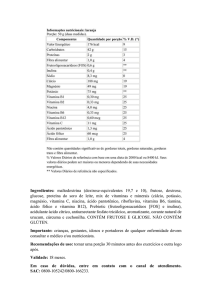
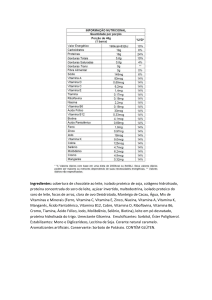
IMPACTO DA NUTRIÇÃO VITAMÍNICA SOBRE A
RESPOSTA IMUNOLÓGICA DAS AVES
Fernando Rutz
Viviane Lorenzato Bermudez
Geferson Fischer
Ederson Adriano Pan
Universidade Federal de Pelotas
As vitaminas são nutrientes essenciais que atuam em mais de 30 reações
metabólicas celulares (Marks, 1979). Embora o conceito de vitaminas menciona
que as mesmas não podem ser sintetizadas pelo organismo, dependo da espécie
animal e do ambiente onde se encontram, algumas podem ser sintetizadas, como é o
caso da vitamina D (caso o animal tiver acesso ao sol), a vitamina C (caso o animal
tenha a enzima gulonolactona oxidase), a niacina (provém do triptofano) e a colina.
As vitaminas estão presentes nos ingredientes utilizados nas dietas ou podem ser
suplementadas em forma de premix (aproximadamente 0,05% da dieta).
Geralmente nutricionistas fornecem níveis mínimos necessários para o máximo
desempenho e lucro, acrescidos de margem de segurança baseados em experiências
práticas. Os critérios utilizados para determinar as exigências são: 1) determinar
os fatores na granja que influenciam a suplementação vitamínica; 2) determinar os
objetivos a serem alcançados com o mínimo custo; 3) determinar uma margem de
segurança para fatores estressantes que possam vir a aparecer entre lotes e locais
(Coelho e McNaughton, 1995).
Dentre outras funções, as vitaminas participam no metabolismo como imunomoduladores para melhorar as funções imunológicas e a resistência a infecções em aves e
outros animais domésticos.
Considerações sobre o sistema imunológico das aves
O sistema imunológico das aves pode ser dividido em imunidade humoral e
celular. A imunidade humoral é caracterizada por uma função adaptativa do sistema
imunológico através do qual os anticorpos são produzidos em resposta a um antigeno.
A imunidade celular envolve mecanismos através dos quais células infectadas com
agentes estranhos, tais como vírus, são destruídas, o que é feito diretamente por um
efetor (ex. célula T ativada) em contato com a célula alvo (Weinstock et al., 1989).
Os sistemas linfóides e não linfóides constituem duas amplas categorias estruturais
do sistema imunológico das aves.
Entre os componentes linfóides, a bursa
de Fabricius, sitio de desenvolvimento e diferenciação de linfócitos B e T, são
considerados órgãos linfóides primários, enquanto que o baço é considerado órgão
linfóide secundário. Além disso, estruturas linfóides distribuídas ao longo do intestino
representam uma barreira imunológica. As estruturas do sistema imunológico
representam uma barreira imunológica importante porque muitos patógenos economicamente importantes replicam no epitélio intestinal. Os componentes não linfóides do
sistema imune incluem células que propiciam uma defesa imunológica não específica
para o hospedeiro. Atuando como primeira linha de defesa imunológica, as células do
1
sistema fagocítico mononuclear são as que atuam primeiro nesta categoria. Monócítos
sanguíneos e macrófagos são importantes devido a sua ampla distribuição nos fluídos
corpóreos, órgãos e cavidades (Quershi et al., 1998).
Tendo natureza fagocítica, os macrófagos de aves se ligam, internalizam e
degradam antigenos, tais como bactérias. Por exemplo, macrófagos destroem mais
do que 80% da salmonela internalizada dentro de 15 minutos após o engolfamento
(Quershi et al., 1986). Além disso, células não linfóides com potencial fagocítico em
aves incluem heterófilos (neutrófilos em mamíferos) e trombócitos (Chang e Hamilton,
1979; Harmon et al., 1992).
Klasing (1998) comenta que fatores relacionados a genética das aves, a freqüência
de sua exposição a patógenos e a eficácia dos programas de vacinação são
detrimentes a incidência de doenças infecciosas em aves. Entretanto, características
dietéticas podem modular a suceptibilidade das aves para doenças infecciosas
devido ao nível de nutrientes ou dos tipos de ingredientes utilizados nas dietas que
eventualmente podem apresentar importância crítica.
Do ponto de vista prático, a susceptibilidade da ave a uma infecção pode ser
subdividida em dois componentes, resistência e persistência de produção. Resistência
se refere a capacidade de uma variedade de sistemas anatômicos e fisiológicos,
incluindo o sistema imune em excluir patógenos. Persistência de produção se refere
a capacidade manter a sua produtividade (ex. crescimento, eficiência alimentar,
produção de ovos) durante o desafio sanitário (Klasing, 1998). Há muitas situações
nas quais as dietas (incluindo níveis vitamínicos) que maximizam o desempenho
produtivo, não propiciam a máxima proteção imunológica. Isto pode ser facilmente
verificado ao examinar tabelas de exigências nutricionais que otimizam o desempenho
produtivo em condições de ausência quase que completa de desafio sanitário.
Em uma apreciação sobre a interação entre mecanismos de modulação nutricional
e resistência a doenças infecciosas, Klasing (1998) estabeleceu o papel de nutrientes
e ingredientes em várias categorias:
Tabela 1 — Mecanismos de modulação nutricional e resposta
imune
Mecanismo
Nutrientes
Desenvolvimento do sistema imune Ácido linoleico,
Fe, Vit. A
Substrato para o sistema imune
Todos nutrientes
Imunidade nutricional
Fe e biotina
Alteração hormonal
Energia e proteína
Ação regulatória direta de nutrientes Vit. A, D, E, ácidos
sobre o sistema imune
graxos poliinsaturados
Redução de patologia
Vit. E
Ação fisico-química de alimentos
Fibra, lectinas,
no intestino
lipideos peroxidados
Klasing (1998)
É importante ressaltar que a deficiência crônica severa de micronutrientes
(vitaminas e minerais) são mais debilitantes ao sistema imune do que energia
2
e proteína. O presente trabalho examinará o papel da nutrição vitamínica na
determinação da resistência a doenças das aves e sua relação com a produção
animal.
Vitamina E
A vitamina E é um termo utilizado para descrever duas gamas de compostos:
tocoferois e tocotrienóis. A diferença estrutural básica entre eles reside no número
de ligações duplas da cadeia lateral. Esta particularidade resulta em alteração na
atividade biológica da vitamina. Em geral, considera-se o α-tocoferol como padrão de
comparação para a determinação da atividade de outras formas de vitamina E.
A vitamina E pode ser encontrada na forma natural ou sintética. Ela é encontrada
em óleos vegetais, ovos, fígado, legumes e, em geral, plantas verdes. Já as
formas comerciais mais comuns são o acetato de d-α-tocoferol ou acetato de
d-a-tocoferol. A forma esterificada confere estabilidade a vitamina, mas não permite
ação antioxidante. É importante salientar que a forma alcoólica (tocoferol), que é a que
ocorre naturalmente, está sujeita a destruição no trato digestivo. Na forma esterificada
(acetato), isto não ocorre. O acetato é prontamente clivado na parede intestinal e a
forma alcoólica é então absorvida, permitindo assim que a vitamina exerça a sua ação
antioxidante (McDowell, 1989).
Por sua natureza lipossolúvel, a vitamina E está localizada em nível de membrana,
sabidamente de constituição lipoprotéica. Estruturalmente, ela reside entre os ácidos
graxos componentes de fosfolipídeos. Ali localizada, a vitamina E exerce a sua
função mais conhecida, qual seja a de antioxidante natural. Nesta função também
atuam outras substâncias lipossolúveis denominadas de carotenóides. Além destes
compostos, este papel biológico é coadjuvado intracelularmente por compostos
reconhecidamente de natureza hidrossolúvel como a vitamina C e a glutationa
peroxidase dependente de selênio.
Todos os elementos do sistema antioxidante interagem entre si de forma eficiente.
Esta interação provavelmente inicia ao nível de absorção de nutrientes e se
continua no metabolismo. Por exemplo, o selênio dietético poupa a vitamina E, de
forma que galináceos (Thompson e Scott, 1970) e patos (Dean e Combs, 1981)
apresentam concentrações mais elevadas de vitamina E no plasma ao receberem
dietas suplementadas com selênio. Por outro lado, a vitamina E mantém o selênio
no organismo de uma forma ativa, impedindo a sua perda do organismo. Além
disso, ao impedir a destruição dos lipídeos de membrana, impedindo a produção de
hidroperóxidos, reduz a quantidade de glutationa peroxidase necessária para destruir
peróxidos dentro do citosol das células (McDowell, 1989). A atividade da glutationa
peroxidase é superior quando o animal recebe selênio orgânico comparativamente a
inorgânico (Anônimo, 2000). Este aspecto pode justificar o fato de que concentrações
de vitamina E no fígado, cérebro e plasma dos pintos de um dia de idade
apresentam aumento significativo em decorrência da suplementação materna com
selênio orgânico (Surai e Sparks, S D).
A carência destes nutrientes em pintos tem sido relacionada à diátese exudativa
e distrofia muscular (vitamina E e selênio) e encefalomalácea (vitamina E) (Combs,
1991). No caso em específico da vitamina E, Combs e Scott (1974) observaram
3
que no mínimo 30 UI de vitamina E/kg de dieta eram necessários para minimizar a
susceptibilidade de microssomas hepáticos contra a peroxidação. Já em perus, Sell
(1996) demonstrou que eritrócitos das aves recebendo dietas sem a suplementação
de vitamina E apresentavam 100% de hemólise, enquanto que 60 ppm de vitamina E
eram necessários para manter a integridade da membrana das células vermelhas do
sangue.
As células do sistema imunológico e fagocitário são caracterizadas por apresentarem mitose acelerada podendo sofrer alterações provocadas por radicais livres,
peróxidos e superóxidos. Esta situação é freqüentemente observada em processos
de fagocitose, onde macrófagos produzem e secretam compostos reativos para
atacar organismos invasores. Parte deste efeito refere-se a produção de óxido
nítrico produzido por macrófagos ativados, resultando em molécula imunoregulatória
importante e citotóxica para células tumorogênicas e microorganismos (Lorsbach
et al., 1993; Nathan e Xie, 1994). A síntese de óxido nítrico por macrófagos de
aves é inibida durante infecção viral (Lyon e Hinshaw, 1993). Além disso, Gore e
Qureshi (1997) observaram que macrófagos de embriões de aves recebendo 10 UI de
vitamina E no ovo produziram mais ácido nítrico quando expostos a lipopolissacarídios
bacterianos in vitro. Os autores postularam que a vitamina E pode aumentar a
afinidade dos receptores da membrana de macrófagos para ativação, tal como
lipopolissacarídios, ou aumenta a atividade de síntese de óxido nítrico, resultando em
maior produção de óxido nítrico. Assim, a inoculação de vitamina E no ovo aumenta
o sistema imune humoral e celular. Infelizmente, estes compostos não só destroem
os agentes invasores como também produzem lesões nas suas próprias estruturas e
em tecidos adjacentes. A vitamina E neutraliza o efeito autodestruidor que o oxigênio
reativo exerce ao manter a integridade celular (Finch e Turner, 1996).
Mecanismo de ação alternativo da vitamina E junto ao sistema imunológico esta
relacionado com a sua participação na síntese de eicosanóides que modulam a produção de prostaglandinas e leucotrienos. As prostaglandinas são imunosupressoras,
enquanto que os leucotrienos são estimulantes do sistema imunológico (Still, 1995).
Tendo em vista que os macrófagos são o principal tipo de célula responsável pela
produção de prostaglandinas (Aderem et al., 1985), a vitamina E apresenta papel
vital na redução da produção de prostaglandinas ao antagonizar a peroxidação do
ácido araquidônico, limitando a entrada de precursores na via das prostaglandinas.
Likoff et al. (1981) demonstraram que a função fagocítica de aves recebendo 300
ppm de vitamina E era aumentada. Este fenômeno foi atribuído a redução nos
níveis endógenos de prostaglandinas em pintos arraçoados com dietas contendo altos
níveis de vitamina E, comparativamente aos animais que receberam dietas sem a
suplementação desta vitamina.
O terceiro envolvimento da vitamina E com o sistema imunológico está associado
com a síntese de interferon, que é uma glicoproteína que inibe de maneira não
específica a replicação viral. Quando uma célula for invadida por um vírus, o seu
material genético desencadeia a produção de interferon. Este tem a capacidade de
sair da célula infectada e situar-se em células vizinhas, nas quais é capaz de induzir
um estado antiviral ao gerar uma diminuição da atividade nuclear que impede, ou pelo
menos dificulta seriamente a invasão de células vizinhas por outro vírus. Franchini
et al. (1990) registraram que, após a vacinação com antígenos indutores de interferon
para vírus de Newcastle e Pasteurella antipestifer, os pintos arraçoados com 325 ppm
4
de vitamina E apresentaram maior síntese de interferon do que os arraçoados com
dietas contendo 25 ppm de vitamina E.
Efeito da vitamina E na dieta da matriz sobre a imunidade do pinto
A proteção da ação oxidativa já começa no período embrionário, onde a dieta
materna apresenta efeito determinante. Este efeito não só ocorre no período
embrionário, mas também durante o período inicial pós-eclosão (Surai, 2000). O autor
observou que dietas de matrizes contendo altos níveis de vitamina E (100 a 200 ppm)
propiciaram aumento na concentração de glutationa no fígado do recém nascido. A
glutationa é considerada um dos antioxidantes hidrossolúveis mais importantes para a
célula (Bains e Shaw, 1997) e sua concentração elevada pode ser considerada como
um indicativo de aumento da proteção antioxidante aos tecidos.
Nos tecidos das aves, um balanço entre a atividade pró-oxidante e antioxidante
apresenta papel crucial no desenvolvimento e viabilidade do pinto. Assim, tendo
em vista o efeito protetor da vitamina E contra a peroxidação dos lipídeos durante
o desenvolvimento embrionário e durante o período pós-eclosão, a possibilidade de
enriquecer a gema e tecidos embrionários pelo selênio e vitamina E é um fator a ser
considerado na suplementação da dieta da matriz de corte.
O entendimento da importância do papel da vitamina E na dieta da matriz e seu
possível efeito benéfico em pintos pós-eclosão foi provavelmente a razão de aumentar
o nível desta vitamina na dieta de matrizes de corte de 28 ppm em 1988 para 100 ppm
em 1995 (Surai, 1999). Dentro deste contexto, Sell (1996) comentou que os níveis de
vitamina E eram inadequados durante as 3 primeiras semanas pós-eclosão em perus.
Assim, níveis mais elevados de vitamina E deveriam ser considerados na dieta das
matrizes de perus.
A administração de vitamina E pode ser feita através da dieta ou parenteralmente.
A suplementação de vitamina E aumenta a proliferação de linfócitos. Haq et al. (1996)
suplementaram ou não dietas de matrizes de corte com níveis de 300 ppm de vitamina
E, associado ou não com β-caroteno. Os autores não observaram diferença no ganho
de peso e titulo de anticorpos em pintos com três semanas de idade. Entretanto, os
pintos oriundos de matrizes suplementadas com vitamina E apresentaram maior nível
de anticorpos a 1 e 7 dias de idade dos pintos. Estes dados são semelhantes aos de
Jackson et al (1978) que demonstraram melhora no título de anticorpos em pintos a 2
e 7 dias de idade, quando as matrizes imunizadas receberam altos níveis de vitamina
E (150 ppm). Portanto, é possível que a suplementação de vitamina E sobre o sistema
imune seja mais significativo durante os primeiros dias de vida.
Tengerdy et al (1972) registraram que pintos e matrizes arraçoados com dietas
suplementadas com 130 ppm de vitamina E apresentaram aumento significativo na
resposta imune, medido pelo teste de hemoaglutinação, comparativamente a aves
arraçoadas sem a suplementação de vitamina E.
Tengerdy e Nockels (1973) suplementaram (132 ppm vitamina E) dietas de
matrizes Leghorn e observaram aumento na resposta humoral em pintos a 2 e
4 semanas de idade. A mesma suplementação não apresentou efeito sobre a
eclodibilidade. Os autores concluíram que o nível de 15 a 30 ppm de vitamina E é
5
adequado para otimizar a eclodibilidade e produção de ovos, mas níveis de 100 a 150
ppm de vitamina E são recomendados para a resistência máxima a doenças.
Portanto, a imunidade passiva é um fator de grande importância na resistência das
aves a doenças, especialmente nas primeiras semanas de vida. As matrizes devem
ser alimentadas não só para maximizar os parâmetros produtivos, mas também para
otimizar a resposta imune da progênie.
Baseado nas propriedades da vitamina E em melhorar a resposta imunológica,
Gore e Qureshi (1997) avaliaram a inoculação de vitamina E no ovo e resposta imune.
Os autores demonstraram um aumento na resposta de anticorpos e macrófagos e
sugeriram que a inoculação de vitamina E no ovo melhora a qualidade de pintos e
perus pós-eclosão.
Importância da vitamina E sobre a potencialização das
vacinas
Tengerdy e Brown (1977) arraçoaram pintos com dietas contendo altos níveis de
vitamina E (300 mg/kg) ou vitamina A (60 000 UI/kg). O fornecimento individual
de cada um dos nutrientes em separado, mas não na forma combinada, reduziu
a mortalidade causada por E. coli. Esta proteção foi atribuída a um aumento
na produção de anticorpos e aumento na fagocitose. A proteção funcionou bem,
independente da imunização, mas tendo em vista que a proteção foi maior nas
imunizadas, os autores sugeriram o uso destas vitaminas junto a vacinas, quer seja
na forma de suplemento dietético, quer seja junto à vacina por si só. Vários trabalhos
confirmaram esta hipótese (Franchini et al., 1986, 1988, 1991 e 1995).
Em um destes estudos, Franchini et al. (1991) inocularam frangos (21 dias de
idade) com vacinas para Newcastle e E. coli. Dentro do conteúdo vacinal, a fração
óleo mineral foi parcialmente substituída por vitamina E, na forma de acetato de
α-tocoferol. A vitamina E aumentou a resposta imune para o antígeno viral, mas
não para o bacteriano quando a sua presença na fase oleosa não excedeu a 30%.
É possível que a vitamina E possa ter aumentado a resposta imune ao interagir com
células imuno-competentes envolvidas na reação inflamatória.
Franchini et al (1995) avaliaram se a vitamina E adicionada a vacinas inativadas e
emulsificadas aumentavam a resposta imune a antigenos virais (Newcastle, síndrome
da queda de postura e Gumboro) em aves. A fração óleo mineral foi substituída em
10, 20 e 30% pela vitamina E. Os autores observaram que vacinas com vitamina E,
especialmente quando o nível de substituição do óleo mineral foi de 20 a 30% pela
vitamina E, propiciou uma resposta na produção de anticorpos mais rápida e maior que
a vacina aplicada sobre um grupo controle. Isto provavelmente esta relacionado com
um bom balanço entre efeito inflamatório do óleo mineral e a influência da vitamina E
na atividade de células que atuam no sistema imunológico (linfócitos e macrófagos)
no ponto de inoculação viral.
6
Vitamina E durante o período pós-eclosão e imunidade
Conforme supracitado, a vitamina E é importante no desenvolvimento do sistema
imunológico da ave, ao aumentar, por exemplo, a proliferação de linfócitos (Corwin
e Gordon, 1982). A vitamina E também estimula a proliferação de linfócitos T,
particularmente as “helper T- cells” (Tanaka et al., 1979), inibe o efeito de substâncias
que impedem a mitogênese de linfócitos T (Corwin e Gordon, 1982) e influencia
positivamente a cooperação entre linfócitos T e B (Tanaka et al., 1979).
Likoff et al. (1981) avaliaram o uso de uma dieta com 300 ppm de vitamina E
sobre a mortalidade, imunidade humoral e fagocitose em pintos infectados com E.
coli. Os autores observaram uma melhora na resposta imune, medida por meio
de anticorpos aglutinantes (imunidade humoral) e número de células no sangue
(atividade fagocítica). Este trabalho foi corroborado pelo de Franchini et al. (1986), que
observaram que a produção de anticorpos responde positivamente a suplementação
de níveis crescentes de vitamina E. Os autores demonstraram que o benefício
máximo sobre o sistema imune de aves expostas ao vírus de Newcastle e Pasteurella
antipestifer foi obtido quando estas se alimentavam com níveis altos de vitamina E
durante os primeiros 28 dias. Após este período, o efeito benéfico sobre o sistema
imune é menor.
Colnago et al. (1984) avaliaram o efeito da suplementação de 100 ppm de vitamina
E sobre o rendimento produtivo e lesões causadas por E. tenella. A suplementação
com vitamina E propiciou um maior ganho de peso e menor presença de lesões no
ceco do que em frangos não imunizados. Embora de forma não significativa, os pintos
não imunizados e suplementados com vitamina E apresentaram melhor eficiência
alimentar que os não suplementados. Já no trabalho de Heinzerling et al. (1974),
a adição de 250 a 300 ppm de vitamina E aumentou a proteção dos pintos contra a
infecção por E. coli.
Benefício econômico da suplementação da vitamina E
A utilização de teores dietéticos mais elevados de vitamina E em nível de aviário
comercial só é possível de justificar caso ocorrer retorno financeiro. Dentro deste
contexto, Mclroy et al. (1993) observaram que o rendimento de aves naturalmente
expostas a infecção subclínica de Gumboro foram superiores quando estas se
alimentaram com dietas que continham 178 ppm de vitamina E, ao comparar com
aves recebendo 48 ppm de vitamina E. Os autores registraram que os lotes com
problema infeccioso apresentaram um retorno econômico de 4,56% a mais com a
suplementação superior de vitamina E.
Vitamina A
Aves deficientes em vitamina A ficam mais propensas a aumento na freqüência
e severidade de infecções bacterianas, virais e de protozoários. Parte da resistência
a doença devido a vitamina A está relacionada a manutenção da mucosa das membranas e funcionamento das glândulas adrenais para produção de corticosteróides
7
necessários para prevenir as doenças. A capacidade de um animal resistir a uma
doença depende do sistema imune, sendo que a deficiência de vitamina A propicia
uma redução do sistema imune (McDowell, 1989).
Em experimentos conduzidos em animais domésticos e de laboratório, os efeitos
das deficiências clínicas e subclínicas da vitamina A sobre a produção de anticorpos e
resistência de diferentes tecidos contra a infecção microbiana tem sido demonstrados.
Um suprimento inadequado de vitamina A aumenta a incidência de infecções
espontâneas nos animais. Krishnan et al (1974) demonstraram que pintos deficientes
em vitamina A apresentaram perda de linfócitos. A mortalidade devido a Salmonella
gallinarum foi reduzida ao suplementar vitamina A. Além disso, os níveis de anticorpos
no soro de pintos foi aumentado de 2 a 5 vezes ao suplementar altos níveis de vitamina
A Davis e Sell (1989) compararam o efeito do retinol e do ácido retinóico sobre o
sistema imunológico de frangos de corte. A resposta em IgG, IgM, IgA séricos e IgA
biliar foi superior em pintos recebendo 2 mcg de ácido retinóico/g da dieta do que em
aves recebendo quantidades equivalentes de vitamina A em forma de retinol. Sklan
et al. (1994) estabeleceram que níveis de até 6660 mcg de vitamina A/kg da dieta
propiciaram aumento na produção de anticorpos de pintos de corte.
Eventos importantes que atuam sobre a imunocompetencia começam no período
de desenvolvimento embrionário e continuam durante a primeira semana pós eclosão.
A primeira semana de vida é um período de rápida expansão da população de
leucócitos (órgãos linfóides) e eventos que produzem clones de linfócitos que
medeiam a imunidade com o avançar da idade. Trata-se de um momento crítico
durante o qual deficiências nutricionais ou excesso de nutrientes podem influenciar
o sistema imune. Deficiências nutricionais que prejudicam o desenvolvimento do
sistema imune incluem, além da vitamina A, o ácido linoleico, o ferro, selênio e
vitaminas do complexo B (Cook, 1991; Latshaw, 1991; Dietert et al., 1994). Por
exemplo, é sabido que o desenvolvimento embrionário do pinto é muito sensível à
deficiência de vitamina A, além do que pintos oriundos de matrizes deficientes em
vitamina A apresentam imunidade prejudicada e redução na resistência para uma
gama de doenças infecciosas. Entretanto, surpreendentemente Coskun et al (1998)
estudando os efeitos de níveis dietéticos da vitamina A sobre a produção de ovos e
resposta imunológica de matrizes semi-pesadas, observaram que a suplementação
de vitamina A não era necessária em premix para alcançar o máximo desempenho e
resposta imunológica. Além disso, os autores argumentam que a suplementação de
vitamina A na dieta das matrizes não melhorou a imunidade materna dos pintos.
O nível de vitamina A que maximiza o crescimento e a eficiência alimentar de
frangos de corte (500 mcg/kg) é insuficiente para o desenvolvimento do sistema
imune. Um nível para o crescimento máximo dos pintos em condições ótimas,
sem desafio sanitário, de instalações em universidades foi utilizado para estabelecer
exigências do NRC, mas níveis 10 a 20 vezes superiores são necessários para
maximizar a imunocompetencia (Sklan et al., 1994; Friedman e Sklan., 1994).
Entretanto, excesso de vitamina A pode prejudicar igualmente a imunocompetencia,
provavelmente por causar deficiências secundárias de outras vitaminas lipossolúveis
(Veltman et al., 1984). Está claro que o sistema imune é sensivel a deficiências
como a excesso de nutrientes. O mecanismo através do qual as vitaminas A e
D influenciam o desenvolvimento do sistema imune é provavelmente através de
8
influencias diretas sobre a diferenciação de células precursoras (Kline e Sanders,
1993; Woods et al., 1995).
A vitamina A, juntamente com as vitaminas E e D apresentam papel regulatório
sobre as células do sistema imune. Em roedores, níveis dietéticos acima das
exigências máximas de crescimento, mas dentro de um nível aceitável, apresentam
propriedades imunomodulatórias. Influencia semelhante parece ocorrer em aves.
Segundo Halevy et al. (1994), a vitamina A, agindo através de receptores para o ácido
retinóico, aumenta a resposta de linfócitos T para antigeno específico. Além disso,
a produção de citocinas, blastogênese de linfócitos, células natural killer e fagocitose
aumentam ao elevar os níveis de vitamina A na dieta (Ross, 1992).
O beta-caroteno é considerado uma pró-vitamina A, porque o corpo pode
transformá-la em forma de vitamina A ativa (McDowell, 1989). Okotie-Eboh e Bailey
(1991) avaliaram os efeitos do beta-caroteno e da cantaxantina sobre a resposta
imunológica de galos leghorn recebendo altas doses de aflatoxinas. Os autores não
observaram qualquer alteração na produção de anticorpos e peso relativo da bursa
de Fabricio devido a suplementação de beta-caroteno. Portanto, o beta-caroteno não
reduz a suceptibilidade dos galos para aflatoxicose.
Vitamina D
A vitamina D fornecida através da dieta ou produzida na pele por irradiação do
7-desidrocolesterol por ação da luz ultravioleta é convertida a sua forma hormonal
ativa (calcitriol) através de uma seqüência de hidroxilações no fígado e rins (Norman
et al., 1982). O calcitriol não é armazenado nos tecidos (Reddy e Tserng, 1989).
O calcitriol atua principalmente no osso, intestino e rins (Norman et al., 1982).
Entretanto, existem receptores para a vitamina D e seus metabólitos em células
do sistema imunológico, incluindo monócitos ou macrófagos (Bhalla et al., 1983;
Provvedini et al., 1983; Manolagas et al., 1985). Células linfóides também expressam
receptores para o calcitriol, mas somente em alguns estágios de diferenciação (Bhalla
et al., 1983). Mamíferos deficientes em vitamina D apresentam redução na ativação
de macrófagos, quimiotaxia e produção de citocinas (Aslam et al., 1998).
A manifestação da deficiência de vitamina D é um fenômeno complexo. Em geral,
os animais são considerados deficientes caso demonstrarem raquitismo, redução no
transporte intestinal de cálcio, hipocalcemia ou níveis não detectáveis de 25 (OH)D3
ou 1,25 (OH)2 D3 (Walters et al., 1992). Em frangos de corte, Aslam et al. (1998)
demonstraram que a deficiência de vitamina D altera a resposta imunológica. O peso
do timo, ajustado para peso corporal, foi menor em pintos deficientes em vitamina
D. Os autores também observaram redução de macrófagos e concluiram que a
deficiência de vitamina D deprime a resposta imune celular.
Ácido ascórbico
O ácido ascórbico melhora a resposta imunológica e a resistência a doenças em
aves (Pardue et al., 1985; Rund, 1989). Na primeira linha de defesa contra patógenos,
a fagocitose por neutrófilos envolve o aumento da utilização de ácido ascórbico e
9
desidroascórbico (Stankova et al., 1975; Rund, 1989). Além disso, infecções virais
causam depleção de ácido ascórbico em leucócitos, resultando em vários graus de
imunossupressão não específica (Thomas e Holt, 1978). O ácido ascórbico pode
modular a atividade das células B e a adição de ácido ascórbico na dieta antes da
imunização resulta em aumento da produção de anticorpos (McCorkle et al., 1980).
Amakye-Anim et al. (2000) concluiram que a suplementação de 1000 ppm de ácido
ascórbico na dieta apresenta efeitos benéficos na produção de anticorpos em aves
vacinadas contra gumboro.
A vitamina C e a vitamina E interagem metabolicamente. A vitamina C melhora
a atividade antioxidante da vitamina E ao reduzir os radicais de tocoferoxila para a
forma ativa da vitamina E (Jacob, 1995) ou ao poupar a vitamina E disponível (Retsky
e Frei, 1995). Yin et al. (1993) registraram que uma mistura de alfa-tocoferol e ácido
ascórbico retardava a oxidação de mioglobina, enquanto que o ácido ascórbico e o
alfa-tocoferol sozinhos não retardavam a formação de metamioglobina. Schaefer et al.
(1995) demonstraram que a oxidação de mioglobina esta propensa a retardar quando
o radical de alfa-tocoferoxil na face interna do retículo sarcoplasmático é reduzida pelo
ascorbato. Com relação a atividade antioxidante, existe um efeito sinérgico da vitamina
E e C sobre a resposta imunológica. A resposta imunológica de cobaias aumenta
quando receberam dietas contendo níveis elevados de vitaminas E e C (Bendich
et al., 1984). Gonzalez-Veja-Aguirre et al. (1995) demonstraram que a combinação
de 200 ppm de vitamina C e 75000 UI/tonelada de vitamina E melhorava os níveis de
anticorpos de frangos de corte contra a Brucella abortus e para o vírus vivo e morto
da Newcastle. Além da ação antioxidante, a vitamina C propicia melhora na resposta
imunológica ao modificar a síntese de corticosteróide nas glândulas adrenais (Pardue
e Thaxton, 1984).
Puthpongsiriporn et al (2001) avaliaram o efeito da suplementação das vitaminas E
a C sobre a proliferação de linfócitos em poedeiras expostas a estresse por calor.
Os autores concluíram que a suplementação de 65 UI de vitamina E/kg da dieta
melhorava a resposta imunológica durante o estresse de calor ao elevar a proliferação
de linfócitos. A suplementação de 65 UI/kg de acetato de dl-alfa-tocoferol e 1000 ppm
de vitamina C propiciou uma melhora mais acentuada na proliferação de linfócitos
em aves expostas a estresse térmico do que aquelas que receberam somente a
suplementação de vitamina E.
Complexo polivitamínico
Vários autores (Thaxton e Siegel, 1970; Brake, 1989; Siegel, 1989; Miller e
Quershi, 1991) demonstraram que aves expostas a estresse ambiental de várias
naturezas apresentavam depressão do sistema imunológico. O desempenho e a
função imunológica de aves submetidas a estresse térmico melhora significativamente
ao aumentar os níveis de vitamina C (Pardue e Thaxton, 1984; Pardue et al., 1985),
vitamina E (El-Boushy, 1988) e piridoxina (Blalock et al., 1984). A administração
de vitamina E em excesso apresenta um efeito estimulatório na atividade fagocítica
do sistema reticuloendotelial (Heinzerling et al., 1974) e na produção de anticorpos,
particularmente IgG (Jerne e Nordin, 1963; Tengerdy et al., 1973).
10
Ferket e Qureshi (1992) avaliaram a imunidade de frangos de corte expostos
a estresse térmico recebendo níveis elevados de vitaminas (vitaminas A, D, E e
complexo B) e minerais na água. Os autores observaram melhora na produção de
anticorpos IgG e macrófagos, mas não na capacidade fagocítica de macrófagos nas
aves que receberam a suplementação acima.
Deyhim e Teeter (1993) submeteram frangos de corte a estresse térmico e
avaliaram o efeito da retirada de vitaminas e minerais da dieta durante o período de
28 a 49 dias de idade. A retirada dos premixes não afetou a compêtencia imunológica,
quando avaliada por títulos de anticorpos. É provavel que o conteúdo de vitaminas
e minerais presentes no milho e farelo de soja apresentassem níveis vitamínicos e
minerais suficientemente elevados para manter a resposta imunológica. Entretanto,
para que uma decisão desta natureza seja tomada, cabe avaliar a genética das aves
o desafio sanitário e o manejo oferecido.
Embora não avaliando diretamente parâmetros imunológicos, Coelho e McNaughton (1995) submeteram frangos de corte a diferentes condições de estresse
associados a diferentes níveis vitamínicos e avaliaram o desempenho produtivo
dos animais. As condições ambientais consistiram de cama usada, densidade de
aves, desafio com coccidiose, gordura peroxidada, contaminação com micotoxinas
e densidade da dieta. Os autores concluíram que em condições de alto desafio
ambiental, níveis vitamínicos superiores são necessários para o máximo desempenho
das aves.
Conclusão
O efeito benéfico da vitamina E sobre a resposta imunológica e doenças está bem
estabelecido. Para alcançar tal êxito, níveis dietéticos de 150 a 250 mg de vitamina
E/kg são recomendados. Estes níveis são superiores aos normalmente utilizados para
maximizar o desempenho produtivo. Para a vitamina A, níveis de 6660 mcg /kg e 1000
ppm de vitamina C são recomendados para máxima resposta imunológica.
Bibliografia
ADEREM, A. A.; S. D. WRIGHT.; S. C. SILVERSTEIN e Z. A. COHN. J. Exp. Med.,
161: 617–622 (1985).
AMAKYE-ANIM, J. T. L. LIN, P. Y. HESTER, D. THIAGARAJAN, B. A WATKINS e C. C.
Wu. Poultry Sci. 79:680–688 (2000).
ANÔNIMO. Feeding Times, 5(2): 20–21, (2000).
ASLAM, S. M., J. D. GARLICH E M. A QURESHI. Poultry Sci. 77: 842–849 (1998).
BAINS, J. S. e C. A. SHAW. Brain Res. Reviews, 25: 335–243 (2000).
BENDICH, A P., P. D. APOLITO, E. GABRIEL e J. L. MACHLIN. J. Nutr. 114:1588–
1593. (1984).
BHALLA, A K., E. P. AMENTO, T. L. CLEMENS, M. F. HOLICK e S. M. KRANE. J. Clin.
Endocrinol. Metab. 57:1308–1310. (1983).
BLALOCK, J. L. , J. P. THAXTON e J. D. GARLICH J. Nutr. 114:312–322 (1984).
BRAKE, J. T. Zootech. Int. January 1989:37–40. (1989).
11
CHANG, C. F. e P. B. HAMILTON. J. Reticuloendothelial Soc. 25: 585–590. (1979).
COELHO, M. B., J. L. McNAUGHTON. J. Appl. Poultry Research 4:219–229 (1995).
COMBS, G. F. Jr The Vitamins. Academic Press, INC. SanDiego. 528 p. (1991).
COMBS, G. F. Jr. E M. SCOTT. J. Nutr, 104: 1297–1303 (1974).
COOK, M. E. Crit. Ver. Poultry Biol. 3:167–190. (1991).
CORWIN, L. M. e R. R. GORDON. N. Y. Acad. Sci., 393: 437–451 (1982).
COSKUN, B., F. INAL, I. CELIK. Poultry Sci. 77:542–546. (1998).
DAVIS, C. Y. e J. SELL. Poultry Sci. 68:136–144 (1989).
DEAN, W. F. e G. F. COMBS. Poultry Science, 60: 2655 (1981).
DEYHIM, F. e R. G. TEETER. J. Appl. Poultry Res. 2:347–355. (1993).
DIETERT, R. R., K. A GOLEMBOSKI, R. E. Austic. Poultry Sci. 73:1062–
1076. (1994).
EL-BOUSHY, A R. Feedstuffs 60(4):20–26. (1988).
FERKET, P. R. e M. A QUERSHI. Poultry Sci. 71:88–97. (1992).
FINCH, J. M. e R. J. TURNER. Res. Vet. Sci., 60: 97–106 (1996).
FRANCHINI, A.; S. BERTUZZI.; G. MANFREDAL e A. MELUZZI. Archiv - fur Geflugelkunde, 54: 143–146 (1990).
FRANCHINI, A..; S. BERTUZZI.; C. TOSARELLI e G. MANFREDA. Poultry Sci., 74:
666–671 (1995).
FRANCHINI, A.; M. CANTI.; G. MANFREDA e S. BERTUZZI. Poultry Sci., 70:
1709–1715 (1991).
FRIEDMAN, A, D. SKLAN. World’s Poultry Sci. J. 53:186–195. (1997).
GONZALES-VEJA-AGUIRRE, D., B. P. A CONTRERAS, R. KLEIN e H. BOHMWALD.
Veterinária 26:333–340. (1995).
GORE, A. B. e M. A. QURESHI. Poultry Sci., 76: 984–991 (1997).
HAQ C.; A. BARLEY e A. CHINNAH. Poultry Sci, 75: 1092–1097 (1995).
HARMON, B. G., J. R. GLISSON e J. C. Nunnally. Avian Dis. 36:986–991. (1992).
HAVELY, O, Y. ARAZI, D. MELAMED. A D. FRIEDMAN e D. SKLAN. J. Nutr.
124:2139–2146. (1994).
HEINZERLING, R. H., C. F. NOCKELS, C. L. QUARLES e R. P. Tangerdy. Proc. Soc.
Exp. Biol. Med. 146:279–283. (1974).
JACKSON, D. W.; G. R. J. LAW e C. F. NOCKELS. Poultry Sci., 57: 70–73 (1978).
JACOB, R. A Nutr. Res. 15:755–766. (1995).
JERNE, N. K. e A A Nordin. Science 140:405 (1963).
KLINE, K. e B. G. SANDERS. Nutr. Cancer 19:241–252. (1993).
KLASING, K. C. Poultry Sci. 77: 1119–1125 (1998).
KRISHNAN, S., U. N. BHUYAN, G. P. TALWAR e V. RAMALINGASWAMI. Immunology
27, 383–392 (1974).
LATSHAW, D. J. Vet. Immunol Immunopathol. 30:111–120 (1991).
LIKOFF, R. O.; D. R. GUPTILL.; C. C. MEKAY.; M. M. MATHIAS.; C. F. NOCKELS e R.
P. TENGERDY. Am. J. Clin. Nutr., 34: 245–251 (1981).
LORSBACH, R. B.; W. J. MURPHY.; C. J. LOWENSTEIN. S. H. SNYDER e S. W.
RUSSELL. J. Biol. Chem, 268: 1908–1913 (1993).
LYON, J. A. e V. S. HINSHOW. Avian Dis, 37: 868–873 (1993).
McCORKLE, F. R., R. TAYLOR, E. SINSON, B. Glick Poultry Sci. 59:1324–
1327 (1980).
McDOWELL, L. R. Vitamins in Animal Nutrition. Academic Press. Sandiego. 486
pp. (1989).
12
MANOLAGAS, S. C., D. M. PROVVEDINI e C. D. TSOUKAS MOL. Cell. Endocrin.
43:113–122 (1985).
MARKS, J. A Guide to the Vitamins: Their role in Health and Disease MTP, Medical
and Tech. Publishing Co. Ltd. England. (1979).
MILLER, L., M. A Quershi. Vet. Immunol, Immunopathol. (1991).
NATHAN, C. Q. e Xie. Cell. 78: 915–918 (1994).
NORMAN, A W., J. ROTH e L. Orci. Endocrin. Review 3(4): 331–366 (1982).
OKOTIE-EBOH, G. O, C. a BAILEY, L. F. KUBENA Poultry Sci. 70 (Suppl):89. (1991).
PARDUE, S. L., J. P. THAXTON. Poultry Sci. 63:1262–1268. (1984).
PARDUE, S. L., J. P. THAXTON e J. BRAKE. J. Appl. Physiol. 58:1511–1516. (1985).
PROVVEDINI, D. M. C> D. TSOKAS, J. L. DEFTOS e S. C. MANOLAGAS. Science
221:1181–1183. (1983).
QUERSHI, M. A, R. R. DIETERT e L. D. BACON. Proc. Soc. Exp. Biol. Med.
181:560–568. (1986).
QURESHI, M. A, I. HUSSAIN, C. L. HEGGEN. Poultry Sci. 77:1126–1129. (1998).
REDDY, G. S. e K. Y. TSERNG. Biochemistry 28: 1763–1769. (1989).
RETSKY, K L. e B. FREI. Biochem. Biophys. Acta. 1257:279287. (1995).
ROSS, A C. Proc. Soc. Exp. Biol. Med. 200:303–320 (1992).
RUND, B. Poultry Dig. 48:44–55. (1989).
SCHAEFER< D. M. Q. LIU, C. FAUSTMAN, M. C. YIN. J. Nutr. 125(suppl. 6):S1792–
S1798. (1995).
SELL, J. L. Animal Feed Science and Tecnology, 60: 229–240 (1996).
SIEGEL, P. B. Stress and Immunity. Pg 157-160 In: Proceedings 14th National
Meeting on Poultry Health and Condemnations, College Park, MD. (1989).
SKLAN, D. , D. MELAMED e A FRIEDMAN. Poultry Sci. 73:843–847. (1994).
STANKOVA, L., N. B. GERHERDT, L. NAGEL, R. H. BIGLEY. Infect. Immunol.
12:252–255. (1975).
STILL. P. Pet Forum’95. April, 3–4, (1995).
SURAI, P. E. Poultry and Avian Biology Reviews, 10: 1–60 (1999).
SURAI, P. E. e N. H. C. SPARKS. Efeito do teor de selênio na dieta materna sobre
o sistema antioxidante do pinto recém-eclodido, (S D).
TANAKA, J.; H. FUJIWARA e M. TORISU. Immunology, 38: 727–734 (1979).
TENGERDY, R. P. e J. C. BROWN. Poultry Sci., 56: 957–963 (1977).
TENGERDY, R. P.; R. H. HEINZERLING.; G. L. BROWN.; M. M. MATHIAS. Int. Anch.
Allergy, 44: 221–232 (1973).
TENGERDY, R. P.; P. A. HEINZERLING e C.F. NOCKELS. Infection and Immunity, 5:
987–989 (1972).
TENGERDY, R. P., R. H. HEINZERLING, G. L. BROWN e M. M. MATHIAS Int. Arch.
Allergy Appl. Immunol. 44:221–231. (1973).
THAXTON, J. P., e H. S. SIEGEL Poultry Sci. 49:202–205. (1970).
THOMAS, W. R. e P. G. HOLT CLIN. Exp. Immunology. 32:370–379.
THOMPSON, J. N. e M. L. SCOTT. J. Nutr, 100: 797 (1970).
VELTMANN, J. R., L. S. JENSEN, G. N. Rowland Avian Dis. 29:446–452. (1984).
YIN, M. C., C. FAUSTMAN, J. W. RIESEN e S. N. WILLIAMS J. Food Sci. 58:1273–
1276. (1993).
WALTERS, M. R., U. KOLLENKIRCHEN e J. Fox. Proc. Soc. Exptl. Biol. Med.
199:385–393. (1992).
WEINSTOCK, D., K. A SCHAT e B. W. CALNET, 1989. Eur. J. Immunol. 19:267-272.
WOODS, C. C., C. DOMENGET, F. SOLARI. Endocrinology 136:85–95. (1995).
13