O SISTEMA E A SÍNDROME PIRAMIDAIS
Dedico este opúsculo ao meu mestre, Luís Barraquer i Bordas, pela sua contribuição ao estudo do
sistema e da síndrome piramidais.
Introdução
A organização geral dos sistemas motores cerebrais depende da organização dos impulsos
motores superiores que chagam a unidade motora na medula e, se esquematiza, de acordo com
a diferenciação que existe entre dois sistemas: a) o sistema piramidal, b) o sistema ou sistemas
extrapiramidais.
A motricidade voluntária (MV) da qual o sistema piramidal faz parte é a mais evoluída das três
formas de atividade motora. Caracteriza-se pelo mais alto grau de independência em relação a
influencia imediata do meio, estando intimamente presa à formação da personalidade do
indivíduo. Pessoas de ambos os sexos têm formas especificas para realizar movimentos, os quais
foram aprendidos durante suas vidas dentro de um contexto cultural. Mas, nem todo movimento
vem a partir do sistema piramidal. Os movimentos automáticos originados filogeneticamente
antes do piramidal podem aparecer durante a marcha em várias situações. Numa atração
sensual os movimentos no homem são exacerbados nos ombros, enchendo o peito, abrindo os
braços, colocando uma das mãos na cintura; enquanto a mulher afasta um pouco os braços do
corpo, mantém o tronco superior mais fixo para mexer melhor os quadris, passa a mão no
cabelo, joga-o para trás ou para o lado.
Os centros motores somáticos trabalham juntos com os centros vegetativos durante a excitação
cortical para haver vasodilatação periférica devido ao maior gasto de energia, a fim de auxiliar na
contração muscular sem fadiga. Mas, a MV distingue-se da automática, principalmente pelo fato
de sua realização provocar muito maior fadiga. O cansaço produzido pelos movimentos
propositais, repetidos, de flexão e extensão dos membros inferiores, é muito maior do que o
determinado pelo mesmo conjunto de movimentos no ato automático da marcha: pensar em
andar cansa mais do que andar sem pensar.
1
Generalidades
O trato piramidal (TP) com seus 20 milhões de fibras é um dos sistemas motores que convergem
para as células do corno anterior da medula espinhal, embora, existem outros neurônios
motores superiores que não fazem parte desse sistema. O TP descrito em 1851, por Türck,
parece ser o primeiro conjunto de fibras reconhecido como um trato específico do encéfalo. O
trato piramidal é formado pelo trato córticoespinhal (TCE) e o trato córticobulbar (TCB) que são
usados quando for feita referências a eles. O TCB supre as estruturas do tronco cerebral no
mesmo caminho do TCE, que inerva a medula espinhal, mas não passa através da pirâmide
bulbar e, portanto, não é piramidal. Isso significa que existem neurônios que se projetam do
córtex para a medula espinhal sem fazer parte do sistema piramidal.
Anatomicamente o termo piramidal é relativamente bem definido, mas isso não justifica o termo
síndrome piramidal, para designar uma constelação de sintomas determinados. O termo
piramidal e extrapiramidal tem sido desprezado por alguns anatomistas, mas os neurologistas
continuam usando-os para diferenciar as lesões diretas ou piramidais das indiretas ou
extrapiramidais. Por definição, o trato piramidal são todas as fibras que cursam
longitudinalmente pela pirâmide bulbar, não importando seu local de origem no córtex cerebral
ou destino final na medula. As células piramidais fazem parte de quase todo o córtex cerebral,
mas apenas uma parte dos axônios dessas células passa pelas pirâmides bulbar. Portanto, as
únicas lesões que provocariam uma síndrome piramidal pura seriam interrupções à passagem
das fibras, onde as mesmas se encontram isoladas na pirâmide bulbar.
Do ponto de vista clínico o TCE é o principal sistema eferente através e, pelo qual, movimentos
voluntários são iniciados e realizados. Esse trato não é o único mecanismo cortical para a
realização dos movimentos; ele atua primariamente para integrar os movimentos discretos, finos
e altamente qualificados nas extremidades, como: mãos, pés e boca. O TCE é responsável pela
contração e inibição de músculos agonistas, ou graduar o relaxamento dos músculos
antagonistas necessária à realização de ações qualificadas. Por meio de sua integração e controle
as contrações de músculos individuais são fundidos dentro de uma ação motora complexa. O TCE
não funciona independentemente. Normalmente, e na presença de doença, está intimamente
integrado com outros níveis de atividade motora, como também com um fluxo constante de
2
impulsos sensitivos. O TCE no seu percurso com outras vias cortical e do tronco cerebral,
constantemente supre centros inferiores com impulsos que têm um efeito geralmente inibidor.
Doenças envolvendo os caminhos piramidais resultam na liberação do efeito inibidor, resultando
na hiperatividade e função autônoma dos segmentos afetados da medula espinhal.
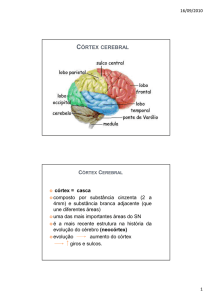
Córtex motor cerebral
Não existem critérios exatos para definir uma área cerebral como motora. O que sabemos é que
as camadas corticais II e IV recebem as vias sensitivas; as camadas II e V é ponto de partida para
as vias motoras; e as camadas I e VI estão envolvidas com as vias de associação. Atualmente,
uma área motora deve reunir os seguintes critérios: a) ter projeções para os motoneurônios
espinhais ou para os núcleos motores no tronco cerebral; b) possuir uma representação do
sistema motor, ou seja, dos músculos; c) ser ativada durante a execução ou o planejamento dos
movimentos voluntários. Baseados nesses critérios foram determinados as áreas motoras que
têm ligação com o sistema piramidal: 1. Córtex motor primário (MI); 2. Área prémotora (APM); 3.
Área motora ocular (AMC); 4. Área motora suplementar (AMS); 5. Área motora do sulco do
cíngulo (AMC); e 6. Área supressora (AS) (Fig.1).
Hanna Damasio, 05
3
Fig.1
Histologia do córtex motor cerebral
O córtex cerebral é formado por seis camadas. Segundo Ramon y Cajal o neocórtex possui dois
tipos distintos de células: neurônios piramidais, que constituem 72%, e os não piramidais, que
formam 28% restantes. Há uma grande variedade de tamanho entre esses neurônios, que
alcançam dimensões entre neurônios pequenos a gigantes. Os neurônios piramidais se
distribuem entre as camadas, II e IV do córtex cerebral. Embora a maioria dos neurônios
piramidais esteja situada nas camadas III e V do córtex cerebral. Existem dois tipos de células
piramidais, as rápidas, que transmitem impulsos fisiológicos a 27 m\s, e as lentas, que o fazem a
19 m\s. As rápidas caracterizam-se por apresentar poucas espinhas dendríticas, baixa resistência
de membrana, potenciais de ação (PA) breves e hiperpolarização póspotencial de curta duração,
o que lhes permitem responder a estímulos repetitivos de alta frequência. As lentas possuem
muitas espinhas dendríticas, alto resistência de frequência, PA de longa duração,
hiperpolarização póspotencial longa, que as fazem disparar com baixa frequência.
Os neurônios não piramidais são interneurônios, em forma de estrelas, que se subdivide em três
tipos. 1. Células granulares com espinhas dendríticas, que ocupam a camada IV cortical e
distribuem-se por todo neocórtex, mas não no motor; 2. Células em cesto, sem espinhas
dendríticas, cujo axônio tem direção horizontal, que inerva outros neurônios, inibindo-os
mediante ação do GABA que são retroalimentados pelos neurônios piramidais; 3. Células
bipolares, cujos axônios dividem-se em forma de T para inervar verticalmente os neurônios
superficiais e profundos. O neurotransmissor cortical excitatório é o glutamato e o aspartato,
embora alguns possam usar neuropeptídeos.
As áreas cerebrais da classificação de Brodmann que mais estão relacionadas com o sistema
piramidal são as seguintes: A área 8 – a excitação da porção superior da desta região (áreas
8αβδ) determina o aparecimento de movimentos conjugados dos olhos para o lado oposto,
abertura da rima palpebral e, às vezes, midríase. Além de responder a excitação do campo ocular
intuitivo. O estímulo da porção inferior da área 8γ produz inibição dos movimentos rítmicos
induzidos pela excitação da área 6bα durante a mastigação, a estimulação da área 8γ, provoca
inibição, além de receber estímulos de áreas corticais não intuitivas. A área 6 – a excitação da
área 6aα origina contralateral: movimentos de músculos isolados, devido à difusão cortical do
4
estimulo à área 4; e movimentos secundários prolongados e mais altamente organizados, cuja
distribuição somática varia segundo o ponto de excitação (na porção superior movimentos
apendiculares; na inferior, movimentos da face, mandíbula e língua). A área 6aβ determina
respostas semelhantes às produzidas na área 6aα, porém o seu limiar é mais elevado. São
característicos movimentos rítmicos e estereotipados dos membros, independentes da área 4, e
os movimentos adversivos da cabeça, olhos e tronco - campo adversivo de Foerster (Fig.2).
Fig.2
A área 6bα dá origem a movimentos
prolongados rítmicos e coordenados destes
territórios somáticos para mastigar, lamber,
deglutir, gritar e estalar a língua. A área 6bβ
determina lentificação do ritmo respiratório
até apnéia por transmissão do estímulo para
a área 13 de Brodmann, do lobo orbitário,
conhecida
Adams, 05
como
centro
respiratório
primário do córtex cerebral. A área 1, 3 e 2
estão relacionadas com o controle piramidal nos núcleos de Goll e Burdach no bulbo e no corno
posterior da medula espinhal, responsável pelo feed-back entre o estímulo sensitivo e o
movimento. Esses movimentos são bilaterais, embora que assimétricos, comunicam-se com as
áreas 6 e 4 via corpo caloso, cujos axônios saem das camadas IV a VI de ambas as áreas cerebrais.
As áreas 5 e 7 estão relacionadas com movimentos de praxia ideativa, ideomotora e cinestésica.
As áreas visuais enviam os estímulos
Fig. 3
para organização ocular ou motora
antes
de
descer
pelo
sistema
piramidal. A área 22 de Wernicke
envia estímulos sensoriais complexos
para as áreas oculares e motoras (Fig.
2 e 3).
O reflexo transcortical ocorre quando
Adams, 05
5
um músculo está contraindo e surge
uma sobrecarga ou uma mínima diminuição repentina da carga, os neurônios corticoespinhais,
que estão ativos durante a execução do movimento, aumentam ou diminuem seus disparos,
respectivamente. Isso se reflete nos músculos por um aumento ou uma diminuição da atividade
registrada no eletromiograma. Ou seja, existe uma compensação automática da força muscular
necessária para a realização de um movimento. A função desse mecanismo servo-assistido é
manter constante a força muscular. Sua origem reside nas aferências do fuso muscular ativadas
pela contração muscular. Esse reflexo transcortical, descrito por Phillips, é um reflexo miotático
ou de estiramento em paralelo com o da medula espinhal. Sua importância é demonstrada na
lesão do funículo posterior, que o suprime juntamente com o qual desaparece o ajuste
automático e a manutenção da força muscular. Isso impede que um indivíduo possa sustentar
uma xícara de café na mão pelo fato de a força de apreensão flutuar constantemente, e então
termine por soltá-la. A lesão dos fascículos cuneiformes suprime a ativação das células piramidais
do córtex motor contralateral, produzida pelo movimento passivo das articulações e pela
estimulação das aferências cutâneas. Isso indicaria que o funículo posterior seria o encarregado
de levar essas aferências.
Área motora primária (AMP)
Localiza-se no giro précentral, anterior ao sulco central ou a fissura de Rolando. O córtex da área
AMP ou MI é do tipo agranular e heterotípico e sua maior característica é a presença de
neurônios piramidais gigantes de Vladimir Betz, situadas na camada V do córtex cerebral da área
4 de Brodmann. Essa região tem o limiar de excitação mais baixo, para que a estimulação cause a
contração dos músculos do lado oposto do corpo mais rapidamente. Possui uma representação
dos músculos contralaterais do corpo; os da cabeça e da face ocupam a parte inferior do giro
précentral, enquanto que parte dos músculos do membro (pé) localiza-se na face interna do
hemisfério. Uma característica dessa representação é que os neurônios que vão inervar os
músculos distais localizam-se nas porções posteriores do giro précentral, em sua maioria na
borda anterior do sulco central e descem pelo feixe córticoespinhal lateral, enquanto os que vão
inervar os músculos axiais situam-se na parte anterior da área 4 de Brodmann descem pelo feixe
corticoespinhal anterior ou de Türck. Outra característica é de que nem todas as partes dessa
6
área têm uma representação igual, resultando numa figura distorcida. A face e a mão ocupam
uma grande porção da área 4 de Brodmann (Fig.4).
Fig. 4
O que está representado no
córtex
cerebral
são
os
movimentos realizados por
músculos
específicos
aquelas áreas e, não, os
músculos.
Isso
ocorre
porque as células da mesma
coluna do córtex motor se
projetam
na
espinhal
sobre
motoneurônios
medula
os
de
diferentes músculos, sendo
nesse caso, a coluna cortical
a entidade funcional do
DeJong, 05
movimento: para fazer a
flexão do membro superior
entram em ação o bíceps braquial, braquial e o braquiorradial. As aferências sensitivas para cada
coluna partem da superfície cutânea ou articular situada na direção do movimento que será
realizado, obedecendo ao padrão geral de somatotopia para o antebraço e mão. No
funcionamento do córtex motor, à estimulação de uma coluna se segue a inibição das colunas
vizinhas. Portanto, há inibição dos músculos para a extensão do antebraço enquanto são
ativados os músculos para a flexão do mesmo, fenômeno de inervação recíproca entre coluna
facilitadora e inibidora. Mas foi observado que um neurônio piramidal inerva mais de um
músculo agonista. Isso se deve a divisão do axônio na medula ou princípio da divergência, o que
contribui para superposição da representação. Isso permite que unidades motoras de um
músculo sejam ativadas enquanto outras do agonista facilitadas, podendo realizar uma ação
associada.
7
Área prémotora (APM)
Fig.5
A
definição
de
córtex
prémotor foi proposta por
A. W. Campbell, em 1905,
e a área correspondente
denominada como tal em
1929, por M. Hines. A APM
situa-se na porção inferior
da área 6 de Brodmann e
anterior
a
Brodmann,
intimamente
área
4 de
estando
relacionada
DeJong, 05
com a AMP ou MI, tanto
anatômica quanto funcionalmente. O córtex prémotor é histologicamente similar ao córtex
motor, mas faltam as células piramidais gigantes de Betz. Algumas fibras da área 6 de Brodmann
passam para a área 4 de Brodmann pelas fibras arciformes e depois descem com o TCE; outras
descem com as fibras piramidais. Existem mais fibras cruzadas do córtex motor do que do córtex
prémotor. Além do mais, a APM comunica-se com os gânglios da base e outras porções do
sistema extrapiramidal, incluindo o núcleo subtalâmico, núcleo rubro, colículo superior, núcleos
vestibulares, oliva inferior, e formação reticular (Fig. 5).
Estimulação da área 6 de Brodmann causa movimentos contraversivos da cabeça e do tronco. A
APM está envolvida com os movimentos guiados pelos estímulos visuais, auditivos e sensitivos. A
área motora suplementar é parte da área 6 que situa-se no aspecto medial do lobo préfrontal
anterior ao lobo paracentral (área 6aβ). Comunica-se com o córtex motor primário e com o
córtex motor suplementar contralateral. A APM parece estar envolvida com o planejamento e a
integração dos movimentos bilaterais do corpo. A APM também é somatotopicamente
organizada, mas o homúnculo é grosseiro e menos detalhado do que o homúnculo da AMP ou
MI. Na lesão da APM, o paciente apresenta apraxia ideomotora e dificuldade na programação de
movimentos guiados pela informação sensorial, nos movimentos iniciados por estímulos
8
sensoriais e, em alguns casos, em certos movimentos autoiniciados no espaço extrapessoal.
Participa na modificação dos programas motores já estabelecidos, com base na informação
sensorial l, como ocorre durante a exploração de objetos para seu reconhecimento estereognosia.
Área motora ocular (AMO)
Os olhos são controlados por duas áreas corticais, localizadas fora das áreas corticais motoras. O
chamado campo ocular frontal (COF) encontra-se na região inferior da área 8 de Brodmann e o
campo ocular suplementar (COS) está situado na parte rostrodorsal da área 6 de Brodmann.
Ambos os campos recebem informações das áreas visuais, com exceção do campo ocular frontal
que não recebe projeções do córtex estriado, mas apenas das áreas extraestriadas. Ambos os
campos projetam-se para os centros subcorticias óculomotores. A estimulação produz
movimentos conjugados para o lado oposto, podendo ser acompanhado de dilatação pupilar e
abertura palpebral. O COF é especializado na seleção de movimentos oculares determinados por
estímulos visuais. É ativado quando o indivíduo realiza movimentos sacádicos durante a leitura
ou quando um objeto chama a sua atenção. O COS intervém na seleção de movimentos oculares
não determinados por alvos visuais, o indivíduo decide onde olhar autoregerando seus
movimentos oculares. Lesão destrutiva unilateral do COF origina desvio transitório dos olhos
para o lado da lesão e impossibilita de movê-los e, lesão irritativa, desvio transitório para o lado
oposto da lesão, veja a fig. 4. Para maiores detalhes sobre AMO, veja neste site em: nervos óculomotores.
Área motora suplementar (AMS)
Descrita por Penfield e Welch, em 1949, foi assim denominada porque sua estimulação produz
movimentos complexos, variáveis e com limiares mais altos do que os da AMP ou MI. A área
motora suplementar situa-se na face medial dos hemisférios cerebrais superior ao giro do
cíngulo na área 6 de Brodmann, adiante do lóbulo paracentral. A AMS conecta-se com a medula
espinhal pela via corticoespinhal, a qual contribui com 12 a 18 % das suas fibras. Geralmente sua
função é bilateral ao corpo, embora ainda predomine as contralaterais. Uma lesão unilateral da
AMS durante primeira semana produz afasia transcortical motora e alterações dos movimentos
seqüenciais e simultâneos com o membro contralateral à lesão. Ao solicitar ao paciente que
feche o punho e, em seguida, que flexione o cotovelo, o paciente realiza o primeiro movimento e
9
Fig.6
demora executar o segundo
ou não realiza; ou bater na
mesa em sequência com a
mão fechada, o lado da mão
e a palma da mão. Não
realiza
movimentos
simultâneos como fletir o
cotovelo e fechar a mão.
Após a primeira semana da
lesão
DeJong, 05
incoordenação
permanece
bimanual,
que consiste na realização
de movimentos em espelho com ambas as mãos, ainda que possa realizar movimentos
individuais corretos. O paciente reproduz involuntariamente com a outra mão qualquer
movimento voluntário realizado com a mão volitiva. Isso dificulta o paciente andar de bicicleta,
dirigir um automóvel, etc. Na mesma lesão também persiste o reflexo de apreensão forçado.
Além do mais, a AMS é responsável por movimentos posturais, rítmicos; vocalização, bloqueio de
linguagem, dilatação pupilar e taquicardia. A AMS estaria relacionada com a programação dos
movimentos autoiniciados, complexos, sequenciais e simultâneos, assim como a coordenação
bilateral desses (Fig.6).
Área motora do sulco do cíngulo (AMC)
Localiza-se na face interna do hemisfério cerebral, no bordo inferior e no fundo do sulco do
cíngulo. É uma área descrita recentemente, constituída de uma zona anterior, que ocupa a área
24c, e outra posterior, a área 23c. Em ambas encontra-se uma representação somatomotora,
com os músculos da face na parte anterior seguidos do membro superior e inferior. A
estimulação dessas áreas provoca movimentos complexos nas expressões orofaciais, da cabeça e
da mastigação. Na área 23c, os neurônios se ativam 500 ms antes de começar o movimento
autoiniciado. A área 24c está relacionada com o significado do movimento autoiniciado. A AMC
da metade anterior do giro do cíngulo projeta-se para o córtex motor primário e também
10
contribui com as fibras descendentes do TCE e TCB. Existe também uma área motora secundaria
na profundidade do sulco central onde o giro précentral e o póscentral emergem, veja a fig. 6.
Áreas supressoras (AS)
A estimulação das áreas cerebrais de Mac Culloch, especialmente uma pequena região situada
entre as áreas 4 e 6 de Brodmann, identificada por Hines como área 4s, inibe o movimento
desencadeado pela área piramidal. Essa inibição pode ser obtida em outras áreas como 1s, 19s e
24s. As vias procedentes da área 4s descem juntas com as da via piramidal até a cápsula interna,
em seguida se separam do contingente piramidal para dirigir-se a substância reticular central do
bulbo. Quando esse centro inibidor está desconectado das áreas cerebrais supressoras, é incapaz
de manter uma atividade inibidora. Do centro bulbar nascem às fibras retículoespinhais que
terminam na medula espinhal, veja abaixo feixe retículoespinal.
Resumo
Metade a três quartos de todas as fibras piramidais originam-se nas áreas 4 e 6 de Brodmann
situadas anterior ao sulco central, e o resto vem das áreas posteriores do sulco central no lobo
parietal (áreas de Brodmann 1, 3, 2, 5, 7 e 40). A área 4 de Brodmann apresenta uma
somatotopia mais específica do que a da área 6 de Brodmann. Muitos neurônios da área
suplementar e partes da área prémotora e motora cortical foram ativadas em um programa de
descarga que precede o movimento planejado. Assim, os neurônios da área motora 4 foram
preparados para dar continuidade a ativação; foram desencadeado por um número de impulsos
corticais do lobo parietal, préfrontal, prémotor, e córtex auditivo e visual. Esse programa de
impulsos pode ocorrer na ausência de impulsos segmentar. A fonte desse programa de sinal foi
identificada principalmente na própria área suplementar do córtex motor, que parece estar sob
influência direta dos estímulos de prontidão, alçando a área préfrontal para planejar
movimentos e córtex parietal para iniciar atividade motora por percepção visual, auditiva e tátil.
Também existem fibras que alcançam área motora do sistema límbico, facilitando motivação e
atenção.
11
Anatomofisiologia do sistema piramidal
O
Fig.7
sistema
corticoespinhal
é
filogeneticamente o sistema motor
mais
recente,
desenvolvido
sendo
nos
totalmente
mamíferos
e
alcança seu mais alto grau nos
macacos e no homem. A aquisição
filogenética
da
fala
e
funções
complexas das mãos resultaram na
expansão
de
áreas
corticais
representando a língua, a boca, os
lábios, o dedo polegar, e demais
dedos das mãos, deslocando a
representação do córtex da mão e
perna para cima, e do pé e região
DeJong, 05
sacral sobre a superfície medial do
hemisfério. Áreas para a língua, face, e dedos são excepcionalmente grandes e fora de proporção
quando comparadas com as áreas da musculatura proximal. A extensão do giro précentral sobre
o aspecto medial do lobo frontal forma a porção anterior do lóbulo paracentral. Os neurônios
dessas áreas são importantes na regulação e regulação dos intestinos e do esfincter da bexiga.
Existem conexões recíprocas o córtex motor e o córtex somatosensitivo no giro póscentral. A
área MI recebe associação de fibras da área prémotora, motora suplementar e da ínsula. Essas
conexões são envolvidas na preparação e planejamento para os movimentos voluntários que são
depois executados pelo córtex motor primário. Existem também conexões entre o córtex
primário de ambos os hemisférios cerebrais. A divisão posterior do núcleo ventrolateral do
tálamo recebe input do cerebelo e envia para área 4 de Brodmann (Fig.7).
O termo trato piramidal surgiu porque a maioria de suas fibras passa pelas pirâmides bulbares. O
número de fibras piramidais que passam por cada pirâmide vai de 749.000 a 1.391.000, com
média de 1.087.200; somente e 20% a 30% originam-se da AMP ou MI, sendo que, somente 3%
12
Fig.8
das fibras piramidais ou 34.000 por AMP originamse das células de Betz. Além da contribuição da
área MI, o TCE contem fibras do córtex prémotor
(área 6 de Brodmann), a área motora suplementar
e as regiões adjacentes, e o lobo parietal (áreas 3,
1, 2, 5, 7, 40 e 43 de Brodmann), além das áreas
temporais e occipitais. Cerca de 60% das fibras
grossas são mielinizadas e 40% são pouco e não
mielinizadas; as últimas procedem de áreas
vegetativas corticais com importante papel nas
adaptações
atividade
vegetativas
motora
que
acompanham
a
(regulação
vasomotora).
A
maioria do TCE origina-se em partes iguais de 30%
DeJong, 05
para a área 4, área 6 de Brodmann e giro
póscentral, com uma contribuição adicional do córtex parietal (área 5) e outras porções do
cérebro, incluindo os lobos temporal e occipital, giro do cíngulo, e certos centros subcorticais. O
TCE é importante no controle discreto, das respostas motoras isoladas dos movimentos
voluntários finos individuais dos dedos. O TCE provê velocidade e agilidade dos movimentos
distais das extremidades. A APM e suas vias são destinadas para grandes respostas motoras
coordenadas, com mais movimentos estereotipados que são parcialmente automáticos e
envolvem o tronco e a porção proximal dos membros, e também com os mecanismos de
postura; sendo o principal componente cortical do sistema extrapiramidal (Fig. 8).
Na cápsula interna (CI), as fibras piramidais se projetam sob uma seguinte somatotopia. A CI é
formada por dois braços, um anterior e outro posterior, e um joelho. As fibras piramidais
oriundas da região craniofacial do córtex motor, área 4 de Brodmann, projetam-se no joelho da
CI; as demais fibras piramidais do membro superior, o tronco e membro inferior são distribuídas
na mesma ordem no braço posterior da CI. O restante do braço posterior da CI é ocupado por
fibras na sequência sensitivas, temporais, auditivas e visuais. O braço anterior da CI é ocupado
13
por fibras na ordem frontopontina e frontotalâmica. A próxima projeção importante das fibras
piramidais é no mesencéfalo ao formar parte do pedúnculo cerebral. O pedúnculo é uma massa
constituída de fibras piramidais e não piramidais que descem do córtex para terminar no tronco
cerebral e na medula espinhal. Considera-se que os dois terços mediais do pedúnculo contém o
TCE e o TCB. As fibras mais laterais do pedúnculo estão relacionadas com os membros inferiores,
as mais mediais com a musculatura da face e da laringe, e as intermediárias com os membros
superiores. As fibras frontopontinas são mediais, sendo que no encontro superior entre o terço
medial e o médio no pedúnculo, formando meia lua, passam as fibras do fascículo córticooculocefalico; já as fibras frontotemporo-occipitais são laterais, sendo que essas fibras se
distribuem em forma de triângulo: as temporais estão na base lateral, às occipitais ocupam o
ápice e as parietais a base medial (Fig. 9).
O TCB também conhecido
por fascículo corticonuclear
ou
geniculado
que
exaustivamente
foi
estudado
por Déjerine (1914) tem uma
dupla origem cortical; por um
lado, um quinto da área 4 de
Brodmann e, por outro, a
parte posterior da segunda
circunvolução frontal ou área
8 de Brodmann. O TCB
distribui-se
em
dois
fascículos: o corticonuclear e
o
fascículo aberrante de
Déjerine (FAD). a) O feixe
geniculado segue o curso do
contingente corticoespinhal,
do qual se separa no nível
Fig.9
14
dos núcleos de origem dos
nervos bulbopontinos, fazendo cruzamento parcial, já que certo numero de fibras acabam nos
núcleos ipsilaterais. Mediante essa inervação bilateral se consegue uma dupla representação
cortical de importantes funções de nervos cranianos. b) As fibras corticobulbares aberrantes
dividem-se em: interna, externa e direta ou feixe aberrante de Pick, esse último destinado ao
núcleo do facial do lado oposto, veja abaixo. O FAD cruzado é mais denso do que o direto e
contém maior numero de sinapse também. O FAD desce junto com o corticonuclear, mas o
cruzamento não se faz de uma vez, mas gradualmente, separam-se do anterior por pequenos
fascículos a distintos níveis do tronco
cerebral, realizando sinapses tanto
ipsilateral
como
contralateral.
Os
principais fascículos aberrantes que
chegam ao tronco cerebral, que se
distribuem de cima para baixo e no
sentido
anteroposterior
são
os
seguintes:
a) fibras aberrantes do pedúnculo
formam
dois
corticobulbar
feixes,
o
medial
e
fascículo
o
lateral.
Descem pelo território do leminisco
medial e proporcionam fibras aos
núcleos do III, VI e XI nervos cranianos.
Junto com esses fascículos correm
algumas fibras destinadas até aos
segmentos
medula.
cervicais
Esse
superiores
grupo
de
da
fibras
aberrantes governa os movimentos dos
olhos e os movimentos associados à
cabeça. b) As fibras aberrantes da
Fig.10
Puvanendran, 78
ponte que se unem as anteriores no
leminisco medial, terminam nos núcleos motores do trigêmio que envia axônios para os
15
músculos mastigadores; para os núcleos do hipoglosso e o núcleo ambíguo que enviam axônios
para os músculos da laringe e rotadores da cabeça. Mas, também pelas fibras aberrantes
pontinas passam fibras destinadas ao núcleo do facial, desde o pedúnculo cerebral, essas fibras
abandonaram a via piramidal para situar-se na parte interna da cinta de Reil. Contudo, os
músculos que recebem fibras diretas do TCB são a porção superior da coluna cinzenta do facial e
do núcleo do hipoglosso. c) As fibras aberrantes bulbopontinas deixam o tronco principal do
sistema piramidal próximo do nível do sulco entre a ponte e o bulbo. As fibras anteriores são
reforçadas ao proporcionar fibras ao núcleo motor do facial e, também, enviar outras ao núcleo
ambíguo e o nervo hipoglosso. Ainda merece destaque, algumas fibras do feixe corticobulbar
direto que descem pelo tronco cerebral até a porção ventromedial do bulbo em nível superior e
voltam para o núcleo do facial do lado oposto, consagradas como as fibras do trato aberrante de
Pick (Fig.10).
Fig.11
DeJong, 05
As fibras do TCB terminam de duas
maneiras:
diretamente
nos
motoneurônios α dos núcleos cranianos e
fazendo sinapse com células da formação
reticular próximas aos centros de controle
dos nervos. As fibras do TCB que alcançam
os núcleos sensitivos do trigêmio e nos
núcleos de Goll e de Burdach no bulbo,
exercem
um
controle
centrífugo
da
entrada sensitiva, como um tipo de feedback. Antes de chegar aos núcleos
motores respectivos, às fibras do TCB se
entrecruzam na sua maioria na rafe
mediana. Essa topografia particular de
como terminam as fibras do TCB, nos
explica porque a lesão da cinta de Reil
mediana, geralmente ocasiona o desvio conjugado da cabeça e dos olhos, veja abaixo a fig. 11.
16
Os axônios dos neurônios motores do giro précentral descem através da coroa radiada e do
braço posterior da cápsula interna, onde as fibras corticobulbares são anteriores, seguidas
posteriormente por aquelas das extremidades superiores, tronco, e extremidades inferiores. As
fibras piramidais são de distintos calibres; as de maior diâmetro vindas das células gigantes de
Betz conduzem uma velocidade de 60 a 65 metros por segundos. As fibras piramidais mais finas
conduzem os impulsos volitivos em média de 18 metros por segundo. Cerca de 90% das fibras do
TCE são de axônios pouco mielinizados com diâmetro que varia de 1 a 4 milimicra, e a maioria
com diâmetro de 5 a 10 milimicra. Um pequeno número de fibras que surgem das células de Betz
tem diâmetros de 10 a 22 milimicra. O membro posterior da cápsula interna é a porção entre o
núcleo lenticular e o tálamo, e a parte retrolenticular situa-se posterior para o núcleo. Na parte
rostral da cápsula interna, o TCE situa-se na porção anterior do membro posterior. Na cápsula as
fibras do TCE distribuem-se posteriormente e ocupam a posição no terço posterior do membro
posterior da cápsula interna. Em nível do mesencéfalo, o trato piramidal contém 1 milhão de
fibras que atravessam os três quintos mediais do pedúnculo cerebral, distribuídas na seguinte
somatotopia da linha media para lateral: face, braço, tronco e perna. A maioria das fibras do TCB
decussa antes de fazer sinapse com os núcleos dos nervos cranianos, mas a maioria da inervação
cortical para os centros do tronco cerebral é cruzada e não cruzada. Depois, as fibras
descendentes passam através da porção basilar da ponte em forma de fascículos distribuídos
difusamente, entremeados com as fibras arciformes e os núcleos pontinos da ponte para entrar
no bulbo. Na porção caudal do bulbo, o TCE forma duas colunas chamadas de pirâmides bulbares
na base do bulbo e divide-se em três tratos distintos (Fig.11).
Aproximadamente 90% das fibras cruzam na decussação das pirâmides ou cruzamento de
Mistichelli, com aquelas destinadas aos membros superiores mais anteriores e mediais e aquelas
para os membros inferiores posteriores e laterais. Existe considerável variação na proporção do
cruzamento e do não cruzamento das fibras do TCE no homem que varia de indivíduo para
indivíduo, fato importante na recuperação motora. As pirâmides também contêm outros tipos de
fibras que não são piramidais. Ressonância magnética funcional, potencial evocado motor e
somatosensitivo confirmaram hemiplegia ipsilateral em um paciente que teve hemorragia
cerebral. Foram capazes de demonstrar que os membros afetados eram controlados pelo córtex
cerebral ipsilateral. Em alguns casos um TCE não cruzado está associado à anomalia congênita de
17
vários
tipos.
As
pirâmides
bulbares
apresentam somatotopia da passagem do
trato piramidal em cada lado do bulbo. No
corte axial do bulbo, em nível das
pirâmides,
existe
distribuição
somatotópica das fibras piramidais no
sentido pósteroanterior: perna, tronco,
braço e face. Acima das pirâmides bulbares
Fig.12
Brodal, 84
aparece a somatotopia do leminisco
medial: perna, tronco, braço e face
(Fig.12).
No homem as fibras piramidais que chegam à medula são divididas em três fascículos
independentes: lateral ou cruzado, anterior ou direto e o homolateral. As fibras que decussam
descem pelo funículo lateral da medula no TCE lateral, situado ânterolateral ao corno posterior,
medial ao trato espinocerebelar posterior,
e posterior ao plano dos ligamentos
denteados para suprir os músculos do lado
oposto do corpo. Na medula lombossacra,
onde não existe o trato espinocerebelar
posterior, o TCE limita a superfície da
medula. O TCE lateral pode também conter
outras fibras corticofugais, como algumas
fibras ascendentes. Cerca de 50% das
fibras do TCE lateral terminam na região
cervical, 20% na torácica e 30% na
lombossacra, mas o trato termina no nível
de S3 a S4. O TCE anterior ou feixe de
Türck é menor do que o TCE lateral,
formado por uma porção menor das fibras
Fig.13
18
Capenter, 80
piramidais, geralmente contém cerca de
8% das fibras; não cruza nas pirâmides, desce pelo funículo anterior da medula e geralmente não
se estende além da porção medial da região torácica. O feixe de Türck cruza nos segmentos
medulares pela comissura branca anterior e termina na lâmina VII para suprir os músculos axiais.
As fibras do TCE ânterolateral de Barnes ou fascículo piramidal homolateral de Dèjérine não são
cruzadas, contam com 2%, são de calibre pequeno, e descem em posição mais anterior no
funículo lateral que as fibras cruzadas do TCE lateral, também não cruzam na medula e terminam
na base da coluna posterior e substância cinzenta intermédia da medula. A existência dessas
fibras homolaterais nos explica o motivo pelo qual a destruição do fascículo piramidal em curso
de seu trajeto encefálico determina não somente uma hemiplegia do lado oposto, mas também
do lado correspondente ao da lesão, certa debilidade muscular e reflexos vivos (Fig.13).
As fibras anteriores da região précentral terminam na região ventromedial da medula, enquanto
as mais próximas do sulco précentral terminam na região dorsolateral da medula. Os axônios dos
TCE e TCB terminam nos núcleos motores dos nervos cranianos e nas células do corno anterior
da medula, terminando primariamente nas lâminas IV a VI, lâmina VII, lâmina VIII, e lâmina IX do
lado oposto ao hemisfério cerebral de origem. Fibras das áreas 4 e 6 terminam mais
ventralmente, principalmente na lâmina VII e VIII, e são concentradas nas intumescências
cervical e lombossacra. A maioria das fibras do TCE faz sinapse nos neurônios γ do núcleo
intermédiomedial de Cajal, que modifica a descarga dos fusos musculares e se mede o
movimento. A intervenção do sistema gama retarda (30 milisegundos) a iniciação do movimento,
enquanto a força do mesmo é graduada constantemente. A informação proprioceptiva é de
grande valor para a realização dos movimentos para o sistema γ atuar nos movimentos motores
finos, qualificados e discretos da porção mais distal das extremidades. Depois, impulsos viajam
dos núcleos motores do tronco cerebral e das células do corno anterior da medula para as
junções musculares estriadas. Mas, cerca de 10% a 20% das fibras piramidais termina
diretamente sobre os neurônios motores α na lâmina IX de Rexed. Essas fibras formam a via
corticomotoneuronal que projetam diretamente do córtex motor primário ou área 4 para o
corno anterior da medula que permite a realização de movimentos bruscos, rápidos,
possibilitando a interrupção rápida da atividade periférica. Uma única fibra do TCE inerva mais de
um neurônio na medula, e alguns provavelmente inervam muitos. O trato piramidal afeta a
atividade, tanto dos neurônios motores α como γ. A coativação α-γ serve para manter um nível
19
consistente
de
estiramento
e
relaxamento das fibras
extrafusais. As fibras do
lobo parietal que descem
no
trato
piramidal
terminam
sobre
neurônios sensitivos no
corno
Fig.14
Netter, 87
posterior
medula,
regulando
entrada
de
da
a
impulsos
sensitivos ascendentes. Essas fibras constituem, provavelmente, um mecanismo sensitivo do tipo
feed-back, capaz de influenciar os grupos celulares secundários, da qual o córtex recebe
informação (Fig.14).
RESUMO ANATOMOFISIOLÓGICO DAS VIAS MOTORAS DESCENDENTES
Do ponto de vista filogenético, o controle motor voluntário é: extrapiramidal em espécies
submamíferos (nos vertebrados inferiores não existe feixe piramidal), pirâmido-extrapiramidal
em mamífero não humano e, essencialmente piramidal no homem; fica mielinizado nos
primeiros 2 anos de vida. O sistema piramidal aparece tardiamente tanto na filogênese como na
ontogênese do sistema nervoso. Para alguns anatomistas, a classificação clínica de sistema
piramidal e extrapiramidal dificulta a compreensão das vias motoras descendentes. A partir da
década de 60, estuda-se uma classificação anatomofisiológica em sistema lateral e medial. Os
feixes que se originam no córtex cerebral e no mesencéfalo são geralmente cruzados e fazem
parte do: sistema lateral.
Do córtex cerebral, áreas 4 e 6 de Brodmann, sai o TCE que desce pela coroa radiada, dois terços
posteriores do braço posterior da cápsula interna, o terço médio do pedúnculo mesencefálico
lateral ao lado do TCB; o TCE organiza-se em fascículos distribuídos difusamente na ponte e
entremeados pelas fibras arciformes da ponte; e no bulbo forma um feixe compacto em forma
de cone que ao chegar na porção caudal do bulbo cruza a linha média com a maioria das fibras
20
para o lado oposto;
desce pelo funículo
lateral
da
medula,
termina nas lâminas
IV, V, VI, VII e IX,
sendo que a lâmina IV
e V recebem as fibras
do
TCE
que
se
originaram das áreas
1, 2 e 3 de Brodmann
no giro póscentral do
Fig.15
Greenstein & Greenstein, 2000
lobo
parietal
para
controlar a entrada de informação sensitiva superficial e profunda; os axônios que saem das
lâminas inervam os músculos das extremidades do corpo, especialmente os músculos intrínsecos
das mãos, do pés e os orbiculares da boca, para a realização de movimentos fino, suave,
qualificado e hábeis, prevalecendo a destreza (Fig.15).
Do mesencéfalo, da porção magnocelular do núcleo rubro origina-se o feixe rubroespinhal (FRE)
ou feixe prépiramidal de André Thomas. O núcleo rubro (NR) contém alto teor de ferro e é
coberto por uma espécie de cápsula que consiste de fibras aferentes e eferentes. A partir dos
primatas, o NR subdivide-se em uma parte cranial ou parvocelular (constituído de células
pequenas) e outra caudal ou magnocelular (formado por células grandes multipolares). A partir
do homem, existem quatro grupos de células na parte magnocelular com 150 a 200 células
grandes, mas na escala filogenética a proporção de células grandes decresce. O FRE ou feixe de
von Monakow é formado por fibras de vários diâmetros com velocidade de condução que varia
de 31 a 120 m\s. Logo após originar-se cruza no mesencéfalo na decussação tegmentar ventral
de Meynert, em seguida desce ipsilateral para integrar o funículo lateral da medula. O FRE tem
somatotopia no seu núcleo e medula espinhal. As fibras que terminam na região cervical
originam-se na porção dorsal e dorsomedial do núcleo rubro, as fibras de projeção lombossacra
originam-se na porção ventral e ventrolateral do núcleo rubro, as fibras do segmento torácico da
medula originam-se da porção intermediaria do núcleo rubro. A estimulação do NR ativa
21
motoneurônios flexores contralaterais com potenciais excitatórios póssinapticos registrados
intracelular, enquanto motoneurônios extensores contralaterais emitem potenciais inibitórios
póssinapticos. Os motoneurônios estáticos gama dos músculos flexores facilitam e os dos
músculos extensores inibidos. O FRE termina nas lâminas V, VI e VII da medula onde ficam
entremescladas com as do feixe corticoespinhal lateral. O FRE controla o tônus na musculatura
flexora, atuando nos motoneurônios alfa tanto flexores como extensores, excitando os flexores e
inibindo os extensores, além de inibir a ação das fibras aferentes musculares vindas dos ONT de
Golgi e das vindo da pele que terminam na asa posterior, controlando o aparelho motor sem
interferência das vias reflexas da medula (Tabela 1).
Características dos dois Sistemas de Vias Descendentes – Tabela 1 - Lent, 05
Sistema Lateral
Origem
Feixe
Lateralidade
Terminação
Função
Córtex cerebral
(áreas 6 e 4
Brodmann)
Córticoespinhal
Lateral
Contralateral
(decussação piramidal)
Moto- e
interneurônios
laterais
Movimentos
apendiculares voluntários
Núcleo rubro
Rubroespinhal
(feixe de von
Monakow)
Contralateral (cruza no
tegmento ventral
mesencefálico)
Moto- e
interneurônios
laterais
Movimentos
apendiculares voluntários
Sistema Medial
Origem
22
Feixe
Lateralidade
Terminação
Função
Córtex cerebral
(áreas 6 e 4
Brodmann)
Córticoespinhal
medial (feixe de
Türck)
Moto- e
Bilateral (cruzamento parcial
interneurônios
na medula)
mediais
Movimentos axiais
voluntários
Colículo Superior
Tectoespinhal
Contralateral (cruza no
tegmento dorsal do
mesencéfalo)
Moto- e
interneurônios
mediais
Orientação
Sensoriomotora da cabeça
Formação Reticular
pontina
Retículoespinhal
pontino
Ipsilateral
Moto- e
interneurônios
mediais
Ajustes posturais
antecipatórios
Formação reticular
bulbar
Retículoespinhal
bulbar
Ipsilateral
Moto- e
interneurônios
mediais
Ajustes posturais
Antecipatórios
Núcleo Vestibular
Lateral (Núcleo de
Deiters)
Vestíbuloespinhal
lateral
Ipsolateral
Moto- e
interneurônios
mediais
Ajustes posturais para a
manutenção do equilíbrio
corporal
Núcleo Vestibular
Medial
(Núcleo de Schwalbe)
Vestíbuloespinhal
medial
Bilateral
Moto- e
interneurônios
mediais
Ajustes posturais da cabeça e
tronco
Os feixes que se originam no mesencéfalo, ponte e bulbo são ipsilaterais e fazem parte do:
sistema medial.
Do córtex cerebral, áreas 4 e 6 de Brodmann, sai um pequeno contingente de fibras que cruza
parcialmente no bulbo, menos do que as fibras do TCE, desce pelo funículo ventromedial
formando o trato corticoespinhal ventral, cruza na comissura branca anterior da medula e
termina na lâmina VII, para inervar os músculos proximais dos apêndices, braços e coxas, e a
musculatura do tronco. Origina-se do mesencéfalo o feixe tectoespinhal (FTE) ou fascículo de
Löwenthal, a partir das camadas mais profundas dos colículos superiores. O FTE cruza na
decussação tegmentar dorsal, e desce junto do FLM no tronco cerebral e na medula pelo
funículo anterior até os quatro primeiros segmentos cervicais; termina nas lâminas VI, VII e VIII
sem fazer sinapse direta nos motoneurônios α e, sim, por via dos neurônios internunciais. Essas
fibras constituem o braço de descarga de um arco reflexo óptico através do qual se produzem
movimentos defensivos e reflexos de fuga. Esse feixe tem como principal função mediar os
movimentos posturais reflexos, em resposta aos estímulos visuais e, talvez, auditivos (girando os
olhos e a cabeça para o lado). Recebe influência do córtex cerebral, principalmente da área 18 de
Brodmann do lobo occipital.
O feixe vestíbuloespinhal (FVE) forma dois feixes: o lateral e o medial. O lateral origina-se no
núcleo de Deiters (situado no assoalho do IV ventrículo, na altura da área vestibular com células
grandes ou de Deiters e células pequenas) e descendente do nervo vestibular. O FVE lateral
desce ipsilateral pela parte mais anterior do funículo lateral da medula numa velocidade de
condução que varia de 24 a 140 m\s. As fibras que saem da parte dorsolateral do núcleo
vestibular lateral projetam-se no segmento lombossacral da medula espinhal e as fibras que
saem da porção rostrocaudal projetam-se para os segmentos cervicais da medula. O FVE lateral
termina nos neurônios α e γ na lâmina VIII, e parte das lâminas VII e VIII. A influência do sobre os
neurônios gama parece ser exercida sobre neurônios estáticos apenas, monossinápticos, sendo
excitatórios. Motoneurônios gama flexores e extensores parecem ser facilitados e inibidos,
respectivamente. Motoneurônios α e γ dentro de um pool de motoneurônios parecem ser
ativados em paralelo pelo núcleo de Deiters ao utilizar as células grandes e pequenas, e suas
atividades parecem ser intimamente relacionadas. O exemplo comum ocorre durante a marcha
23
automática, quando um membro está em flexão o outro em extensão. O FVE lateral transmite
aos motoneurônios os influxos que chegam à área vestibular desde o labirinto posterior, o que
permite interferir na produção dos reflexos estáticos e cinéticos do equilíbrio, ou seja, os reflexos
que mantém a atitude ou postura do corpo. Ainda influencia no funcionamento do arco reflexo
miotático, reforçando a sua atividade. Considera-se que o FVE lateral exerce influencia
facilitadora sobre a atividade reflexa somática da medula e sobre os mecanismos que controlam
o tônus dos músculos extensores. O feixe vestibuloespinhal (FVE) medial origina-se dos núcleos
vestibulares mediais bilaterais, núcleos de Schwalbe, situados na ponte inferior, cruzam em
seguida para formar o FVE medial, desce pela parte medial da medula e terminam nos
motoneurônios e neurônios internunciais na porção média da medula torácica, principalmente
na lâmina VII de Rexed. Sua principal função está relacionada com os ajustes posturais da cabeça
e membros superiores.
O feixe retículoespinhal (FRE) é dividido em três: ventral, medial e lateral. O FRE lateral origina-se
nos neurônios dos núcleos pontinos oral e caudal da formação reticular (zona facilitadora). O FRE
pontino é quase ipsilateral e desce pela parte medial do funículo anterior e cobre toda extensão
da medula numa velocidade de condução de 101 m\s; associa-se ao FLM até a medula cervical
superior, exercendo uma função facilitadora sobre diferentes tipos de motoneurônios da lâmina
VIII, e parte da lâmina VII. Sua principal função é a de controlar os movimentos posturais dos
músculos axiais e da parte proximal dos membros, principalmente do pescoço. O feixe
retículoespinhal (FRE) medial e o ventral originam-se da porção bulbar da formação reticular
(zona inibidora ou supressora) principalmente no núcleo reticular gigantocelular. O FRE medial
desce pela parte mais interna do funículo anterior da medula. O FRE ventral desce pelo funículo
anterior da medula numa velocidade de condução de 70 m\s. O FRE ventral e o medial
constituem um circuito neuronal numa via inibidora do arco reflexo miotático que começa no
sulco prérolandico do córtex frontal na área 4S de Marion Hines, e termina nos motoneurônios
da asa anterior da medula, passando através da zona inibidora da formação reticular. Essa via
tem dois neurônios: do córtex cerebral e da formação reticular bulbar. O axônio da primeira é
uma fibra justapiramidal que desce junto co o feixe piramidal até a ponte de Varolio, em cujo
ponto se separa para alcançar a formação reticular bulbar. Na medula o FRE ventral e o medial
24
terminam na lâmina VII do sistema γ (juntas com fibras do TCE e rubroespinhal), e algumas na
lâmina I que controlam a entrada de informação para a formação reticular para regular o tônus.
Fig. 16
O feixe solitário espinhal
de Rothman ou fascículo
bulboespinhal visceral de
Allen é parte integrante
do FRE lateral, mas ás
vezes
está
separado.
Trata-se de um grupo de
células nascidas no setor
da
Lent, 05
formação
reticular
próximas ao núcleo do
nervo vago, as quais se comportam como centro respiratório e vasomotor. As fibras desse
fascículo terminam nos centros da asa lateral da medula que governa a respiração e a função
vasomotora, integradas aos motoneurônios que inervam o diafragma e os músculos intercostais.
Além do mais, fibras autônomas descendentes dos níveis superiores acompanham o FRE na
regulação noradrenérgica na pressão arterial. A classificação do sistema piramidal e
extrapiramidal, descrita acima, ainda não tem sustentação clínica, mas pode ser útil para pessoas
experientes (Fig.16).
FISIOPATOLOGIA E SEMIOLOGIA DA SÍNDROME PIRAMIDAL
Do ponto de vista fisiopatológico o sistema piramidal é constituído por dois componentes:
ortopiramidal e justapiramidal. O primeiro é um feixe que se inicia no córtex e termina nos
segmentos da medula, nos neurônios internunciais, neurônios α e γ, sem fazer sinapse durante
seu percurso. O segundo é um feixe que se inicia nas áreas supressoras da área 4 e área 6 ou
suplementar; ao descer pelo tronco cerebral não passa pelas pirâmides bulbares; faz sinapse na
formação reticular inibidora situada na porção ventromedial do bulbo; projetando-se para os
segmentos medulares pelo FRE inibidor com função de inibir o reflexo miotático. Um terceiro
componente contíguo ao primeiro inicia-se das áreas 1, 2 e 3 do lobo parietal para terminar na
25
asa posterior dos segmentos da medula e, parte dele, termina no núcleo do trigêmio e de Goll e
Burdach e recebe o nome de TCB. Sua função é a de controlar os impulsos que chegam à medula
para em seguida subirem ao córtex parietal, realizando um feed-back negativo.
O termo plegia serve para motricidade e sensibilidade: hemiplegia motora e hemiplegia sensitiva.
Ao longo do tempo, o nome hemiplegia tornou-se um sinônimo de déficit motor no hemicorpo,
mas não déficit de sensibilidade. Conforme a distribuição somática das paralisias o déficit motor
pode ser chamado de: monoplegia braquial e crural; hemiplegia quando todo o dimídio corporal
está comprometido, incluindo a hemitesta; plegia dos membros homolaterais, incluindo o facial
supranuclear ou hemiplegia poupando a hemitesta; paraplegia braquial e paraplegia crural;
triplegia é a fraqueza motora em três membros causada por uma mesma lesão (veja a
decussação das pirâmides), quando causado por duas lesões trata-se de hemiplegia e
monoplegia; tetraplegia é a paralisia de quatro membros provocada por uma lesão (meningeoma
do forame magno pode começar a dar sintomas de fraqueza espástica de um membro, seguido
em forma de “U” ou síndrome em “U”); diplegia é uma hemiplegia dupla provocada por duas
lesões distintas. Conforme a distribuição da proporção a hemiplegia pode ser classificada como:
desproporcionada de predomínio braquial, crural, facial, faciobraquial (formas semiológicas
características das embolias cerebrais que comprometem a artéria silviana na região da coroa
radiada); as hemiplegias podem ser alternas quando os membros homolaterais comprometidos
estão associados no lado contralateral a um nervo craniano: paralisia facial ou estrabismo
divergente ou desvio da língua, etc; a hemiplegia cruzada ocorre quando o déficit motor alterna
entre os membros sem afetar os nervos cranianos: um membro superior homolateral à lesão e
um membro inferior contralateral à lesão; a hemiplegia pode ser pura, quando exclusivamente
motora; complicada quando acompanhada de síndrome sensitiva, atáxica, amiotrófica, distônica,
etc.
Semiologia da síndrome piramidal
Costuma-se designar como síndrome piramidal deficitária ou de liberação ou ambas, ao conjunto
de sintomas e sinais observáveis em patologias que afetam a via piramidal. Geralmente, a fase
deficitária aparece em episódios agudo da lesão piramidal, fase de choque; posteriormente
seguida pela fase de liberação que surge com a presença do sinal de Babinski. A síndrome
26
piramidal destrutiva foi descrita por Barré, sem que essa falha da função piramidal, implique
necessariamente na presença de uma lesão objetiva. A síndrome piramidal irritativa ou clássica
foi descrita primeiro, especialmente por Babinski, a qual determina descarga epiléptica focal.
Sintomas deficitários da síndrome piramidal de Barré
A síndrome piramidal deficitária por lesão ortopiramidal ocorre por perda da inibição. O déficit
de energia pode ser cinético com debilidade dos movimentos voluntários e, estático, com
incapacidade de manutenção de atitudes forçadas. O déficit motor cortical ortopiramidal
distribui-se na região dos dedos polegar e indicador, dificultando o movimento em pinça,
observado na manobra dinâmica de Müller Fischer. O déficit motor córticosubcortical
ortopiramidal distribui-se nos músculos extensores do punho, dificultando a extensão do punho
detectado pela manobra de Strauss. paciente estende o punho contra a resistência da mão do
médico, havendo déficit motor. Os músculos extensores do punho são filogeneticamente mais
recentes do que os músculos flexores, portanto alteram-se antes. Os músculos flexores do punho
são inervados pelo feixe rubroespinhal que é controlado, principalmente pela área 6 de
Brodmann. O déficit motor peculiar da SP se distribui habitualmente segundo uma topografia
eletiva, que realiza a postura de Wernicke-Mann, constituída por adução do braço, flexão e
pronação do antebraço, flexão do punho e dos dedos da mão com o polegar aduzido; extensão
do membro inferior, pé varo e em ligeiro equinismo.
Os reflexos podem ser divididos em proprioceptivos, exteroceptivos e visceroceptivos. Os
proprioceptivos dependem da excitação de receptores com sede nos músculos, tendões, sáculo,
utrículo e canais semicirculares. Os exteroceptivos são captados em receptores situados na pele,
ou em mucosas externas como córnea, faringe, etc. Os visceroceptivos relacionam-se com o
setor vegetativo. Os proprioceptivos são: reflexos de tração muscular ou miotáticos que são
captados pelos receptores do fuso muscular e pelos órgãos neurotendíneos de Golgi, conduzidos
à medula pelas fibras mielínicas tipo I e II de Lloyd; e pelos reflexos labirínticos ou tônicos. Ao
serem pesquisadas duas respostas podem ser obtidas: clônica e tônica. Na primeira, os reflexos
proprioceptivos estimulam-se as terminações anuloespirais que disparam aos motoneurônios
medulares α-1, provocando os reflexos profundos.
27
Os reflexos profundos se dividem em apendiculares e axiais. Entre os primeiros existem: nos
membros superiores, o bicipital realiza flexão e supinação do antebraço, inervado pelo
musculocutâneo e raízes de C5-C6; o tricipital realiza a extensão do antebraço, inervado pelo
radial e raízes de C7-C8 e, na resposta invertida do tricipital, existe flexão no antebraço por lesão
aferente do arco reflexo nas lesões nos segmentos C7-C8, principalmente quando existe
elemento de espasticidade como na espondilose cervical com rediculomielopatia; o estilorradial
realiza pronação e ligeira flexão do antebraço, inervado pelo radial e raízes de C5-C6
braquiorradial e C7,C8,T1 flexores dos dedos; nos membros inferiores o patelar realiza a
extensão da perna, inervado pelo femoral e as raízes de L2-L4; e o aquileu realiza a extensão do
pé, inervado pelo tibial e as raízes L5-S2. A resposta clínica desses reflexos pode ser: abolido,
diminuído, presente, vivo (caracteriza-se por resposta rápida e amplitude aumentada) e exaltado
(acrescenta-se ao reflexo vivo, o aumento da área reflexógena). Outra obtenção é usada como: 0
abolido, 1 + (presente diminuído), 2 + (normal), 3 + (aumentado mas não necessariamente
patológico) e 4 + (marcadamente hiperativo, patológico, com aumento da área de percussão). O
sinal + após o número é mais tradicional do que informativa.
Os reflexos axiais incluem os axiais da face e do tronco. Os primeiros são: o nasopalpebral que
realiza oclusão da rima palpebral, inervação aferente trigêmio e eferente facial; o orbicular dos
lábios realiza protusão dos lábios, inervado pelo trigêmio e facial; e o mandibular realiza
elevação da mandíbula, inervado pela aferência e eferência do trigêmio - todos os três têm seu
centro reflexo na ponte. Os reflexos do tronco são os reflexos abdominais profundos, sendo mais
importante o mediopúbco, que realiza contração ipsilateral dos músculos retos abdominais,
desvio da cicatriz umbilical e adução das coxas, inervado pelos intercostais inferiores, ilíoinguinal,
ilíohipogástrico, obiturador e as raízes T6-T12.
Os reflexos tônicos são segmentares e cervicais. Os primeiros sustentam as atividades posturais e
participam na regulação dos movimentos regulada pelos circuitos reverberantes γ e as vias
descendentes, originadas na formação reticular, núcleo vestibular lateral, cerebelo, etc., cujos
influxos atingem os músculos nas fibras extrafusais, seja diretamente pelos motoneurônios α,
seja indiretamente pelo sistema γ. Os reflexos tônicos cervicais consistem de reações de
músculos apendiculares desencadeadas pela movimentação passiva da cabeça. Os corpúsculos
28
de Vater-Pacini situados nos ligamentos das articulações cervicais, particularmente na atlantooccipital. Na rotação da cabeça para um lado provoca a extensão do membro superior ipsilateral
e a flexão contralateral (postura do esgrimista em guarda). A anteflexão passiva da cabeça
determina a flexão dos membros superiores e a extensão dos inferiores. A retroflexão passiva da
cabeça determina extensão dos membros superiores e flexão dos membros inferiores. Os
reflexos labirínticos são desencadeados pela estimulação do ouvido interno resultando
movimentação óculocefalica e do equilíbrio. Os influxos são transmitidos aos neurônios motores
periféricos através do FLM e do trato vestíbuloespinhal, estabelecendo conexões entre os
núcleos vestibulares, os nervos motores oculares, o núcleo espinhal do acessório e as colunas
cinzentas ventrais da medula cervical; o trato vestíbuloespinhal está correlacionado com o tono
dos músculos extensores do pescoço e dos membros inferiores, e músculos antigravitários das
costas.
Os reflexos exteroceptivos são polissinápticos com maior período de latência, maior
fatigabilidade, necessidade de somação de estímulos e falta de resposta a autopesquisa. Entre os
reflexos cutâneos que se admite sejam normalmente ativados pelo sistema piramidal: os
cutâneoabdominais superior, médio e inferior realizam a contração ipsilaterais dos músculos
abdominais, os quais são inervados pelas raízes de T6-T9 para os superiores, T9-T11 para os
médios e T11-T12 para os inferiores; o cremastérico realiza a contração do músculo cremastérico
na dependência do músculo oblíquo interno do abdome, inervado pelo ilíohipogastrico e
ilíoinguinal e raízes de L1L2; e os reflexos cutâneoplantares em flexão (veja abaixo sinal de
Babinski). A perda transitória ou permanente dos reflexos superficiais cutâneoabdominais,
cremastéricos e cutâneoplantares ocorre pela retirada da inibição ortopiramidal durante o
período de choque do sistema piramidal. Os reflexos visceroceptivos são estudados com o
sistema nervoso autônomo.
Os reflexos exteroceptivos são polissinápticos com maior período de latência, maior
fatigabilidade, necessidade de somação de estímulos e falta de resposta a autopesquisa. A perda
dos reflexos cutâneoabdominais, cremastéricos, cutâneoplantares, e o aparecimento do reflexo
palmomentual acontecem durante o período de choque por lesão ortopiramidal, causando perda
da facilitação do sistema piramidal. Os reflexos abdominais e cremastéricos estão presentes no
29
nascimento, embora com uma área reflexógena maior; no adulto, a estimulação exteroceptiva
do abdômen provoca contração da musculatura abdominal e relaxamento recíproco da
musculatura do tórax. Acredita-se que esses reflexos sejam reflexos de defesa, integrados em
níveis espinhal via internunciais e facilitados pelo trato piramidal. Apesar dos estímulos
piramidais produzirem um potencial excitatório pós-sináptico nos neurônios internunciais
relacionados com esses reflexos, não há evidencia de que a abolição desses reflexos esteja
diretamente dependente de lesão do trato piramidal. A estimulação cutânea rápida, em
condições favoráveis da parede abdominal no sentido láteromedial à altura das regiões
epigástricas, obtém-se os reflexos: abdominais superiores – raízes de T6-T9; na região umbilical,
abdominais médios – raízes de T9-T11; e na região hipogástrica, e os abdominais inferiores –
raízes de T11-T12. Ocorre a contração ipsilateral dos músculos reto do abdome, oblíquo externo,
oblíquo interno e transverso, com desvio da linha alba e da cicatriz umbilical no sentido
médiolateral.
O reflexo cremastérico superficial aparece pela estimulação cutânea do terço superior da face
medial da coxa e, o reflexo cremastérico profundo, pela pressão digital sobre a face medial da
musculatura da coxa, entre o terço superior e médio. A contração do músculo cremaster está na
dependência do músculo oblíquo interno do abdome com elevação do testículo ipsilateral ou
retração do grande lábio ipsilateral. Em caso do reflexo cremastérico exaltado, pode ser
percebido contralateral. Sua inervação é feita pelos nervos iliohipogástrico e ilioinguinal com
nível medular em L1-L2. Esse reflexo não deve ser confundido com o reflexo dartóico ou escrotal,
que consiste em leve corrugamento da bolsa escrotal, devido ao a contração do darto um
músculo liso de inervação simpática, e provocado por excitação cutânea do escroto, períneo ou
coxa, como também por ação do frio. Nas lesões cordonais posteriores aparece o sinal de Tolosa
devido à dissociação entre os dois reflexos cremastéricos: presença do superficial e abolição do
profundo. Os reflexos cutâneoplantares em flexão (veja abaixo sinal de Babinski).
O reflexo palmomentoniano é possuidor de um arco reflexo multissegmentar inibido pelos
sistemas motores superiores, tendo fisiopatologia similar aos reflexos axiais da face. Pesquisa-se
pela estimulação, principalmente da eminência tênar; pode está presente em indivíduos
normais, por contração ipsilateral dos músculos mentual e orbicular, com ligeira elevação do
lábio inferior; mostra-se vivo nas lesões piramidais bilaterais, principalmente corticonuclear, e
30
nas síndromes paleoestriadas; e mostra-se abolido na paralisia facial central ou periférica; sua
interpretação depende do contexto sindrômico. Entretanto, existem duas proposições: A. O
reflexo palmomentual normal segue tais características – 1. Zona reflexógena limitada à
eminência tênar; 2. Resposta débil e de curta duração; 3. Extinção da resposta entre 2 a 4
estimulações; e 5. Variações de um a outro momento. B. O reflexo palmomentual patológico se
caracteriza por - 1. Zona reflexógena que ultrapassa a eminência tênar; 2. Resposta de média a
intensa; 3. Sem habituação após 2 a 4 estimulações; 4. Constancia da resposta entre um a outro
momento. Os reflexos visceroceptivos são estudados com o sistema nervoso autônomo.
Sintomas de liberação da síndrome piramidal de Babinski
As lesões piramidais condicionam uma desintegração complexa da função motora, que
associadas ao déficit motor realizam modificações qualitativas, por sua conta, do campo de ação
motor onde ocorrem os movimentos associados anormais. Portanto, a exaltação do sistema
extrapiramidal tende a predominar pela falta de inibição do sistema piramidal e origina a
síndrome de liberação extrapiramidal. Essa síndrome é constituída por dois componentes: por
lesão da via ortopiramidal e por lesão da via justapiramidal. Na lesão da via ortopiramidal,
aparece liberação do sistema extrapiramidal com sincinesias globais e de coordenação; e a
liberação dos sistemas polissinápticos nociceptivos da medula decorrentes da liberação pura do
sistema ortopiramidal com sinal de Babinski e automatismo medular. Nas lesões da via
justapiramidal por liberação do sistema vestibuloespinhal aparecem sinais de espasticidade; e na
liberação do sistema reticuloespinhal facilitador ocorre hiperreflexia e clonus.
Sincinesias ocorrem quando os membros contraturados, mais ou menos incapazes de
movimentar-se pela vontade, podem apresentar movimentos involuntários diante de
movimentos voluntários ou passivos do lado oposto. Por imaturidade do sistema piramidal as
crianças realizam movimentos similares as sincinesias com a mão esquerda, face, língua quando
aprendem a escrever. Posteriormente desaparecem. Entretanto, algumas pessoas os mantêm
quando realizam movimentos mais finos. As sincinesias patológicas podem ser classificadas em:
A sincinesia global caracteriza-se pela contração global dos músculos do lado paralisado com
consequente contratura observada nos hemiplégicos (flexão da extremidade superior e extensão
31
do inferior). Pode ser obtida quando: a) quando se reforça a contração do esforço, o fechamento
com força da mão do lado sadio; b) no hemiplégico a sincinesia global é influenciada pelo reflexo
tônica cervical, ao girar a cabeça para o lado normal aumenta o tônus flexor e diminui ao girar a
cabeça para o lado doente por aumento do tônus extensor; c) durante a marcha na hemiplegia
piramidal clássica, ao abduzir a extremidade inferior, o paciente flexiona o joelho contra a
resistência, não pode fazê-lo sem ao mesmo tempo realizar uma flexão dorsal com adução do pé
(fenômeno de Strümpell ou do tibial anterior); d) durante movimentos automáticos como
espirro, tosse, riso, etc.
Sincinesias de coordenação é a que melhor expressa à situação condicionada pelo transtorno
piramidal. A sincinesia pode ser homolateral e heterolateral ao déficit: a) A sincinesia
homolateral no tibial anterior (Strümpell) ocorre quando o paciente com lesão piramidal tenta
fletir a perna sobre a coxa e a coxa sobre a pelve, aparece dorsoflexão e inversão do pé. O sinal
dos interósseos de Souques ocorre quando da elevação voluntária do membro superior lesado
acompanha-se de hiperextensão e abdução dos dedos da mão paralisada. b) A sincinesia
heterolateral realizada pela prova de Raimiste na abdução associada. Coloca-se o paciente em
decúbito dorsal com os membros inferiores em extensão, tenta-se abduzir o membro normal
contra a resistência e o membro lesado abduz por associação. O mesmo principio é obtido ao
fazer a adução dos membros inferiores.
O sinal de Babinski (SB) descrito em 1896 e os reflexos de defesa chamados de automatismo
medular têm uma unidade funcional e fisiopatológica. O SB é a flexão dorsal ou extensão do
hálux produzida por estimulação do tegumento de qualquer que seja a superfície plantar que
não seja a do hálux. O sinal de Babinski em forma de leque, descrito em 1903, ocorre em lesão
predominante na área prémotora, pesquisado similar ao tradicional. Mas, é bem aceita a
estimulação de trás para frente, roçando a parte externa do pé e a base dos dedos, sem feri-lo. O
SB é o primeiro sinal de liberação a aparecer na lesão piramidal, podendo ser seguido pelos
reflexos de defesa. Em certos casos de lesão piramidal pode aparecer o SB da mão, que consiste
no movimento de flexão, oposição e adução do polegar ao ser estimulada a eminência hipotenar.
A margem cubital no adulto, apesar de medial, corresponde à lateral do pé no processo de
ontogênese. Em casos raros, o SB pode ser obtido pela estimulação do território do nervo
32
trigêmio com resposta extensora do hálux contralateral. Mas, interpretações errôneas do SB
existem. O pseudosinal de Babinski ou Babinski periférico pode ocorrer em casos de neuropatia
periférica, poliomielite com atrofia dos extensores curtos dos artelhos, principalmente o flexor
curto, adutor e abdutor do hálux. Como os músculos extensores estão íntegros pode haver
resposta extensora da primeira falange do hálux. Mas, o SB não traduz necessariamente a
existência de uma lesão anatômica piramidal, também pode existir por distúrbio funcional no
sono profundo, intoxicação por barbitúricos, etc.
A extensão do hálux não é mais do que o componente mais distal da resposta sinérgica
nociceptiva flexora da extremidade inferior, como visto no reflexo medular cruzado. Portanto, a
flexão dorsal reflexa do pé estimulada pela planta do pé não é uma resposta reflexa patológica.
Normalmente, a cronaxia sensitiva é igual à dos flexores, mas é o dobro da dos extensores. A via
sensitiva contém fibras finas do tipo A e do tipo C, sendo as últimas de condução mais lenta.
Portanto, o reflexo cutâneoplantar tem dois componentes, um mais rápido e outro mais lento.
Na resposta motora, primeiro aparece com a flexão do bíceps crural e depois com a extensão do
tibial anterior, respectivamente. No caso de lesão piramidal a cronaxia sensitiva não se altera,
mas as motoras se invertem, de forma que a dos flexores se divide e a dos extensores se duplica.
Como os influxos nervosos caminham por vias isócronas, a excitação plantar provocará não mais
flexão e, sim, extensão: primeiro o tibial anterior e extensor longo do hálux, depois o bíceps. A
retirada da inibição tônica desses músculos pelo sistema piramidal é responsável pela inversão
funcional desses músculos.
Resumo do sinal de Joseph Francois Felix Babinski - A área reflexógena para o reflexo
cutâneoplantar é a raiz de S1 com as terminações nervosas alcançando a pele. O nervo aferente
é o tibial anterior, os segmentos medulares envolvidos no arco reflexo vão de L4 até S2. Para
pesquisá-lo, deve-se dizer ao paciente que a planta do seu pé vai ser estimulada, ele deve ficar
com a perna relaxada sobre a cama com leve flexão na dobra do joelho e o pé em discreta
eversão. Utiliza-se um estilete de ponta rombuda e pressiona-se qualquer área da planta do pé
menos o próprio hálux e, num tempo máximo de 5 segundos, realiza-se o movimento uniforme;
não se deve ferir a pele do pé ou repetir o procedimento logo em seguida. O verdadeiro sinal de
Babinski ocorre pela extensão do hálux decorrente do recrutamento de todos os componentes
33
envolvidos com o músculo longo extensor do hálux. A resposta fisiológica do reflexo
cutâneoplantar ocorre quando indivíduos sentem uma sensação desagradável durante a
estimulação ou são muito sensíveis ao estímulo; esse fenômeno já tinha sido demonstrado antes
de Babinski, por Leonardo da Vinci e Raphael, nas suas clássicas pinturas. O sinal de Babinski
pode existir por lesão fisiológica ou anatômica do feixe ortopiramidal. Quando a via
justapiramidal está envolvida separa da ortopiramidal a resposta em extensão não tem o mesmo
significado. Quando ambas as vias estão lesadas há extensão rápida de todos os dedos do pé;
quando na via ortopiramidal, apenas do hálux faz extensão; e na lesão da via justapiramidal a
extensão é em leque e mais devagar. A extensão do hálux só é patológica se causada pela
contração do músculo extensor do hálux; a contração do músculo extensor do hálux é patológica
somente se ele ocorrer sincronicamente com atividade reflexa em outros músculos flexores; o
sinal de Babinski não implica necessariamente que a atividade concorrente dos outros músculos
flexores seja muito viva ou vice-versa.
Com relação aos reflexos de defesa ou de automatismo medular precisa-se ser rigoroso na
diferenciação entre os autênticos reflexos de defesa, que tem uma significação de disfunção
piramidal com liberação da atividade nociceptiva espinhal, dos reflexos de reação de defesa. O
sinal patológico é a flexão dorsal reflexa do pé produzida por estimulação fora da superfície
plantar; sempre hiperexcitável e estereotipado. Um beliscão no dorso do pé pode desencadear
uma resposta em tríplice flexão (do pé, da perna e da coxa), principalmente nas lesões
medulares. Enquanto, os reflexos de reação de defesa ocorrem por hiperestesia, excitação
psicomotora, instabilidade emocional, redução do controle de vigília, etc; poder ser excitáveis ou
hiperexcitáveis, mas a resposta não é estereotipada.
A espasticidade é uma hipertonia essencialmente constituída por um excesso de atividades
reflexas que utilizam o arco reflexo. É decorrente da liberação do feixe vestibuloespinhal pela
lesão do sistema justapiramidal que deixa de inibir os motoneurônios do sistema γ dinâmico,
tendo reflexos vivos ou exaltados. Na espasticidade existe um desequilíbrio entre as influências
facilitadoras e inibidoras supraespinhais que chegam aos motoneurônios γ, desequilibra o sinal
positivo através do circuito periférico, determina a potenciação présinaptica dos motoneurônios
α tônico. A espasticidade apresenta-se diferente em topografias distintas. Nas lesões corticais e
34
capsulares não existe a quebra do princípio da inervação recíproca; nas lesões de tronco
encefálico há redução da inibição recíproca com atividade aumentada do antagonista, tanto
durante a tração como ao longo da sua manutenção; e nas lesões da medula espinhal não existe
inervação recíproca, em virtude da descarga prolongada e irradiada, deixando de ser
espasticidade. Nas lesões corticais, capsulares e de tronco cerebral da via piramidal aparece o
sinal de Babinski simples ou em leque, a depender da área cortical lesada, área 4 ou área 6 de
Brodmann, respectivamente; nas lesões da medula espinhal além do sinal de Babinski, aparecem
os reflexos de defesa em tríplice flexão. Na síndrome piramidal pura a hipertonia é seletiva,
atingindo os músculos flexores dos membros superiores e os extensores dos músculos inferiores.
A hipertonia é elástica e liberta o músculo passivamente distendido. Nas lesões que predominam
a via ortopiramidal a presença do sinal do canivete pode ser mais evidente do que quando
predomina os sinais de lesão da via justapiramidal.
A espasticidade do sistema γ dinâmico é diferente da rigidez devido à falta de inibição do sistema
γ tônico, com o aparecimento dos reflexos tônicos exaltados. O reflexo tônico do músculo tibial
anterior encontrado nos parkinsonianos é um dos exemplos de reflexo tônico de postura. O
bloqueio seletivo das fibras finas, que respeitam tanto as fibras eferentes α como as aferentes Ia,
produz alguma diminuição transitória do reflexo tônico exagerado. Enquanto a rigidez do sistema
α pode ser vista principalmente nas lesões do vermis anterior ou paleocerebelo e, não
desaparece com secção das raízes dorsais, como desaparece na espasticidade do sistema γ
dinâmico.
Do ponto de vista semiológico, a espasticidade é diferente da rigidez. A hipertonia para
movimentos passivos difere da rigidez por não ser uniforme aos vários tipos de movimentos e
variar com a velocidade do movimento. Além do mais, a rigidez tende a afetar todos os músculos
no mesmo grau, enquanto a hipertonia varia de um grupo de músculos para outro. Na
espasticidade, se o movimento for realizado lentamente, pode existir pouca resistência. Mas, se
o movimento for rápido, haverá aumento rápido da resistência. Com o cotovelo em 90 graus e o
antebraço pronado o médico supina lentamente a mão do paciente, a menos que a espasticidade
seja severa, nesse caso haverá clonus do músculo pronador. Na hemiparesia ou hemiplegia, a
espasticidade é mais marcada nos músculos flexores e pronadores da extremidade superior e
35
extensores da inferior. A postura apresenta-se com flexão do membro superior, tendo flexão do
antebraço sobre o braço, do punho e dos dedos da mão; no membro inferior haverá extensão
completa, podendo haver espasmo dos músculos adutores do quadril. Existe mais resistência
passiva no movimento de extensão do que no de flexão na extremidade superior e, na
extremidade inferior, mais resistência passiva no movimento de flexão do que no de extensão.
Os reflexos miotáticos fásicos são inibidos pelo sistema justapiramidal. Mas, a sua lesão libera o
sistema reticuloespinhal ventral com a expressão dos reflexos vivos e exaltados; o clonus do pé
pela distensão do tendão do tríceps sural; da rótula pelo estiramento do tendão do quadríceps;
da mão e dedos pela extensão passiva do punho ou dedos; e da mandíbula pelo estiramento do
masseter. Clonus é uma serie de contrações clônicas e rítmicas, involuntárias, do respectivo
músculo, cuja duração é variável, subordinada ao tempo em que se mantém a distensão
provocada passivamente pela distensão brusca de um tendão. Fazem parte desse cortejo, as
“trepidações epileptóides”, que consistem em abalos clônicos repetidos de todo um membro. As
trepidações aparecem ao se pesquisar determinado reflexo ou durante um esforço voluntário.
Esses sinais são devido a um marcapasso dos reflexos, neles intervêm os estímulos originados
antagonistas pelos sucessivos estiramentos que sofre cada contração do agonista.
Resumo da síndrome piramidal
Lesão na área motora primária ou área 4 de Brodmann dificulta a realização de movimentos
finos, sem espasticidade, podendo haver hipotonia e hiporreflexia, mas está presente o sinal de
Babinski. Lesão na área 4s de Hines, limite entre as áreas 4 e 6 de Brodmann, de onde sai o feixe
de Türck, provoca espasticidade principalmente nos segmentos axiais. Lesão em parte da área
prémotora 6 de Brodmann produz fraqueza contralateral e dificuldade na realização de
movimentos finos, aumento da resistência à movimentação passiva, sem características
espástica, e a presença de grasping. Lesão na área motora suplementar área 6 de Brodmann leva
a aumento do tônus passivo, sem características espástica e sem sinal de Babinski. Pode haver
moderada e transitória hiperreflexia e fraqueza muscular que desaparece em grande parte,
persistindo apenas para movimentos finos. Presença do reflexo de grasping ou preensão forçada,
além do sinal de Babinski em leque. Em qualquer nível do neuroeixo, do córtex até a medula
36
espinhal, uma lesão acarretará uma sintomatologia que varia em grau, mas similar ao que foi
descrito acima.
MANIFESTAÇÕES CLÍNICAS DE LESÕES NO SISTEMA PIRAMIDAL
A via do TCE pode ser envolvida em uma série de processos mórbidos, incluindo doença vascular,
neoplasia, degeneração, trauma, e outros. Dentre esses, as doenças vasculares são as mais
comuns. A hipertonia e a hiperreflexia observadas em lesões de diferentes níveis do SNC têm
muitas divergências entre si. Por exemplo, a hipertonia das lesões medulares se acompanha de
reflexos nociceptivos flexores; a atitude de Wenicke-Mann é observada nas hemiplegias por
lesão da capsula interna; nas lesões da coroa radiada a hemiparesia é desproporcionada e a
hipertonia é menos intensa. A atitude hemiplégica é dinâmica, dependendo a distribuição do
tono de reflexos labirínticos e cervicais, pois se modifica conforme o paciente é colocado em
posição bípede, quadrúpede ou em decúbito lateral.
1. Síndromes envolvendo as fibras piramidais no córtex cerebral e centro semioval:
a) Síndrome motora cortical por predomínio de lesão na área 4 de Brodmann, ocorre por lesão
piramidal com fraqueza focal ou segmentar e hipotonia. As paralisias corticais são parciais e
incompletas. Quando lesa o opérculo rolândico a paralisia afeta os músculos da metade inferior
do rosto; quando lesa a parte média o membro superior; e quando lesa o lóbulo paracentral
afeta o membro inferior.
b) Síndrome cortical por predomínio de lesão na área 6 de Brodmann, o transtorno motor não é
do tipo deficitário, mais próxima da área 4 mostra lesão piramidal com fraqueza frustra,
espasticidade e sinal de Babinski em leque. Lesão na região prémotora, área 6 de Brodmann, há
incoordenação chamada ataxia frontal de Bruns (1892). A síndrome é formada por lesão
piramidal frustra e incoordenação contralateral. Ao deambular para frente à láteropulsão ocorre
para o lado da incoordenação e, andando para trás, para o lado da lesão. Esse detalhe faz o
diagnóstico diferencial com a incoordenação de origem cerebelar.
c) A área 6 de Brodmann, especialmente a área 6aβ provoca uma variedade de mal epiléptico.
Quando o fenômeno inicia-se pela área 6b, consistem de mastigação, salivação, deglutição e
37
sensação constritiva na região da faringe; às vezes aparece gagueira, desejo de falar sem poder,
etc.
d) A área 6 de Brodmann quando lesada, pode apresentar a síndrome acinética hipertônica ou
síndrome pseudopalidal. Nessa síndrome a lesão é puramente cortical e pode também combinarse com fenômeno de catalepsia relacionada com hipertonia extrapiramidal. Existe disartria e
amimia, quando a lesão é esquerda, existe afasia tipo Broca e agrafia. O diagnóstico diferencial
pode ser feito com as síndromes extrapiramidais de origem nos gânglios da base, porque a
síndrome descrita acima é do tipo piramidal e compromete apenas o córtex cerebral prémotor.
e) Síndromes subcorticais e no centro semioval ocorrem por lesão nas fibras piramidais, dando
fraqueza e espasticidade no dimídio contralateral à lesão. São desproporcionadas de predomínio
facial, braquial ou crural. Lesão no ramo superficial da artéria cerebral média é responsável por
hemiplegia de predomínio faciobraquial com prejuízo da linguagem quando à esquerda. O
mecanismo fisiopatogênico mais comum nessas síndromes é o de embolia cerebral, advinda do
coração, principalmente em casos de aneurisma de ponta do ventrículo esquerdo após enfarte
do miocárdio, ou estenose da valva mitral como sequela de febre reumática. Lesão na artéria de
Heubner, ramo da artéria cerebral anterior, é responsável por hemiplegia de predomínio crural
sem afetar a linguagem. Essa síndrome pode estar relacionada com trombose sifilítica.
2. Síndromes envolvendo as fibras piramidais que passam na cápsula interna: nessa topografia
a característica semiológica é a de que o déficit motor é completo e a área lesada é pequena.
a) Síndrome do andar superior acontece por lesão dos ramos profundos da artéria cerebral
média, que afeta as radiações talâmicas e as fibras piramidais. No quadro clínico aparece
hemianestesia e hemiplegia contralateral à lesão.
b) Síndrome da porção anterior do andar inferior ocorre por lesão da artéria coroidéa anterior,
apresentando uma síndrome extrapiramidal.
c) Síndrome porção pósteroventral do andar inferior acontece por lesão da artéria coroidéa
posterior, mostrando hemiplegia pura com espasticidade adquirindo postura de Wernicke Mann.
38
d) Síndrome da porção pósterodorsal do andar inferior ocorre por lesão da artéria coroidéa
anterior que compromete o trato geniculocalcarino com hemiplegia e hemianopsia.
e) Síndrome talâmica de Déjerinè-Roussy acontece por lesão dos ramos artéria cerebral
posterior, acometendo o pedúnculo talâmicogeniculado. O quadro clínico é formado por
hemianestesia, principalmente profunda, dor espontânea (anestesia dolorosa), desproporção
entre intensidade de estímulo e sensação, tonalidade afetiva nas emoções ou hiperpatia, sinais
cerebelares e movimentos coreicos e atetósicos, semiflexão das primeiras falanges e extensão
das demais, mão talâmica.
3) Síndromes mesencefálicas: a irrigação do mesencéfalo depende de ramos da artéria cerebral
posterior, formada da subdivisão da artéria basilar, emite uma série de artérias colaterais que se
destinam sobretudo ao mesencéfalo e são constituídas pelas artérias interpendiculares, as quais
atravessam o espaço perfurado posterior e irrigam a substancia negra; pelas artérias
pedunculares externas, que se aprofundam no pedúnculo; pela artéria média do tubérculos
quadrigêmios; pelas artérias do corpo geniculado; e pela artéria anterior
dos tubérculos
quadrigêmios. As síndromes clínicas do mesencéfalo são clássicas.
a) Síndrome do topo da basilar caracteriza-se por apresentar hemiparesia contralateral à lesão,
cegueira cortical, déficit do óculomotor, dificuldade com a memória, e déficit sensitivo
contralateral. A lesão acomete os lobos occipitais, núcleos do óculomotor, pedúnculo cerebral,
porção medial do lobo temporal e tálamo.
b) Síndrome de Weber depende de isquemia da porção ventral da base do pedúnculo cerebral,
afetando as fibras piramidais e as fibras radiculares do III nervo que transitam nesse segmento. O
quadro clínico caracteriza-se no lado da lesão por oftalmoplegia homolateral do III nervo e
contralateral à lesão hemiplegia, afetando o facial central. O prejuízo do III nervo poder ser
incompleto, assim como o grau da hemiplegia. Lesões nessa área são geralmente causadas por
enfartes lacunares em hipertensos devido à oclusão das pequenas artérias paramedianas.
c) Síndrome de Benedikt ocorre por lesão do paleorubro, porção magnocelular. O quadro clínico
caracteriza-se no lado da lesão por comprometimento do III nervo e, contralateral, por síndrome
hipercinética e hipertônica, associada à rigidez. As hipercinesias são tremores cerebelares ou do
39
tipo parkinsoniano ou movimentos coreoatetósicos. Eventualmente, existem alterações de todos
os tipos de sensibilidade.
d) Síndrome de Claude depende da obstrução de uma das arteríolas medianas que se dirigem
para o núcleo do III nervo, atravessa o pedúnculo cerebelar superior, fornecendo um ramúsculo
à base do pedúnculo e lesa a porção parvocelular do núcleo rubro de Stilling. No quadro clínico
há no lado da lesão oftalmoplegia completa e, contralateral, manifestações cerebelares pela
lesão do pedúnculo cerebelar superior sobre o núcleo rubro.
e) Síndrome de Chiray, Foix e Nicolesco ou síndrome peduncular da calota ocorre por lesão da
parte mais superior do núcleo rubro, parvocelular, sem prejudicar as fibras do III nervo. No
quadro clínico existe assinergia, tremores intencionais, alterações sensitivas e hipotonia no
hemicorpo oposto à lesão. Ás vezes existe miose homolateral.
f) Síndrome de Fovile peduncular depende de lesão na porção medial da base do pedúnculo
cerebral e consiste de hemiplegia contralateral à lesão e desvio da cabeça e dos olhos para o lado
da lesão. Pode ocorrer paralisia facial central no mesmo lado da hemiplegia, pelo
comprometimento do feixe aberrante peduncular profundo de Déjerine. O paciente não
contempla a sua hemiplegia.
g) Síndrome de Raymond-Cestan caracteriza-se no lado da lesão do pedúnculo cerebelar por
hemisíndrome cerebelar associada a tremor, ataxia e assinergia e hemiplegia contralateral.
h) Síndrome de Gelle ou acústica alterna apresenta hipoacusia, paralisia facial homolateral à
lesão e hemiplegia contralateral.
4. Síndromes protuberanciais: a irrigação da protuberância é formada por três grupos: ramos
paramedianos, ramos circunferenciais curtos e longos. As artérias paramedianas, em numero de
3 a 5, penetram na ponte muito próxima à linha mediana e irrigam os tratos corticoespinhal e
corticobulbar, os núcleos pontinos, as fibras ponto cerebelares, parte do leminisco medial e
raízes do VI nervo. As artérias circunferenciais curtas irrigam alguns núcleos pontinos, os
segmentos mais laterais do trato corticoespinhal e do leminisco lateral, os núcleos do V e VII
nervos, e o pedúnculo cerebelar médio. As artérias circunferenciais longas são representadas por
dois vasos: artéria cerebelar superior e artéria cerebelar anterior inferior que irrigam o trato
40
espinotalâmico lateral, o núcleo espinhal do V nervo e o trato espinhal do trigêmio, os núcleos
cocleares e do facial, os feixes espinocerebelares e o pedúnculo cerebelar superior.
a) Síndrome protuberancial paramediana ocasiona hemiplegia e\ou hemianestesia contralateral.
Em caso de lesão bilateral, pode ocorrer apoplexia por hemorragia da porção paramediana da
ponte com diplegia ou síndrome pseudobulbar.
b) Síndrome do encarcerado ou loked-in syndrome ocorre por trombose da artéria basilar na
porção anterior da ponte. O quadro clínico é formado por tetraplegia, afonia, paralisia do olhar
horizontal.
c)
Dysarthria-Clumsy hand
syndrome
ocorre
por
lesão
no
trato
corticobulbar e
corticopontocerebelar devido a enfarte lacunar nos dois terços inferiores da ponte similar ao
quadro clínico que pode ocorrer no joelho da cápsula interna. O quadro clínico é formado por
disartria e inquietação da mão.
d) Síndrome lateral da ponte corresponde à oclusão das artérias circunferenciais curtas e se
traduz por: síndrome cerebelar ipsilateral, eventualmente com hemiplegia contralateral. A
depender da extensão da lesão pode haver alterações dissociadas de sensibilidade no mesmo
lado da hemiplegia, além de sinais de nervos cranianos da ponte.
e) Síndrome de Foville superior é ocasionada por lesão da porção superior da ponte devido ao
comprometimento do feixe aberrante pontino de Déjerine. No quadro clínico há hemiparesia
contralateral à lesão e paralisia conjugada do olhar horizontal para o lado contrário ao da lesão; a
cabeça e os olhos costumam desviar para a hemiparesia. O paciente contempla a sua hemiplegia.
f) Síndrome de Foville inferior acontece por lesão na porção caudal da ponte. No quadro clínico
ocorre hemiplegia alterna com paralisia facial periférica contralateral, hemianestesia no lado
hemiplégico, e desvio da cabeça e dos olhos para o lado contrário à paralisia facial periférica. O
paciente contempla sua hemiplegia parcialmente.
g) Síndrome de Millard–Gubler ocorre na porção caudal do tegmento da ponte. No quadro
clínico homolateral a lesão há paralisia facial periférica e, às vezes, estrabismo convergente, e
hemiparesia contralateral à lesão.
41
5. Síndromes bulbares: uma forma simplista de mostrar a irrigação do bulbo é dizer que a artéria
vertebral fornece quatro ramos. a) ramo meningeo se destina as meninges do rombencéfalo; b)
artéria espinhal posterior (não caracteriza síndrome neurológica conhecida) que se destaca da
artéria cerebelar posteroinferior; c) artérias espinhais anteriores (juntam-se para formar uma, e
desce pela fissura anterior da medula), irrigam as pirâmides bulbares, os leminiscos mediais e, às
vezes, fibras do hipoglosso e porções ventrolaterais dos núcleos olivar; d) artéria cerebelar
pósteroinferior irriga a fosseta lateral do bulbo e hemisfério cerebelar.
a) Síndrome da artéria espinhal anterior (1908) manifesta-se com dormência e redução de força
no dimídio contralateral à lesão. No exame há sinais piramidais e distúrbio da sensibilidade
profunda. Pode haver língua com fibrilações e nistagmo por lesão do FLM posterior no lado da
lesão.
b) Síndrome de Wallenberg na maior parte das vezes ocorre por lesão na artéria vertebral e, mais
raro, na artéria cerebelar pósteroinferior. A região da fosseta lateral é uma área cuneiforme com
o ápice voltado pósteromedial. Essa região abrange os tratos espinocerebelares ventral e dorsal,
trato espinotalâmico lateral e o núcleo do vago e do glossofaríngeo (porção superior do núcleo
ambíguo), parte das formações reticulares, inclusive as vias simpáticas, a raiz descendente do
trigêmio, os dois terços anteriores da substância gelatinosa de Rolando, e o pólo superior da
oliva bulbar. A síndrome completa do lado da lesão existe hemissíndrome cerebelar, paralisia
velofaringea, síndrome oculosimpática de Claude Bernard-Horner, e alteração da sensibilidade
térmica e dolorosa na hemiface: do lado oposto da lesão existe anestesia dos membros e tronco,
com dissociação siringomiélica.
c) Síndrome de Babinski-Nageotte ou síndrome d hemibulbo contem quatro focos de lesão
vasculares. O quadro clínico do lado oposto à lesão apresenta hemiplegia poupando a face,
hemianestesia dos membros e troncos; do mesmo lado da lesão hemissíndrome cerebelar,
paralisia velofaringea, síndrome de Claude Bernard-Horner e alterações de sensibilidade na
hemiface.
42
d) Síndrome de Avellis ou síndrome do tegumento
bulbar. É mais comum em idosos por trombose da
artéria vertebral, comprometimento do território do X
nervo, trato espinotalâmico lateral e núcleo ambíguo;
ocasionalmente envolve o leminisco medial. Os achados
clínicos homolaterais à lesão são fraqueza da corda
vocal e do palato, pode haver síndrome de Horner;
contralateral à lesão há alteração da hemisensibilidade
Fig.17
térmica, dolorosa, e pode haver hemiparesia alteração
do tato e propriocepção profunda.
e) Síndrome de Goukowsky ocorre devido à isquemia na
distribuição artéria espinhal anterior. Compromete as
raízes do XII nervo e a pirâmide bulbar. O quadro clínico
apresenta paralisia da hemilíngua homolateral à lesão e, contralateral, hemiparesia poupando a
face.
f) Síndrome da decussação piramidal lateral ocorre por
lesão na porção lateral da pirâmide bulbar. Evidencia-se
por monoplegia no membro superior homolateral à lesão
e monoplegia no membro inferior contralateral à lesão
(Fig. 17 & 18).
g) Síndrome da decussação medial e posterior ocorre por
lesão na porção central e posterior do bulbo na região da
pirâmide bulbar. No exame há paraplegia braquial, veja a
fig. 18.
i) Síndrome de Schneider ocorre por lesão no nível do
segmento cervical inferior na metade anterior da medula
Fig.18
por oclusão da artéria espinhal anterior. O quadro clínico
é de tetraplegia, sendo flácida nos membros superiores e
espástica nos membros inferiores. Existe dor no pescoço, dormência nas mãos.
43
LEITURAS RECOMENDADAS
1. BARRAQUER-BORDAS, L. NEUROLOGÍA FUNDAMENTAL. 3ª EDIÇÃO, BARCELONA, TORAY, 1976, 1144.
2. BRODAL, A. ANATOMIA NEUROLÓGICA COM CORRELAÇOES CLÍNICAS. 3ª EDIÇÃO, SÃO PAULO, ROCA,
1984, p.888.
3. CAMPBELL, W.W. DeJONG: THE NEUROLOGIC EXAMINATION. SIXTH EDITION, PHILADELPHIA,
LIPPINCOTT WILLIAMS & WILKINS, 2005, p.671.
4. CANELAS, H.M; ASSIS, J.L; E SCAFF, M. FISIOPATOLOGIA DO SISTEMA NERVOSO. SÃO PAULO, SARVIER,
1983, p.476.
5. CARPENTER, M.B. FUNDAMENTOS DE NEUROANATOMIA. QUARTA EDIÇÃO, MARYLAND,
PANAMERICANA, 1999, p. 458.
6. CASAS, A.P & GONZÁLEZ, M.E.B. MORFOLOGÍA, ESTRUTURA Y FUNCIÓN DE LOS CENTROS NERVIOSOS.
TERCERA EDICIÓN, MADRID, EDITORIAL PAZ MONTALVO, 1977, p. 901.
7. GOETZ, C.G. TEXTBOOK OF CLINICAL NEUROLOGY. THIRD EDITION, PHILADELPHIA, SAUNDERS
ELSEVIER, 2007, p.1364.
8. CINGOLANI, H.E; HOUSSAY, A. B. FISIOLOGIA HUMANA. SÉTIMA EDIÇÃO, SÃO PAULO, ARTMED
EDITORA, 2004, p.1124.
9. KUMAR, S.P; RAMASUBRAMANIAN. THE BABINSKI SIGN – A REAPPRAISAL. NEUROL INDIA, 2000;
48:314-318.
10. LENT, R. CEM BILHÕES DE NEURÔNIOS: CONCEITOS FUNDAMENTAIS DE NEUROCIÊNCIAS. SÃO
PAULO, EDITORA ATHENEU, 2005, p. 698.
11. PEDRO-PONS, A. ENFERMEDADES DEL SISTEMA NERVIOSO, NEUROSIS Y MEDICINA PSICOSOMATICA
ENFERMEDADES MENTALES. TOMO IV, SALVAT, BARCELONA, 1973, p.1191.
12. PUVANENDRAN, K; WONG, PK; RANSOME, GA. SYNDROME OF DÉJERINE’S FOURTH REICH. ACTA
NEUROL SCAND. 1978;57:349-353.
13. RANSON, S.W & CLARK, S.L. ANATOMIA DEL SISTEMA NERVIOSO. DECIMA EDICIÓN, MEXICO,
EDITORIAL PANAMERICANA, 1963, p.573.
14. ROPPER, A.H and BROWN, R.H. ADAMS and VICTOR’S: PRINCIPLES OF NEUROLOGY. EIGHTH EDITION,
NEW YORK, McGRAW HILL, 2005, p.1382.
15. SURÓS, J. SEMIOLOGIA MÉDICA & TÉCNICA EXPLORATÓRIA. 6ª EDIÇÃO, RIO DE JANEIRO, GUANABARA
KOOGAN, 1981, p.986.
16. TESTUT, L & LATARJET, A. ANATOMÍA HUMANA. TOMO II. NOVENA EDICIÓN, BARCELONA, SALVAT
EDITORES, 1977, p.1237.
17. TOLOSA, A & CANELAS, H. PROPEDÊUTICA NEUROLÓGICA. 2ª EDIÇÃO, SÃO PAULO, SARVIER, 1975,
526p.
44