Hereditariedade ligada aos CROMOSSOMAS SEXUAIS
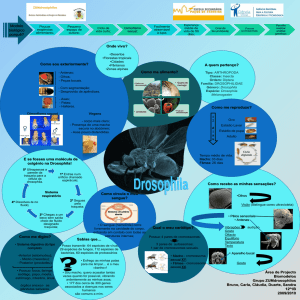
Thomas H. Morgan – embriologista, desenvolveu trabalho com as moscas Drosophila
Melanogaster (mosca da fruta).
Razões para o uso de Drosophila Melanogaster:
Apresenta um curto período de desenvolvimento e apresenta cromossomas muito grandes,
que facilitam o seu estudo e observação.
Aspecto da Drosophila
Melanogaster
Forma selvagem
Corpo cinzento
Olhos vermelhos
Asas longas
Forma mutante
Corpo negro
Olhos brancos
Asas vestigiais
Representamos a constituição genética das formas alternativas pela letra inicial da palavra
inglesa que expressa a característica que elas manifestam.
Exemplo: alelo para olhos brancos -> w (white). Quando é da forma selvagem é w+.
Nas experiências de Mendel não era relevante que um determinado fenótipo pertencesse a
uma fêmea ou a um macho – o cruzamento recíproco não interferia nos resultados.
Os resultados obtidos por Morgan eram diferentes – ao cruzar uma fêmea de olhos
vermelhos com um macho de olhos brancos não obtinha os mesmos resultados de quando
cruzava uma fêmea de olhos brancos com um macho de olhos vermelhos.
No 1º cruzamento todos os indivíduos apresentavam olhos vermelhos, sendo 50% machos
e 50% fêmeas – de acordo com o previsto por Mendel.
No 2º cruzamento as fêmeas têm todas olhos vermelhos e os machos olhos brancos – não
se verifica a uniformidade fenotípica dos indivíduos da primeira geração.
COMO EXPLICAR?
Na Drosophila, como na maioria das espécies animais, existe um par de cromossomas
chamado cromossomas sexuais.
Indivíduos que apresentam dois
HOMOGAMÉTICOS
cromossomas sexuais idênticos
Indivíduos que apresentam dois
HETEROGAMÉTICOS
cromossomas sexuais diferentes
entre si
Como a espécie humana, as fêmeas de Drosophila, para além dos autossomas, apresentam
dois cromossomas sexuais X, enquanto que os machos apresentam, para além dos
autossomas, um cromossoma sexual X e outro, mais curto e praticamente desprovido de
genes, Y.
O sexo heterogamético é portanto o masculino.
Se considerarmos que o alelo da cor branca dos olhos de Drosophila se localiza no
cromossoma X, podemos justificar o resultado dos dois cruzamentos.
As características hereditárias que dependem de genes localizados no cromossoma X são
características ligadas ao sexo.
Nestes casos, os resultados obtidos no cruzamento directo e no seu recíproco são
diferentes. Estes resultados devem-se ao facto de o cromossoma Y do macho não possuir os
alelos correspondentes do cromossoma X, dado que os dois cromossomas não são totalmente
homólogos.
A maior parte dos genes localizados no cromossoma X não têm alelo correspondente no
cromossoma Y, pelo que existe um único alelo para esse gene e esse alelo exprime-se sempre
no fenótipo dos machos, que são hemizigóticos.
Os genes presentes no cromossoma Y são transmitidos DE PAI PARA FILHO. Os genes
presentes no cromossoma X são transmitidos de PAI PARA FILHA e de MÃE PARA FILHO OU
FILHA.
Transmissão de um alelo DOMINANTE
ligado ao cromossoma X
O carácter exprime-se sempre nos
homens, de uma forma mais severa do
que nas mulheres
O carácter exprime-se nas mulheres
homozigóticas dominantes e
heterozigóticas
Um homem afectado tem uma mãe
afectada
Uma mulher afectada tem uma mãe
afectada ou um pai afectado
Síndrome de Rett, hipertricose
Transmissão de um alelo RECESSIVO
ligado ao cromossoma X
O carácter exprime-se sempre nos
homens
O carácter exprime-se apenas nas
mulheres homozigóticas recessivas ( e
nos homens?)
Um homem afectado tem uma mãe
afectada ou portadora
Uma mulher afectada tem
obrigatoriamente um pai afectado e
uma mãe afectada ou portadora
Daltonismo, hemofilia, diabetes
insípidos
Os trabalhos de Morgan são excepções às Leis de Mendel – contudo, apoiaram a teoria
cromossómica da hereditariedade.
Ligação
factorial
Descrição
Exemplos de situações em que
se verifica
Os genes localizados no mesmo cromossoma, não
sofrem geralmente, segregação independente na
meiose – ficam juntos nos mesmos gâmetas e, por
isso, os fenótipos da descendência não seguem as
proporções previstas pelas Leis de Mendel.
Os fenómenos de crossing-over podem separar genes
ligados, o que faz com que se comportem como se
estivessem localizados em cromossomas diferentes e
apareçam recombinados na descendência. Quanto
mais distantes estiverem dois genes no mesmo
cromossoma, maior a probabilidade de serem
separados por crossing-over.
O gene do grupo sanguíneo Rh e
o gene da eliptocitose (uma
forma de anemia) estão
localizados
no
mesmo
cromossoma.
Estes
dois
cromossomas são herdados em
bloco por 96% dos indivíduos e
4%
dos
indivíduos
são
recombinantes.
Explicação do manual
Cada cromossoma tem de ter muitos genes.
Os genes que se dispõem linearmente ao longo do mesmo cromossoma dizem-se em
linkage e constituem um grupo de ligação factorial – são transmitidos em bloco.
Cruzam-se indivíduos de duas linhas puras com características antagónicas.
Fenotipicamente
Corpo negro e
x Corpo cinzento e asas
asas vestigiais
longas
+ + + +
Genotipicamente
bbvgvg
x
b b vg vg
b – símbolo do alelo responsável pela cor negra (black)
vg – símbolo do alelo responsável pelas asas vestigiais ( vestigials)
b+ - símbolo do alelo responsável pela cor selvagem (cinzenta – DOMINANTE)
vg+ - símbolo do alelo responsável pela forma selvagem das asas (longas – DOMINANTE)
O cruzamento destas linhas puras resulta numa geração F1 – cujos resultados
correspondem a uma situação normal de diibridismo, em que os descendentes são
HETEROZIGÓTICOS e manifestam as características do ALELO DOMINANTE.
Fenótipo – Corpo cinzento e asas longas
Genótipo – heterozigótico b+bvg+vg
para que se mantivessem as previsões mendelianas, deviam agora surgir quatro classes
fenotípicas, que seriam:
Fenótipo dos
descendentes
Corpo cinzento e
asas longas
Corpo negro e asas
longas
Corpo cinzento e
asas vestigiais
Corpo negro e asas
vestigiais
Resultados
esperados em
diibridismo
9/16
Resultados
observados
3/16
-
3/16
-
1/16
1/4
3/4
Estes resultados são explicados pelo
facto de os alelos do corpo negro e asas
vestigiais estão situados no mesmo
cromossoma – são transmitidos em
conjunto
(não
há
segregação
independente prevista por Mendel) –
correspondem a um cruzamento de
Linhas puras em monibridismo.
Nem sempre os genes em linkage se comportam como uma unidade inseparável – pode
acontecer, que como resultado de crossing-over durante a meiose, os genes se separarem,
como se estivessem em cromossomas separados.
Assim, obtém-se uma descendência qualitativamente igual à prevista numa segregação
independente (em que os alelos são segregados de forma aleatória). Contudo, estes genes só
se transmitem deste modo quando há a sua separação em crossing-over, e isto ocorre muito
menos frequentemente que a transmissão em bloco.
Embora possam surgir as 4 classes fenotípicas esperadas, as suas proporções são
completamente aleatórias.
Organização e regulação do material genético
Genoma – totalidade do material genético de um indivíduo (contém todos os genes).
Gene – sequência de nucleótidos de uma molécula de DNA que origina uma molécula de
RNA funcional.
Organização do Material Genético
Genoma dos procariontes
Molécula circular de DNA associada a proteínas
não histónicas, que forma o seu único
cromossoma e se concentra na região do
nucleóide.
Algumas bactérias também possuem moléculas
circulares de DNA chamadas plasmídeos.
Genoma dos eucariontes
Várias moléculas lineares de DNA nuclear
associadas a uma grande quantidade de
proteínas, especialmente histonas, formando a
cromatina. Cada molécula de DNA associada a
proteínas constitui um cromossoma.
Também possui material genético extranuclear.
As mitocôndrias e cloroplastos contêm DNA que
codifica produtos essenciais à sua função
biológica e que é muito semelhante ao DNA
bacteriano.
Um cariótipo organiza os cromossomas metafásicos aos pares com base no seu tamanho e
noutras marcas físicas, como a posição do centrómero.
O cariótipo humano tem 46 cromossomas, organizados em 23 pares.
44 são autossomas e são idênticos nos dois sexos (possuem os mesmos genes, na mesma
sequencia) e 2 são heterossomas (ou cromossomas sexuais).
A análise do cariótipo é útil para confirmar diagnósticos clínicos de certas doenças de
transmissão hereditária – a comparação de cariótipos de diferentes espécies permite
encontrar relações evolutivas.
Regulação do material genético
Temos muitos genes no nosso corpo, mas só apenas alguns se manifestam. Tem de haver
portanto uma regulação dos genes. Este processo foi estudado pelos franceses François Jacob
e Jacques Monod.
Os organismos unicelulares reagem às variações do meio ambiente, variando a expressão
dos genes e ajustando o seu metabolismo – desenvolveram mecanismos de resposta RÁPIDOS
face às alterações das condições do meio, das quais dependem muito.
Nos eucariontes multicelulares, o controlo da expressão dos genes torna possível que as
células com o mesmo DNA possam divergir (em forma e função), tornando-se especializadas.
A transcrição do DNA para mRNA é um exemplo da regulação da expressão dos genes.
Modelo do Operão
(principal mecanismo de controlo da expressão dos genes em bactérias)
Unidade funcional constituída pelos elementos descritos abaixo.
Operão
Conjunto de genes que codificam proteínas com funções relacionadas.
Genes
estruturais Ex.: enzimas de uma determinada via metabólica
Sequência específica de nucleótidos do DNA à qual se liga a RNA
Gene
promotor polimerase e onde tem início a transcrição
Sequência de DNA que controla o acesso da RNA polimerase ao
Gene
operador promotor e que permite activar ou desactivar a transcrição de todos os
Gene
regulador
Repressor
genes estruturais
Encontra-se a uma determinada distância do operão, tem o seu
próprio promotor e codifica o repressor
É uma proteína alostérica com duas formas, uma activa e uma inactiva.
É específico, reconhece e liga-se apenas ao operador de um
determinado operão.
Explicação do funcionamento do operão lac.:
Funcionamento de um operação do tipo indutivo
NA AUSÊNCIA DE LACTOSE
O gene regulador determina a síntese de
um repressor;
O repressor bloqueia o promotor, ao
ligar-se ao operador;
A enzima RNA polimerase não se liga ao
promotor;
Os genes estruturais não são transcritos;
Não ocorre a síntese das três
enzimas.
NA PRESENÇA DE LACTOSE
A lactose liga-se ao repressor,
inactivando-o;
O operador fica desbloqueado;
A enzima RNA polimerase liga-se ao
promotor;
Os genes estruturais são transcritos;
Dá-se a síntese de enzimas.
Explicação do funcionamento do operão trp.:
Funcionamento de um operação do tipo repressivo
NA AUSÊNCIA DE TRIPTOFANO
O gene regulador produz um repressor
que está inactivo;
O operador está livre;
A RNA polimerase pode ligar-se ao
promotor;
Dá-se a transcrição;
NA PRESENÇA DE TRIPTOFANO
O triptofano liga-se ao repressor,
activando-o;
O repressor liga-se ao operador;
A RNA polimerase não pode ligar-se
ao promotor;
Não se dá a transcrição;
Ocorre a síntese de enzimas.
Não se sintetizam as
enzimas.
Muitos genes de um genoma se destinam a regular o funcionamento de outros genes.
Os genes que se expressam numa determinada situação dependem das interacções que o
ambiente estabelece com o DNA.
Transmissão Genética de Genes Mitocôndriais
O material genético contido nas mitocôndrias é transmitido pela mãe para os filhos e filhas.
A razão para este facto é simples: o citoplasma (e todos os seus constituintes) que vai dar
origem ao zigoto é proveniente do oócito (tem, portanto, todos os organelos celulares da mãe
– incluindo a mitocôndria!); o espermatozóide, apenas contribui com o núcleo para a formação
do zigoto, pelo que não são transmitidas as mitocôndrias do progenitor masculino.
Diferenças e semelhanças entre o DNA mitocondrial e o DNA nuclear.
DNA mitocondrial
Não possui exões
Não ocorre crossing-over
Possui várias cópias de DNA em cada
mitocôndria, permitindo que na mesma
célula existam diferentes alelos para o
mesmo gene
Taxa de mutação muito elevada
Não possui enzimas que reparam o DNA
DNA nuclear
Possui exões
Ocorre crossing-over
Só possui uma cadeia (com dupla hélice) de
DNA no núcleo da célula
Taxa de mutação pouco elevada
Possui enzimas que reparam o DNA
Mutações
Mutação – alteração permanente no material genético que afecta a expressão de um ou
mais genes.
Apesar de se darem centenas de
alterações do DNA por dia, as células
possuem enzimas capazes de corrigir
ou eliminar porções mutadas do DNA,
diminuindo a hipótese de esta ser uma
mutação
que
se
manifeste
fenotipicamente. Podem ser génicas
ou cromossómicas.
m. génicas – alteram a estrutura do
DNA;
m. cromossómicas – alteram a
estrutura/número de cromossomas;
m. silenciosas – não alteram a
proteína ou a sua acção;
m. letais – provocam a morte ou
doenças e anomalias;
m. benéficas – levam à evolução
das espécies;
m. prejudiciais – provocam a morte
do indivíduo.
Agentes mutagénicos são factores do meio que provocam mutações em genes e/ou
cromossomas.
As mutações podem ocorrer em células somáticas ou germinativas.
Mutação somática
Ocorre durante a replicação do DNA que precede uma divisão mitótica.
Pode originar um conjunto ou um clone de células mutantes identicas entre si, que se
distinguem das restantes células do indivíduo.
A descendência do indivíduo não é afectada.
Este tipo de mutação está na origem de certos cancros.
Mutação nas células germinativas
Ocorre durante a replicação do DNA que precede a meiose.
A mutação afecta os gâmetas e todas as células que dela descendem após a fecundação.
Mutações génicas
Ocorrem quando se dá uma alteração pontual ao nível dos nucleótidos de um gene,
constituindo-se uma nova versão do gene.
Alteram a sequencia de nucleótidos do DNA, por substituição, adição (inserção) ou
remoção (delecção) de bases.
Estas mutações podem conduzir à modificação da molécula de mRNA que é transcrita a
partir do DNA e à alterção da proteína produzida. O efeito desta alteração é imprevisível,
dependendo de qual o tipo de mutação e qual a proteína que passa a ser codificada. Pode ter
efeitos benéficos e levar à evolução da espécie, ou pode ser prejudicial e causar a morte do
indivíduo ou um grande numero de doenças e anomalias. Pode também ter um efeito neutro,
não causando quaisquer modificações.
Mutações génicas
Substituição
Inserção
Delecção
Ocorre a troca de um ou mais pares de bases.
Acontece quando uma ou mais bases
são adicionadas ao DNA, modificando a
ordem de leitura da molécula durante a
replicação ou transcrição.
Acontece quando uma ou mais bases
são retiradas do DNA, modificando a
ordem de leitura, durante a replicação
ou transcrição.
A adição/remoção de um numero
que não seja múltiplo de três
altera completamente a
mensagem do gene.
Mutações cromossómicas
Traduzem-se numa alteração da estrutura ou do número de cromossomas. Podem afectar
uma determinada região de um cromossoma, um cromossoma inteiro ou todo o complemento
cromossómico de um indivíduo.
Mutações cromossómicas numéricas
Aneuploidia
Poliploidia
Tipo de
mutação
Definição/causas
Consequências/exemplos
Existe pelo menos um conjunto completo de É comum nas plantas. As
plantas poloplóides podem
cromossomas a mais.
Entre as causas possiveis:
-fecundação
de
um
oócito
por
2
espermatozóides;
-fecundação de um gâmeta diplóide (triploidia);
-falta de divisão do zigoto após a replicação dos
cromossomas
autopolinizar-se ou cruzar-se
com plantas semelhantes.
Nos
humanos
embiões
poliplóides não se desenvolvem
e
são
abortados
espontaneamente.
Algumas
células somáticas podem ser
poliplóides.
Existem cromossomas a mais ou a menos
Anuploidias mais comuns em
em relação ao numero normal.
seres
humanos
são
as
Geralmente, envolve apenas um par de
cromossomas e pode ser autossómica ou nos
cromossomas sexuais.
Podem distinguir-se:
Polissomia – um ou mais cromossomas extra;
Monossomia – um cromossoma em falta;
As aneuploidias são causadas pela não-disjunção
dos cromossomas homólogos ou dos cromatídeos
na anafase da meiose I ou II.
Um gâmeta recebe 2 cromossomas do mesmo
par e outro não recebe nenhum.
trissomias dos cromossomas 21,
13, 18 e a monossomia do X.
Aneuploidias
de
outros
cromossomas não permitem o
desenvlvimento
até
ao
nascimento e resultam num
aborto espontâneo.
As
aneuploidias
nos
cromossomas
sexuais
são
melhor toleradas que as dos
autossomas. Síndrome.
Sindromes estudadas:
Trissomia 21 – (47,XX) ou (47,XY) – SÍNDROME DE DOWN
Cromossoma extra no ‘lote’ 21.
Monossomia do X – (45,X0) – SÍNDROME DE TURNER
Afecta apenas mulheres, que carecem de um dos cromossomas sexuais.
(47,XXY) – SÍNDROME DE KLINEFELTER
Mutações cromossómicas estruturais
Tipo de
mutação
Definição/Causas
Delecção
Representa
uma
perda
no
material
cromossómico.
As delecções visíveis de cromossomas humanos
estão
sempre
associadas
a
grandes
incapacidades.
Duplicação
Translocação
Inversão
Consequências/Exemplos
Caracteriza-se pela repetição de uma porção de
cromossoma.
As duplicações são alterações cromossómicas
muito importantes sob o ponto de vistaa
evolutivo, porque fornecem informação genética
complementar, potencialmente capaz de assumir
novas funções.
Ocorre uma inversao quando um segmento
cromossómico experimenta uma rotação de 180º
em relação à posição normal, sem alterar a sua
localização no cromossoma.
A transferencia de uma porção de um
cromossoma, ou mesmo de um cromossoma
inteiro para outro não homólogo designa-se por
translocação simples.
As translocações mais comuns são as
translocações recíprocas, havendo troca de
segmentos entre cromossomas não homólogos.
As translocações podem alterar drasticamente o
tamanho dos cromossomas, assim como a
posição do centrómero.
Poliploidia
Os inivíduos poliplóides são indivíduos em que o número de conjuntos completos de
cromossomas é multiplo do numero haploide primitivo existente nos gâmetas. Apresentam
cariótipos triploides (3n), tetraploides (4n) ou mesmo numeros mais elevados de
cromossomas.
A poliploidia surge:
- acidentalmente;
- a partir da não-disjunção dos cromossomas durante a meiose ou mitose. Também pode
acontecer que não há citocinese na repartição dos cromossomas pelas células filhas.
- cruzamento entre indivíduos de espécies diferentes (o que é muito comum entre as
plantas) – os indivíduos resultantes deste processo são naturalmente estéreis, uma vez que
não possuem cromossomas homologos, não podendo estes emparelhar durante a meiose.
Como é que estes indivíduos se reproduzem então?
Através de reprodução assexuada – no caso dos individuos que resultam do cruzamento
entre espécies diferentes, estes acabam por tornar-se ferteis apos algumas gerações, devido a
uma ocorrencia de uma duplicação cromossómica resultante de uma não-disjunção dos
cromossomas na divisão celular.
A poliploidia é muitas vezes provocada em laboratório para que se obtenham plantas mais
resistentes, com grandes frutos, sem caroço ou sementesm grãos de trigo maiores, etc.
As Mutações, a tecnologia e a vida
Um agente mutagénico é qualquer agente responsável por uma mutação.
O processo que conduz ao aparecimento de mutações pelo agente mutagénico é a
mutagénese.
As nossas células tem a capacidade de reparar alguns danos causados ao DNA. Há portanto,
um equilíbrio entre a proliferação celular, em que as células se renovam e multiplicam e entre
a morte das células.
Apesar disso, este equilíbrio por vezes perde-se – umas das consequências é o
aparecimento de um cancro.
Um cancro (neoplasia maligna/tumor maligno) é um conjunto muito heterogéneo e
multifactorial de doenças que têm em comum o facto de apresentarem sempre o crescimento
de um tecido neoformado.
Outra definição
O cancro é uma doença genética que resulta da perda de controlo do ciclo celular. A divisão
da célula com mais frequência dá origem a uma população de células em proliferação
descontrolada e forma um tumor.
As células cancerosas:
-são pouco especializadas e com forma arredondada;
-dividem-se continuamente;
-invadem os tecidos adjacentes;
-podem instalar-se noutros lugares do organismo.
O aparecimento de cancros está normalmente associado a alterações dos mecanismos que
regulam a divisão celular.
Necrose – as células morrem devido à acção de substâncias tóxicas ou à falta de nutrientes
essenciais. Apesar de manterem o núcleo intacto, aumentam de volume, rompe-se a
membrana plasmática e verte-se o conteúdo da célula no meio extracelular, causando uma
pequena inflamação.
Apoptose – ocorre um conjunto de fenómenos
programados geneticamente e que levam à morte da
célula – processo mais comum. Quando as células
apresentam anomalias – sobretudo genéticas – ou já
não são necessárias ao organismo, desencadeia-se
um “suicídio” por parte das células.
1. A cromatina começa a condensar;
2. A célula isola-se das células vizinhas,
compactando o citoplasma e a cromatina;
3. Uma enzima (endonuclease/enzima de
restrição) fragmenta o DNA em pequenas unidades;
4. A célula fragmenta-se sem que ocorra
ruptura nem resposta inflamatória.
Quando este equilíbrio, entre a divisão celular e a apoptose é quebrado, pode surgir um
cancro.
As neoplasias têm origem genética, pois resultam de
alterações no DNA.
No caso de as alterações se
darem
a
nível
dos
proto-oncogenes:
Estes são genes que
estimulam a divisão celular,
mas que estão inactivos em
células que não se dividem.
Devido à acção de agentes
mutagénicos podem tornar-se
activos, e passam a estimular
permanentemente a divisão
celular,
passando
a
oncogenes.
No caso de as alterações se dares ao nível dos genes supressores tumorais:
Estes genes têm a função de regulam a proliferação celular, contrabalançando a acção dos
proto-oncogenes, inibindo-os. Estes genes estão normalmente activos (bloqueiam a divisão
celular), mas devido à influencia de agentes mutagénicos podem desactivá-los, fazendo com
que as células se continuem a dividir.
As infecções por vírus contribuem para o aparecimento de cancro pela integração do
material genético do vírus no DNA das células infectadas. O DNA viral pode ser inserido num
local onde destrua a actividade de um gene supressor tumoral ou converta um protooncogene num oncogene.
Todos os cancros são genéticos, mas quase nenhuns são hereditários. Nestes casos, a
alteração genética está presente em todas as células do indivíduo, manifestando-se muito
cedo.
A maioria dos cancros é esporádica (95%) e surgem como resultado de mutações nas
células somáticas. Estas alterações são promovidas pela interacção entre o genoma do
indivíduo e o ambiente.
As componentes genética e ambiental estão sempre presentes, apesar de nem sempre
assumirem igual importância.
Ex: melanoma – radiações solares + alteração de um gene supressor tumoral (MTS)
localizado no cromossoma 9.
Todos os dias surgem neoplasias no nosso corpo, que são eliminadas por apoptose. Quando
isto não acontece, inicia-se um cancro, que corresponde ao momento em que estas células se
proliferam e invadem tecidos vizinhos.
Pode seguir-se um processo de metastização, em que as células cancerosas se podem
movimentar através da corrente sanguínea ou linfática e continuar a desenvolver-se noutras
partes do corpo.
Fundamentos da Engenharia Genética
A engenharia genética permite manipular directamente os genes de determinados
organismos com objectivos práticos.
Após a descoberta de que também o DNA podia ser manipulado, a primeira “ferramenta”
da engenharia genética foram as enzimas de restrição (ou endonucleases).
Estas enzimas cortam a hélice dupla do DNA em zonas específicas, sempre que as
encontram.
Funcionamento das enzimas de restrição
Os vírus invadem as bactérias e afectam o seu DNA.
Algumas bactérias têm um mecanismo de defesa contra os vírus, que consiste na produção
de enzimas de restrição.
Ou seja:
1. As enzimas cindem a cadeia de DNA do vírus quando encontram uma determinada
sequência de pares de bases.
2. Estas enzimas actuam em pontos específicos (ZONAS DE RESTRIÇÃO), catalisando o
desdobramento do DNA em fragmentos menores.
3. Estes fragmentos possuem nas extremidades a sequência de nucleótidos reconhecida
pela enzima de restrição – são constituídos por cadeia simples ligada a cadeia dupla e
chamam-se extremidades coesivas.
As extremidades coesivas podem ligar-se por complementaridade a outro DNA. Intervêm as
ligases do DNA, que catalisam o processo que permite que fragmentos de DNA se voltem a
ligar.
Para a transferência destes genes, é também necessária a existência de um vector, que será
a entidade que leva o material genético do genoma de onde foi retirado para o genoma que o
vai receber.
Os plasmídeos das bactérias são exemplos de vectores.
Técnica do DNA recombinante
A técnica do DNA recombinante permite combinar na mesma molécula de DNA genes
provenientes de fontes diferentes, mas não necessariamente de espécies diferentes, obtendo
uma molécula de RNA recombinante (rDNA).
Nesta técnica, recorre-se a enzimas de restrição para cortar o DNA em pontos específicos e
a ligases do DNA para reconstruir a molécula.
Obtenção e expressão da molécula de rDNA:
1. Selecção de uma molécula de DNA (a integrar) contendo um gene com interesse, que
se pretende transferir e clonar; selecção de um vector adequado (plasmídeo);
2. A molécula de DNA e o vector são tratados com a mesma enzima de restrição, que
corta as duas moléculas em regiões com a mesma sequência de nucleótidos;
3. Misturam-se os fragmentos de restrição da molécula de DNA e o vector, adicionando
ligases do DNA. O vector e os fragmentos de restrição emparelham pelas extremidades
coesivas e a ligase estabelece a ligação entre eles;
4. O vector, contendo o DNA dador, é transferido para uma célula/organismo receptor;
5. O DNA dador é incorporado no genoma da célula/organismo receptor, que passa a
possuir um DNA recombinante;
Os plasmídeos possuem genes que lhes conferem resistência a um antibiótico, permitindo
localizar as bactérias que têm o DNA recombinante. O cultivo de bactérias que foram
misturadas com plasmídeos num meio com esse antibiótico, é possível isolar as bactérias que
resistem – essas têm certamente os plasmídeos recombinantes, porque as que não têm
desaparecem com a aplicação do antibiótico.
Os vírus também podem ser utilizados como vectores. As células hospedeiras dos genes já
não são só bactérias, mas podem ser outras células, como leveduras e mesmo células
eucarióticas.
São comuns as plantas e os animais em cujo genoma foram introduzidos genes que
determinam características vantajosas, constituindo os OGM.
A técnica do rDNA é utilizada, por exemplo, na produção de insulina humana.
Técnica do DNA complementar
Os procariontes são organismos muito utilizados em Engenharia Genética como receptores
de DNA estranho porque:
São fáceis de cultivar,
Têm um crescimento rápido,
Processos bioquímicos bem conhecidos.
No entanto, os seres procariontes não processam o mRNA e se, em alternativa, recebem
genes com intrões, não são capazes de os retirar e a proteína produzida não é funcional.
Este problema é ultrapassado pela obtenção e transferência de DNA complementar ou
cDNA.
Para a técnica de DNA complementar são necessários:
Molécula de mRNA;
Transcriptase reversa (enzima que catalisa a formação da cadeia complementar do
DNA – transcriptase porque é um processo de transcrição, reversa porque é inverso
ao processo de transcrição da molécula de DNA em mRNA);
DNA polimerase – que catalisa a formação da cadeia complementar de DNA;
Nucleótidos livres.
O cDNA é uma molécula de DNA sem intrões, que é directamente transcrita numa molécula
de mRNA funcional. O processo de obtenção de cDNA é o seguinte:
1. Isola-se uma molécula de mRNA funcional das células;
2. Adiciona-se a trancriptase reversa e nucleótidos livres;
3. Junta-se uma enzima que degrada o mRNA que serviu de molde e DNA polimerase,
que catalisa a formação da cadeia complementar do DNA.
O cDNA pode ser inserido num procarionte através de um vector contendo o promotor e
sequências reguladoras.
Reacções de polimerização em Cadeia – PCR
O PCR é uma técnica que permite amplificar qualquer porção de DNA fora das células.
Esta técnica é útil para quando é necessária uma determinada quantidade de DNA que não
se possui, mas que pode ser obtido através desta técnica.
Esta técnica consiste nas seguintes etapas:
1. O fragmento de DNA a amplificar é aquecido de modo a separar as duas cadeias da
dupla hélice, quebrando as ligações entre os aminoácidos - DESNATURAÇÃO;
2. Obtêm-se duas cadeias simples;
3. São adicionados nucleótidos livres e DNA polimerase resistente ao calor – esta DNA
polimerase é obtida a partir de microrganismos termófilos, uma vez que vivem a temperaturas
muito elevadas, e aguentam ser mantidos às mesmas, enquanto a DNA polimerase
normalmente usada acaba por sofrer também DESNATURAÇÃO quando sujeita a temperaturas
muito elevadas;
4. A DNA polimerase catalisa a formação das cadeias complementares, restituindo a
dupla hélice, formando duas moléculas de DNA a partir de uma;
5. Arrefecimento das novas moléculas;
6. Repetição do processo – em cada ciclo a quantidade de DNA é duplicada.
Esta técnica permite a obtenção de biliões de cópias de uma porção de DNA em poucas
horas e é executada por aparelhos
DNA fingerprint
No genoma humano existem sequências de DNA repetitivas que são reconhecidas e
cortadas por determinadas enzimas de restrição.
O DNA fica então fragmentado – estes fragmentos apresentam tamanhos e composição
diferentes, variando de pessoa para pessoa.
Quando submetidos a técnicas como a electroforese, o resultado é um padrão de bandas
que difere de indivíduo para individuo, sendo possível identificar uma pessoa através destas
bandas, com quase (ou mesmo) 100% de certezas.
O processo de identificação por DNA fingerprint é feito da seguinte forma:
1. Obtenção de fragmentos da molécula de DNA, colocando em recipientes amostras de
DNA e enzimas de restrição, que a fragmentam nas respectivas zonas de restrição;
2. Os fragmentos obtidos são colocados num meio apropriado (por exemplo gel) e
quando submetidos a um campo eléctrico, deslocam-se até à extremidade oposta de onde
foram inseridos, a velocidades diferentes, consoante o tamanho e “peso” do fragmento;
3. Ao fim de algum tempo, os fragmentos localizam-se em diferentes secções do gel,
permitindo identificar um indivíduo pelo padrão obtido por electroforese.
Técnica
Aplicações
DNA
Recombinante
(rDNA)
Investigação fundamental – torna possível isolar genes de organismos
complexos e estudar as suas funções a nível molecular
Obtenção de organismos geneticamente modificados (OGM) –
organismos em cujo genoma foram introduzidos genes que conferem
características vantajosas. São usados:
-na produção de alimentos em maior quantidade e qualidade;
-na produção de grandes quantidades de substancias com aplicação
médica ou farmacêutica;
-na com aplicação industrial;
-biorremediação.
DNA
Complementar
(cDNA)
Polimerização
por reacção em
cadeia (PCR)
DNA fingerprint
Obtenção de cópias de genes que codificam produtos com interesse.
Obtenção de grandes quantidades de DNA em pouco tempo.
-Investigação criminal, forense e histórica;
-Determinação de paternidade.