Universidade de São Paulo
Faculdade de Medicina de Ribeirão Preto
Programa de Pós-Graduação em Imunologia Básica e Aplicada
Autores: Daiani Cristina Cilião Alves e Taíse Natali Landgraf
Imunidade Adaptativa Humoral
A imunidade adaptativa humoral tem como principal molécula efetora o anticorpo. Em uma
infecção, o antígeno é transportado pela via linfática para o linfonodo, e linfócitos B vindos da
circulação ou residentes nos linfonodos reconhecem o antígeno, na forma solúvel ou não, através do
seu receptor de célula B (B Cell Receptor-BCR). Dessa forma, os linfócitos B são ativados
transformando-se em plasmócitos capazes de secretar anticorpos que entram na circulação até
encontrar o local de infecção e exercer suas funções efetoras.
1. Estrutura do Anticorpo
Conhecer a estrutura do anticorpo é muito importante para estudarmos sua formação e
funções. O anticorpo é composto por duas cadeias pesadas idênticas e duas cadeias leves idênticas.
As cadeias pesadas são unidas entre si e unidas às cadeias leves por ligações dissulfeto. As cadeias
leves e pesadas têm uma porção constante e uma porção variável. A porção variável apresenta o
sítio de ligação ao antígeno, contendo ainda regiões hipervariáveis denominadas Regiões
Determinantes de Complementariedade-1 (CDR-1), CDR-2 e CDR3, responsáveis pelas regiões de
maior diversidade e especificidade de reconhecimento do antígeno. A porção constante do anticorpo
contém o fragmento Fc (Fragment crystallizable) responsável pelas funções diferentes dos isotipos
IgM, IgD, IgG, IgA e IgE. O fragmento Fc liga-se a receptores Fc existentes em diversas células do
sistema imune e em plaquetas. O fragmento Fab (Fragment antigen binding) contém o domínio de
imunoglobulina variável (V) da cadeia leve e pesada (já descrito como responsável pela ligação ao
antígeno) e um domínio constante adjacente. Alguns isotipos de imunoglobulina apresentam ainda
subclasses como a IgA (IgA1 e IgA2) e IgG (IgG1 a IgG4) e podem ser monoméricas (IgG, IgD,
IgA e IgE), dimérica (IgA) ou pentamérica (IgM) (Figura 1).
Figura 1: A estrutura e os isotipos de anticorpos.
Sítio de ligação
ao antigeno
Cadeia pesada
N
Cadeia leve
N
N
N
vh
vL
CH1
CL
Região Fab
Dobradiça
Receptor de Fc /
sítio de ligação
do complemento
CH2
Região Fc
CH3
Ligação dissulfeto
Peça caudal
N
IgG
C
Domínio de
imunoglobulina
N
vh
N
C
N
N
N
N
IgA
CL
N
N
CL
CL
N
vL
CH3
C2
CH
3
CH2
N
CH3
CH4
CH
4
C
N
C
vh
Cadeia J
vL
C
N
N
C
vh
CL
CH3
CH2
CC
C
Cμ
1
C
C
C
C
C
C
C
C
C
C
C
CL
CH
3
C CH
4
C
C
CH4
Cμ
1
Cadeia J
CH3
CH
4
N
CH
3
N
C2
C2
Cμ
1
CL
CL
CH1
CL
CH4
C
C
N N
vL
vh
vh
vL
Cμ1
vL
N
N
C
C
N
N
N
vL
N
CH3
vh
C2
C2
N
CH2
CH
4
CH
3
CL
N
vL
CH1
C
C
C
C
N
N
C
C
C
IgE
N
C2
Cμ
1
CL
C
N
vL
vh
N
CH1
CH3
N
N
vh
Cμ1
vL
vh
CH2
C
Ig
M
N
N
vL
CH1
N
N
vh
N
N
N
2. Maturação da célula B
As células B são produzidas a partir de células tronco precursoras da medula óssea. Durante
a fase fetal, o fígado faz a função de maturação das células B até que a medula óssea seja formada,
sendo esses órgãos caracterizados como órgãos linfóides primários. Dessa forma, a célula B
apresenta as seguintes fases de maturação na medula óssea: célula tronco precursora, célula pró-B,
célula pré-B, e célula B imatura; e na periferia: célula B madura, plasmócito e célula de memória
(Figura 2).
Figura 2: Maturação da célula B e imunodeficiências relacionadas.
Periferia
Medula óssea
Pré-BCR
RAG 1/2
IgM-BCR
IgM
IgM-BCR
BTK
IgE
IgA
IgG
IgA
IgD
CD40L
CVID
Plasmócito
IgD-BCR
B
Célula tronco
Célula pró-B
Recombinação
somática da
cadeia pesada
Célula pré-B
Célula B imatura
Recombinação
somática da
cadeia leve
Seleção
negativa
Adição de
nucleotídios
Edição do
receptor
Ausência de antígenos
Célula B madura
Interação com célula T
Troca de isotipo
Célula B memória
Hipermutação somática
Antígenos próprios
Antígenos estranhos
Em amarelo, eventos da maturação da célula B e em verde, imunodeficiências (vide texto).
Abreviações: BCR- receptor de célula B; BTK-tirosina kinase de Brutton; Ig-imunoglobulina; CVID- imunodeficiência
variável comum.
A célula tronco precursora dá origem a todas as células hematopoéticas mielóides e
linfóides através de estímulos de fatores de crescimento. A célula B é da linhagem linfóide e
durante a maturação da célula pró-B e pré-B ocorre o processo de recombinação somática, em que
as enzimas RAG 1 e 2 (proteínas gênicas ativadoras da recombinase) promovem o rearranjo dos
segmentos gênicos variáveis (V), de junção (J), de diversidade (D) e constantes (C) para formação
da cadeia pesada do receptor de célula B (BCR) e dos segmentos V, J e C para formação da cadeia
leve, formando assim o BCR IgM, que é expresso na célula B imatura. A recombinação somática
representa o mecanismo responsável pela formação da diversidade de BCR e anticorpos, que passa
ainda por um processo de adição de nucleotídeos entre a junção dos segmentos V-D ou D-J,
podendo, dessa forma, apresentar aproximadamente 1011 tipos diferentes de anticorpos, que não
seria possível se esses segmentos gênicos não fossem rearranjados, pois temos somente 4x104 genes
no genoma humano (Figura 3).
Na fase de diferenciação de célula pró-B para pré-B e de pré-B para imatura, caso ocorram
defeitos gênicos da enzima RAG 1/2, o individuo poderá ser acometido por uma imunodeficiência,
denominada imunodeficiência combinada grave (IDCG) (T e B), que leva à redução dos números
de células B e T e consequente redução do número de anticorpos. Mutações da enzima tirosina
kinase de Brutton (BTK), responsável pela formação da célula B imatura, acarreta a
agamaglobulinemia ligada ao X, caracterizada por baixos níveis séricos de anticorpos e células B
(Figura 2).
Figura 3: recombinação somática das cadeias leves e pesadas para formação do BCR e anticorpos.
Cadeia leve (Lκ e Lλ)
Cadeia pesada (H)
V = 51
V1 V2
D = 27
J=6
D1-n
Vn
κ
J1-n
V = 40
C1-n (μ,δ,γ,ε,α)
J1-n
C1-n (μ,δ,γ,ε,α)
5´
Cromossomo 14
J=5
V1
V2
Vn
3´
5´
3´
Cromossomo 2
λ
V = 30
J=4
C1-n (μ,δ,γ,ε,α)
J1-n
Recombinação somática
(junção V-D-J)
V1
RAG1/2
V2
Vn
3´
5´
Cromossomo 22
Adição de nucleotídeos
(entre V-D e D-J)
TdT
Recombinação somática
(junção V-J),
RAG1/2
DH1 JH1
VH1
5
´
VHn
VH2
3´
Nucleotídeos
DH2
DH3 JH5
CμH1
CμH1
3´
5´
Nucleotídeos
Nucleotídeos
JH2
CμH1
5´
3´
Nucleotídeos
Jκ1
Vκ1 Cμκ1
Jκ2
Vκ2 Cμκ1
5´
3´
5’
Jλ1
Vλ1
3´
5
´
Cμλ1
3´
VH1
DH1
Nucleotídeos
JH1
Vκ1
VH1
DH1
JH1
Vκ1
Jκ1
Cμκ1
Jκ1
Cadeia leve
Cμκ1
CμH1
CμH1
Cadeia pesada
IgM
Durante a fase de célula B imatura ocorre importante processo de seleção negativa para que
essas células não reajam contra antígenos próprios. Assim, se a interação com antígenos próprios
ocorrer com baixa afinidade, dá-se sequência à maturação desse clone de linfócito B na periferia, se
ocorrer interação com alta afinidade o linfócito B pode: a) tornar-se anérgico (não funcional), b)
sofrer morte celular por apoptose, ou c) passar pelo processo de edição do receptor, ou seja, há
reativação da RAG e recombinação dos segmentos gênicos V-J da cadeia leve do outro
cromossomo. Dessa forma, a célula B imatura tem a chance de passar novamente pela seleção
negativa. Se ocorrerem falhas durante a seleção negativa e esses antígenos próprios forem
reconhecidos fortemente pelo BCR pode ocorrer doenças autoimunes (link Mecanismos de Escape
de Tolerância Imunológica).
A célula B madura, presente na periferia, co-expressa os BCR IgM e BCR IgD e é capaz de
reconhecer o antígeno estranho, iniciando uma cascata de sinalização intracelular que leva à
ativação de fatores de transcrição, responsáveis por uma série de eventos na célula B.
Diferentemente das células T, as células B maduras reconhecem o antígeno estranho na sua
forma solúvel e nativa, no sítio de infecção ou no linfonodo e, além de reconhecerem antígenos
protéicos, reconhecem também antígenos não protéicos, como polissacarídeos, ácidos nucléicos e
glicolipídios.
A resposta de célula B a antígenos protéicos é dependente da interação com célula T que
ocorre na interface entre a zona de célula B e zona de célula T do linfonodo. A ligação cruzada do
antígeno ao BCR leva à endocitose e processamento do antígeno e à expressão de B7.1 (CD80) e
B7.2 (CD86) na superfície da célula B. Ocorre o reconhecimento do peptídeo ligado ao MHC de
classe II da célula B com o TCR da célula T, e do B7 da célula B com o CD28 da célula T, com
consequente estimulação da expressão de CD40L (Ligante de CD40) pela célula T e secreção de
citocinas. Ocorre dessa forma, a ativação da célula B pelas citocinas e interação do CD40 da célula
B com o CD40L da célula T, responsável por promover proliferação e diferenciação das células B
(Figura 4). Mutações no gene do CD40L na célula T leva à Síndrome de Hiper-IgM que é
caracterizada por níveis séricos normais de IgD e elevados de IgM, com ausência dos outros
isotipos de imunoglobulinas e número normal de células B e T (Figura 2).
Figura 4: Interação da célula B com célula T
Folículo
primário
B7
CD40
B7
BT
Linfonodo
CD28
CD40L
CD28
TCR
MHC II
B7
CD40
TCR
MHC II
MHC II
Citocinas
As citocinas do microambiente são responsáveis pela troca de isotipo, que ocorre pelo
processo de edição alternativa do segmento constante (C) Cμ, (responsável pela produção de IgM),
por segmentos Cδ (IgD), Cγ (IgG), Cε (IgE) ou Cα (IgA). A célula B se diferencia em plasmócitos
capaz de secretar anticorpos que através da circulação sanguínea chegam ao foco de infecção para
exercer suas funções efetoras. Nessa fase, pode ocorrer doença de base genética desconhecida
denominada imunodeficiência variável comum (CVID), caracterizada pela ausência da formação de
plasmócitos. Podem ocorrer falhas na diferenciação de todos os isotipos de imunoglobulinas
levando a imunodeficiências, como a deficiência de IgA e a de IgG (Figura 2).
Os diferentes isotipos de imunoglobulina são distribuídos pelo organismo de acordo com sua
função, IgG e IgM predominam no plasma, IgA dimérica predomina nas secreções mucosa e
epitélios, incluindo o leite materno. IgE é encontrada principalmente associada com os mastócitos
logo abaixo de superfícies epiteliais (especialmente do trato respiratório, trato gastrointestinal e
pele). O feto recebe IgG da mãe por transporte transplacentário e o cérebro é normalmente
desprovido de imunoglobulina.
Outro evento que ocorre na célula B madura presente no linfonodo é a hipermutação
somática. A célula B entra novamente em contato com o antígeno peptídico, seja na superfície de
células dendríticas foliculares ou na forma solúvel, e sofre mutações nas regiões hipervariáveis da
imunoglobulina (CDR-1, CDR-2 e CDR-3). Esse processo é responsável por gerar maior afinidade
pelo antígeno e se as mutações gerarem menor afinidade, essas células B entram em apoptose.
Durante a maturação da célula B ocorre a formação de células B de memória que
permanecem após termino da resposta imune e que no caso de uma reinfecção têm características de
resposta mais rápida, secreção de anticorpos de maior afinidade e de diferentes isotipos.
A resposta da célula B a antígenos não protéicos é independente da interação com células T,
podendo ocorrer ativação da célula B por duas formas distintas: a) ligação cruzada do antígeno em
mais de um BCR ou b) o antígeno opsonizado pelo complemento é reconhecido simultaneamente
pelo BCR e pelo receptor de complemento CR2, ambos expressos na membrana da célula B. A
ativação da célula B independente de célula T leva à restrição em eventos de troca de isotipo
(somente IgG), maturação por afinidade e formação de célula de memória.
3. Mecanismos de ação.
Como os anticorpos protegem nosso corpo? Os anticorpos desempenham diversas funções
efetoras: neutralização, opsonização, ativação do complemento e citotoxicidade celular dependente
de anticorpo.
Figura 5: Mecanismos de ação dos anticorpos
Neutralização de
Antigenos e toxinas
Opsonização e
fagocitose de microbios
N
K
alvo
Citotoxicidade celular
dependente de
anticorpo
Ativação do
complemento
Neutralização: neutraliza toxinas bacterianas, agentes virais, entre outros antígenos
solúveis, o que impede a ligação desses antígenos a receptores específicos na superfície
celular e, consequentemente, o processo infeccioso. Os principais isotipos que
desempenham essa função são IgG1, IgG2, IgG3, IgG4 e IgA;
Opsonização: os anticorpos ligam-se aos antígenos e suas porções Fc são reconhecidas
pelos receptores Fc localizados na superfície de fagócitos. Esse complexo antígeno–
anticorpo é endocitado e destruído nas vesículas endossômicas. IgG1 e IgG3 são as
principais subclasses de IgG que opsonizam antígenos e facilitam a fagocitose;
Ativação do complemento: alguns isotipos de anticorpos contêm um sítio de ligação ao
complemento presente nos domínios de imunoglobulina CH2 (IgG) ou CH3 (IgM) que
interage com as regiões globulares da molécula C1q do complemento, ativando-a (link
Sistema do Complemento);
Citotoxicidade celular dependente de anticorpo (ADCC, do inglês Antibody-Dependent
Cell-Mediated Cytotoxicity): Esse mecanismo consiste na interação da porção Fc do
anticorpo ligado à célula alvo (pela porção Fab) com o receptor Fc da célula efetora
(eosinófilos, mastócitos e células NK) e liberação de grânulos que causam a lise das células
alvo. Os isotipos que desempenham essa função são principalmente as subclasses de IgG1
e IgG3, que se ligam a receptores Fc das células Natural Killer, e IgE, que se ligam a
receptores dos mastócitos e IgG e IgA em receptores de eosinófilos.
4. Regulação
Após eliminação do antígeno, as respostas imunes necessitam ser finalizadas para que não
ocorram danos ao organismo causados por resposta imune exacerbada, de forma que seja
recuperada a homeostase. A própria ausência do estimulo antigênico leva ao processo de apoptose.
No caso da resposta imune humoral, ocorre mecanismo de regulação negativa de anticorpos, que
consiste na ligação de complexos antígeno–anticorpos a receptores Fc localizados na superfície dos
linfócitos B, gerando sinais negativos, impedindo a ativação desses linfócitos.
5. Implicações clínicas
Acerca da imunidade adaptativa humoral, são relatadas diversas imunodeficiências,
diagnosticadas especialmente entre 5 a 7 meses de idade, algumas em crianças maiores ou na idade
adulta. As principais manifestações dessas imunodeficiências incluem infecções recorrentes,
principalmente, por patógenos como Streptococcus, Staphylococcus, Haemophilus, Campylobacter,
Enterovirus e Giardia, e ainda, outras alterações como autoimunidade, linfomas, timoma,
poliomielite pós-vacinação.
As imunodeficiências mais importantes relacionadas ao sistema imune humoral estão
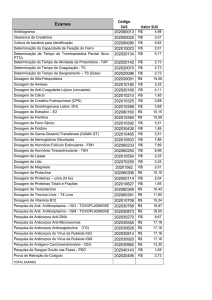
representadas na tabela abaixo.
Tabela 1: Imunodeficiências da resposta imune humoral
Deficiências
Imunodeficiência Combinada Grave (IDCG)
(T-B-)
Agamaglobulinemia ligada ao X
Síndrome de Hiper-IgM
Deficiência de IgA
Deficiência de IgG
Imunodeficiência variável comum (CVID)
Ig sérica
Linfócito B
Linfócito T
Defeito
↓
↓
↓
Defeitos da RAG 1/2
↓
↓
-
Mutações da BTK
Normal
Normal
IgM e IgD ↑ ou
normal
↓ ou ausência
↓ ou ausência
de subclases
Variável
Mutações no gene do
CD40L
Normal
Diferenciação isotipica
Normal
Diferenciação isotipica
Normal ou ↓
-
Desconhecido
Referências:
Abbas, A. K.; Lichtman, A. H. Imunologia celular e molecular, 6a ed. Rio de Janeiro. Elsevier,
2008.
Voltarelli, J.C.; Donadi, E.A.; de Carvalho, I.F.; Arruda, L.K.; Louzada Jr., P.; Sarti, W.
Imunologia Clínica na Prática Médica, 1ª ed. São Paulo. Editora Atheneu, 2009.
Charlotte Cunningham-Rundles and Prashant P. Ponda. Molecular defects in T and B cell
primary immunodeficiency diseases. Nature reviews immunology