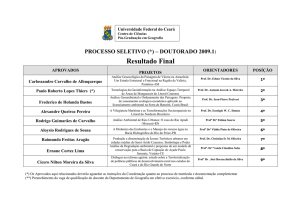
Curso de Embriologia Ocular / 2003 - Profª Drª Márcia Reis Guimarães
Hospital de Olhos de Minas Gerais – Belo Horizonte/MG
Introdução à Embriologia Ocular
O código genético é comum a todas as células e o que as diferencia entre si é a
porção do genoma que é expresso, segundo a “interpretação” das circunstâncias
que as rodeiam em dado momento da seqüência embriológica. Assim, cada uma
delas segue representando seu papel na sociedade multicelular que é o nosso
organismo.
A fecundação, que corresponde à união entre um óvulo e um espermatozóide leva
à formação de um ovo unicelular, que passa então a se dividir em dois, quatro, oito
e enfim, 16 células que passa a ser a mórula envolvida pela membrana pelúcida.
A liberação de fatores de crescimento que intervêm no metabolismo de
fosfolípides intracelulares (inositol), ativam enzimas do tipo proteína C – quinase
que vão alcalinizar o meio intracelular ativando síntese de DNA e proteínas. Além
disso, estes fatores influenciarão na coesão das células mais externas da mórula,
isolando-a do meio exterior enquanto as pressões osmóticas criadas pela saída dos
íons Na+ através das membranas celulares criam uma cavidade (blastocele) e um
pólo de células compactadas, o embrião.
Graças à elongação, adesão e contração, o pólo celular se transforma em uma
estrutura trilaminar com um eixo central e simetria bilateral: gástrula. Destas 3
lâminas se originarão os folhetos fundamentais comuns a todos os organismos
superiores: Ectoderma: Que origina a pele, SNC e periférico, epitélio sensorial do
olho, ouvido e nariz, glândulas mamárias, hipófise, glândulas subcutâneas e
esmalte dentário.
Endoderma: Que origina os intestinos e pulmões.
Mesoderma: Que origina os músculos, inclusive os extra-oculares, rins e ossos.
A morfogênese destes 3 folhetos germinativos depende de 4 fenômenos de
indução a nível molecular:
a) modificação do citoesqueleto celular condicionando a deformação da célula
e seus movimentos;
b) coordenação entre as células por um sistema de comunicação intercelular
(gap junctions);
c) as aderências intercelulares (glicoproteínas, neuroglia, etc., conhecidos
como moléculas de adesão celular – CAM ou Cell Adhesion Molecules)
d) o substrato pelo qual se deslocam (papel das fibronectinas e seus
receptores).
As CAM têm papel fundamental nos deslocamentos e posicionamento dos folhetos
embrionários. Graças à uma matriz rica em fibronectinas é possível o
deslocamento de um grupo de células próximo ao ectoderma, a crista neural que
Profª Drª Márcia Reis Guimarães Pág 1
Curso de Embriologia Ocular / 2003 - Profª Drª Márcia Reis Guimarães
Hospital de Olhos de Minas Gerais – Belo Horizonte/MG
irá se distribuir estrategicamente ao longo dos folhetos embrionários originando os
melanócitos, o sistema nervoso periférico e estruturas oculares fundamentais.
Neste esquema observarmos a variação da concentração molecular das adesinas
(CAM) e fibronectinas durante as 5 fases da morfogênese.
A: determinação na periferia da placa neural
B: células da crista neural se isolam da placa neural
C: migração celular
D: condensação das células após a constituição dos gânglios nervosos
E: diferenciação dos neurônios
LCAM:
glicoproteínas contráteis presentes nas células embrionárias do fígado (L=fígado)
fazem parte da família das adesinas, juntamente com a N. caderina
NCAM:
glicoproteínas presentes nas células do sistema nervoso central e periférico tendo
características semelhantes às imunoglobinas. A função principal é promover a
adesão celular.
FN:
fibronectina
NgCAM: glicoproteínas das células da neuróglia
A análise destas curvas de concentração nos mostra que a fibronectina
intermédia, por mecanismos de adesão e/ou migração, o comportamento da crista
neural. Esta massa de células, localizada lateralmente à placa neural, se comporta
ora como células migratórias isoladas ora como uma massa epitelial. A
fibronectina é tão importante que se injetarmos precocemente anticorpos
monoclonais anti-fibronectina não ocorre a gastrulação. Em sua etapa mais
avançada, haveria o alongamento do ectoderma e mesoderma, porém, este último
Profª Drª Márcia Reis Guimarães Pág 2
Curso de Embriologia Ocular / 2003 - Profª Drª Márcia Reis Guimarães
Hospital de Olhos de Minas Gerais – Belo Horizonte/MG
não se contactaria com o ectoderma nem haveria indução para formação da placa
neural.
A partir de um ovo único fecundado, a natureza vai permitir que, em algumas
semanas, seja elaborado um organismo multicelular composto de células
diferenciadas segundo um plano preciso. Os tipos celulares são distribuídos
geograficamente e na etapa seguinte, o de proliferação com diferenciação, cria-se
um ecossistema estabilizado onde certos tecidos se renovam, mas mantêm sua
diferenciação sem alterar a organização global.
O desenvolvimento intrauterino envolve a execução precisa de um programa de
apoptose; assim, o crescimento fetal emprega em sua sofisticada escultura a
modulação pela morte celular. As células de um organismo vivo estão em
intercomunicação constante com os tecidos e órgãos à distância através de um
intricado sistema de sinalização química. Estas informações são analisadas
continuamente e a célula opta por cometer o suicídio, uma cascata de morte se
instala com morte intracelular, fagocitose da célula apoptótica e a reabsorção dos
restos celulares.
O mecanismo de morte celular pela apoptose é geneticamente controlado e
por ela a célula ativa uma seqüência de instruções que leva à sua “desconstrução”
de dentro para fora; processo de evolução da própria vida, tal como as folhas que
caem no outono para a renovação da própria folhagem das árvores. Este código de
“morte celular” é modulado pelo próprio DNA que controla o desenvolvimento
dos organismos multicelulares através da eliminação estratégica de células
selecionadas SEM a ocorrência de edema, lise celular e inflamação como é o caso
na necrose.
Um exemplo clássico de apoptose é observado no crescimento das extremidades
do ser humano quando os dedos se separam pela remoção das membranas
interdigitais. Horovitz, Brenner e Sulston ganharam o Prêmio Nobel de Fisiologia
e Medicina em 2002 por suas contribuições neste campo. As proteínas mediadoras
da programação de morte programada, a apoptose, são as “caspases”. Estas
proteínas atuam como “tesouras” moleculares que quebram certas proteínas
fisiológica e estruturalmente importantes desencadeando a morte intracelular. A
degeneração macular senil, retinopatias congênitas, retinite pigmentar, doença de
Alzheimer, Aids, etc., são exemplos de patologias relacionadas à desequilíbrios no
mecanismo de apoptose. Outros exemplos serão mencionados no curso de
embriologia e malformações.
O desenvolvimento do ovo, fertilizado através dos estágios de clivagem, mórula,
blástula, gástrula e tubo neural, a formação das três camadas germinativas
(ectoderma, mesoderma e endoderma) com o estabelecimento dos órgãos
rudimentares primários, fazem parte do conhecimento básico na Embriologia
Geral.
Profª Drª Márcia Reis Guimarães Pág 3
Curso de Embriologia Ocular / 2003 - Profª Drª Márcia Reis Guimarães
Hospital de Olhos de Minas Gerais – Belo Horizonte/MG
Resumidamente podemos descrever a seqüência embriológica humana nas
seguintes etapas: crescendo interiormente, o embrião com 28 dias tem a forma
encurvada em “C”. Seu cérebro e coração primitivos estão muito ativos e o
embrião tem em sua porção apical um espessamento no tamanho de 1 cabeça de
alfinete, correspondente ao cérebro. Nos 3 arcos abaixo deste espessamento estão
os aparelhos essenciais para as futuras funções da audição, fala, respiração,
alimentação e expressão facial.
Como numa revelação fotográfica em banho de imersão; o embrião se torna mais
distinto a cada dia. Uma estria em curva aparece acima da saliência facial, trata-se
do ouvido primitivo. Nesta mesma fase, o cristalino se aprofunda na taça óptica
concluindo mais uma etapa no desenvolvimento do globo ocular.
Duas placas espessadas da camada ectodérmica externa original se expandem,
depois se dobram formando depressões rodeadas por anéis celulares em forma de
ferradura; estas se tornarão o nariz.
Profª Drª Márcia Reis Guimarães Pág 4
Curso de Embriologia Ocular / 2003 - Profª Drª Márcia Reis Guimarães
Hospital de Olhos de Minas Gerais – Belo Horizonte/MG
Com grande velocidade, o origami humano vai se organizando cada vez mais em
formas específicas, o cérebro é agora uma série de 5 cavidades revestidas por
tecido embrionário neural, e as células da parte apical iniciam sua diferenciação
formando o córtex cerebral, sede do intelecto. As depressões auditivas se
expandem e a nasal se aprofunda e sobe em direção aos olhos. Como o coração e o
cérebro até aqui eram mais necessários do que o aparelho digestivo, a parte
inferior do corpo se desenvolveu mais lentamente que a superior.
O fígado, estômago e esôfago começam a se desenvolver assim como projeções
dos ombros, membros e partes distais.
Células sexuais migram do saco vitelino para os locais onde os órgãos sexuais se
desenvolverão. A cauda vestigial atinge seu comprimento máximo antes de
começar sua involução completa.
O embrião humano ainda dobrado, com a cabeça e cauda a se tocar, parece ser o
mesmo que qualquer outro vertebrado no mesmo estágio. Por fora, poderia ser
uma ave, rato ou um porco, mas internamente a máquina embrionária está
totalmente ativada e direcionada à confecção de um ser humano e os efeitos
cumulativos deste trabalho começarão a aparecer neste ponto.
Os setores faciais começam a se juntar rapidamente, um sulco se forma próximo a
depressão nasal – futuro maxilar e lábio. Dois dias depois, as hemi-partes
superiores e inferiores se fundem simetricamente. Mais 48 horas, a musculatura
facial e os dentes iniciam sua formação com a involução dos arcos branquiais
vestigiais.
Os olhos iniciam sua pigmentação e menos de 24 horas após, os músculos
extraoculares – dentre os mais delicados do organismo – começam a sua
diferenciação.
Profª Drª Márcia Reis Guimarães Pág 5
Curso de Embriologia Ocular / 2003 - Profª Drª Márcia Reis Guimarães
Hospital de Olhos de Minas Gerais – Belo Horizonte/MG
Fibras nervosas conectam-se com região olfatória do cérebro, pavimentando a via
para o sentido do olfato.
Enquanto isso, o coração começa a se subdividir em câmaras bombeadoras
enquanto o fígado inicia a produção de células sangüíneas a partir do saco vitelino
que começa a se dissolver.
Se o 1º mês foi o do grande planejamento, no 2º a execução atinge seu ápice.
Embora haja riscos no desenvolvimento do embrião, estes já são bem menores à
medida em que o embrião assume cada vez mais sua forma humana.
Embriologia Ocular
O conhecimento da ontogenia adquire para o oftalmologista uma importância
muito além do aspecto acadêmico de sua formação, uma vez que nenhum outro
órgão do corpo humano as anomalias de embriogênese são tão aparentes quanto
em sua especialidade. O diagnóstico diferencial entre uma anomalia do
desenvolvimento e uma patologia adquirida torna-se em muitos casos questão
crucial e extremamente delicada pois a partir dele serão tomadas decisões clínicas
de considerável responsabilidade.
A seqüência da evolução embriológica pode ser dividida em 3 fases:
embriogênese, organogênese e diferenciação.
Na fase de embriogênese:
a) Na fase de embriogênese, individualização dos folhetos embrionários
(ecto, meso e endoderma) até o aparecimento do olho primitivo (1ª até a 3ª
semana após fecundação)
27 dias / 4mm
Profª Drª Márcia Reis Guimarães Pág 6
Curso de Embriologia Ocular / 2003 - Profª Drª Márcia Reis Guimarães
Hospital de Olhos de Minas Gerais – Belo Horizonte/MG
b) Na fase da organogênese, camadas se organizam em padrões específicos
formando órgãos (4ª até a 8ª semana).
c) Na fase da diferenciação, elaboração das estruturas características de cada
órgão, fase mais tardia que se prolonga até depois do nascimento como no
caso da retina (3º ao 9º mês) em geral, da mácula (16º mês) e fovéola (45º
mês) em especial.
Pela sua complexidade, uma vez que a seqüência embriológica se faz com eventos
em cascata e com interações tridimensionais, entraremos em descrições detalhadas
de algumas etapas da embriogênese, organogênese e diferenciação.
Durante a fase da embriogênese (1ª até o final da 3ª semana), ressaltamos alguns
detalhes: a migração das células da crista neural e a formação do tubo neural, a
região situada medialmente do mesoderma (chamada de mesoderma paraxial)
forma somitos pareados que se alinham ao longo de ambos os lados do canal
neural.
Lateralmente aos somitos, o mesoderma se divide em duas camadas, uma
associada ao ectoderma situado acima e a outra ao endoderma situado abaixo do
mesmo.
A – Mesoderma paraxial
B – Somitos ventrais
C – Área pericardíaca
D – Arco branquial
Profª Drª Márcia Reis Guimarães Pág 7
Curso de Embriologia Ocular / 2003 - Profª Drª Márcia Reis Guimarães
Hospital de Olhos de Minas Gerais – Belo Horizonte/MG
O sulco óptico está contido no prosencéfalo. O canal neural ainda não se fechou
anteriormente. A segmentação do mesoderma em somitos se inicia posteriormente
à cefálica da notocorda que fica debaixo do canal neural. O 1º par de somitos é o
occipital e cada um dos 42 pares se formam sucessivamente no sentido crâniocaudal.
As células da crista neural (CCN) aparecem dorsalmente, a partir das células
neuroepiteliais das extremidades laterais da placa neural.
Células da crista neural marcadas com timidina triciada
Profª Drª Márcia Reis Guimarães Pág 8
Curso de Embriologia Ocular / 2003 - Profª Drª Márcia Reis Guimarães
Hospital de Olhos de Minas Gerais – Belo Horizonte/MG
Durante a fase primária as células mesodérmicas e a CCN não tem diferenças
bioquímicas ou citológicas significativas. A distinção principal está na sua
capacidade de migração: enquanto as células mesodérmicas primárias ficam
imobilizadas, as CCN são capazes de ampla movimentação, graças à ação da
fibronectina.
Os tecidos do olho são derivados do ectoderma neural, crista neural, ectoderma
superficial e mesoderma. O termo mesênquima abrange o mesoderma e os tecidos
conjuntivos de origem diversa, como é o caso da crista neural. Os tecidos que se
originam da crista neural são chamados de mesectoderma. Os músculos extraoculares e os componentes vasculares particularmente o endotélio, tem origem no
mesoderma primitivo.
A formação do globo ocular acompanha o desenvolvimento do sistema nervoso
central e o esboço deste último ocorre ainda na 3ª semana de desenvolvimento,
partir do tubo neural.
No final da 3ª semana, a porção craniana do tubo neural apresenta dilatações
separadas por duas constrições. São as vesículas cerebrais primárias:
prosencéfalo, mesencéfalo e rombencéfalo. O olho primitivo, também chamado
de sulco óptico aparece como um par de estrias em cada lado da linha média da
placa neural antes de seu fechamento tubular, exatamente entre o diencéfalo e o
telencéfalo ao 22º dia da embriogênese. Com o aparecimento do sulco óptico
considera-se encerrado o período da embriogênese.
A organogênese perdurará entre a 4ª até o final da 8ª semana.
Ao final da 4ª para a 5ª semana, o prosencéfalo subdivide-se em telencéfalo
anteriormente e diencéfalo posteriormente.
Na 4ª semana o olho primitivo sofre uma escavação e se torna uma vesícula com a
formação de uma dobra inferior que se denomina fissura embrionária ou
coroidiana.
A vesícula e o canal óptico invaginam formando
a fissura coroidiana inferior
Profª Drª Márcia Reis Guimarães Pág 9
Curso de Embriologia Ocular / 2003 - Profª Drª Márcia Reis Guimarães
Hospital de Olhos de Minas Gerais – Belo Horizonte/MG
Esta fissura se fechará no centro e depois ântero-posteriormente até a íris e o nervo
óptico respectivamente ao fim da 6ª semana.
Como dissemos, as vesículas ópticas crescem e sofrem uma invaginação
adquirindo aspecto de taça. Ao se dobrar, o neuroectoderma passará a apresentar
uma dupla camada: as camadas interna e externa da futura retina. E entre elas,
persiste um espaço virtual intraretiniano que se comunica com o 3º ventrículo pelo
canal óptico.
A invaginação da vesícula afeta o canal ou haste óptica formando uma fissura, à
qual já nos referimos no parágrafo anterior como fissura embrionária.
A artéria central da retina, penetra no canal óptico através desta fissura.
Profª Drª Márcia Reis Guimarães Pág 10
Curso de Embriologia Ocular / 2003 - Profª Drª Márcia Reis Guimarães
Hospital de Olhos de Minas Gerais – Belo Horizonte/MG
A artéria hialóidea, futura artéria central da retina, penetra no canal óptico através
desta fissura. A invaginação da vesícula óptica está sincronizada com a
invaginação de uma placa de ectoderma superficial correspondente ao cristalino.
Com a indução da placa ectodérmica e sua invaginação para formar a vesícula
cristaliniana a vesícula óptica se invagina para formar a taça óptica. Após esta fase
a retina neural se desenvolverá relativamente independente dos outros tecidos.
Normalmente o acúmulo do vítreo primário dentro do olho é rápido o suficiente
para impedir dobras na retina se está se distendendo.
Quando a indução do ectoderma superficial inicia a formação do espaço
lenticuloretiniano e do cristalino; o epitélio pigmentar da retina se diferencia na
camada externa da taça óptica.
Inicialmente, as 2 paredes da taça óptica são monocelulares (1ª fase do
desenvolvimento retina, embrião com 4-10min). As células da face interna, através
de induções químicas se divide em várias camadas de células formando a camada
neuroepitelial ou retina sensorial.
Entre as camadas interna e externa da futura retina, persiste um espaço virtual
intraretiniano que se comunica com o III ventrículo pelo canal óptico.
O desenvolvimento da retina sensorial inicia-se ainda no estágio de vesícula
óptica, com a migração dos núcleos para a superfície da retina sensorial.
Profª Drª Márcia Reis Guimarães Pág 11
Curso de Embriologia Ocular / 2003 - Profª Drª Márcia Reis Guimarães
Hospital de Olhos de Minas Gerais – Belo Horizonte/MG
No segundo estágio do desenvolvimento retiniano (10 a 70mm), obesrvamos uma
zona de fibras primitivas entre as camadas interna e externa da retina sensorial ou
neuroblástica, trata-se da camada transitória da Chievitz que desaparecerá.
O desenvolvimento subseqüente da retina se caracterizará pela formação de novas
camadas através da divisão celular e migração posterior destas células. A retina se
desenvolve de dentro para fora: as células ganglionares formam-se precocemente e
os fotorreceptores são os últimos a atingir plena maturação.
No 3º estágio de desenvolvimento retiniano (70 a 200mm) ocorre a diferenciação
das células: da camada neuroblástica interna vêem as células ganglionares, células
de Muller, amácrimas e horizontais. A neuroblástica externa origina as células
bipolares, cones e bastonetes. As alterações na morfologia retiniana ocorrem pela
formação simultânea de conecções intercelulares múltiplas e de alta complexidade.
No 5º mês de gestação, a maior parte das conecções neurais já estão estabelecidas
(Mann, 1964).
As sinapses funcionais são feitas quase que exclusivamente nas 2 camadas
plexiformes e o pericarion das células nervosas está distribuído pelas três camadas
nucleares.
A maturação dos fotorreceptores começa pela formação dos segmentos externos
contendo o pigmento visual nas múltiplas pregas de sua membrana plasmática.
Com a definição dos segmentos completa, o olho se torna fotossensível à luz em
torno do sétimo mês de gestação. A última porção a se maturar é a fóvea. O
espessamento da camada ganglionar inicia-se no 4º mês de gestação. A camada
nuclear externa é mais espessa aqui do que no restante da retina e consiste quase
inteiramente em cones em desenvolvimento.
As células ganglionares migram radialmente para fora em círculo, deixando a
fóvea livre de seus núcleos. As conecções célula-célula pré-existentes e os cones
da fóvea alteram sua forma para acomodar o movimento das células ganglionares.
O desenvolvimento foveal se prolonga com rearranjos celulares a alterações na
forma dos cones até os 4 anos pós-nascimento (Hendrickson and Yondelis, 1984;
Curciot e Hendrickson, 1991).
A percepção visual da forma ocorre após o 1º ano de vida, o senso cromático se
inicia no 2º ano e só estará maturo na puberdade.
O desenvolvimento das duas camadas de neuroectoderma (retina sensorial e
epitélio pigmentar) é feito independente do cristalino mas a presença dele é
necessária para o crescimento normal da coróide e esclera e do epitélio pigmentar
da retina. Além disto a vesícula cristaliniana promove a diferenciação corneana e o
Profª Drª Márcia Reis Guimarães Pág 12
Curso de Embriologia Ocular / 2003 - Profª Drª Márcia Reis Guimarães
Hospital de Olhos de Minas Gerais – Belo Horizonte/MG
crescimento vítreo, existe ainda uma forte conexão organogenética entre a íris e o
cristalino.
A córnea tem dupla origem embriológica: seu epitélio é formado pelo ectoderma
superficial e os ceratócitos do estroma e o endotélio pela crista neural.
A crista neural tem papel vital na formação da câmera anterior atuando em três
ondas migratórias: na primeira, forma-se o endotélio corneano, em seguida os
ceratócitos ocupam o espaço entre o ectoderma superficial e o endotélio e, na
terceira onda migratória as células da crista neural formarão o estroma da íris no
espaço retrocorneano esvaziado de suas células mesenquimais através da apoptose.
Desenvolvimento da Córnea (Porção Central)
a) Ectodema Superficial contribui para a formação do Epitélio Corneano
b) Com 7 semanas, o Mesectoderma da periferia migra entre o Epitélio e o
Endotélio (precursor do futuro estroma corneano)
c e d) Os Ceratócitos (Mesectoderma) começam a produzir colágeno
e) Algumas células aladas aparecem no epitélio e uma membrana acelular,
começa a se diferenciar abaixo da lâmina basal (Bowman’s). A membrana de
Descemet já está bem evidenciada
f) Córnea com morfologia definitiva
Profª Drª Márcia Reis Guimarães Pág 13
Curso de Embriologia Ocular / 2003 - Profª Drª Márcia Reis Guimarães
Hospital de Olhos de Minas Gerais – Belo Horizonte/MG
A esclera evidencia-se anteriormente na 11ª semana pelo acúmulo de colágeno e
fibras elásticas e vai se estendendo posteriormente até que no 5º mês já existe uma
membrana ao redor do globo ocular com melanócitos começando a aparecer na
face interna. O canal de Schlemm (sinus venosus sclerae) começa com um plexo
venoso ao final do 3º mês e suas extensões mais tarde formarão as veias aquosas.
Inicialmente não há poros no endotélio que reveste o canal de Schlemm mas no 8º
mês poros intracelulares começam a se fenestrar e depois se abrem.
A conjuntiva se origina do ectoderma das pálpebras e vai recobrir a parte anterior
do globo. A diferenciação das células caliciformes (ou mucosas) e sua proliferação
só se fará após a abertura palpebral no fim do 5º mês. As dobras e o fórnice só
ficarão evidentes ao fim da gestação.
O vítreo se origina das células da crista neural. Estas células mesenquimais
invadem a taça óptica onde se fundem com processo protoplastmáticos e, na 6ª
semana, a vasculatura do vítreo primário já é evidente e os processos das células
de Muller estão em continuidade com as fibras do vítreo. Anteriormente as fibras
da artéria hialóidea se espalham formando a túnica vascular do cristalino. O vítreo
primário cessa seu desenvolvimento em torno da 9ª semana, sendo então
substituído por um vítreo secundário, mais fino, avascular e composto por células
finas em disposição paralela. Na 12ª semana as fibras da borda da taça óptica
extendem em filamentos até o equador do cristalino formando o vítreo terciário.
À medida em que o vítreo secundário aumenta seu volume, comprime o primário
para o centro do eixo antero-posterior (futuro canal de Cloquet). Ao final do 4º
mês, a túnica vascular cristaliniana anterior se atrofia e um filamento glial
substitui a art. hialóidea onde não há mais circulação sangüínea.
Profª Drª Márcia Reis Guimarães Pág 14
Curso de Embriologia Ocular / 2003 - Profª Drª Márcia Reis Guimarães
Hospital de Olhos de Minas Gerais – Belo Horizonte/MG
O vítreo posterior está aderido à membrana limitante interna da retina onde estão
as células de Muller. Os vasos sangüíneos do vítreo nutrem a retina imatura e ao se
obliterarem fazem-no posterior e externamente facilitando a expansão do vítreo
secundário.
A presença do cristalino é necessária para que o vítreo se acumule normalmente.
O desenvolvimento do cristalino começa na 4ª semana de gestação, com o
aparecimento de um espessamento no ectoderma superficial e prossegue com a
formação de uma vesícula, e assim por diante, conforme o esquema abaixo:
Profª Drª Márcia Reis Guimarães Pág 15
Curso de Embriologia Ocular / 2003 - Profª Drª Márcia Reis Guimarães
Hospital de Olhos de Minas Gerais – Belo Horizonte/MG
A Crista Neural contribui para a formação dos elementos de tecido conjuntivo do
olho e anexos e corresponde a uma massa celular situada nos dois lados da placa
neural e representa a delaminação do ectoderma superficial vizinho das pregas
neurais. A medida em que vai havendo a evaginação do olho primitivo, as células
da crista neural o envolvem e são chamadas de Mesectoderma ou
Ectomesênquina.
Os derivados do Mesectoderma (ou células da crista neural) são responsáveis pelas
seguintes linhagens de células oculares: ceratócitos, endotélio da córnea e
trabeculado, estroma da íris e coróide (células pigmentadas e não pigmentadas),
músculo ciliar liso, fibroblastos da esclera e meninges nervo óptico e maior parte
da esclera.
Os derivados da Crista Neural estão igualmente envolvidos na formação dos
tecidos orbitários: fibrogordurosos, nervos (inclusive o trigêmeo), cartilagem e
ossos, pericitos dos canais vasculares da órbita e células de Schwann perineurais.
O Mesoderma, anteriormente, responsabilizado pela maior parte dos tecidos
conjuntivos oculares participa na verdade de um modo bem menos significativo:
apenas os músculos estriados extra-oculares e o endotélio vascular dela são
derivados.
Profª Drª Márcia Reis Guimarães Pág 16
Curso de Embriologia Ocular / 2003 - Profª Drª Márcia Reis Guimarães
Hospital de Olhos de Minas Gerais – Belo Horizonte/MG
O Ectoderma Neural ou Neuroectoderma se deriva da placa neural e fornece
células para a formação do epitélio pigmentar da retina, músculos esfíncter da íris
e dilatador, neuroepitélio posterior da íris, epitélio pigmentar e não pigmentar do
corpo ciliar e fibras do nervo óptico e glia.
O epitélio pigmentar da retina atua sobre o mesênquima ao seu redor induzindo o
seu espessamento com posterior diferenciação em coróide e pequena porção da
esclera.
Desenvolvimento da Coróide
Neste esquema do desenvolvimento da coróide, observamos que no 2º mês, a rede
vascular primitiva mesenquimal ao redor do epitélio pigmentar da retina
(neuroectoderma) se conecta com pequenos vasos arteriais dos precursores das
artérias ciliares posteriores curtas (ACPC) que se originaram da artéria oftálmica
como 2 troncos juntamente com as ciliares posteriores longas (ACPL). Estas
últimas vão se estender até a parte anterior do corpo ciliar para formar as
tributárias dos plexos venosos infra e supra-orbitários. Ao 3º mês começam as
conexões das ACPC com a coriocapilar, 2 ou 3 ramos das ACPC participarão do
futuro círculo de Haller-Zinn. As ACPL’s formaram o círculo arterial maior na
região ciliar anterior.
Profª Drª Márcia Reis Guimarães Pág 17
Curso de Embriologia Ocular / 2003 - Profª Drª Márcia Reis Guimarães
Hospital de Olhos de Minas Gerais – Belo Horizonte/MG
Esquema do Desenvolvimento do Nervo Óptico e Vascularização Periocular
Profª Drª Márcia Reis Guimarães Pág 18
Curso de Embriologia Ocular / 2003 - Profª Drª Márcia Reis Guimarães
Hospital de Olhos de Minas Gerais – Belo Horizonte/MG
Esquema do Desenvolvimento do Corpo Ciliar e Íris e sua Interrelação com
as Posições do Canal de Schlemm’s e Ora Serrata
Origem do corpo ciliar: A vesícula óptica está circundada por mesênquima
indiferenciado e células da crista neural. As células da crista neural migrarão
para a frente do cristalino em várias ondas, sendo que, na segunda, forma-se a
íris primitiva e se divide a região em câmara anterior e posterior. Depois que o
estroma iriano se forma, duas camadas da borda da taça óptica movem
anteriormente posicionando-se em frente ao cristalino, formando inicialmente, a
camada epitelial do corpo ciliar e em seguida, o epitélio da íris. O músculo ciliar
se forma a partir das células da crista neural vizinhas do epitélio ciliar primitivo.
Profª Drª Márcia Reis Guimarães Pág 19
Curso de Embriologia Ocular / 2003 - Profª Drª Márcia Reis Guimarães
Hospital de Olhos de Minas Gerais – Belo Horizonte/MG
As pálpebras rudimentares se esboçam como dobras de mesênquima recobertas
por ectoderma nos dois lados. Subseqüentemente as dobras superior e inferior
crescem em direção ao centro e se fundem na 8ª - 9ª semana. A diferenciação do
ectoderma superficial leva à formação dos cílios e ceratinização da borda
palpebral, resultando na separação das pálpebras entre o 6º e o 7º mês de gestação.
No 2º trimestre de gestação aparecerão os cílios na borda de fusão das pálpebras
em 2 a 3 fileiras com folículo piloso correspondente. À frente, a pele se
queratiniza e posteriormente à ele forma-se o tarso e os músculos. Em
continuidade, a pele posteriormente não se ceratiniza formando a conjuntiva.
Entre o 3º e o 4º mês no ângulo interno, aparece uma prega de epitélio com
mesectoderma interiormente correspondendo à prega semilunar. Logo junto à ela,
internamente o epitélio da pálp. inf. se prolifera formando a carúncula.
As gls. de Meibomius aparecem no 3º trimestre e há secreção sebácea a partir do
8º mês.
Endereço para correspondência:
Hospital de Olhos de Minas Gerais – Clínica Dr. Ricardo Guimarães
Rua da Paisagem, 220 – Vila da Serra
Belo Horizonte/MG – CEP 34000-000
Telefone: (31)3289-2000 / Fax: (31)3289-2150
[email protected]
Profª Drª Márcia Reis Guimarães Pág 20