes. A primeira centrifugação foi realizada utilizando
água destilada por três minutos a uma velocidade de 2500 rpm e em seguida as
amostras foram novamente centrifugadas utilizando solução de sacarose 50% por um
minuto a uma velocidade de 2500 rpm. O sobrenadante formado foi peneirado e as
amostras contendo os esporos foram armazenadas em freezer para posterior
quantificação utilizando esteromicrocopio no aumento de 40X.
4.15 Analise estatística
Os dados foram submetidos à análise de variância a 5 % de probabilidade. Foi
realizado teste de Tukey a 5 % de probabilidade para o fator qualitativo e regressão
linear para o fator quantitativo. Foram geradas equações para observar como cada
nível do fator qualitativo (FMA) se comportou dentro de cada dose do fator quantitativo
(Composto orgânico). As analises foram realizadas nos pacotes estatísticos R
(FERREIRA et al., 2011) e STATISTICA 8.0 (STATSOFT, 2002).
20
5. RESULTADOS
5.1
Altura e diâmetro do coleto das plantas
Houve interação significativa entre os tratamentos micorrízicos aplicados e as
doses do composto orgânico incorporadas ao solo (p<0,05) para as variáveis altura e
diâmetro do coleto das mudas de gravioleira (Tabela 1).
Tabela 1. Resumo da Análise de Variância para altura e diâmetro do coleto das mudas de
gravioleira cultivadas em solo com diferentes doses de composto orgânico e inoculadas com
E. colombiana, A. scrobiculata e a mistura desses dois fungos micorrízicos
Quadrado Médio
FV
GL
Bloco
FMA
Composto
FMA*Composto
Erro
CV (%)
3
3
4
12
57
Altura
Diâmetro do coleto
96,43
577,18*
42,99
122,52*
57,73
26,31
0,1591
6,3512*
3,6059*
0,9290*
0,2307
8,88
*Significativo a 5% de probabilidade pelo teste de F.
Os valores médios de altura e diâmetro do coleto das mudas de gravioleira
inoculadas com FMAs e submetidas a doses de composto orgânico, podem ser
observados na Tabela 2.
Tabela 2. Valores médios de altura e diâmetro do coleto de mudas de gravioleira cultivadas
em solo com diferentes doses de composto orgânico e inoculadas com E. colombiana, A.
scrobiculata e a mistura desses dois fungos micorrízicos (MIX), e não inoculadas (controle)
Altura (cm)
Tratamento micorrízico
Controle
Acaulospora scrobiculata
Entrophospora colombiana
MIX
0
15,5 b
17,5 ab
20,6 a
18,2 ab
Doses composto (g dm-3)
5
10
20
15,9 b 16,3 c 16,8 b
19,7 ab 19,7 bc 27,3 a
21,0 a 23,4 ab 26,0 a
19,9 ab 26,8 a 27,3 a
Diâmetro do coleto (mm)
Doses composto (g dm-3)
Tratamento micorrízico
0
5
10
20
Controle
4,1 c
4,4 b
4,5 c
4,6 b
Acaulospora scrobiculata
4,7 bc 5,0 ab
5,2 bc 6,3 a
Entrophospora colombiana
5,0 ab 5,7 a
5,9 b
6,1 a
MIX
5,6 a
5,4 a
6,8 a
6,6 a
30
18,5 c
24,2 b
18,7 c
29,1 a
30
5,0 c
6,1 ab
5,6 bc
6,6 a
Médias seguidas das mesmas letras nas colunas não diferem estatisticamente pelo teste de Tukey a
5% de probabilidade.
21
Em todas as doses do composto orgânico aplicadas ao tratamento não
micorrizado (plantas controle), a altura e diâmetro do coleto apresentaram valores
inferiores aos das plantas inoculadas com micorrizas. Nas doses iniciais do composto
orgânico (0 e 5 g dm-3) as plantas submetidas ao tratamento micorrízico com
Entrophospora colombiana apresentaram os maiores valores para altura (Tabela 2),
mas este não foi estatisticamente superior aos do tratamento inoculado com
Acaulospora scrobiculata e a mistura dos fungos micorrízicos (MIX).
Nas maiores doses do composto (10; 20 e 30 g dm -3) o tratamento MIX foi o
que apresentou os maiores valores de altura, sendo estatisticamente superior aos
tratamentos controle e Acaulospora scrobiculata na dose central (10 g dm-3). O
tratamento MIX foi estatisticamente superior aos demais na maior dose do composto
orgânico (30 g dm-3).
5.2
Produção de matéria seca
Houve interação entre as doses do composto orgânico e os FMAs inoculados
em gravioleira para a produção de matéria seca total das plantas, conforme pode ser
observado na Tabela 3.
Tabela 3. Resumo da Análise de Variância para matéria seca total das mudas de gravioleira
cultivadas em solo com diferentes doses de composto orgânico e inoculadas com E.
colombiana, A. scrobiculata e a mistura desses dois fungos micorrízicos
FV
GL
Quadrado Médio
Bloco
FMA
Composto
FMA*Composto
3
3
4
4
1,846
135,624*
27,362*
6,382*
Erro
57
1,297
CV (%)
18,8
*Significativo a 5% de probabilidade pelo teste de F.
Os valores médios de matéria seca total das mudas de gravioleira submetidas
a doses de composto orgânico (0; 5; 10; 20 e 30 g dm -3) e inoculadas com FMAs,
podem ser observados na tabela 4.
22
Tabela 4. Valores médios de matéria seca total (MST) de mudas de gravioleira cultivadas em
solo com diferentes doses de composto orgânico e inoculadas com E. colombiana, A.
scrobiculata e a mistura desses dois fungos micorrízicos (MIX), e não inoculadas (controle)
Tratamento micorrizico
Controle
Acaulospora scrobiculata
Entrophospora colombiana
MIX
0
2,20 b
6,09 b
6,69 a
6,68 a
Doses composto (g dm-3)
5
10
20
2,68 c
2,91 c
3,70 b
6,44 b
7,30 b
8,17 a
7,67 a
9,58 a
7,92 a
7,71 a
9,81 a
8,48 a
30
3,94 b
7,97 a
6,82 a
8,19 a
Médias seguidas das mesmas letras nas colunas não diferem estatisticamente pelo teste de Tukey a
5% de probabilidade.
A análise estatística apresentou diferenças entre os tratamentos micorrizícos
em todas as doses de fertilização orgânica aplicadas ao solo. Na ausência de
adubação, os maiores valores de matéria seca foram das plantas inoculadas com E.
colombiana, sendo iguais estatisticamente ao tratamento MIX e superiores aos
demais. Nas plantas fertilizadas, independente da dose aplicada o tratamento MIX
obteve a maior produção de matéria seca, igualando-se estatisticamente a E.
colombiana nas doses 5 e 10 g dm-3. Nas duas doses subsequentes não houve
diferença estatística entre os tratamentos com inoculação de FMAs.
14
Materia seca total (g)
12
Controle Y** = 1,0963+0,0819*x R² = 0,88
E. colombiana Y** = 6,4913+0,2712*x-0,0086*x^2 R² = 0,71
A. scrobiculata Y** = 5,6389+0,1141*x R² = 0,91
MIX Y** = 3,6591+0,6534*x-0,0167*x^2 R² = 0,80
10
8
6
4
2
0
0
5
10
20
30
Dose do composto orgânico (g dm-3)
Figura 2. Gráfico da análise de regressão para a média da massa seca total das mudas de
gravioleira submetido a diferentes doses de composto orgânico e inoculado com
Entrophospora colombiana, Acaulospora scrobiculata e a mistura desses dois fungos
micorrízicos (MIX), e não inoculados (controle).
23
As plantas que receberam a mistura dos inóculos E. colombiana e A.
scrobiculata (MIX) apresentaram um comportamento quadrático. As equações obtidas
pela regressão foram derivadas para obter a dose do composto orgânico onde a
produção de biomassa seca será máxima. A dose ajustada para E. colombiana e Mix
foram 15,67 g dm-3 e 19,56 g dm-3 respectivamente. O tratamento A. scrobiculata,
assim como o tratamento controle apresentaram linearidade no modelo matemático
ajustado, tendo os maiores valores de matéria seca total na maior dose de composto
orgânico (30 g dm-3).
5.3
Dependência micorrízica
Houve interação entre as doses do composto orgânico e os FMAs inoculados
em mudas de gravioleira para a análise da dependência micorrizica da planta,
conforme pode ser observado na Tabela 5.
Tabela 5. Resumo da Análise de Variância para dependência micorrízica de mudas de
gravioleira cultivadas em solo com diferentes doses de composto orgânico e inoculados com
E. colombiana, A. scrobiculata e a mistura desses dois fungos micorrízicos
FV
GL
Quadrado Médio
Bloco
FMA
3
3
7
33260*
Composto
4
388*
FMA*Composto
4
69*
Erro
37
5
CV (%)
3,77
*Significativo a 5% de probabilidade pelo teste de F.
Os valores médios de dependência micorrízica das mudas de gravioleira,
inoculadas com FMAs e submetidas a doses de composto são apresentados na
Tabela 6.
24
Tabela 6. Valores médios de dependência micorrizica das mudas de gravioleira submetidas
diferentes doses de composto orgânico e inoculadas com E. colombiana, A. scrobiculata e a
mistura desses dois fungos micorrízicos (MIX)
Tratamento micorrízico
Acaulospora scrobiculata
Entrophospora colombiana
MIX
0
84,08 a
87,69 a
84,89 a
Doses composto (g dm-3)
5
10
20
84,34 a
82,96 b
78,33 a
87,82 a
86,26 ab
77,01 a
87,90 a
88,05 a
79,30 a
30
75,78 a
65,62 b
73,06 a
Médias seguidas das mesmas letras nas colunas não diferem estatisticamente pelo teste de Tukey a
5% de probabilidade.
Foram observadas diferenças estatísticas (p<0,05), na porcentagem de
dependência micorrízica. Na ausência de adubação foram obtidos os maiores valores
de dependência micorrízica em mudas de gravioleira, com redução proporcional ao
aumento da dose de composto orgânico incorporado ao solo. Esperavam-se reduções
nos valores nas doses mais elevadas de composto orgânico, entretanto os valores de
dependência micorrizica foram elevados, inclusive na dose mais elevada (Tabela 6).
Tabela 7. Níveis categóricos (segundo HABTE e MANJUNATH, 1991) de dependência
micorrízica de gravioleiras submetidas a diferentes doses de composto orgânico e inoculadas
com E. colombiana, A. scrobiculata e a mistura desses dois fungos (MIX)
Tratamento micorrizico
Acaulospora scrobiculata
Entrophospora colombiana
MIX
0
ED
ED
ED
Doses composto (g dm-3)
5
10
20
ED
ED
ED
ED
ED
ED
ED
ED
ED
30
AD
AD
ED
ED: Excessivamente dependente; AD: Altamente dependente
De acordo com os níveis categóricos (Tabela 7), a gravioleira foi classificada
quanto à dependência micorrízica, onde pelos resultados apresentados foi possível
observar que a espécie Annona muricata apresentou um elevado grau de
dependência micorrízica em todas as condições de fertilidade do solo apresentadas
no experimento, sendo quase sempre excessivamente dependente a simbiose, com
exceção dos tratamentos E. colombiana e A. scrobiculata na dose mais elevada do
composto orgânico (30 g dm-3), onde a gravioleira foi classificada como altamente
dependente segundo os níveis categóricos de HABTE e MANJUNATH (1991).
25
5.4
Grau de colonização micorrizica
Houve interação entre as doses do composto orgânico e os FMAs inoculados
em gravioleira para a colonização micorrízica das raízes, conforme pode ser
observado na Tabela 8.
Tabela 8. Resumo da Análise de Variância para porcentagem de colonização micorrízica de
mudas de gravioleira cultivadas em solo com diferentes doses de composto orgânico e
inoculadas com E. colombiana, A. scrobiculata e a mistura desses dois fungos micorrízicos
FV
GL
Quadrado Médio
Bloco
FMA
3
3
118,4*
23677,7*
Composto
4
1874,8*
FMA*Composto
4
251,5*
Erro
57
281,7
CV (%)
11,13
*Significativo a 5% de probabilidade pelo teste de F.
Os valores médios da porcentagem de colonização micorrízica e da presença
de estruturas nas raízes de mudas de gravioleira, inoculadas com FMAs e submetidas
a doses de composto orgânico (0; 5; 10; 20 e 30 g dm-3), encontram-se expostos na
Tabela 9.
Tabela 9. Valores médios do grau de colonização micorrízica de mudas de gravioleira
cultivadas em solo sob diferentes doses de composto orgânico e inoculadas com E.
colombiana, A. scrobiculata e a mistura desses dois fungos micorrízicos (MIX), e não
inoculadas (plantas controle)
Tratamento micorrizico
Controle
Acaulospora scrobiculata
Entrophospora colombiana
MIX
0
00,00 c
59,34 a
40,27 b
57,82 a
Doses composto (g dm-3)
5
10
20
00,00 c
00,00 b
00,00 b
69,92 ab
81,03 a
75,25 a
63,46 b
87,29 a
78,71 a
74,99 a
89,40 a
82,82 a
30
00,00 b
52,34 a
55,11 a
59,63 a
Médias seguidas das mesmas letras nas colunas não diferem estatisticamente pelo teste de Tukey a
5% de probabilidade.
Na ausência do composto orgânico e com a adição de 5 g dm -3 de composto
orgânico foi observado que a porcentagem de colonização das raízes das plantas
inoculadas com Acaulospora scrobiculata e com a mistura dos dois FMAs foram
estatisticamente superiores ao das plantas inoculadas com Entrophospora
colombiana. Nas demais doses do composto não houve diferença estatística entre os
tratamentos, exceto ao tratamento controle (não micorrizado). Vale destaca-se que a
26
taxa de colonização micorrízica nas plantas do tratamento MIX apresentou os maiores
valores.
100
Colonização micorrizica (%)
80
60
40
Controle Y = Não houve ajuste
E. colomb iana Y** = 41,4060+5,6577*x-0,1753*x^2 R² = 0,93
A. scrob iculata Y** = 59,0158+3,0789*x-0,1104*x^2 R² = 0,98
MIX Y** = 58,5815+4,0966*x-0,1365*x^2 R² = 0,96
20
0
0
5
10
20
30
Dose do composto orgânico (g dm-3)
Figura 3. Gráfico da análise de regressão para o grau de colonização micorrízica da
gravioleira submetida a diferentes doses de composto orgânico e inoculada com
Entrophospora colombiana, Acaulospora scrobiculata e a mistura desses dois fungos
micorrízicos (MIX), e não plantas não inoculadas (controle).
Os modelos matemáticos significativos (p<0,05) que mais se ajustaram a
variável grau de colonização micorrizica foram quadráticos (Figura 3). Nos
tratamentos micorrízicos Acaulospora scrobiculata, Entrophospora colombiana e MIX
observou-se um comportamento semelhante à medida em que se aumentou a dose
do composto orgânico. A colonização micorrízica na ausência do composto orgânico
variou de 40 a 60%, e apresentou aumento até a dose central do composto orgânico
(10 g dm-3), com médias variando entre 80 e 90%, seguidas de redução na
colonização.
Pelo fato dos dados terem ajustados modelos matemáticos quadráticos foi
possível inferir a dose do composto orgânico que propiciou a maior colonização
radicular para cada tratamento de FMA. Dessa forma os tratamentos Entrophospora
colombiana, Acaulospora scrobiculata, e MIX apresentam sua máxima colonização
27
nas doses 16,14 g dm-3; 13,94 g dm-3 e 15,00 g dm-3 do composto orgânico,
respectivamente.
Morfologicamente as micorrizas da gravioleira observadas no presente
trabalho (Figura 4), apresentaram semelhança aos da série Arun. Micorrizas inclusas
nessa serie apresentam arbúsculos bem desenvolvidos, os quais são as estruturas
onde ocorre a troca de nutrientes entre o fungo e a planta (SMITH e READ, 1997).
28
A
B
C
D
E
F
Figura 4. Raízes de gravioleira dos tratamentos controle e inoculado com A. scrobiculata,
Entrophospora colombiana e a mistura desses fungos micorrízicos arbusculares (FMAs)
submetida a diferentes doses de composto orgânico. A. Vista geral de uma raiz não
micorrizada do tratamento controle; B. Vista geral de uma raiz micorrizada com E. colombiana
mostrando a frequência dos arbúsculos (setas); C. Raiz micorrizada do tratamento com A.
scrobiculata, onde pode ser observados arbúsculos conspícuos (setas); D. Segmento de raiz
do tratamento inoculado com a mistura dos FMAs mostrando os arbúsculos (seta); E. Detalhe
das hifas extraradiculares (seta) e dos tricomas da raiz (ponta de seta); F. Detalhe dos
arbúsculos (setas) de A. scrobiculata no interior das células corticais.
29
5.5
Densidade de esporos
Houve interação entre as doses do composto orgânico e os FMAs inoculados
em mudas de gravioleira para a produção de esporos de FMA em 50g de solo
rizosférico, conforme pode ser observado na Tabela 10.
Tabela 10. Resumo da Análise de Variância para densidade de esporos em 50 g de solo
rizosférico de gravioleiras cultivadas em solo com diferentes doses de composto orgânico e
inoculadas com E. colombiana, A. scrobiculata e a mistura desses dois fungos micorrízicos
FV
GL
Quadrado Médio
Bloco
FMA
3
3
293
845968*
Composto
4
227956*
FMA*Composto
4
29712*
Erro
57
1109
CV (%)
10,53
*Significativo a 5% de probabilidade pelo teste de F.
Os valores médios para o número de esporos em 50 g de solo rizosférico onde
foram cultivadas mudas de gravioleira inoculadas com FMAs e submetidas a doses
de composto orgânico podem ser observados na Tabela 11.
Tabela 11. Valores médios do número de esporos no solo após o plantio com mudas de
gravioleira inoculadas com E. colombiana, A. scrobiculata e a mistura desses dois fungos
micorrízicos (Mix), e não inoculadas (controle) submetidas a diferentes doses de composto
orgânico.
Doses composto (g dm-3)
Tratamento micorrízico
0
5
10
20
30
Nº médio esporos/50 g de solo rizosférico
Controle
Acaulospora scrobiculata
Entrophospora colombiana
MIX
17,0 d
219,7 b
112,0 c
302,2 a
14,5 c
360,7 a
254,0 b
410,7 a
16,2 c
585,7 b
576,7 b
689,5 a
17,5 c
495,5 b
492,2 b
594,5 a
16,50 b
361,2 a
392,2 a
399,2 a
Médias seguidas das mesmas letras nas colunas não diferem estatisticamente pelo teste de Tukey a
5% de probabilidade.
Diferenças estatísticas foram observadas na esporulação de Acaulospora
scrobiculata, Entrophospora colombiana e a mistura de ambos FMAs (MIX).
Independente da dose do composto orgânico aplicada, o tratamento com a presença
das duas espécies de fungos micorrizícos (MIX) apresentou os maiores valores, não
sendo estatisticamente superiores aos tratamentos Acaulospora scrobiculata,
30
Entrophospora colombiana nas doses 5 g dm-3 e 30 g dm-3 do composto orgânico,
apresentando na dose 5 g dm-3 do composto orgânico uma produção de esporos que
não diferiu estatisticamente ao tratamento Acaulospora scrobiculata, que apresentou
um incremento de 64% no número de esporos quando comparado a produção na
ausência do composto orgânico. Por outro lado, na dose de 30 g dm-3 do composto
orgânico foi observada redução no número de esporos em todos os tratamentos, não
havendo diferença estatística entre os tratamentos onde houve inoculação de FMAs.
O número médio de esporos encontrados no solo rizosférico das gravioleiras
submetidas a doses crescentes de composto orgânico e inoculadas com FMAs está
representado na Figura 5.
900
Densidade de esporos
800
Controle Y = Não houve ajuste
E. Colombina Y** = 105,8067 + 49,9806X - 1,3684X² R² = 0,85
A. scrobiculata Y** = 222,8488 + 40,9061X - 1,2281 ² R² = 0,87
MIX Y** = 287,2790 + 45,7348X - 1,4125X² R² = 0,86
700
600
500
400
300
200
100
0
0
5
10
20
30
-3
Dose composto orgânico (g dm )
Figura 5. Análise de regressão para o número médio de esporos no solo após o plantio com
mudas de gravioleira inoculadas com E. colombiana, A. scrobiculata e a mistura desses dois
fungos micorrízicos (Mix), e não inoculadas (plantas controle) submetidas a doses crescentes
de composto orgânico.
A aplicação de doses crescentes do composto orgânico resultou no incremento
no número de esporos para todos os inóculos testados. Os maiores valores em
esporulação para todos FMAs inoculados ocorreu na dose central do composto
orgânico (10 g dm-3). Modelos matemáticos com coeficientes significativos
(quadráticos) foram ajustados para todos FMAs testados, possibilitando obter a dose
onde cada fungo obtenha sua maior produção de esporos nessas condições de
31
fertilidade. A dose ótima para Acaulospora scrobiculata, Entrophospora colombiana e
a mistura de ambos (MIX) foi de 18,47 g dm-3, 16,65 g dm-3 e 15,91 g dm-3,
respectivamente.
5.6
Glomalina no solo
Houve interação entre as doses do composto orgânico e os FMAs inoculados
em gravioleira para o teor de glomalina facilmente disponível no solo após cultivo com
gravioleira inoculada com FMAs e submetida a doses crescentes de composto
orgânico, conforme pode ser observado na Tabela 12.
Tabela 12. Resumo da Análise de Variância para teor de glomalina no solo após o plantio
com mudas de gravioleira inoculadas com E. colombiana, A. scrobiculata e a mistura desses
dois fungos micorrízicos e submetida a doses crescentes de composto orgânico
FV
GL
Quadrado Médio
Bloco
FMA
3
3
7
33260*
Composto
4
388*
FMA*Composto
4
69*
Erro
57
5
CV (%)
3,77
*Significativo a 5% de probabilidade pelo teste de F.
Os valores médios do teor de glomalina facilmente disponível em 2 g de
agregados de solo com diâmetro < 2 mm podem ser observados na Tabela 13.
Tabela 13. Valores médios para a concentração de glomalina no solo após o plantio com
mudas de gravioleira inoculadas com E. colombiana, A. scrobiculata e a mistura desses dois
fungos micorrízicos (Mix), e não inoculada (controle) e submetida a diferentes doses de
composto orgânico
Tratamento micorrizico
Controle
Acaulospora scrobiculata
Entrophospora colombiana
MIX
0
13,94 b
17,25 a
17,27 a
17,26 a
Doses composto (g dm-3)
5
10
20
14,50 b
14,37 b
14,53 b
19,96 a
20,31 a
21,04 a
20,92 a
20,39 a
20,36 a
20,21 a
20,34 a
20,71 a
30
13,96 b
20,92 a
20,45 a
21,22 a
Médias seguidas das mesmas letras nas colunas não diferem estatisticamente pelo teste de Tukey a
5% de probabilidade.
Diferenças estatísticas para o fator qualitativo do experimento (FMA) foram
observadas em todas as doses do fator quantitativo (doses do composto orgânico).
As plantas dos tratamentos inoculados com E. Colombiana, A. scrobiculata e a mistura
32
dos dois FMAs não diferiram ente si, diferindo apenas dos valores encontrado para o
tratamento controle (não micorrizado).
A análise de regressão para o teor de glomalina nos agregados do solo
apresentou ajuste de modelos matemáticos para todos os tratamentos (Figura 6). A
partir das equações quadráticas ajustadas foi possível chegar à dose ideal em que
cada tratamento micorrízico produziu maiores concentrações de glomalina.
23
Glomalina Facilmente Extraivel ( mg g-1 solo )
22
21
20
19
18
17
16
15
14
13
Controle Y = não houve ajuste
E. Colombina Y** = 18,0561 + 0,3560X - 0,0104X² R² = 0,64
A. scrobiculata Y** = 17,6079 + 0,3906X - 0,0103X² R² = 0,92
MIX Y** = 17,7912 + 0,3507X - 0,0089X² R² = 0,83
12
11
10
0
5
10
20
30
-3
Dose do composto orgânico (g dm )
Figura 6. Análise de regressão para a concentração de glomalina no solo após o
plantio com mudas de gravioleira inoculadas com E. colombiana, A. scrobiculata e a mistura
desses dois fungos micorrízicos (Mix), e não inoculada (controle) submetida a diferentes
doses de composto orgânico.
As plantas inoculadas com E. Colombiana, dentre todos tratamentos, foram as
que apresentaram a menor dose ótima do composto orgânico (19,27 g dm -3), para a
produção do maior teor de glomalina no solo. As amostras de solo onde as plantas
foram cultivadas com a inoculação de A. scrobiculata apresentaram 21,36 g dm-3 como
a dose ótima de composto orgânico para a máxima concentração de glomalina no
solo. O teor dessa glicoproteína também foi avaliado para o tratamento onde as duas
espécies fúngicas foram proporcionalmente inoculadas (MIX), sendo que este
tratamento foi o que apresentou a maior dose (22,15 g dm-3) necessária do composto
orgânico para a máxima concentração de glomalina no solo.
33
5.7
Nutrientes na folha
5.7.1 Macronutrientes
Houve interação entre as doses do composto orgânico e os FMAs inoculados
em mudas de gravioleira para o teor de N, P, K, e Mg observados nas folhas (Tabela
14). O teor de Ca nas folhas não apresentou interação entre os fatores de variação,
entretanto cada fator apresentou significância separadamente. O teor desses
nutrientes foi significativamente influenciado pelo inóculo micorrízico, sendo que as
médias desses teores foram comparadas dentro de cada dose de composto orgânico.
Tabela 14. Resumo da análise de variância para o macronutrientes na folha da gravioleira
submetida a diferentes doses de composto orgânico e inoculada com Entrophospora colombiana,
Acaulospora scrobiculata e a mistura desses dois fungos micorrízicos (Mix), e não inoculada
(controle)
Quadrado Médio
FV
GL
N
P
K
Ca
Mg
Bloco
2
13,22
1,22
0,07
3,104
0,0147
FMA
3
717,4*
3,58*
110.94*
14,140* 13,5316*
Composto
4
148,48*
0,4586*
595,19*
201,616* 15,3514*
FMA*Composto
12
28,46*
0,29*
119,83*
3,587
0,3892*
Erro
37
4,71
0,669
0,85
2,504
0,1821
CV (%)
8,02
15,19
8,24
10,09
6,97
*Significativo a 5% de probabilidade pelo teste de F.
Os valores médios dos teores foliares de N, P, K, Ca e Mg nas folhas
diagnostico das gravioleiras inoculadas com FMAs e submetidas a doses crescentes
de composto orgânico (0; 5; 10; 20 e 30 g dm -3), encontram-se expostos na Tabela
15.
34
Tabela 15. Média do teor de macronutrientes nas folhas da gravioleira submetida a diferentes
doses de composto orgânico e inoculada com Entrophospora colombiana, Acaulospora
scrobiculata e a mistura desses dois fungos micorrízicos (Mix), e não inoculada (controle)
Nitrogênio (g kg-1)
Tratamento micorrízico
Controle
Acaulospora scrobiculata
Entrophospora colombiana
MIX
0
8,40 c
31,64 a
20,98 b
24,46 b
Doses composto (g dm-3)
5
10
20
12,22 b
18,34 c
22,00 b
32,26 a
34,41 a
31,35 a
29,08 a
28,69 b
30,09 a
31,22 a
31,18 ab
32,27 a
30
23,60 b
32,02 a
35,21 a
32,16 a
Fosforo (g kg-1)
Tratamento micorrízico
Controle
Acaulospora scrobiculata
Entrophospora colombiana
MIX
0
0,37 c
2,16 a
1,57 b
1,81 ab
Doses composto (g dm-3)
5
10
20
0,63 b
0,91 b
1,18 c
2,38 a
2,58 a
1,95 ab
2,04 a
2,20 a
2,28 a
1,91 a
2,17 a
1,71 bc
30
1,34 a
1,75 a
1,64 a
1,43 a
Potássio (g kg-1)
Tratamento micorrízico
Controle
Acaulospora scrobiculata
Entrophospora colombiana
MIX
0
3,11 b
5,51 a
5,28 a
5,37 a
Doses composto (g dm-3)
5
10
20
4,65 b
5,31 b
8,17 b
7,29 a
9,25 a
15,41 a
6,81 a
8,88 a
15,62 a
7,38 a
10,46 a
16,37 a
30
14,46 c
23,34 b
25,68 a
25,34 ab
Magnésio (g kg-1)
Tratamento micorrízico
Controle
Acaulospora scrobiculata
Entrophospora colombiana
MIX
0
3,86 c
4,53 bc
4,81 ab
5,72 a
Doses composto (g dm-3)
5
10
20
4,24 c
4,82 c
5,62 b
4,95 bc
5,91 b
6,46 b
5,61 b
6,05 b
6,55 b
7,04 a
7,47 a
8,11 a
30
6,51 b
8,36 a
7,51 a
8,33 a
Cálcio (g kg-1) ***
Tratamento micorrízico
Controle
Acaulospora scrobiculata
Entrophospora colombiana
MIX
Médias
14,31 b
16,56 a
15,92 a
15,97 a
Médias seguidas das mesmas letras nas colunas não diferem estatisticamente pelo teste de Tukey a
5% de probabilidade. *** Sem interação entre os fatores.
As folhas das gravioleiras não micorrizadas apresentaram os menores teores
de N, P, K, Ca e Mg em todas as doses do composto orgânico testadas. Para o
nutriente Ca, as plantas controle apresentaram os menores teores do nutriente na
35
parte aérea da gravioleira, ocorrendo diferença estatística entre os tratamentos
micorrizicos com a adição do composto orgânico (Tabela 15).
O teor de nitrogênio nas folhas das mudas de A. muricata (Tabela 15) foi
influenciado pela adição do composto orgânico ao solo, e inoculação com FMAs, onde
a ausência da inoculação levou a teores inferiores a quaisquer do tratamento com
inoculação com FMA.
O teor de K nas plantas inoculadas com FMAs diferiu de forma significativa do
controle não micorrizado com ou sem aplicação do composto orgânico. Diferenças no
teor de K nas folhas das plantas onde as espécies fúngicas foram inoculadas
ocorreram apenas nas doses mais elevadas do composto orgânico aplicado ao solo.
A regressão foi significativa para o elemento K, sendo os modelos lineares os que
melhores se ajustaram em todos os tratamentos.
30
Teor de K nas folhas (g kg-1)
25
Controle Y = Y** = 3,5174 - 0,0621X + 0,0098X² R² = 0,98
E. Colombina Y** = 3,5765 - 0,6831X R² = 0,96
A. scrobiculata Y** = 4,3824 + 0,5982X R² = 0,97
MIX Y** = 4,3558 + 0,0221X R² = 0,99
20
15
10
5
0
-5
0
5
10
20
30
Dose do composto organico (g dm-3 )
Figura 7. Análise de regressão para o teor de K nas folhas da gravioleira inoculada com E.
colombiana, A. scrobiculata e a mistura desses dois fungos micorrízicos (Mix), e não inoculada
(controle) submetida a diferentes doses de composto orgânico.
Para o nutriente Mg (Figura 8), as plantas não inoculadas também
apresentaram os menores teores em todas as doses de composto orgânico
36
incorporado ao solo. As plantas inoculadas com a mistura de FMAs foram quase
sempre superiores estatisticamente aos demais tratamento micorrízicos, não sendo
apenas na dose mais elevada do composto orgânico, onde apesar de apresentar os
maiores valores foi superior apenas ao controle não micorrizado. Houve ajustes
lineares para os modelos matemáticos para os tratamentos micorrízicos, ou seja, o
aumento da quantidade do composto orgânico se correlacionou positivamente com os
teores de Mg nas folhas da gravioleira.
10
Teor de Mg nas folhas (g kg-1 )
9
Controle Y = Y** = 3,8621 - 0,0885X + 0,0098X² R² = 0,99
E. Colombina Y** = 5,0311 + 0,0827X R² = 0,97
A. scrobiculata Y** = 4,4501 + 0,1227X R² = 0,96
MIX Y** = 6,319 + 0,0781X R² = 0,82
8
7
6
5
4
3
2
0
5
10
20
30
Dose do composto orgânico (g dm-3 )
Figura 8. Análise de regressão para o teor de Mg nas folhas de gravioleira inoculada com E.
colombiana, A. scrobiculata e a mistura desses dois fungos micorrízicos (Mix), e não inoculada
(controle) submetida a diferentes doses de composto orgânico.
Os teores de P, nas plantas não micorrizadas foram os menores em todas as
doses de composto orgânico incorporado ao solo. Na ausência do composto orgânico
as plantas inoculadas com A. scrobiculata apresentaram os maiores teores de P,
tendo comportamento semelhante até a dose central do composto orgânico. Com a
adição de 20 g dm-3 do composto, as plantas inoculadas com E. colombiana
apresentaram os maiores teores de P foliar, diferindo do controle não micorrizado e
da mistura (MIX). Não houve diferença entre os tratamentos na dose mais elevada do
composto orgânico.
37
3,5
Teor de P nas folhas ( g kg-1 )
3,0
Controle Y = 0,4731 + 0,0318X R² = 0,93
E. Colombina Y** = 1,5852 + 0,0981X - 0,0032X² R² = 0,99
A. scrobiculata Y** = 2,2504 + 0,0279X - 0,0015X² R² = 0,75
MIX Y** = 1,8391 + 0,3169X - 0,0015X² R² = 0,82
2,5
2,0
1,5
1,0
0,5
0,0
-0,5
0
5
10
20
30
Dose do composto orgânico (g dm-3 )
Figura 9. Análise de regressão para o teor de P nas folhas de gravioleira inoculada com E.
colombiana, A. scrobiculata e a mistura desses dois fungos micorrízicos (Mix), e não inoculada
(controle) submetida a diferentes doses de composto orgânico.
Com relação aos teores de P na parte aérea das plantas inoculadas com A.
scrobiculata e com E. colombiana e a mistura de ambos FMAs, os modelos
matemáticos que melhor se ajustaram foram quadráticos, e para as plantas controle
foi o ajuste linear (Figura 9). Os teores de P nas folhas das plantas controle de A.
muricata se elevaram conforme o aumento da dose de composto orgânico
previamente incorporado ao solo. As plantas inoculadas com A. scrobiculata, assim
como as inoculadas com a mistura de inóculos apresentaram teores mais elevados
de P com a adição de 10 g dm-3. Entretanto, as mudas de gravioleira inoculadas com
E. colombiana obtiveram teores de P mais elevados com a incorporação de 20 g dm 3
do composto orgânico. Por apresentar modelos de regressão de ajuste quadrático,
as equações foram derivadas para obtenção da dose ótima (X) e consequentemente
do teor ótimo (Y) de P nas folhas de gravioleira. A dose ótima do composto orgânico
para E. colombiana, A. scrobiculata e Mix foram 16,4 g dm-3, 9,3 g dm-3 e 10,6 g dm-3
respectivamente. Esses valores quando aplicados nas equações ajustadas
anteriormente geraram o teor máximo de P, sendo este 3,14 g kg-1 para E. colombiana;
38
2,5 g kg-1 para A. scrobiculata e 2,16 g kg-1 para MIX. Dessa forma, observou-se que
em relação ao nutriente P, o FMA E. colombiana apresentou uma simbiose mais
eficiente.
Os teores de cálcio presentes nas folhas da gravioleira não foram
influenciados pela interação FMA x composto orgânico. Entretanto, as duas fontes de
variação citadas exerceram influência separadamente sob o teor foliar deste mineral.
A inoculação de FMAs influenciou positivamente os teores de Ca na gravioleira
(Tabela 15). A inoculação com A. scrobiculata foi a que resultou na maior média do
teor de Ca nas folhas, entretanto ela não diferiu estatisticamente das médias dos
teores de Ca obtidos pela gravioleira inoculada com E. colombiana e com a mistura
das duas espécies fúngicas. Dessa forma gravioleiras submetidas a inoculação com
FMAs apresentaram teores de Ca foliar estatisticamente superiores aos das plantas
controle não inoculadas.
24
Teor de Ca nas folhas ( g kg -1 )
22
20
Y** = 11,2821 + 0,3390X R² = 0,99
18
16
14
12
10
8
0
5
10
20
30
Dose do composto orgânico (g dm-3 )
Figura 10. Análise de regressão para o teor de Ca nas folhas da gravioleira submetidas a
diferentes doses de composto orgânico.
Assim como a inoculação com FMAs, a incorporação de doses crescentes do
composto orgânico ao solo exerceu influência sobre o teor de cálcio nas folhas da
gravioleira (Figura 10). A análise de regressão apresentou ajuste de modelo
matemático linear significativo (p < 0,05), onde foram apresentados incrementos no
39
teor de Ca diretamente proporcionais a dosagem do composto orgânico aplicado ao
solo no experimento.
5.7.2 Micronutrientes
Houve interação entre as doses do composto orgânico e os FMAs inoculados
em gravioleira para o teor de Zn nas folhas (Tabela 16). O teor desse nutriente foi
significativamente influenciado pelo inóculo micorrízico, sendo que as médias desses
teores foram comparadas dentro de cada dose de composto orgânico. O teor de Fe,
Cu e Mn nas folhas não apresentou interação entre os fatores de variação (Tabela
16), entretanto cada fator apresentou significância separadamente para Fe e apenas
a inoculação de FMAs influenciou os teores de Cu e Mn observados na folha de
gravioleira (Tabela 16).
Tabela 16. Resumo da análise de variância para os micronutrientes na folha de gravioleira
submetida a diferentes doses de composto orgânico e inoculada com Entrophospora
colombiana, Acaulospora scrobiculata e a mistura desses dois fungos micorrízicos (Mix), e
não inoculada (controle)
Quadrado Médio
FV
GL
Fe
Zn
Cu
Mn
Bloco
3
679,21
28,06
1,23
587,32
FMA
3
1192,48*
342,22*
3,09*
2100,21*
Composto
4
2285,06*
165,82*
17,52
396,03
FMA*Composto
12
89,98
29,12*
7,79
5028,2
Erro
37
241,59
8,81
1,34
600,36
CV (%)
13,91
11,89
13,55
10,07
*Significativo a 5% de probabilidade pelo teste de F.
Os valores médios dos teores foliares de Fe, Zn, Cu e Mn nas gravioleiras
inoculadas com FMAs e submetidas a doses crescentes de composto orgânico
encontram-se na Tabela 17.
40
Tabela 17. Média do teor de micronutrientes nas folhas da gravioleira submetida a diferentes
doses de composto orgânico e inoculada com Entrophospora colombiana, Acaulospora
scrobiculata e a mistura desses dois fungos micorrízicos (Mix), e não inoculada (controle)
Ferro (mg kg-1) ***
Tratamento micorrízico
Médias
Controle
Acaulospora scrobiculata
Entrophospora colombiana
MIX
Zinco (mg kg-1)
Tratamento micorrízico
Controle
Acaulospora scrobiculata
Entrophospora colombiana
MIX
0
14,11 b
20,68 a
20,62 a
21,90 a
100,94 b
121,31 a
110,62 ab
114,15 ab
Doses composto (g dm-3)
5
10
20
16,93 b
20,32 c
19,01 b
28,01 a
28,54 b
25,13 ab
30,53 a
30,75 b
28,85 a
26,75 a
41,06 a
27,35 a
30
20,19 b
23,17 ab
27,36 a
25,91 ab
Cobre (mg kg-1) ***
Tratamento micorrizico
Controle
Acaulospora scrobiculata
Entrophospora colombiana
MIX
Médias
1,89 b
3,53 a
3,48 a
3,78 a
Manganês (mg g-1) ***
Tratamento micorrízico
Controle
Acaulospora scrobiculata
Entrophospora colombiana
MIX
Médias
56,06 b
76,35 a
84,39 a
72,47 a
Medias seguidas das mesmas letras nas colunas não diferem estatisticamente pelo teste de Tukey a
5% de probabilidade. *** Sem interação entre os fatores.
A inoculação de FMAs influenciou de forma significativa no teor do
micronutriente Fe encontrado nas folhas da gravioleira, onde o controle não
micorrizado apresentou teores inferiores aos tratamentos com inoculação. As plantas
inoculadas com A. scrobiculata apresentaram as maiores médias, sendo
estatisticamente superior ao controle não micorrizado e não diferindo dos tratamentos
com inoculação de E. colombiana e da mistura de FMAs (MIX). Estes tratamentos
mesmo apresentando teores mais elevados de Fe nas folhas não apresentaram
incrementos significativos em relação ao controle não micorrizado.
41
160
Teor de Fe nas folhas (mg kg-1 )
150
Y** = 96,5825 + 3,5734 - 0,1097X R² = 0,68
140
130
120
110
100
90
80
0
5
10
20
30
Dose do composto orgânico (g dm -3 )
Figura 11. Análise de regressão para o teor de Fe nas folhas da gravioleira submetidas a
diferentes doses de composto orgânico.
A
adição
de
doses
crescentes
do
composto
orgânico
influenciou
significativamente o teor de Fe, onde a análise de regressão apresentou ajuste de um
modelo de natureza quadrática com coeficientes significativos. Incrementos em
relação a ausência do composto foram observados até a dose central, com reduções
consideráveis nas doses mais elevadas. A partir da equação quadrática ajustada a
dose onde o teor de Fe seria máximo é de 16,28 g dm -3 do composto orgânico.
Em relação ao teor de Zn nas folhas da gravioleira (Tabela 17), as plantas não
micorrizadas (controle) tiveram teores desse micronutriente estatisticamente inferiores
aos demais em todas as doses do composto orgânico testadas. Dentre os tratamentos
com inoculação de FMAs o teor de Zn não diferiu nas menores doses do composto
orgânico (ausência e 5 g dm-3), ocorrendo diferenças nas demais doses, chamando a
atenção a dose central do composto orgânico, onde a inoculação com as duas
espécies fúngicas propiciou um incremento próximo a 50% no teor de Zn.
42
50
Teor de Zn nas folhas (mg kg-3 )
45
Controle Y = Não houve ajuste
E. Colombina Y** = 22,8682 + 1,0443X - 0,0309X² R² = 0,66
A. scrobiculata Y** = 23,7539 + 0,6021X - 0,0216X² R² = 0,65
MIX Y** = 22,7605 + 1,5872X - 0,0514X² R² = 0,51
40
35
30
25
20
15
10
5
0
5
10
20
30
Dose do composto orgânico (g dm-3 )
Figura 12. Análise de regressão para o teor de Zn nas folhas da gravioleira inoculada com E.
colombiana, A. scrobiculata e a mistura desses dois fungos micorrízicos (Mix), e não inoculada
(controle) submetida a diferentes doses de composto orgânico.
A análise de regressão da interação FMA x composto orgânico para os teores
de Zn das folhas de gravioleira apresentou coeficiente R² com valores médios porém
significativos (Figura 12). Não houve ajuste de regressão para o controle não
micorrizado. Os demais tratamentos apresentaram teores de Zn nas folhas que foram
ajustados a modelos matemáticos quadráticos com incrementos até a dose central de
composto orgânico previamente incorporado ao solo, seguidas de redução dos
valores nas doses mais elevadas.
43
Para os teores de Mn no tecido vegetal das gravioleiras, apesar das plantas
controle terem apresentado os menores valores para o micronutriente em questão,
não houve diferenças estatisticamente significativas entre estas e as demais
inoculadas com fungos micorrízicos. No caso do teor de Cu nas folhas, o tratamento
inoculado com a mistura dos FMAs apresentou teores significativamente superiores
aos demais tratamentos na dose de 10 g dm-3 do composto orgânico.
44
6. DISCUSSÃO
6.1
Altura e diâmetro do coleto
As doses do composto orgânico aplicadas ao solo influenciaram no crescimento
inicial (altura e diâmetro do coleto) da gravioleira. Houve também o incremento em
altura e diâmetro do coleto das plantas inoculadas com E. colombiana e A.
scrobiculata em relação às plantas controle. O incremento na altura das plantas
inoculadas com A. scrobiculata adubadas com 20 g dm-3 do composto orgânico foi de
63% enquanto que em E. colombiana o incremento foi de 55% na mesma dose do
composto. Nessa mesma dose, o incremento em diâmetro do coleto para a gravioleira
micorrizada inoculadas com A. scrobiculata foi de 37% com E. colombiana
apresentando 31%. Soares et. al (2012) estudando mudas de jenipapeiro inoculadas
com A. scrobiculata encontraram incremento de 54,8% em seu diâmetro em relação
a testemunha.
Os maiores incrementos no crescimento da planta hospedeira geralmente são
observados em condições de baixa fertilidade do solo (SILVA et al. 2008; CHU et al.
2001). Tal comportamento foi observado por Coelho et al. (2012) no crescimento de
mudas de pinheira (Annona squamosa L.), em que as plantas na ausência de
adubação fosfatada mas inoculadas com o FMA Gigaspora albida apresentaram um
melhor desempenho. Na ausência de adubação orgânica o incremento para altura e
diâmetro do coleto das plantas inoculadas com E. colombiana foi de 17 e 20%,
respectivamente sendo superior à A. scrobiculata, que apresentou 13 e 15% de
incremento, respectivamente. Esses dados indicam que E. colombiana estabelece
uma simbiose mais eficiente que A. scrobiculata em níveis reduzidos de fertilidade do
solo.
Independente da espécie de FMA inoculada, a gravioleira apresentou redução
nos valores de altura e diâmetro do coleto, na dose de composto orgânico mais
elevada. Resultados diferentes foram apresentados por Samarão et al. (2011), onde
a gravioleira inoculada com fungos micorrizícos arbusculares e submetida a adubação
fosfatada apresentou incrementos em altura e diâmetro do coleto, entretanto não
obteve-se o ponto máximo de crescimento, indicando que doses mais elevadas de P
em plantas micorrizadas proporcionariam incrementos ainda maiores. Vale ressaltar,
entretanto que esses autores utilizaram adubação mineral.
45
6.2
Produção de massa seca
A aplicação conjunta de substratos orgânicos e FMA pode resultar em efeito
sinérgico positivo no crescimento vegetal (SILVA et al. 2008). Isso ocorreu no presente
estudo, sendo que ambos os fatores estudados (inóculos de FMAs e adubação
orgânica) atuaram concomitantemente e de forma positiva para a produção de matéria
seca da gravioleira.
A melhor dose do composto orgânico quando associado à inoculação com E.
colombiana foi a de 10 g dm-3, enquanto as plantas inoculadas com A. scrobiculata
apresentaram uma maior produção de matéria seca total com 20 g dm -3. Dessa forma,
tomando como base os parâmetros de crescimento da gravioleira, pode-se afirmar
que a inoculação com E. colombiana ou da mistura dos inoculados foram mais
eficientes que com A. scrobiculata. Esse comportamento pode ser comprovado na
taxa de colonização micorrizica com de adubação orgânica de 10 g dm -3 de solo, onde
a raízes colonizadas por E. colombiana apresentaram valores superiores às
colonizadas por A. scrobiculata.
Foram observadas reduções nos valores de matéria seca total das plantas para
todos os tratamentos com inoculação nas doses mais elevadas do composto orgânico.
Esse comportamento em plantas é esperado para experimentos que testam
fertilizantes orgânicos e inorgânicos. Segundo Challen e Hemingway (1965) a
promoção de crescimento está ligada a fatores como AIA e AIB (auxinas), giberelinas
(A e B), micronutrientes e macronutrientes presentes nos adubos orgânico, enquanto
que os efeitos de decréscimo do crescimento das plantas podem ser atribuídos, nas
maiores concentrações desses adubos, ao excesso de fitohormônios ou à alta
concentração de minerais presentes que provocam toxidez nas plantas.
6.3
Dependência micorrízica
A dependência micorrízica das mudas de gravioleira foi influenciada pelas
doses do composto orgânico aplicado ao solo e pelas espécies de FMA inoculadas.
Apesar da redução no grau de dependência micorrízica com o aumento das
doses do composto orgânico no solo, as mudas inoculadas com A. scrobiculata e E.
colombiana foram consideradas excessivamente dependentes na dose mais levada
46
de fertilização orgânica, indicando de forma notória que a gravioleira necessita do
desenvolvimento dessa simbiose.
De acordo com os resultados obtidos, a gravioleira pode ser considerada uma
espécie micotrófica obrigatória, ou seja, é uma planta que depende da simbiose
micorrízica independentemente do nível de fertilidade do solo que essa se encontra,
principalmente em relação ao nutriente P, devido ao seu baixo grau de mobilidade no
solo.
Resultados similares foram apresentados por Brandão (2003), onde a
dependência micorrízica da gravioleira foi influenciada pela fertilização fosfatada no
solo, considerando a A. muricata altamente dependente à simbiose com inóculos
nativos, mesmo nas doses mais elevadas de P. Estudos envolvendo FMAs em A.
cherimoia, espécie do mesmo gênero da gravioleira, foi classificada por Azcón-Aguilar
et al. (1994), como sendo micotrófica obrigatória.
Costa et al. (2005), observou uma maior dependência micorrizica da
mangabeira em solo com baixos teores de P. Dessa forma, a dependência micorrizica
foi estabelecida por Moreira e Siqueira (2006), como sendo inversamente proporcional
à disponibilidade de P no solo. No presente estudo resultados com essa tendência
foram encontrados, onde a porcentagem de dependência micorrizica da gravioleira
diminuiu conforme o aumento das doses do composto orgânico.
6.4
Grau de colonização micorrízica
O efeito das micorrizas na planta, na maioria das vezes, está positivamente
correlacionado ao percentual de colonização das raízes (SHENG et al., 2004),
fenômeno que foi observado nas gravioleiras inoculadas com A. scrobiculata e E.
colombiana que tiveram percentual de colonização micorrizica que alcançou 80% das
raízes mais finas, valor superior ao observado por Chu et al. (2001) em mudas de
gravioleira em substrato não fumigado, na qual as plantas inoculadas com
Scutellospora heterogama, Gigaspora margarita, Entrophospora colombiana e
Gigaspora sp apresentaram taxa de colonização micorrizica de 30,6%, 28,6%, 30,2%
e 36,5% respectivamente. Diversas outras frutíferas vêm apresentando desempenho
semelhante, como pessegueiro (NUNES et al., 2008), jenipapeiro (SOARES et al.,
2012), meloeiro (SILVA JÚNIOR et al.,2010), aceroleira (BALOTA et al., 2011).
47
Samarão et al. (2011), não obteve ajuste de modelos de regressão com
coeficientes significativos para o percentual de raízes colonizadas de gravioleira, onde
a inoculação com Glomus clarum e Glomus magarita em função das doses de P
aplicada, manteve a taxa de colonização em 19 e 30% respectivamente, independente
da dose de P aplicada. Assim como na gravioleira, a inoculação do FMA A.
scrobiculata em mudas de jenipapeiro (SOARES et al,2012) apresentou uma
porcentagem de colonização radicular de 74,3% que pode ser considerada alta.
O teor de P disponível no solo ou substrato vem sendo frequentemente
documentado como o principal fator para a redução da colonização micorrizica,
podendo diminuir a produção de esporos e formação de hifas (MENGE et al, 1978;
BRUCE et al, 1994; De MIRANDA e HARRIS, 1994; LU et al., 1994; VALENTINE et
al., 2001). Altos teores de P no tecido da raiz de plantas reduzem a secreção de
moléculas de sinalização que são responsáveis pela ramificação de hifas
(NAGAHASHI e DOUDS, 2000) e modificam os fosfolipídios das células que
influenciam a permeabilidade da membrana e libertação do composto de carbono que
é essencial para a manutenção dos FMA’s nas raízes (GRAHAM et al, 1981; SCHWAB
et al., 1991). No presente estudo, a taxa de colonização micorrízica da gravioleira não
foi reduzida de forma consistente, sendo observados valores acima de 50% de
micorrização na dose mais elevada (30 g dm -3) do composto orgânico, o que
corresponde a uma dose de P de 680 mg dm-3.
6.5
Densidade de esporos
A aplicação de material orgânico de qualidade podem otimizar os efeitos dos
fungos micorrízicos arbusculares nas plantas (CARAVACA et al., 2004). Para Silveira
et al. (1998) os mesmos fatores que influenciam na taxa de colonização micorrízica
também podem influenciar na esporulação dos FMAs. Esse comportamento foi
observado no presente estudo, onde ocorreu uma correlação positiva entre
colonização e esporulação dos FMAs.
A densidade de esporos em 50 g de solo rizosférico nos tratamentos com
inoculação de FMA esteve sempre entre 100 e 700 esporos, a depender da dose de
composto aplicada ao solo. Amostras dos solos sem inoculação apresentaram poucos
esporos envelhecidos.
48
Balota et al. (2011), obtiveram em aceroleira, correlações altamente
significativas entre a colonização e esporulação de G. margarita (r=0,94; p≤0,01), G.
manihotis (r=0,96; p≤0,01) e G. clarum (r=0,70; p≤0,01). No presente estudo
resultados semelhantes foram obtidos para E. colombiana (r=0,91; p≤0,01), A.
scrobiculata (r=0,76; p≤0,01) e a mistura dos inoculos (r=0,77; p≤0,01). Esses mesmo
autores observaram diminuição na esporulação de G. margarita G. manihotis e G.
clarum em doses com adubação fosfatada superior a 100 mg kg-1 com esporulação
variando 252 a 388; 15 a 368 e 75 a 91 respectivamente. O comportamento no
presente estudo foi semelhante para a gravioleira inoculada com A. Scrobiculata e
Entrophospora colombiana, assim como na presença de ambos. Isso pode ser
explicado pelo P presente em cada dose do composto orgânico, como foi sugerido por
Moreira e Siqueira (2006), onde elevadas teores de P no solo influenciaram
negativamente na ação de FMAs. Smith e Read, (1997) relataram que o P pode estar
relacionado a taxa de disseminação dos FMA e sobre o desenvolvimento do micélio
extra–radicular, sendo este efeito regulado por fatores ambientais e pela espécie
hospedeira.
6.6
Glomalina no solo
Silva et al. (2008), avaliaram a influência da adubação com vermicomposto
sobre a produção de glomalina pelos FMAs Gigaspora albida e Acaulospora longula,
observando incrementos superiores a 100% em comparação ao solo não adubado.
Wuest et al. (2005) também registraram aumento na produção de glomalina em solo
adubado com esterco bovino. Esses aumentos apresentados em solos adubados com
manejo orgânico refletem melhorias na qualidade do solo, dada a correlação existente
entre glomalina e agregação de partículas (RILLIG, 2004).
O teor de glomalina facilmente (GFE) disponível em agregados seguiram a
mesma tendência apresentada pelos autores citados anteriormente. O incremento nos
teores de glomalina no solo sem adição do composto orgânico esteve em torno de
23%, independente do FMA inoculado. O maior incremento ocorreu na presença A.
scrobiculata e E. colombiana e na dose mais elevada de composto aplicada ao solo.
O teor de GFE correlacionou-se de forma altamente significativa com o grau de
colonização radicular da gravioleira em todos os tratamentos micorrizicos: A.
49
scrobiculata (r = 0,65; p≤0,01); E. colombiana (r = 0,71; p≤0,01) e Mix (r = 0,67;
p≤0,01). Com relação ao número de esporos, os teores de glomalina apresentaram
correlação altamente significativa para A. scrobiculata (r = 0,61; p≤0,01) e E.
colombiana (r = 0,61; p≤0,01) e significativa na presença das duas linhagens de FMA
(r = 0,45; p≤0,05). Essa correlação positiva pode ser perfeitamente explicada,
considerando que 80% da glomalina presente no solo seriam oriundas da
decomposição de hifas e esporos de FMAs (DRIVER et al., 2005).
6.7
Nutrientes na folha
Com relação aos nutrientes analisados (N, P, K, Ca, Mg, Fe, Cu, Zn e Mn) os
valores encontrados nas plantas não micorrizadas foram significativamente menores
do que os das plantas micorrizadas. Os dados indicaram uma correlação significativa
entre a porcentagem de colonização radicular e os teores de P (r=0,77; p≤0,01) Ca
(r=0,34; p≤0,01), Mg (r=0,59; p≤0,01), e Zn (r=0,73; p≤0,01). Soares et al. (2012),
também encontraram correlações positivas com N (r=0,71; p≤0,001), P (r=0,79;
p≤0,001), K (r=0,43; p≤0,01), Mg (r=0,58; p≤0,001) e Cu (r=0,52; p≤0,01) em
jenipapeiro.
Correlações positivas entre o teor de nutrientes e colonização radicular por
fungos micorrizicos arbusculares podem resultar na de até 80% de fósforo, 60% de
cobre, 25% de nitrogênio, 25% de zinco e 10% de K para a planta (MARSCHENER e
DELL, 1994).
O teor do macronutriente N nas folhas da gravioleira foi influenciado pela
inoculação de FMAs. A absorção desse elemento por plantas micorrizadas, está
relacionada a forma como este está disponível, já que a forma amoniacal é
considerada menos móvel que a do nitrato (CANTARELLA, 2007).
Os maiores teores de P e Zn encontrados nas folhas da gravioleira inoculadas
com A. scrobiculata, E. colombiana e a mistura desses inóculos podem ser explicados
pela atuação das hifas extraradiculares dos FMAs que aumentam a área radicular
explorada (micorrizosfera), promovendo a absorção desses nutrientes (LAMBERT et
al. 1979; EL-SHAIK e MOHAMMED, 2009), de baixa mobilidade no solo e que tem
absorção via difusão e interceptação radicular.
A adição de doses crescentes do composto orgânico propiciou aumentos
lineares nos teores de K e Mg. Os menores teores desses nutrientes foram sempre
50
das plantas controle não inoculadas com FMAs, independente da dose de composto
incorporada ao solo. Dessa forma inoculação com A. scrobiculata, E. colombiana e a
misturas deste propiciou incrementos de K e Mg, podendo então se dizer que a
simbiose micorrizica na gravioleira exerce influência sobre a absorção desses
nutrientes. Resultados semelhantes foram observados por Samarão et al. (2011) os
quais citam conteúdo de K 47,8% superior ao tratamento sem inoculação. Altoé et al.
(2008) concluíram que a inoculação com A. scrobiculata não influenciou o conteúdo
de P, Mg e S na parte aérea de tangerineira ‘Cleópatra’, elevando apenas o conteúdo
de K.
Os nutrientes fósforo, zinco e cobre compõe o grupo dos nutrientes pouco
móveis e a forma de transporte dificulta a absorção de nutrientes pelo sistema
radicular das plantas (CARDOSO et al., 2010). Os teores de P e Zn apresentaram
redução nas doses mais elevadas do composto orgânico incorporado ao solo. Esse
comportamento pode perfeitamente ser explicado, já que a aplicação de níveis
elevados de nutrientes, pode reduzir a taxa de colonização e seus benefícios em
promover a nutrição da planta podem até ser anulados (BOLAN et al., 1984). Esse
comportamento não foi observado por Samarão et al. (2011) na gravioleira, onde
foram encontrados incrementos no conteúdo de P na parte aérea inoculadas com
FMAs.
Em relação ao micronutriente zinco Lehmann et al. (2014) afirmaram que a taxa
de colonização radicular influencia as concentrações de Zn independente do tecido
celular, já que a simbiose acaba por diminuir a limitação imposta pelo transporte por
difusão.
51
7. CONCLUSÃO
Houve influência da interação composto orgânico e inoculação micorrízica
sobre o crescimento de mudas de gravioleira.
A gravioleira foi considerada altamente dependente a simbiose micorrízica e a
aplicação do composto orgânico influenciou o grau de colonização micorrízica e a
esporulação de FMAs, com reduções a partir nesses parâmetros nas doses mais
elevadas (20 e 30 g dm-3).
A inoculação com FMAs estimulou a absorção de nutrientes, com redução nos
teores de P foliar nas doses mais elevadas.
A inoculação de fungos micorrízicos elevou os teores de glomalina facilmente
disponível do solo.
52
8. REFERÊNCIAS
ABIOLA, S. S.; TEWE, O. O. Chemical evaluation of cocoa by-products. Tropical
Agriculture, Trinidad, v. 68, n. 4, p. 335-336, 1991.
ADAB. Agência de Defesa Agropecuária do Estado da Bahia. Disponível em:
<http://www.adab.ba.gov.br/modules/news/article.php?storyid=480>. Acesso em: 01
Nov. 2014.
ALLEN, M. F. The ecology of arbuscular mycorrhizas: a look back into the 20th
century and a peek in to the 21 st. Mycological Research, v. 100, n.07, p. 769 782,
1996.
ALTOÉ, J. A.; MARINHO, C. S.; MUNIZ, R. A.; RODRIGUES, L. A.; GOMES, M. M.
A. Tangerineira ‘Cleópatra’ submetida a micorrização e a um análogo de
brassinosteróide. Acta Scientiarum. Agronomy, v. 30, n. 1, p. 13-17, 2008.
AZCÓN-AGUILAR, C.; ENCINA, C.L.; AZCÓN, R.; BAREA, J.M. Mycotrophy of
Annona-cherimola and the morphology of its mycorrhizae. Mycorrhiza 4:161168.1994.
BÁEZ-PÉREZ, A.; GONZALEZ-CHAVEZ, A.; ETCHEVERS-BARRA, J. D.; PRAT, C.;
HIDALGO-MORENO, C. Glomalina y secuestro de carbono em tepetates cultivados.
Agrociencia, v. 44, n. 5, p. 517-529, 2010.
BALOTA, E. L.; MACHINESKI, O.; STENZEL, N. M. C. Resposta da acerola à
inoculação de fungos micorrízicos arbusculares em solo com diferentes níveis de
fósforo. Bragantia, Campinas, v. 70, n. 1, p.166-175, 2011.
BERBARA, R. L. L.; SOUZA, F. A.; FONSECA, H. M. A. C. Fungos micorrízicos
arbusculares: muito além da nutrição. In: FERNANDES, M. S. (Ed). Nutrição
Mineral de Plantas, Viçosa: SBCS, 432p., 2006.
BERBERT, P. R. Estudo da pectina do mel e da casca do fruto do cacau. Revista
Theobroma, Ilhéus, v. 2, n. 49, p. 51, 1972.
BOLAN, N. S.; ROBSON, A. D.; BARROW, N.J. Increasing phosphorus supply can
increase the infection of plant roots by vesicular-arbuscular mycorrhizal fungi. Soil
Biology and Biochemistry 16: 419–420, 1984.
BONILLA, J. A. Fundamentos da agricultura ecológica: sobrevivência e
qualidade de vida. São Paulo: Nobel, 260 p., 1992.
BORGES, A. L.; TRINDADE, A. V.; SOUZA, L. da S.; SILVA, M. N. B. da. Cultivo
orgânico de fruteiras tropicais – manejo do solo e da cultura. Cruz das Almas:
EMBRAPA – CNPMF, Comunicado Técnico 64, 2003.
BORIE, F.; RUBIO, R.; ROUANET, J. L.; MORALES, A.; BORIE, G.; ROJAS, C.
Effect of tillage systems on soil characteristics, glomalin and mycorrhizal propagules
53
in a Chilean Ultisol. Soil & Tillage Research, Netherlands, v. 88, n. 1-2, p. 253-261,
2006.
BRADFORD, M. M. A rapid and sensive method for the quantification of microgram
quantities of protein utilizing the principle of protein-dye binding. Analytical
Biochemistry, v. 72, n. 1-2, p. 248-254, 1976.
BRANDÃO, J. A. C. Simbiose micorrízica arbuscular de gravioleiras (Annona
Muricata) em solo infestado por pratylenchus coffeae. 74f. Dissertação
(Programa de Pós-Graduação em Biologia de Fungos), Universidade Federal de
Pernambuco. Recife, 2003.
BRUCE, A.; SMITH, W. E.; TESTER, M. The development of mycorrhizal infection in
cucumber: effects of P supply on root growth, formation of entry points and growth of
infection units. New Phytologist, 1276: 507–514, 1994.
CACCO, G. & DELL AGNOLLA, G. Plant Growth Regulator Activitiy of Soluble Humic
Substances. Can. Journal of Soil Science, 64: 25-28, 1984.
CANTARELLA, H. Nitrogênio. In: NOVAIS, R. F.; ALVAREZ, V. H. V.; BARROS, N.
F.; FONTES, R. L. F.; CANTARUTTI, R. B.; NEVES, J. C. L. Fertilidade do Solo.
Viçosa, Sociedade Brasileira de Ciência do Solo, 2007. p. 375-470.
CARAVACA, F.; ALGUACIL, M.M.; AZCON, R.; DIAZ, G. & ROLDAN, A. Comparing
the effectiveness of mycorrhizal inoculation and amendment with sugar beet, rock
phosphate and Aspergillus niger to enhance field performance of the leguminous
shrub Dorycnium pentaphyllum L. Applied Soil Ecology 25: 169-180, 2004.
CARDOSO, E. J. B. N.; CARDOSO, I. M.; NOGUEIRA, M. A.; BARRETA, C. R. D.
M.; de PAULA, A. M. Micorrizas Arbusculares na aquisição de nutrientes pelas
plantas. In: SIQUEIRA, J. O.; de SOUZA, F. A.; CARDOSO, E. J. B. N.; TSAI, S. M.
Micorrizas: 30 anos de pesquisas no Brasil. Lavras: Ufla, 2010. 716 p.
CARNEIRO, M.A.C.; SIQUEIRA, J.O.; MOREIRA, F.M.S.; CARVALHO, D.;
BOTELHO, S.A.; JUNIOR, O.J.S. Micorriza arbuscular em espécies arbóreas e
arbustivas nativas de ocorrência no sudeste do Brasil. Cerne, v.4, p.129-145, 1998.
CHALLEN, S. B.; HEMINGWAY, J. C. Growth of higher plants in response to
feeding with seaweed extract. Proceedings of 5 International Seaweed
Symposium, p. 57, 1965.
CHU, E. Y.; MÖLLER, M. R. F.; CARVALHO, J. G. Efeitos da inoculação micorrízica
em mudas de gravioleira em solo fumigado e não fumigado. Pesquisa
Agropecuária Brasileira, v. 36, n. 4, p. 671–680, 2001.
COELHO, I. R.; CAVALCANTE, U. M. T.; CAMPOS, M. A. S.; DA SILVA, F. S. B.
Uso de fungos micorrízicos arbusculares (FMA) na promoção do crescimento de
mudas de pinheira (Annona squamosa L., Annonaceae). Acta Botanica Brasilica,
26(4): 933-937, 2012.
54
CORNEJO, P.; MEIER, S.; BORIE, G.; RIILING, M. C.; BORIE, F. Glomalin-related
soil protein in a Mediterranean ecosystem affected by a copper smelter and its
contribution to Cu and Zn sequestration. Science of the Total Environment, v. 406,
n. 1-2, p. 154-160, 2008.
COSTA, A. M. G.; COSTA, J. T. A.; CAVALCANTI JUNIOR, A. T.; CORREIA D. e
MEDEIROS FILHO S. Influência de diferentes combinações de substratos na
formação de porta-enxertos de gravioleira (Annona muricata L.). Revista de Ciência
Agronômica, v.36, n.3, p.299-305, 2005.
DE MIRANDA, J. C. C.; HARRIS, P. J. Effects of soil phosphorus on spore
germination and hyphal growth of arbuscular mycorrhizal fungi. New Phytologist,
128: 103–108, 1994.
DECLERCK, S.; PLENCHETTE, C.; STRULLU, D. G. Mycorrhizal dependency of
banana (Musa acuminata, AAA group) cultivar. Plant and Soil, Dordrecht, v. 176,
n. 1, p. 183-187, 1995.
DRIVER, J. D.; HOLBEN, W. E.; RILLIG, M. C. Characterization of glomalin as a
hyphal wall componente of arbuscular mycorrhizal fungi. Soil Biology &
Biochemistry, Oxford, v. 37, n. 1, p. 101-106, 2005.
EL-SHAIKH, K. A. A.; MOHAMMED, M. S. Enhancing fresh and seed yield of okra
and reducing chemical phosphorus fertilizer via using Va-mycorrhizaal inoculants.
World Journal of Agricultural Sciences 5 (S), 810-818, 2009.
EMBRAPA – Empresa Brasileira de Pesquisa Agropecuária. Centro Nacional de
Pesquisas de Solos. Manual de métodos de análises de solos. 2.ed. Rio de
Janeiro: Embrapa Solos, 2011. 230p.
EMBRAPA – Empresa Brasileira de Pesquisa Agropecuária. Manual de análises
químicas de solos, plantas e fertilizantes. 2. ed. rev. Amp. Rio de Janeiro:
Embrapa Solos, 2009. 627p.
FERREIRA, E. B.; CAVALCANTI, P. P.; NOGUEIRA, D. A. Experimental Designs:
um pacote R para análise de experimentos. Revista da Estatística da UFOP, V. 1,
N. 1, p.1-9. 2011.
FIGUEIRA, A.; JANICK, J.; BEMILLER, J. N. New products from Theobroma cacao:
Seed pulp and pod gum. In: JANICK, J. and SIMoN, J. E. (Ed.). New crops. New
York: Wiley, 1993. p. 475-478.
FREITAS, G. B. Propagação, florescimento, frutificação e produção da
gravioleira (Annona muricata L.). 1987. 87f. Tese (Doutorado em Fitotecnia).
Universidade federal de Viçosa, Viçosa, 1997.
GERDEMANN, J.W. Vesicular-arbuscular mycorrhizae, In: J.G. Torrey; D.T.
Clarkson; Eds. The development and function of roots, Academic Press, New York,
USA, 1975.
55
GERDEMANN, J.W.; NICOLSON, T.H. Spores of mycorrhizal Endogone species
extracted from soil by wet sieving and decanting. Transactions of British
Mycological Society, 46:235-244, 1963.
GONZÁLEZ-CHAVÉZ, M. C.; CARRILLO GONZÁLEZ, R.; WRIGHT, S. F.;
NICHOLS, K. A. The role of glomalin, a protein produced by Arbuscular mycorrhizal
fungi, in sequestering potentially toxic elements. Environmental Pollution, v. 130, n.
3, p. 317-323, 2004.
GRAHAM, J. H.; LEONARD, R. T.; MENGE, J. A. Membrane-mediated decrease in
root exudation responsible for phosphorus inhibition of vesicular-arbuscular
mycorrhiza formation. Plant Physiology 68: 548–552, 1981.
HABTE, M.; MANJUNATH, A.: Categories of vesicular-arbuscular mycoohizal
dependency of host species. Mycorrhiza 1:3-12, 1991.
HARINIKUMAR, K. M.; BAYARAJ, D. J. Effect of cropping sequence, fertilizers and
farmyard manure on vesicular-arbuscular mycorrhizal fungi in different crops over
three consecutive seasons. Biology and Fertility of Soil. 7(2): 173-175, 1989.
JANOS, D.P. Mycorrhizae influence tropical sucession. Biotropica, Washington,
v.12, n.1, p.56-64, 1980.
KAKULU, S. E.; OSIBANJO O. Pollution studies of Nigerian rivers: Trace metals
levels of surfasse waters in the Niger Delta area. International Journal of
Environmental Studies. 41, 287-292, 1992.
LAMBERT, D. H.; BAKER, D. E.; COLE, H. The role of mycorrhizae in interactions of
phosphorus with zinc, copper and other elements. Soil Society of American
Journal 43, 976-980, 1979.
LEHMANN, A.; VERESOGLOU, S. D.; LEIFHEIT, E. F.; RILLIG, M. C. Arbuscular
mycorrhizal influence on zinc nutrition in crop plants e A meta-analysis. Soil Biology
& Biochemistry 69: 123-131.2014.
LIMA, M. A. C. et al. Comportamento respiratório e qualidade pós-colheita de graviola
(Annona muricata L.) “Morada” sob temperatura ambiente. Revista Brasileira de
Fruticultura, Jaboticabal - SP, v. 25, n. 1, p. 49-52. 2003.
LU, S.; BRAUNBERGER, P. G.; MILLER, M. H. Response of vesicular-arbuscular
mycorrhizae of maize to various rates of P addition to different rooting zones. Plant
and Soil, 158: 119–128, 1994.
MALAVOLTA, E.; PIMENTEL-GOMES, F.; ALCARDE, J.C. Adubos e adubações.
São Paulo: Nobel, 2002. 200p.
MANICA, I. Taxonomia, morfologia e anatomia. In: SÃO JOSÉ, A. R. et al. (eds.).
Anonáceas: Tecnologia de produção e comercialização. Vitória da Conquista:
DFZ/UESB, p. 20- 35.1997.
56
MARSCHNER, H. Mineral nutrition of higher plants. San Diego, Academic Press,
1995. 889p.
MCGONIGLE, T.P.; MILLER, M.H.; EVANS, D.G.; FAIRCHILD, G.L.; SWAN, J.A. A
new method which gives an objective measure of colonization of roots by vesiculararbuscular mycorrhizal fungi. New Phytologist, v.115, p.495-501, 1990.
MEHROTRA, V. S. Arbuscular mycorrhizal associations of plants colonizing coal mine
spoil in Índia. Journal of Agricultural Science, Cambridge, Inglaterra, v. 130, n. 2, p.
125-133, Mar. 1998.
MELO, G. S.; NETO, L. G.; MOURA, R. J. M. Cultivo da gravioleira (Annona
muricata L.). Recife, PE. 1983. 4 p. (Instruções Técnicas do IPA; 13).
MENGE, J. A.; STEIRLE, D.; BAGYARAJ, D. J.; JOHNSON, E. L. V.; LEONARD, R.
T. Phosphorus concentration in plants responsible for inhibition of mycorrhizal
infection. New Phytologist, 80: 575–578, 1978.
Miller RM, Jastrow JD. 2000. Mycorrhizal fungi influence soil structure. In: Kapulnik
Y, Douds DD, eds. Arbuscular mycorrhizas: molecular biology and physiology.
Dordrecht, the Netherlands: Kluwer Academic, 3–18.
MIYASAKA, S.; NAKAMURA, Y.; OKAMOTO, H. Agricultura natural. 2. ed. Cuiabá:
SEBRAE/MT, 1997. 73 p. (Coleção agroindústria).
MOLINA, R.; MASSICOTTE, H.; TRAPPE, J.M. Specific ity phenomena in
mycorrhizal symbiosis: community ecological consequences and practical
implications. In: ALLEN, M.F. Mycorrhizal functioning an integrated plant fungal
process. New York, Chapman and Hall, 357-423, 1992.
MOREIRA, F. M. S.; SIQUEIRA, J. O. Microbiologia e Bioquímica do Solo. 2º
Edição Atualizada e Ampliada - Lavras: Editora UFLA, 2006.
MORORÓ, R. C. Aproveitamento de derivados, Subprodutos e resíduos do cacau.
In: VALLE, R. R.(Ed.). Ciência, tecnologia e manejo do cacaueiro. Ilhéus: Ceplac,
2007. p. 371- 421.
MORTON, J. F. The Soursop of guanábana (Annona muricata L.) Proceedings of
the Florida State Horticultural Society. v. 79, p. 355-366, 1966.
NAGAHASHI, G. & Jr. DOUDS, D. D. Partial separation of root exudates components
and their effects upon the growth of germinated spores of AM fungi. Mycological
Research, 104: 1453–1464, 2000.
NANNIPIERI, P.; GREGO, S.; DELL’AGNOLA, G.; NARDI, S. Proprietà
biochimiche e fisiologiche dela sostanza orgánica. In: NANNIPIERI, P. (ed.).
Ciclo dela sostanza orgánica nel suolo: aspetti agronomici, chimici, ecologici y
selvicolturali. Bologna: Patron, 1993, p. 67-68.
57
NUNES, J.L.S.; SOUZA, P.V.D.; MARODIN, G.A.D.; FACHINELLO, J.C. Incremento
no desenvolvimento do porta-enxerto de pessegueiro 'Aldrighi' por fungos
micorrízicos arbusculares autóctones. Ciência e Agrotecnologia, v.32, p.17871793, 2008.
OLAWUYI, O. J.; EZEKIEL-ADEWOYIN, D. T.; ODEBODE, A. C.; AINA, D. A.;
ESENBAMEN, G.E. Effect of arbuscular mycorrhizal (Glomus clarum) and
organomineral fertilizer on growth and yield performance of okra (Abelmoschus
esculentus). African Journal of Plant Science Vol. 6 (2), pp.84-88, 2012.
OLSSON, P.A.; THINGSTRUP, I.; JAKOBSEN, I. & BAATH, F. Estimation of the
biomass of arbuscular mycorrhizal fungi in a linseed field. Soil Biology and
Biochemistry, 31:1879-1887, 1999.
PETERSON, R. L.; HOWARTH, M. J.; WHITTIER, D. P. Interactions between a
fungal endophyte and gametophyte cells in Psilotum nudum. Canadian Journal of
Botany 59, 711-720, 1981.
PHILLIPS, J.M.; HAYMAN, D.S. Improved procedures for clearing roots and staining
parasitic and vesicular arbuscular mycorrhizal fungi for rapid assessment of infection.
Transactions of British Mycological Society, v. 55, p. 158-161, 1970.
PINTO, A. C. Q.; GENÚ, P. J. C. Contribuição ao estudo técnico-científico da
graviola (Annona muricata). In: CONGRESSO BRASILEIRO DE FRUTICULTURA, 7.
1984. Florianópolis-SC. Anais... Florianópolis: SBF/EMPASC, 1984. v. 2, p. 529546.
PINTO, A. C. Q.; RAMOS, V. H. V.; RODRIGUES, A. A. Aspectos botânicos. In:
OLIVEIRA, M. A. S (ed.). Graviola. Produção: aspectos técnicos. Embrapa Cerrados
(Planaltina, DF). Brasília: Embrapa Informação Tecnológica. 2001c. p. 10-12 (Frutas
do Brasil; 15).
PINTO, A. C. Q.; SILVA, E. M. Graviola para exportação: aspectos técnicos da
produção. Brasília: EMBRAPA-SPI, 1994. 41 p. (Série Publicações Técnicas
FRUPEX; 7).
PLENCHETTE, C.; FORTIM, J. A.; FURLAM, V. Growth response of several plant
species to mycorrhizae in a soil of moderate P-fertility: Mycorrhizal dependency
under field conditions. Plant and Soil, 70:199-209,1983.
PURIN, S.; RILLIG, M. C. The arbuscular mycorrhizal fungal protein glomalin:
limitations, progress, and a new hypothesis for its function. Pedobiologia, v. 51, n. 2,
p.123-130, 2007.
RAMOS, A. C.; MARTINS, M. A. Fisiologia de micorrizas arbusculares. In:
SIQUEIRA, J. O.; SOUZA, F. A.; CARDOSO, E. J. B. N.; MUI TSAI, S (ed).
Micorrizas: 30 anos de pesquisa no Brasil. Lavras: UFLA, 2010. 716 p.
RAMOS, V. H. V.; PINTO, A. C. Q.; RODRIGUES, A. A. Introdução e importância
socioeconômica. In: OLIVEIRA, M. A. S. (ed.). Graviola. Produção: aspectos
58
técnicos. Embrapa Cerrados (Planaltina, DF) Brasília: Embrapa Informação
Tecnológica. 2001. p. 9. (Frutas do Brasil; 15).
RILLIG, M.C. Arbuscular mycorrhizae, glomalin, and soil aggregation. Canadian
Journal of Soil Science 36: 355-399, 2004.
SACRAMENTO, C. K.; MOURA, J. I. L.; COELHO JUNIOR, E. Graviola. In:
SANTOS-SEREJO, J. A. et al. (eds.). Fruticultura tropical: espécies regionais e
exóticas. Brasília, DF: Embrapa Informação Tecnológica, 2009. p. 95-132.
SAMARÃO, S. S.; RODRIGUES, L. A.; MARTINS, M. A. MANHÃES, T. N.; ALVIM, L.
A. DA M. Desempenho de mudas de gravioleira inoculadas com fungos micorrízicos
arbusculares em solo não-esterilizado, com diferentes doses de fósforo. Acta
Scientiarum Agronomy, Maringá, v. 33, n. 1, p. 81-88, 2011.
SÃO JOSÉ, A. R. Cultivo e mercado da graviola. Fortaleza: Instituto Frutal, 2003.
36 p.
SCHWAB, S. M., MENGE, J.; TINKER, P.B. Regulation of nutrient transfer between
host and fungus in vesicular-arbuscular mycorrhiza. New Phytol., 117: 387–398,
1991.
SHENG, Z. W.; FEN, L. Y.; WU, Z. S.; BAO, X. X. Effects of arbuscular mycorrhizal
fungi on plant growth and osmotic adjustment matter content of trifoliate orange
seedling under water stress. Journal of Plant Physiology and Molecular Biology,
30: 583–588, 2004.
SILVA JUNIOR, J. M. T. et al. Desenvolvimento de meloeiro associado a fungos
micorrizicos arbusculares e cultivado em substrato pó de coco. Revista Brasileira
de Ciências Agrárias, v. 05, n. 01, p. 54-59, 2010.
SILVA, D.K.A.; SILVA, F.S.B.; YANO-MELO, A.M. & MAIA, L.C. Uso de
vermicomposto favorece o crescimento de mudas de gravioleira (Annona muricata L.
´Muricata`) associada a fungos micorrizícos arbusculares. Acta Botanica Brasilica
22: 863-869, 2008.
SILVA, I. R., MENDONÇA, E. S., 2007. Matéria orgânica do solo. In: NOVAIS, R. F.
et al. (Org.) Fertilidade do Solo. Viçosa: Sociedade Brasileira de Ciência do Solo,
pp. 275-374, 2007.
SILVEIRA, A.P.D. Ecologia de fungos micorrizicos arbusculares. In: Melo, I.S. &
Azevedo, J.L. (Eds.). Ecologia microbiana. Jaguariuna, Embrapa – CNPMA, 1998.
SMITH, S. E.; READ, D. J. Vesicular Arbuscular mycorrhiza in agriculture and
horticulture. In: Mycorrhiza symbiosis. Second edition, Smith, SE, DJ Read (eds),
Academic Press, London, UK, p. 453-469, 1997.
SMITH, S.E. & READ, D.J. Mycorrhizal symbiosis. 2ed. London. Academic Press.
1997.
59
SOARES, A.C.F.; Sousa, C.S.; Garrido, M.S. e Lima, F.S. Fungos Micorrízicos
Arbusculares no crescimento e nutrição de mudas de jenipapeiro. Revista Ciência
Agronômica, vol. 43, n. 1, p. 47-54, 2012.
SOUSA, C. S.; MENEZES, R. S. C.; SAMPAIO, E. V. S. B.; LIMA, F. S. Influências
da temperatura de armazenamento e de extratores na determinação de glomalina
em solos Paraibanos. Revista Ciência Agronômica, Fortaleza, v. 42, n. 4, p. 837841, 2011.
STATISTICA for Windows. Computer program.Statsoft, Inc; Tulsa, 2002.
THIRUMARAN, G.; ANANTHARAMAN, P.; KANNAN, L. Effect of seaweed extracts
used as a liquid fertilizer in the radish (Raphanus sativus) Journal of Ecobiology,
20(1): 49-52, 2007.
VALENTINE, A. J., OSBORNE, B. A.; MITCHELL, D. T. Interactions between
phosphorus supply and total nutrient availability on mycorrhizal colonization, growth
and photosynthesis of cucumber. Scientia Horticulturae, 88: 177–189, 2001.
VODNIK, D.; GRCMAN, H.; MACEK, I.; ELTEREN VAN, J. T.; KOVACENIC, M. The
contribution of glomalin-related soil protein to Pb and Zn sequestration in polluted
soil. Science of the Total Environment, v. 392, n. 1, p. 130-136, 2008.
WRIGHT, S. F.; UPADHYAYA, A. Extraction of an abundant and unusual protein
from soil and comparison with hyphal protein of arbuscular mycorrhizal fungi. Soil
Science, Madison, v.161, n. 9, p. 575-586, 1996.
WUEST, S.B.; CAESAR-TONTHAT, T.C.; WRIGTH, S.F. & WILLIAMS, J.D. Organic
matter addition, N, and residue burning effects on infiltration biological and physical
properties of na intensively tilled silt-loam soil. Soil and Tillage Research 84: 154167, 2005.