SISTEMA DIGESTIVO
SUMÁRIO
ANATOMIA, HISTOLOGIA E FISIOLOGIA DO SISTEMA DIGESTIVO
1 - Introdução
2 – Cavidade oral: mucosa oral, palato, glândulas salivares menores, dentes e língua
3 – Glândulas salivares maiores: parótida, submandibular e sublingual
4 – Faringe
5 – Esófago
6 – Estômago. Mucosa gástrica e glândulas gástricas. Barreira Mucosa Gástrica. Secreção gástrica ácida e enzimática: as
glândulas gástricas e a regulação da secreção gástrica. Esfíncter gastro-esofágico inferior. Esvaziamento gástrico e esfíncter
pilórico.
7 – Intestino delgado: duodeno, jejuno e íleo. Pregas circulares e vilosidades intestinais. Epitélio intestinal e glândulas de
Lieberkühn.
8 – Intestino grosso: ceco, cólon ascendente, transverso, descendente e sigmóide, recto e canal anal.
9 – Peritoneu
10 – Fígado: lóbulo hepático, tríade portal, canais biliares, vesícula biliar, canal colédoco.
11 – Pâncreas
.
.
ANATOMIA E HISTOLOGIA DO SISTEMA DIGESTIVO
I – INTRODUÇÃO – CONSTITUIÇÃO – ORGANIZAÇÂO GERAL – ASPECTOS FUNCIONAIS
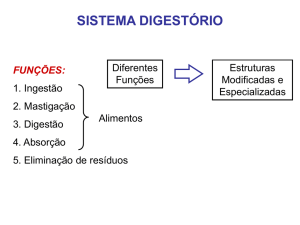
O sistema digestivo compreende todos os órgãos relacionados com a ingestão, mastigação, deglutição,
digestão dos alimentos, absorção dos nutrientes e a eliminação de alimentos não absorvíveis.
Anatomicamente o sistema digestivo é descrito como sendo constituído por um longo tubo que tem início na
fissura oral (a fenda entre os lábios superior e inferior) e termina no ânus, convencionalmente dividido pelas suas características
anatómicas, histológicas e fisiológicas em: cavidade oral, faringe, esófago, estômago, intestino delgado com as três porções
designadas duodeno, jejuno e íleo e intestino grosso com vários segmentos designados ceco, cólon ascendente, cólon
transverso, cólon descendenter, cólon sigmóide, recto e canal anal. Paralelamente ao longo tubo digestivo consideram-se as
1
seguintes estruturas anexas ao tubo digestivo: dentes, língua, glândulas salivares, fígado e pâncreas .
Histologicamente, a parede do tubo digestivo apresenta uma camada mais interna que é a mucosa, cujo epitélio
reveste o lúmen, a camada periférica da parede é uma espessa camada muscular, separada da camada mucosa, por uma
camada de tecido conjuntivo frouxo que constitui a submucosa
1, 2
.
As paredes do tubo digestivo, desde o esófago até ao ânus apresentam uma densa rede de neurónios que
constituem o sistema nervoso entérico dividido em dois plexos: o plexo submucoso ou plexo de Meissner situado na
submucosa e o plexo mioentérico ou plexo de Auerbach situado entre as camadas muscular e longitudinal. O plexo
submucoso e o plexo mioentérico permitem integrar localmente estímulos que desencadeiam actividade secretora e motora,
respectivamente, dando autonomia funcional ao tubo digestivo
3, 4
. Paralelamente, neurónios das componentes simpática e
parassimpática do sistema nervoso autónomo, terminam nestes plexos nervosos permitindo alterar a actividade digestiva
consoante as necessidades. A componente parassimpática aumenta a actividade peristáltica e secretora do tubo digestivo e
relaxa os esfíncteres. A componente simpática tem carácter inibitório, inibindo a actividade peristáltica e secretora e contraindo
os esfíncteres
3, 5
.
Na perspectiva fisiológica, o tubo digestivo assim constituído é capaz de fazer progredir os alimentos ingeridos
ao longo dos seus vários segmentos onde esses alimentos encontram compartimentos com características fisiológicas
diferentes, essas características dependem essencialmente das funções secretora e absortiva das células epiteliais da mucosa.
Nestes compartimentos, os alimentos previamente triturados por acção mecânica dos dentes, são colocados em contacto com
enzimas digestivas, as quais transformam as grandes biomoléculas constituintes dos alimentos em moléculas sufientemente
pequenas para atravessarem a membrana plasmática das células epiteliais, do lúmen em direcção ao espaço intersticial do
tecido conjuntivo de suporte colocado em torno da face basal do epitélio. Estas moléculas entram no sistema vascular e, através
do fluído plasmático chegam a todas as células do organismo. As substâncias que não foram absorvidas serão eliminadas
1
através do ânus .
A principal função do sistema digestivo é transformar os alimentos ingeridos em nutrientes assimiláveis pela
mucosa intestinal para que entrem no sangue. O sistema digestivo cumpre este objectivo porque tem capacidade de: motilidade,
secreção, digestão e absorção. A motilidade refere-se aos movimentos de mistura e propulsão dos alimentos ao longo do tubo
digestivo. A secreção refere-se à produção e libertação de substâncias no interior do tubo digestivo que colaboram na digestão,
humedecimento lubrificação dos alimentos e regulação do processo digestivo. A digestão refere-se à transformação das
grandes moléculas que constituem os alimentos, em pequenas moléculas que facilmente possam ser absorvidas pelo epitélio
3
intestinal. A absorção é a passagem de nutrientes, do lúmen do tubo digestivo para o sangue .
A motilidade permite a progressão dos alimentos ao longo do tubo digestivo, facilita a mistura dos alimentos com
as secreções e permite que os nutrientes fiquem em contacto com a face apical das células epiteliais responsáveis pela
3
absorção . A motilidade deve-se à contracção e relaxamento das fibras musculares lisas que formam a camada muscular do
tubo digestivo, controladas pelo sistema nervoso entérico e por substâncias endócrinas, a maior parte libertadas por células
3, 5
epiteliais da mucosa do tubo digestivo (células entero-endócrinas9
. O movimento propulsivo básico do tubo digestivo é a
4
peristalse ou reflexo peristáltico . A peristalse é uma propriedade inerente de estruturas tubulares com fibras musculares lisas
nas suas paredes. A distensão de um determinado segmento do tubo digestivo devido à acumulação de alimentos nesse
segmento é o principal estímulo que desencadeia a peristalse, caracterizada pela contracção de um anel de fibras musculares
do lado oral e relaxamento das fibras musculares do lado anal ao local da distensão. Por conseguinte os movimentos são
empurrados de oral para anal, passando para outro segmento do tubo digestivo, cuja distensão despoleta novamente a
formação do anel contráctil do lado oral e relaxamento de fibras musculares do lado anal ao novo segmento distendido
4, 5
. Este
movimento repete-se ao longo do tubo digestivo e o resultado é idêntico ao resultado de colocar os dedos à volta de um tubo
5
distendido, deslizando os dedos ao longo desse tubo .
Bibliografia
1 - Williams PL, Warwick R. Gray’s Anatomy. 38th Edition. Edinburgh: Churchill Livingstone; 1995. pp 1683-1815
2 - Junqueira LC, Carneiro J. Histologia Básica. 10ª Edição. Rio de Janeiro: Guanabara Koogan; 2004. pp 284-414.
3 - Berne RM, Levy MN. Physiology. 4th Edition. St Louis: Mosby; 1998. pp 589-674
4 - Kunze WAA, Furness JB. The Enteric Nervous System and Regulation of Intestinal Motility. Annual Review of Physiology. 61:117-142; 1999.
5 - Guyton AC, Hall JE. Textbook of Medical Physiology. 10th Edition. Philadelphia: WB Saunders Company; 2000. pp 718-833
II - A CAVIDADE ORAL
A cavidade oral consiste de uma pequena porção mais exterior ou periférica em forma de fenda designada
vestíbulo e uma porção maior designada cavidade oral propriamente dita. O vestíbulo está limitado externamente pelos
lábios e bochechas e internamente pelos arcos alveolares e dentes. A cavidade oral propriamente dita é o espaço limitado
anteriormente e lateralmente pelos arcos alveolares e dentes. Posteriormente a cavidade oral comunica com a faringe através
do istmo ou orifício orofaríngeo, o espaço limitado pelo arco palatoglosso e que constitui anatomicamente o limite posterior da
cavidade oral
1, 2
. Toda a cavidade oral está revestida por uma membrana mucosa que constitui a mucosa oral cujo epitélio é
pavimentoso estratificado, queratinizado ou não consoante o local da cavidade oral
3, 4
.
1 – LÁBIOS
Constituem a parede anterior da cavidade oral. São duas pregas carnosas revestidas externamente por pele e
internamente por mucosa oral, com uma camada muscular intermédia formada pelo músculo orbicular da boca. O espaço entre
lábio superior e lábio inferior é designado fissura oral, fenda oral ou orifício oral, estando limitada de cada lado pelas comissuras
1, 2
labiais
.
2 – BOCHECHAS
Formam as paredes laterais da cavidade oral, sendo constituídas principalmente pelo músculo bucinador,
revestido internamente por mucosa oral e externamente por pele
1, 2
.
3 – ARCOS ALVEOLARES MANDIBULAR E MAXILAR
Os arcos alveolares, são duas saliências ósseas no interior da cavidade oral, uma inferior que se salienta para
cima e outra superior que se salienta para baixo, apresentando forma de ferradura com concavidade voltada para o interior da
boca. O arco alveolar superior é formado pela apófise alveolar do osso maxilar de cada lado e o arco alveolar inferior é formado
pelo bordo superior do corpo da mandíbula. Os arcos alveolares são responsáveis pela fixação e suporte dos dentes, por isso,
possuem uma série de escavações, designadas alvéolos dentários onde encaixam as raízes dos dentes
1, 2
.
Os arcos alveolares são revestidos pela mucosa oral histologicamente idêntica e contínua com a restante
mucosa da cavidade oral. No rebordo do arco alveolar, também designado crista alveolar, a mucosa oral constitui a gengiva,
com uma porção firmemente aderida à crista alveolar e ao dente através de fibras de colagénio e uma margem livre que forma
um colarinho à volta do dente
2, 3
.
4 – PALATO
Forma o tecto da cavidade oral e, em simultâneo, constitui o soalho da cavidade nasal. Apresenta duas regiões
contínuas uma com a outra. A região mais extensa e mais anterior do palato constitui o palato duro, a região mais posterior
constitui o palato mole. A superfície do palato voltada para a cavidade oral está revestida por mucosa oral com o respectivo
2
epitélio pavimentoso estratificado .
O palato duro é a porção mais anterior e apresenta uma estrutura esquelética formada pela apófise palatina do osso
maxilar de cada lado e pela lâmina horizontal do osso palatino, de cada lado. A mucosa oral reveste esta estrutura esquelética e
apresena uma rafe tendinosa no plano mediano, designada rafe palatina que termina numa saliência anterior designada papila
incisiva. Da rafe palatina partem uma série de rugas ou saliências transversais e na submucosa sob a mucosa que cobre estas
rugas colocam-se inúmeras glândulas palatinas mucosas salivares, que fazem parte do conjunto de glândulas salivares
menores
1, 2
.
O palato mole é uma prega móvel suspensa no bordo posterior do palato duro e estendendo-se para trás e para baixo,
entre a nasofaringe e a orofaringe. Na sua posição normal, relaxada e pendente, a sua superfície inferior, voltada para a
cavidade oral é côncava para diante, enquanto a sua superfície superior voltada para a cavidade nasal é convexa para trás.
Apresenta uma camada muscular entre a camada mucosa da cavidade nasal que reveste a superfície superior, e a camada de
mucosa oral que reveste a face inferior
1, 2
. A camada muscular que entra na constituição do palato mole constitui a musculatura
palatina que permite a elevação do palato mole, no momento da deglutição, impedindo que os alimentos alcancem a
2
nasofaringe .
Da margem inferior e posterior livre do palato mole pende uma estrutura arredondada designada úvula. De cada lado
da base da úvula partem duas pregas para baixo e lateralmente, formando com as do lado oposto dois arcos. Estas pregas são
saliências da mucosa causadas por feixes de fibras musculares que entram na constituição da parede da faringe. O arco
anterior designa-se arco palatoglosso e é o limite anatómico posterior da cavidade oral, limitando o istmo ou orifício
orofaríngeo. O arco posterior já faz parte da faringe e designa-se arco palatofaríngeo. Entre o arco palatoglosso e o arco
palatofaríngeo, colocada na faringe, de cada lado, situa-se a amígdala palatina
1, 2
. Cada amígdala palatina é um agregado de
2
tecido linfóide implicado na defesa contra agentes patogénicos .
5 – LÍNGUA
A língua é um órgão muscular associado à deglutição, fala e paladar, situando-se parcialmente na boca e
parcialmente na faringe. Pelos seus componentes musculares, está inserida: no osso hióide, palato mole, mandíbula, faringe e
apófise estilóide do osso temporal. As porções mais anterior e a mais posterior constiuem o ápice e a raiz da língua,
2
respectivamente, a superfície superior (ou póstero-superior pela posição que adquire) é designada o dorso da língua .
Todas as superfícies da língua estão revestidas por mucosa oral com um epitélio pavimentoso estratificado
suportado por uma lâmina de tecido conjuntivo que separa o epitélio das fibras musculares
2, 3
. O dorso da língua é convexo e
apresenta um sulco em V designado sulco terminal, que divide a língua em duas porções, a porção oral e a porção faríngea
2
1,
.
A porção oral da língua, formando os dois terços anteriores, está colocada sobre o soalho da boca, o seu ápice une-se
ao soalho da boca pelo freio lingual e encosta nas faces posteriores dos dentes incisivos. O dorso da porção oral da língua
está voltado para cima, apresenta um sulco no plano mediano e inúmeras saliências da mucosa, as papilas linguais que
aumentam área de contacto com os alimentos que estão a ser saboreados. As papilas linguais apresentam formas
diversificadas e são classificadas, segundo essa forma, em papilas filiformes, papilas fungiformes, papilas foliadas e papilas
1, 2
valadas
. As papilas valadas são as maiores, estão colocadas imediatamente à frente do sulco terminal. São estruturas
cilíndricas com 1 ou 2 milímetros de diâmetro, com um profundo sulco à volta que separa a papila de uma elevação da mucosa
mais periférica que contorna a papila. Numerosas glândulas serosas, as glândulas de von Ebner, existentes no tecido conjuntivo
1, 2, 3
de suporte ao epitélio, segregam substâncias para o sulco que rodeia a papila valada
. Estas glândulas produzem uma
enzima, a lipase lingual, que previne a formação de uma camada hidrofóbica sobre o dorso da língua que prejudicaria a função
sensitiva. Além disso, esta lipase lingual parece actuar sobre o bolo alimentar permitindo que uma pequena percentagem de
triglicerídeos existentes na dieta sejam digeridos no estômago por esta enzima
3, 5, 6
.
No epitélio da mucosa que reveste o dorso da porção oral da língua aparecem estruturas microscópicas, os botões ou
corpúsculos gustativos com função sensitiva, responsáveis pelo paladar. Cada botão gustativo é um conjunto de 50 a 100
células, colocadas no epitélio da mucosa oral, mas morfologicamente distintas das células epiteliais de revestimento
organizadas de modo a formar um minúsculo órgão em forma de barril. A maior parte das células que formam o botão gustativo
são receptores sensitivos que despolarizam, quando em contacto com determinadas substâncias dissolvidas na saliva. A
despolarização destas células leva à libertação de neurotransmissores que estimulam fibras nervosas aferentes, estas são
responsáveis por transportar estes estímulos nervosos ao sistema nervoso central. O resultado é a percepção de quatro tipos de
sabor: salgado, azedo, doce e amargo, cada um deles despoletado pela presença de determinadas moléculas dissolvidas na
saliva. Para o contacto com estas moléculas, as células sensitivas possuem microvilosidades no seu polo apical, que está em
contacto com um minúsculo poro existente no epitélio da mucosa oral. Cada botão gustativo está em contacto com um poro e
todos os poros espalhados na mucosa oral comunicam livremente com o lúmen da cavidade oral
3, 4
.
A porção faríngea também designada base da língua forma o terço posterior da língua, está colocada na faringe, atrás
do arco palatoglosso e seu dorso fica voltado para trás, constituindo a parede anterior da orofaringe. É desprovida de papilas
linguais mas contém agregados de tecido linfóide que causam elevações arredondadas na mucosa do dorso desta parte da
língua
2, 3
.
6 – SOALHO DA BOCA
O músculo milohioideu é o principal componente do soalho da boca. É revestido internamente pela mucosa oral que se
reflecte e se continua na mucosa que reveste a língua e a face interna do arco alveolar inferior
7 – DENTES
1, 2
.
1, 2, 3
7.1 – Morfologia dentária
Os dentes são órgãos fundamentais para a mastigação, colocados na cavidade oral, inseridos nos arcos alveolares.
Um dente consiste de duas partes: a coroa que é a porção do dente que fica voltada para a cavidade oral e a raiz ou raízes
que constituem a porção do dente que fica inserida no alvéolo, a escavação óssea do arco alveolar onde se insere o dente.
A coroa está revestida perifericamente pelo esmalte tecido altamente mineralizado e acelular que atinge uma
espessura máxima que ronda 2,5 milímetros. Cerca de 96% do esmalte é substância inorgânica constituída essencialmente por
cristais de hidroxiapatite. Estes cristais organizam-se para formar os prismas do esmalte que são colunas que percorrem toda a
camada de esmalte, desde a junção esmalte-dentina, até à superfície da coroa. A coroa pode apresentar bordos incisais, ou
então cúspides separadas por fissuras.
A raiz ou raízes dentárias estão revestidas perifericamente por uma camada de tecido duro, o cimento, que com o
avançar da idade pode atingir 1 milímetro de espessura. O cimento é um tecido mineralizado com um componente inorgânico
que corresponde a cerca de 50% do seu peso, constituído essencialmente por cristais de hidroxiapatite e fosfato de cálcio. Este
tecido é atravessado por fibras de colagénio que se inserem por um lado ao cimento e, por outro ao lado, ao osso alveolar,
constituindo o ligamento periodontal, permitindo a fixação do dente ao osso envolvente.
A zona de junção entre coroa e raiz designa-se colo ou margem cervical.
Num corte longitudinal, o dente revela: um espaço no seu interior, a cavidade pulpar que continua em cada raiz como
canal pulpar. Cavidade e canal pulpares são preenchidos pela polpa, formada por tecido conjuntivo que recebe vasos e
estruturas nervosas provenientes do osso alveolar, através do orifício apical, situado no ápice da raiz. À volta da cavidade e
canal pulpares há uma camada de tecido mineralizado designado dentina. A porção inorgânica da dentina que corresponde a
cerca de 70% do seu peso é constituída essencialmente por cristais de hidroxiapatite. Este tecido possui inúmeros canalículos
preenchidos por prolongamentos citoplasmáticos de células colocadas na polpa e designadas odontoblastos. Estes
prolongamentos citoplasmáticos são responsáveis pela sensibilidade dentária. A matriz orgânica da dentina é produzida pelos
odontoblastos, mais tarde esta matriz mineraliza. A espessura da dentina aumenta com a idade roubando espaço ao
compartimento pulpar.
7.2 – Dentição permanente e dentição decídua
Uma linha imaginária que passa entre os dois incisivos centrais superiores e inferiores permite dividir a boca em quatro
quadrantes: o quadrante superior direito ou 1º quadrante na dentição permanente, 5º quadrante na dentição decídua; o
o
quadrante superior esquerdo ou 2 quadrante na dentição permanente, 6º quadrante na dentição decídua; o quadrante inferior
esquerdo ou 3º quadrante na dentição permanente, 7º quadrante na dentição decídua; o quadrante inferior direito ou 4º
quadrante na dentição permanente, 8º quadrante na dentição decídua.
Num adulto, a dentição completa em cada quadrante é formada por: 2 dentes incisivos, 1 dente canino, 2 dentes prémolares, 3 dentes molares. Este conjunto de dentes nos quatro quadrantes constitui a dentição permanente. No entanto, os
incisivos, caninos e pré-molares vão substituir a designada dentição decídua, formada em cada quadrante por: 2 dentes
incisivos, 1 dente canino, 2 dentes molares.
A substituição dos dentes decíduos faz-se em dois períodos diferentes. Entre os cinco e os sete anos de idade, os dentes
incisivos decíduos são substituídos pelos homónimos da dentição permanente e atrás da dentição decídua irrompem os
primeiros molares permanentes, também designados molares dos seis anos. Entre os nove e doze anos de idade os molares
decíduos são substituídos pelos pré-molares, os caninos decíduos são também substituídos e atrás dos primeiros molares
irrompem os segundos molares também designados molares dos doze anos. Mais tarde, após os dezassete anos, erupcionam
os terceiros molares
Bibliografia
1 - Netter FH. Atlas de Anatomia Humana. Segunda Edición. East Hanover. Novartis; 1999.
2 - Williams PL, Warwick R. Gray’s Anatomy. 38th Edition. Edinburgh: Churchill Livingstone; 1995. pp 1683-1815
3 - Junqueira LC, Carneiro J. Histologia Básica. 10ª Edição. Rio de Janeiro: Guanabara Koogan; 2004. pp 284-414.
4 - Kierszenbaum AL. Histologia e Biologia Celular. Uma Introdução à Patologia. Rio de Janeiro: Elsevier 2004.
5 - Berne RM, Levy MN. Physiology. 4th Edition. St Louis: Mosby; 1998. pp 589-674
6 - Guyton AC, Hall JE. Textbook of Medical Physiology. 10th Edition. Philadelphia: WB Saunders Company; 2000. pp 718-833
III - GLÂNDULAS SALIVARES
1 – ANATOMIA DAS GLÂNDULAS SALIVARES 1, 2
Num sentido lato são células ou órgãos que libertam secreções para a cavidade oral, que no seu conjunto formam a
saliva. Convencionalmente pode fazer-se a distinção entre as glândulas salivares maiores, situadas a alguma distância da
mucosa oral, comunicando com a cavidade oral através de canais extraglandularese e as glândulas salivares menores
espalhadas na mucosa e submucosa da cavidade oral.
Há três pares de glândulas salivares maiores. As glândulas parótidas, uma de cada lado estão situadas abaixo do
orifício do meato acústico externo, entre a mandíbula e o músculo esternocleidomastoideu, estendendo-se para diante, na
superfície do músculo masseter. Seu canal excretor dirige-se para diante na superfície do músculo masseter, perfura o músculo
bucinador e abre-se numa papila da mucosa oral, na bochecha ao nível da coroa do 2º molar superior. As glândulas
submandibulares, uma de cada lado, estão situadas sob o soalho da cavidade oral, encostadas à face interna do corpo da
mandíbula, na designada fossa submandibular, abaixo da linha milohioideia da mandíbula. O canal excretor abre-se no
pavimento da boca, numa papila próxima do freio lingual. As glândulas sublinguais, uma de cada lado, estão situadas sob a
membrana mucosa do soalho da boca, sob a ponta da língua em contacto com a fossa sublingual da face interna da porção
anterior do corpo da mandíbula. Seus canais excretores são vários e abrem-se no soalho da boca sob o ápice ou ponta da
língua.
2 – MICRO-ESTRUTURA DAS GLÂNDULAS SALIVARES
As glândulas salivares maiores são constituídas por aglomerados de células reunidas em lobos separados entre si por
septos de tecido conjuntivo. Cada lobo é uma porção de parênquima constituído por vários lóbulos, por sua vez, cada lóbulo é
formado por um canal excretor que se ramifica no interior do lóbulo e cada ramo terminal acaba numa porção secretora
dilatada e arredondada, o ácino ou alvéolo glandular. O ácino é constituído por um grupo de células com função secretora
cujas secreções são libertadas numa estrutura tubular de diminutas dimensões, este conjunto constitui a terminação secretora
da glândula. As terminações secretoras convergem para o canal intercalar. Vários canais intercalares de um lóbulo convergem
para um canal estriado de dimensões maiores. Os vários canais estriados de um lóbulo convergem para um canal excretor que
abandona o lóbulo. Diferentes canais excretores convergem para formar um ou mais canais excretores de maiores dimensões
que desembocam na cavidade oral
2, 3
.
Nas terminações secretoras encontram-se dois tipos de células secretoras: as células acinares serosas, produtoras de
um fluído de baixa viscosidade, constituído por proteínas entre as quais a amilase, a lisozima e a peroxidase e as células
acinares mucosas, produtoras de substâncias com elevada viscosidade nomeadamente glicoproteínas do grupo das mucinas.
2, 4, 5
Cada ácino pode conter apenas células mucosas, apenas células serosas, ou ambos os tipos de células
. Na periferia das
terminações secretoras e dos canais intercalares encontram-se células mioepiteliais com capacidade contráctil com o objectivo
2
de aumentar o fluxo salivar para os canais estriados e excretores .
3 – A SALIVA E A PRODUÇÃO DE SALIVA PELAS GLÂNDULAS SALIVARES
A secreção das glândulas salivares é um fluido aquoso contendo água, electrólitos e uma mistura de proteínas e mucinas
com grande peso molecular
7, 8
. A água presente na saliva facilita a formação do bolo alimentar e funciona como solvente não só
para diluição das moléculas que estimulam as células sensitivas responsáveis pelo paladar, mas também para que sejam
2
possíveis as reacções enzimáticas digestivas . As mucinas têm como função lubrificar o bolo alimentar facilitando o seu
percurso ao longo do tubo digestivo, evitando lesões da mucosa pelo atrito
amilase, uma enzima responsável pela digestão de polissacarídeos
4, 6
2, 4, 6
. Das proteínas presentes na saliva destaca-se a
, no entanto, outras proteínas estão presentes como a
lisozima, a lactoperoxidase e imunoglobulinas da classe A, todas elas com acção antimicrobiana
4, 5, 6
.
Sendo assim, a saliva providencia hidratação e protecção da mucosa oral, orofaríngea e esofágica, acção
antimicrobiana e acção digestiva enzimática sobre o bolo alimentar. A importância destas funções tornam-se evidentes sempre
que há hipofunção das glândulas salivares, situação que se manifesta clinicamente por dor oral, aumento de ocorrência de cárie
5
dentária e de infecções oportunistas .
A salivação é um processo rigorosamente controlado, ocorrendo a uma taxa muito baixa entre as refeições e quase nula
5
6
durante o sono . O ser humano produz 800 a 1500 mililitros, em média 1000 mililitros por dia e cerca de 80 a 90% ocorre
5
durante as refeições em resposta a estímulos tais como olfacto, paladar e forças mastigatórias
5, 6
.
A produção de saliva pelas glândulas salivares pressupõe dois passos sequenciais. No primeiro passo as terminações
secretoras, provavelmente (?) com a participação dos canais intercalares, segregam um fluído para o lúmen do canal intercalar,
resultante do transporte activo de Na+ e Cl- e do transporte passivo de água constituindo a secreção primária. Este fluido é
isotónico em relação ao plasma com predominância dos iões Na+ e Cl- tal como no fluído plasmático
4, 5, 6
. O segundo passo no
mecanismo de secreção salivar ocorre ao longo dos canais estriados e dos canais excretores, onde a secreção primária é
modificada. A este nível os iões Na+ e Cl- são retirados do lúmen do canal estriado regressando ao plasma e, em simultâneo,
são adicionados iões K+ e HCO3- ao fluído no lúmen do canal estriado
4, 6
.
O fluído resultante é sempre hipotónico em relação ao plasma com concentrações de Na+ e Cl- inferiores e com
concentrações K+ e HCO3- superiores às do fluido plasmático
4, 6
. À medida que aumenta o fluxo salivar também aumenta a
tonicidade da saliva, no entanto, mesmo quando a taxa de secreção salivar atinge o valor máximo a tonicidade salivar
4
permanece inferior à plasmática . O pH é variável consoante as circunstâncias: o pH da saliva produzida entre as refeições
(glândulas em repouso) é ligeiramente ácida, no entanto, quando as glândulas são estimuladas a aumentar a taxa de secreção
salivar, o pH da saliva atinge um valor próximo de 8, devido a um gradual aumento da concentração salivar de ião bicarbonato,
à medida que aumenta a taxa de secreção salivar
4, 6
.
No tecido conjuntivo de suporte do parênquima glandular há plasmócitos que produzem imunoglobulinas da classe A
3
que, através das células epiteliais das terminações acinares e dos canais intercalares estriados são adicionadas à saliva .
3 – REGULAÇÃO DA SECREÇÃO PELAS GLÂNDULAS SALIVARES
A secreção salivar é fortemente estimulada pela componente parassimpática do sistema nervoso autónomo. A
estimulação quer da componente simpática quer da componente parassimpática estimulam a secreção salivar, no entanto, o
4
efeito da estimulação parassimpática é mais forte e duradouro . Estimulação parassimpática aumenta a síntese e secreção de
4
amilase salivar e mucinas , aumenta o transporte de iões e água através do epitélio dos canais intercalares e estriados,
4, 5
aumenta o fluxo sanguíneo facilitando a secreção de iões e água para o lúmen canalicular
e estimula o metabolismo e
4
crescimento da glândula salivar .
A estimulação da componente simpática do sistema nervoso autónomo, da mesma forma que acontece com a
estimulação da componente parassimpática, origina contracção de células mioepiteliais colocadas na periferia dos ácinos
glandulares. Esta contracção permite esvaziar o conteúdo dos ácinos nos canais intercalares, estriados e excretores
2
aumentando o fluxo salivar . No entanto, ao contrário do que sucede com a estimulação da componente parassimpática, a
estimulação da componente simpática causa vasoconstrição acentuada o que restringe o fluxo de sangue na glândula salivar
comprometendo a secreção salivar. Por conseguinte, a estimulação da componente simpática causa um pequeno aumento da
secreção salivar que é de curta duração e que imediatamente reverte para a diminuição da secreção salivar
2, 4, 6
.
Bibliografia
1- Netter FH. Atlas de Anatomia Humana. Segunda Edición. East Hanover. Novartis; 1999.
2 - Williams PL, Warwick R. Gray’s Anatomy. 38th Edition. Edinburgh: Churchill Livingstone; 1995. pp 1683-1815
3 - Junqueira LC, Carneiro J. Histologia Básica. 10ª Edição. Rio de Janeiro: Guanabara Koogan; 2004. pp 284-414.
4 - Berne RM, Levy MN. Physiology. 4th Edition. St Louis: Mosby; 1998. pp 589-674.
5 - Melvin JE. Yule D, Shuttleworth T, Begenisich T. Regulation of Fluid and Electrolyte Secretion in Salivary Gland Acinar Cells. Annual Review
of Physiology. 67:445-469; 2005.
6 - Guyton AC, Hall JE. Textbook of Medical Physiology. 10th Edition. Philadelphia: WB Saunders Company; 2000. pp 718-833
7 - Dockray GJ. Varro A, Dimaline R, Wang T. The Gastrins. Annual Review of Physiology. 63:119-139; 2001.
8 - Geibel JP. Secretion and Absortion by Colonic Crypts. Annual Review of Physiology. 67:471-490; 2005.
IV – FARINGE E ESÒFAGO
1 – Aspectos Anatómicos da Faringe
1, 2
È uma estrutura tubular musculomembranosa, colocada atrás da cavidade nasal, boca e laringe. Faz parte do sistema
digestivo, mas sua porção superior permite a passagem de ar das cavidades nasais para a laringe. Em cima, a faringe está
limitada pelo corpo do osso esfenóide e porção basilar do osso occipital. Atrás está encostada á porção cervical da coluna
vertebral. À frente abre-se na cavidade nasal, boca e laringe, sua parede anterior está incompleta. Caudalmente termina ao
nível da 6ª vértebra cervical, num plano coincidente ao bordo inferior da cartilagem, a partir do qual é contínua com o esófago.
Na faringe são consideradas três partes. A parte nasal ou nasofaringe está colocada atrás da cavidade nasal e acima do
palato mole. Entre a margem livre do palato mole e a parede posterior da faringe, a nasofaringe comunica com a orofaringe pelo
istmo ou orifício faríngeo. De cada lado, nas paredes laterais da nasofaringe, estão as aberturas faríngeas das tubas auditivas,
fazendo a comunicação entre as cavidades timpânicas e a nasofaringe. A parte oral ou orofaringe está colocada atrás da
cavidade oral, entre o palato mole até ao bordo superior da epiglote. Abre-se anteriormente na boca, através do orifício
orofaríngeo. Nas suas paredes laterais estão as amígdalas palatinas constituídas por colecções de tecido linfóide com a
mesma organização dos nódulos linfáticos, colocadas uma de cada lado entre as pregas que formam os arcos palatoglosso e
palatofaríngeo. A parte laríngea da faringe ou laringofaringe, está colocada desde o bordo superior da epiglote ao bordo
inferior da cartilagem cricóide, onde continua com o esófago. Na sua parede anterior está a abertura superior da laringe.
Estruturalmente a faringe é constituída por três camadas. A membrana mucosa é a camada mais interna, apresenta um
epitélio colunar pseudo-estratificado e ciliado ao nível da nasofaringe, mas nas outras partes da faringe o epitélio é pavimentoso
estratificado, idêntico ao da cavidade oral. A camada intermédia é fibrosa. É reforçada posteriormente por uma forte banda
fibrosa, inserida em cima no tubérculo faríngeo da face inferior da porção basilar do osso occipital e que forma a rafe faríngea,
no plano mediano da parede posterior da faringe. A camada muscular é a mais externa e é constituída por três músculos, os
constritores superior, médio e inferior da faringe. Posteriormente os músculos unem-se aos do lado oposto inserindo-se na rafe
faríngea.
2 – Aspectos Anatómicos e Histológicos do Esófago
É um tubo muscular que liga a faringe ao estômago. Tem início no pescoço ao nível do bordo inferior da cartilagem
cricóide da laringe, onde é contínuo com a faringe. Coloca-se logo à frente da coluna vertebral, ao longo de todo o mediastino,
até atingir o diafragma que perfura anteriormente à artéria aorta, alcançando a cavidade abdominal. Termina no orifício cárdico
1, 2
do estômago. Anteriormente ao esófago está colocada a traqueia
.
Histologicamente o esófago apresenta do interior para o exterior: uma camada mucosa com epitélio pavimentoso
estratificado, uma camada submucosa que une a mucosa à camada muscular e que é constituída por tecido conjuntivo com
inúmeras glândulas mucosas, cujos canais se abrem no lúmen do esófago e uma camada muscular constituída por uma
subcamada mais interna com fibras musculares com disposição circular e uma subcamada mais externa com fibras musculares
2, 3
colocadas longitudinalmente
.
Quer a extremidade superior quer a inferior do esófago funcionam como esfíncteres fisiológicos que mantém os
orifícios superior e inferior sempre fechados, excepto no momento da deglutição. Estes esfíncteres designam-se esfíncter
esofágico superior e esfíncter esofágico inferior, este último é também designado esfíncter gastro-esofágico. O primeiro evita a
entrada de ar no tubo digestivo e o segundo evita o refluxo dos alimentos do estômago para o esófago. Este último aspecto
reveste-se de grande importância dado que a mucosa esofágica não tem protecção contra o pH extremamente ácido do
4
conteúdo gástrico .
2
– Aspectos Fisiológicos – Mecanismo da Deglutição
O reflexo da deglutição é é uma sequência rigidamente ordenada de eventos que empurram o bolo alimentar da boca até
ao estômago. O mecanismo da deglutição ocorre em três fases: oral, faríngea e esofágica. A fase oral é voluntária e é iniciada
quando o bolo alimentar, por acção da língua, é empurrado para a porção posterior da cavidade oral. O bolo alimentar é
empurrado para a faringe, onde receptores sensitivos são estimulados despoletando mecanismos reflexos. Durante a fase
faríngea, o palato mole eleva-se fechando a passagem para a nasofaringe. A laringe é deslocada para cima e para diante
devido à elevação do osso hióide em resposta à contracção dos músculos supra-hioideus. Este deslocamento da laringe
permite que a epiglote incline para trás, cobrindo o orifício superior da laringe. Além disso, as pregas vocais aproximam-se
fechando a glote para que os alimentos não entrem na traqueia. Uma onda peristáltica percorre a faringe relaxando o esfíncter
esofágico superior, os alimentos são empurrados para o esófago. Esta fase ocorre em menos de um segundo. A fase esofágica
caracteriza-se por uma onda peristáltica que percorre o esófago causando relaxamento do esfíncter esofágico inferior ou
gastroesofágico. Esta fase tem uma duração de cerca de 10 segundos terminando com a entrada do bolo alimentar no
4
estômago .
Bibliografia
1
- Netter FH. Atlas de Anatomia Humana. Segunda Edición. East Hanover. Novartis; 1999.
2
- Williams PL, Warwick R. Gray’s Anatomy. 38th Edition. Edinburgh: Churchill Livingstone; 1995. pp 1683-1815
3
- Junqueira LC, Carneiro J. Histologia Básica. 10ª Edição. Rio de Janeiro: Guanabara Koogan; 2004. pp 284-414.
4
- Berne RM, Levy MN. Physiology. 4th Edition. St Louis: Mosby; 1998. pp 589-674.
VI – ESTÔMAGO
1 – ASPECTOS ANATÓMICOS
1, 2
É a porção mais dilatada do tubo digestivo, situado entre o esófago e o intestino delgado, na porção mais superior do
abdómen. Tem dois orifícios, dois bordos e duas faces. No estômago estão identificadas quatro regiões: cárdia, fundo, corpo
e piloro
O orifício de comunicação entre esófago e estômago é orifício cárdico situado à esquerda da linha mediana, por trás
da 7ª cartilagem costal, 2,5 cm à esquerda da junção dessa cartilagem costal com o esterno. A curta porção abdominal do
esófago tem forma de cone e curva-se para a esquerda para terminar no estômago, na base deste cone está o orifício cárdico.
O orifício de comunicação entre estômago e intestino delgado é o orifício pilórico, identificado por uma constrição bem
marcada na superfície externa do estômago, correspondente ao esfíncter pilórico. Desde o orifício cárdico ao orifício pilórico há
dois bordos, um do lado direito e outro do lado esquerdo
O bordo direito do estômago constitui a curvatura menor do estômago, voltada para cima e para a direita.
Cranialmente esta curvatura é contínua com o lado direito do esófago ao nível do orifício cárdico e a partir deste, tem um
percurso descendente, apresentando concavidade para a direita. Caudalmente, esta curvatura é atravessada por um sulco
designado incisura angular e, a partir deste ponto, adopta um percurso para a direita para terminar ao nível do orifício pilórico.
O bordo esquerdo do estômago tem um trajecto muito mais longo e constitui a curvatura maior do estômago. Esta
curvatura é contínua com o lado esquerdo do esófago e o local onde este lado do esófago termina para ter início a curvatura
maior do estômago, há um ângulo agudo que constitui a incisura cárdica. Com início na incisura cárdica, a curvatura maior
descreve um arco para cima e para a esquerda do orifíco cárdico, apresentando depois um percurso descendente com
convexidade para a esquerda e, finalmente, volta-se para a direita para terminar ao nível do orifício pilórico
No interior do estômago, apesar de constituir um compartimento único, podem ser identificadas regiões com algumas
características histológicas e funcionais próprias. O segmento mais proximal do estômago constitui a cárdia, um segmento com
2 a 3 centímetros de comprimento a partir do orifício cárdico. A incisura cárdica invagina para o interior deste segmento do
estômago, formando uma prega que, mediante um aumento da pressão intra-gástrica, ajuda a impedir o refluxo gastroesofágico. O segmento do estômago situado acima do nível do orifício cárdico constitui o fundo. O restante do estômago
proximal à incisura angular constitui o corpo. A porção distal à incisura angular constitui o piloro com uma porção mais
proximal e alargada, o antro pilórico e uma porção mais estreita e distal, o canal pilórico que termina no orifício pilórico.
2 - ORGANIZAÇÂO HISTOLÓGICA
A organização histológica da parede do estômago é similar à do restante tubo digestivo consistindo de três camadas
designadas do interior para a periferia: mucosa gástrica, submucosa e muscular, esta última envolvida perifericamente pelo
peritoneu que constitui a camada serosa
2, 3, 4
.
Relativamente à muscular externa, a parede do estômago apresenta três camadas de fibras musculares lisas, sendo a
mais profunda ou interna, designada camada oblíqua, constituida por fibras musculares colocadas obliquamente na parede do
estômago, a camada intermédia é designada camada circular, constituída por fibras musculares colocadas circunferencialmente
e a camada mais periférica é a camada longitudinal constituída por fibras musculares colocadas longitudinalmente. Neste
aspecto, a parede do estômago difere do restante tubo digestivo pela adição da camada oblíqua internamente às outras duas
2, 3
1,
.
A submucosa do estômago, tal como no restante tubo digestivo, consiste de uma camada de tecido conjuntivo contendo
vasos sanguíneos e estruturas nervosas organizadas de forma a constituir um plexo nervoso
2, 3
.
A constituição da mucosa gástrica, embora com características particulares, apresenta uma organização comum à
mucosa do restante tubo digestivo, por conseguinte, é constituída por um epitélio que reveste o lúmen do estômago suportado
por uma camada de tecido conjuntivo frouxo, a lâmina própria, onde se encontram aglomerados de tecido linfóide. A separar a
mucosa da submucosa está um fino estrato de fibras musculares lisas constituindo a camada muscular da mucosa
2, 3
.
3 – A MUCOSA GÁSTRICA
A mucosa gástrica, a camada mais interna da parede do estômago, é espessa e com superfície aveludada. Observando
a olho nu a face interna do estômago vazio, a mucosa apresenta pregas ou rugas que representam elevações da submucosa
em direcção ao lúmen que desaparecem se a parede do estômago for distendida. Quando observada microscopicamente, a
superfície interna da parede do estômago apresenta estrutura em forma de favo de mel, com elevações da mucosa que
delimitam as fossetas ou fovéolas gástricas, estruturas irregulares com cerca de 0,2 milímetros de diâmetro. O soalho de cada
fosseta gástrica apresenta vários orifícios e cada um destes orifícios é a entrada para uma glândula gástrica tubular que se
2
estende ao longo de toda a espessura da lâmina própria (tecido conjuntivo da mucosa), atá à muscular da mucosa .
O epitélio da mucosa gástrica é cilíndrico simples. Reveste as elevações da mucosa e aprofunda-se ao longo da lâmina
2, 3
própria para revestir o lúmen de estruturas tubulares que constituem as glândulas gástricas
.
As células epiteliais que revestem as elevações da mucosa e o soalho das fossetas gástricas são designadas células
2
mucosas superficiais . A sua função é produzir muco e um fluído com elevada quantidade de iões bicarbonato que ficam
aprisionados na camada de muco, conferindo-lhe um carácter alcalino
5, 6
. Esta camada de muco e os iões bicarbonato nele
5
aprisionados constituem a barreira mucosa gástrica, com cerca de 200 micrómetros de espessura , que separa a superfície
apical das células epiteliais, onde o pH ronda o valor 7, do conteúdo gástrico extremamente ácido
5, 6
. A barreira mucosa tem
função essencialmente protectora, qualquer factor que comprometa esta camada, inibindo a secreção de muco ou a de
bicarbonato, proporciona a ocorrência de úlceras gástricas pelo contacto das células da mucosa com o conteúdo ácido e
5
proteolítico do lúmen do estômago .
A partir do soalho da fosseta gástrica o epitélio da mucosa gástrica aprofunda-se no tecido conjuntivo de suporte,
formando estruturas tubulares que constituem as glândulas gástricas. Ao longo da glândula as células epiteliais adquirem
diferentes características morfológicas associadas a secreções diferenciadas
2, 3
. Embora semelhantes em muitos aspectos, as
glândulas gástricas adquirem características distintas nas diferentes regiões do estômago, por conseguinte, costumam
2
3
2
3
distinguir-se três tipos de glândulas gástricas: as glândulas cárdicas , ou da cárdia , as glândulas principais , ou fúndicas ,
5, 6
ou oxínticas
e as glândulas pilóricas
2, 3, 6
.
As glândulas cárdicas estão confinadas à estreita região da cárdia, apresentam lúmen amplo, predominando as células
produtoras de muco. Células produtoras de ácido clorídrico são escassas, no entanto, existentes
2, 3
.
As glândulas pilóricas existentes na região do piloro formam estruturas tubulares curtas onde predominam células
secretoras de muco entre as quais se encontram numerosas células entero-endócrinas, particularmente células-G produtoras
de uma hormona designada gastrina
2, 3, 6
.
As glândulas principais ou fundicas ou oxínticas são as mais numerosas estando espalhadas pelo fundo e corpo do
estômago. Cerca de 3 a 7 destas glândulas abrem-se no soalho de cada fosseta gástrica
2, 3
. Cada uma destas glândulas
7
apresenta quatro regiões distintas à medida que se aprofunda na lâmina própria da mucosa. A porção que abrange a fosseta , a
porção que se abre no soalho da fosseta e que constitui o istmo, a região seguinte é o colo da glândula e a restante e mais
extensa porção da glândula é a base. Cada uma destas glândulas apresenta, pelo menos, cinco tipos distintos de células
epiteliais cuja distribuição não é uniforme ao longo da glândula
2, 3, 7
.
3.1 – Stem cells ou células tronco
São células indiferenciadas existentes no istmo da glândula que apresentam actividade mitótica, as células filhas migram
apicalmente para se diferenciarem em células mucosas superficiais, ou migram basalmente para se diferenciarem nos
2, 3
diferentes tipos de células existentes nas glândulas gástricas
.
3.2 – Células mucosas do colo
Tal como o nome indica estas células estão situadas no colo da glândula e produzem muco, embora com características
histoquímicas distintas do muco produzido pelas células mucosas superficiais
2, 3
.
3.3 – Células principais, pépticas, ou zimogénicas
São células localizadas na base da glândula responsáveis pela secreção de uma enzima proteolítica designada pepsina.
Apresentam características de células secretoras de proteínas, com abundante retículo endoplasmático rugoso, complexo de
Golgi desenvolvido e acentuada basofilia devido à predominância de RNA no seu citoplasma. Apresentam inúmeros grânulos
de secreção no citoplasma contendo pepsinogénio
2,3
.
Quando estimuladas, as células principais libertam o conteúdo dos seus grânulos de secreção para o lúmen da glândula,
libertando o pepsinogénio que, em contacto com o meio ácido, é convertido na sua forma activa, a pepsina. Esta enzima tem
actividade proteolítica máxima em meio com pH 3 ou inferior, tornando-se inactiva se o pH atingir o valor 5. Actua nas ligações
peptídicas, transformando cerca de 20% das proteinas da dieta em pequenos peptídeos
5, 6
.
3.4 – Células parietais ou oxínticas
As células parietais estão presentes na porção da glândula mais próxima do colo. São células arredondadas ou
piramidais que segregam ácido clorídrico e factor intrínseco
3, 5, 6, 8
. Na célula em repouso, observam-se inúmeras estruturas
membranosas tubulo-vesiculares adquirindo a forma de vesículas, túbulos ou cisternas que ocupam o citoplasma próximo da
membrana plasmática apical. Nas células em intensa actividade secretora, a característica mais marcante observável com
microscópio electrónico, é uma profunda invaginação da membrana plasmática apical formando um canalículo intracelular, com
abundantes microvilosidades voltadas para o lúmen do canalículo
3, 7, 8
.
A membrana que envolve as estruturas tubulo-vesiculares da célula em repouso é rica numa proteína de transporte
responsável por deslocar contra gradiente de concentração iões hidrogénio para o exterior e iões potássio para o interior. Esta
proteína de transporte designada H+/ K+ - ATPase, constitui a bomba de protões ou bomba gástrica, ou bomba H+/K+, de
importância fulcral para a secreção de ácido clorídrico pelas células parietais. Na célula em repouso a H+/K+ ATPase está
inactiva. Quando a célula parietal é estimulada para segregar ácido clorídrico, ocorrem profundas alterações morfológicas na
célula com a fusão das estruturas tubulo-vesiculares à membrana plasmática apical que invagina para o interior do célula
7, 8
constituindo o canalículo intracelular
. A cessação da secreção de ácido clorídrico ocorre após o restabelecimento das
7
estruturas tubulo-vesiculares .
A secreção de ácido clorídrico pelas células parietais depende do movimento de iões através da membrana plasmática
das células parietais. O passo inicial é a produção de ácido carbónico no citoplasma da célula pela reacção de H2O com CO2
catalisada pela enzima anidrase carbónica. A dissociação do ácido carbónico dá origem a iões hidrogénio (H+) e iões
bicarbonato (HCO3-). Na membrana plasmática basal as bombas Na*/K+ estão em intensa actividade deslocando, contra
gradiente de concentração, iões sódio para o exterior da célula e iões potássio para o interior aumentando notavelmente a
concentração de iões potássio no citoplasma, algo que é comum a qualquer outra célula do organismo. A elevada concentração
de ião bicarbonato que se forma no citoplasma da célula activa uma proteína de transporte existente na membrana plasmática
basal que, por antiporte, desloca HCO3- para o exterior, ao mesmo tempo que desloca Cl- para o interior da célula, a energia
para este movimento deve-se ao gradiente de concentração do HCO3- e tem como consequência um aumento da concentração
3
de Cl- no citoplasma da célula . Os iões K+ e os iões Cl- abandonam a célula através da membrana plasmática apical, a favor
5
do gradiente de concentração, para os iões Cl- estão previstos canais electrogénicos com este objectivo . O resultado é um
aumento da concentração de iões Cl- e K+ no lúmen do canalículo intracelular. A actividade da H+/K+ - ATPase colocada na
membrana plasmática apical, desloca contra gradiente de concentração, iões potássio do lúmen do canalículo intracelular para
o citoplasma da célula e iões hidrogénio no sentido oposto. O resultado é uma elevada concentração de H+ e Cl- no lúmen do
3, 5
canalículo intracelular formando ácido clorídrico
Os iões H+ são bombeados contra um elevado gradiente de concentração,
bem evidente quando se observa a diferença de valores de pH, cerca de 7 no citoplasma da célula parietal e cerca de 1 no
5
lúmen da glândula gástrica .
Apicação prática: O omeprazol é uma substância que inibe a boma H+/K+ e o resultado é uma diminuição considerável
5
da secreção ácida, por isso esta e outras drogas similares têm sido utilizadas para o tratamento das úlceras pépticas .
As células parietais também segregam factor intrínseco, uma glicoproteína requerida para absorção da vitamina B12 a
nnível do íleo. Quando as células parietais são destruídas na sequência de gastrite crónica, ocorre perda da função de secreção
de ácido clorídrico e desenvolve-se anemia, dada a importância da vitamina B12 na formação de eritrócitos
5, 6
.
3.5 – Células entero-endócrinas.
O sistema entero-endócrino consiste de células endócrinas dispersas no epitélio da mucosa gastro-intestinal que
segregam aminas biogénicas e polipeptídeos, com efeito endócrino ou parácrino, que são responsáveis pelo controlo da
motilidade e da secreção ao longo do tubo digestivo. No estômago estas células aparecem localizadas na porção mais profunda
da base de todas as glândulas gástricas, entre as células zimogénicas
2, 3
.
Muitas das células entero-endócrinas, mas não todas, captam do meio extracelular percursores de aminas, e promovem
no seu citoplasma a descarboxilação dos aminoácidos (APUD – amine precursor uptake and decarboxylation)
2, 3
e, pela
afinidade que estas células apresentam para os sais de crómio ou prata, são designadas células cromafins ou argentafins, que
não sendo exclusivas do sistema digestivo, fazem parte de uma extensa família de células que constituem o sistema neuroendócrino difuso
2, 3, 9
. Estas células encontradas em vários sistemas do organismo têm a função essencial de libertar
3
mediadores químicos que alteram as funções de outras células , as células deste sistema existentes no tubo digestivo, incluindo
2
o estômago, são designadas células enterocromafins .
As principais células entero-endócrinas existentes no epitélio da mucosa gástrica são fulcrais na regulação da secreção
ácida e enzimática das glândulas gástricas e, essencialmente são três tipos de células: as células G localizadas no antro do
piloro e que produzem um polipeptídeo designado gastrina, as células D existentes nas glândulas fundicas do estômago e
antro do piloro e que produzem somatostina e as células ECL existentes nas glândulas fundicas do estômago, idênticas às
células enterocromafins e produtoras de histamina
9, 10, 11
.
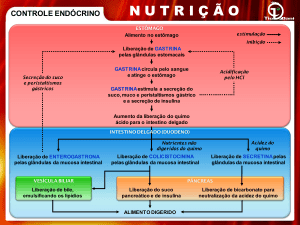
4 – REGULAÇÃO DA SECREÇÃO ÁCIDA E ENZIMÁTICA GÁSTRICA
A secreção de ácido clorídrico pelas células parietais é rigorosamente controlada por uma complexa rede que envolve
factores neuronais, endócrinos e parácrinos
5, 7 8
de ácido clorídrico
7, 8
. A acetilcolina, a gastrina e a histamina são poderosos agonistas da secreção
8
, sendo a estimulação histaminérgica a via mais potente de activação da secreção ácida gástrica . Para
além destes agonistas existe uma substância antagonista que funciona como potente inibidor da secreção ácida, uma
substância designada somatostatina
9, 12
.
4.1 - Histamina
A histamina é um mediador químico que estimula intensamente as células parietais por efeito parácrino, ligando-se a
receptores H2 na membrana plasmática destas células
7, 9
. A histamina é produzida por células idênticas às enterocromafins, as
células ECL (enterocromafin cell like), existentes nas glândulas do fundo do estômago, que captam o aminoácido histidina e
9, 10
promovem a sua descarboxilação para produzir histamina
. A histamina é armazenada em vesículas de secreção e libertada
por exocitose após estimulação das ECL por alguns secretagogos
7, 9
. A ligação da histamina aos receptores H2 das células
9
parietais resulta no aumento da secreção ácida gástrica .
Aplicação prática: Os fármacos antagonistas dos receptores H2, usualmente designados anti-histamínicos, como é o caso da
cimetidina, têm sido utilizados para o tratamento de úlceras pépticas, precisamente porque impedem a actuação estimulante da
9
histamina nas células que sintetizam ácido clorídrico .
10
A estimulação das células ECL para que libertem histamina é feita por vários mediadores químicos . O mediador major é
a gastrina que se liga a resceptores CCK-B na membrana plasmática das células ECL, estimulando estas células a libertar
histamina
9, 10
. A acetilcolina é um neutransmissor libertado por neurónios da componente parassimpática do sistema nevoso
autónomo e constitui um factor que estimula a libertação de histamina pelas células ECL, ligando-se a receptores muscarínicos,
9
no entanto, só 10 a 20% das células ECL são inervadas por fibras nervosas que libertam acetilcolina . Isto explica a fraca
resposta destas células à estimulação pela acetilcolina, como referido por alguns autores. Estes autores referem um outro
neurotransmissor como estimulante das células ECL, esse neurotransmissor é o polipeptídeo pituitário activador da
10
adenilciclase (PACAP) . A epinefrina é um outro mediador químico que actua nos receptores β-adrenérgicos da células ECL
9
estimulando-as .
4.2 - Gastrina
A gastrina é um polipeptídeo sintetizado, armazenado em grânulos de secreção e libertado por exocitose por
células entero-endócrinas designadas células G existentes no antro da região pilórica do estômago, sendo considerada o factor
12
primário que regula a secreção ácida pós prandial .
A gastrina libertada pelas células G entra na circulação sangínea
receptors CCK-B das células alvo
5, 12
10
e tem três principais efeitos pela sua ligação a
. Em primeiro lugar, a gastrina estimula directamente a secreção ácida por se ligar a
receptores CCK-B na membrana plasmática das células parietais
7, 12
. Em segundo lugar, a gastrina estimula indirectamente a
secreção ácida por se ligar a receptores CCK-B na membrana plasmática de células ECL, estimulando a libertação de
histamina que, por sua vez, é o mais forte estimulante da secreção ácida gástrica via receptores H2 na membrana plasmática
7, 10, 12
das células parietais
. Em terceiro lugar, a gastrina actua em receptores CCK-B na membrana plasmática de células D
estimulando-as a libertar somatostatina e este facto, em contraste com os anteriores, levará a uma inibição da secreção ácida
12
pelas células parietais , este efeito será importante para controlar a secreção ácida, para que não se torne excessiva.
A gastrina é libertada mediante estímulos que actuam quer na membrana plasmática apical, quer na membrana
11
plasmática basal das células G . As células G responsáveis pela sua produção apresentam microvilosidades na face apical,
12
voltada para o lúmen da glândula gástrica permitindo a estas células detectar a presença de alimentos no lúmen do estômago .
11
Os estímulos que actuam a este nível são os aminoácidos e aminas presentes nos alimentos ingeridos .
4.3 - Acetilcolina
A acetilcolina é um neurotransmissor libertado pelos terminais axonais dos neurónios da componente parassimpática do
sistema nervoso autónomo que atingem as células da mucosa gástrica via nervo vago e pelos terminais axonais dos neurónios
que constituem o sistema nervoso entérico. A acetilcolina é libertada nas fendas sinápticas entre neurónios e células efectoras e
o seu efeito é mediado pela sua ligação a receptores muscarínicos
5, 12
.
A acetilcolina liga-se a receptores M3 existentes na membrana plasmática das células parietais estimulando-as a
8
produzir ácido clorídrico , liga-se a receptores M1 das células ECL estimulando-as a libertar histamina, o que levará a um
7
aumento da secreção de ácido clorídrico e liga-se a receptores muscarínicos presentes na membrana plasmática das células G
estimulando-as a libertar gastrina
9, 12
, que através das vias directa e indirecta levará à estimulação da secreção ácida.
4.4 - Somatostatina
Para além dos factores agonistas acima descritos, existe um mediador químico com função parácrina, que inibe a
secreção ácida do estômago. Este mediador é a somatostatina produzida por células entero-endócrinas designadas células D
existentes nas glândulas gástricas do fundo e do antro pilórico do estômago
7, 9, 12
. A somatostatina liga-se a receptores S2
existentes na membrana plasmática das células G inibindo a libertação de gastrina, das células ECL inibindo a libertação de
histamina e nas próprias células parietais, o resultado de forma directa ou indirecta é a diminuição da secreção ácida pelas
9
células parietais .
As células D do fundo e do antro pilórico do estômago são estimuladas de forma diferenciada. As células D do antro
9
pilórico são estimuladas a libertar somatostatina pela acidez encontrada no lúmen da glândula gástrica e a somatostatina
proveniente destas células é libertada por prolongamentos citoplasmáticos arredondados em íntimo contacto com as células G,
9, 12
inibindo a libertação de gastrina por estas células
. As células D das glândulas fundicas do estômago são estimuladas a
libertar somatostatina pela gastrina e a somatostatina libertada nestas condições inibe as células ECL próximas, diminuindo
9
assim a libertação de histamina .
A libertação de somatostatina é inibida pela acetilcolina, provavelmente pela ligação desta substância a receptores
9
muscarínicos M2 ou M4 na membrana plasmática das células D .
4.5 – Integração dos diversos factores na regulação da secreção ácida do estômago
Quando o estômago permanece vazio por várias horas, a secreção do ácido clorídrico ocorre a uma taxa basal muito
baixa que representa apenas 10% da taxa máxima de produção de ácido clorídrico. Durante e após a ingestão de alimentos a
taxa de secreção ácida gástrica aumenta imediatamente em três fases sequenciais: a fase cefálica despoletada antes dos
alimentos chegarem ao estômago, a fase gástrica despoletada no momento em que os alimentos chegam ao estômago e a fase
3
intestinal despoletada por mecanismos com origem no intestino delgado .
A fase cefálica é despoletada pela visão, paladar e cheiro dos alimentos e é mediada inteiramente por fibras nervosas da
componente parassimpática do sistema nervoso autónomo que integram o nervo vago. Estas fibras nervosas libertam
essencialmente acetilcolina que estimulam as células G e as células ECL a produzirem gastrina e histamina, respectivamente,
5, 6
estimulando a secreção ácida gástrica, mas também estimula directamente as células parietais a produzirem ácido clorídrico
9
e inibe as células D o que diminuirá a quantidade de somatostatina, tornando mais acentuada a secreção ácida .
A face gástrica é despoletada pela distensão das paredes do estômago, obviamente pela presença de alimento o que
leva ao desencadear de reflexos centrais e locais mediados por neurónios que libertam acetilcolina originando aumento da
secreção de ácido clorídrico. Para além da distensão das paredes do estômago, a presença de amino-ácidos e peptídeos no
lúmen gástrico estimulam as células G através das microvilosidades da membrana plasmática apical voltadas para o lúmen
5, 6, 12
gástrico
e inibem as células D. No entanto, há autores que afirmam que estes estímulos químicos também desencadeiam
reflexos locais mediados por neurónios que libertam acetilcolina, que não só estimulará a libertação de gastrina pelas células G
12
como inibirá a produção de somatostatina pelas células D .
A fase intestinal da secreção ácida gástrica é despoletada pela presença de quimo no duodeno. A distensão da parede
intestinal e a presença de produtos de digestão das proteínas no lúmen intestinal estimulam reflexos via nervo vago que
causam a libertação de acetilcolina que estimulam as células G do antro pilórico a libertar gastrina o que levará a um aumento
5
da secreção ácida gástrica . Por outro lado, a presença de lipídeos no quimo estimulam a libertação de uma hormona
9
designada colecistoquinina que estimula as células D a libertar somatostatina que, por sua vez, inibe a secreção ácida gástrica .
4.6 - Mecanismos celulares de secreção de pepsina pelas células zimogénicas da mucosa gástrica
5, 6
As células zimogénicas ou principais fazem parte do epitélio que reveste as glândulas gástricas e possuem todas as
características de células que sintetizam proteínas. Os grânulos que existem com abundância no seu citoplasma contêm
enzimas sob a forma inactiva, os pepsinogénios. Os pepsinogénios são activados em meio ácido e dão origem às pepsinas,
enzimas proteolíticas cuja actividade enzimática é favorecida pelo meio ácido do conteúdo gástrico.
Muitos dos agentes que estimulam as células parietais estimulam também as células zimogénicas. A acetilcolina e a
gastrina estimulam directamente as células zimogénicas a libertar pepsinogénios. O contacto da mucosa gástrica com o meio
ácido é também um forte estímulo para estas células. A estimulação promovida pelo pH ácido é mediada por reflexos entéricos
da responsabilidade do sistema nervoso entérico. O contacto da mucosa gástrica com o ácido estimula neurónios entéricos que,
por sua vez libertam acetilcolina que estimula as células zimogénicas.
Quando o pH do conteúdo gástrico diminui para valores próximos de 3, o mecanismo da gastrina para
estimulação da secreção gástrica é inibido. A acidez bloqueia a produção de gastrina, por conseguinte bloqueia a secreção
ácida e enzimática da mucosa gástrica. Este feed-back impede a produção excessiva de ácido e enzimas digestivas pela
mucosa gástrica, prevenindo eventuais ulcerações.
4.7 - Mecanismos celulares de secreção de muco e bicarbonato pelas células mucosas superficiais.
As secreções das células mucosas superficiais do epitélio da mucosa gástrica contêm uma glicoproteína designada
mucina que forma o muco que reveste a superfície interna da parede do estômago. O muco é armazenado em grandes
grânulos no polo apical das células mucosas da superfície e libertado por exocitose através da membrana plasmática da face
apical da célula. Dado o carácter viscoso do muco, este forma uma camada gelatinosa na superfície apical das células
epiteliais. As células mucosas da superfície segregam também iões bicarbonato que ficam aprisionados na camada de muco
tornando a sua superfície alcalina. Esta camada de muco alcalino forma a barreira mucosa gástrica, com uma espessura de
cerca de 0,2 milímetros, fundamental para a protecção das paredes do estômago contra o pH extremamente ácido do conteúdo
gástrico. Esta barreira permite que a superfície das células epiteliais se mantenha envolta num pH próximo da neutralidade, em
5
contraste com o pH extremamente ácido do conteúdo do lúmen gástrico .
A barreira mucosa gástrica pressupõe quer a secreção de muco quer a secreção de iões bicarbonato pelas células
mucosas da superfície da mucosa gástrica. Supressão de um ou de outro componente causam destruição desta barreira, por
conseguinte, aumento do risco de úlceras
5, 6
.
O muco é produzido em quantidade significativa, mesmo que não haja alimentos no estômago. No entanto, os mesmos
estímulos que causam aumento da secreção ácida e enzimática da mucosa gástrica, causam também aumento da libertação de
6
muco, sendo a acetilcolina a principal responsável por um aumento substancial da libertação de muco .
Aplicação prática: O ácido acetilsalicílico (aspirina) e outros agentes anti-inflamatórios inibem quer a produção de muco quer
5
de bicarbonato, aumentando a susceptibilidade à formação de úlceras .
A libertação de adrenalina e outras substâncias agonistas adrenérgicas (tal como sucede em situação de stress) inibem a
5
libertação de ião bicarbonato na camada de muco, uma explicação plausível para a patogénese das úlceras por stress . A razão
pela qual isto sucede é a contracção das fibras musculares lisas da túnica média das arteríolas, reduzindo o fluxo de sangue
4
nos capilares da mucosa gástrica, automaticamente, diminui a disponibilidade de iões bicarbonato .
5 – MOTILIDADE GÁSTRICA e ESVAZIAMENTO DO CONTEÚDO GÁSTRICO
Quando o estômago contém alimentos são despoletadas ondas peristálticas de fraca intensidade que empurram os
alimentos em direcção ao antro pilórico. Devido à contracção das fibras musculares que formam o esfíncter pilórico, os
alimentos so empurrados para trás, este tipo de movimento permite misturar os alimentos com as secreções gástricas para a
formação do quimo. Após algumas ondas peristálticas de fraca intensidade ocorrem ondas peristálticas mais intensas. Cada
6
onda peristáltica mais intensa permite que alguns mililitros de quimo passem para o duodeno .
A distensão das paredes do duodeno e a presença de substâncias ácidas em contacto com a mucosa duodenal
desencadeiam reflexos nervosos através do plexo mioentérico que actuam nas paredes do estômago diminuindo o
esvaziamento gástrico.
Bibliografia
1 - Netter FH. Atlas de Anatomia Humana. Segunda Edición. East Hanover. Novartis; 1999.
2 - Williams PL, Warwick R. Gray’s Anatomy. 38th Edition. Edinburgh: Churchill Livingstone; 1995. pp 1683-1815
3 - Junqueira LC, Carneiro J. Histologia Básica. 10ª Edição. Rio de Janeiro: Guanabara Koogan; 2004. pp 284-414.
4 - Kunze WAA, Furness JB. The Enteric Nervous System and Regulation of Intestinal Motility. Annual Review of Physiology. 61:117-142; 1999.
5 - Berne RM, Levy MN. Physiology. 4th Edition. St Louis: Mosby; 1998. pp 589-674.
6 - Guyton AC, Hall JE. Textbook of Medical Physiology. 10th Edition. Philadelphia: WB Saunders Company; 2000. pp 718-833.
7 - Samuelson LC, Hinkle KL. Insights Into the Regulation of Gastric Acid Scretion Through Analysis of Genetically Engineerd Mice. Annual
Review of Physiology. 65:383-400; 2003.
8 - Yao X, Forte JG. Cell Biology of Acid Secretion by the Parietal Cell. Annual Review of Physiology. 65:103-131; 2002.
9 - Sachs G, Zeng N, Prinz C. Physiology of Isolated Gastric Endocrine Cells. Annual Review of Physiology. 59:243-256; 1997.
10 - Prinz C, Zanner R, Gratzl M. Physiology of Gastric Enterochromaffin-Like Cells. Annual Review of Physiology. 65:371-382; 2003.
11 - Dockray GJ. Varro A, Dimaline R, Wang T. The Gastrins. Annual Review of Physiology. 63:119-139; 2001.
12 - Sawada M, Dickinson CJ. The G Cell. Annual Review of Physiology. 59:273-298; 1997.
VII - INTESTINO DELGADO
1 – Aspectos Anatómicos
É um tubo contorcido que se estende do orifício pilórico à válvula ileocecal que o une ao intestino grosso. O seu
comprimento é de 6 a 7 metros e ocupa a porção central e inferior da cavidade abdominal limitado externamente pelo intestino
grosso. Consiste de três partes: duodeno, jejuno e íleo. O duodeno, uma porção curta, cujo comprimento ronda os 20 a 25
centímetros, com um trajecto curvo, com a forma da letra C que circunda a cabeça do pâncreas. O duodeno tem início no piloro
e termina no jejuno. O canal colédoco e o canal pancreático unem-se para formar um curto tubo, denominado ampola
hepatopancreática, que perfura a parede do duodeno para se abrir no lúmen duodenal. O restante intestino delgado é um tubo
longo, preso à parede abdominal posterior por uma prega de peritoneu designada mesentério. Seus dois quintos proximais
constituem o jejuno e os três quintos distais constituem o íleo
1, 2
.
2 – Aspectos Histológicos
2.1 – MUCOSA
O epitélio de revestimento é constituído essencialmente por uma única camada de células cilíndricas, por conseguinte
classificado como epitélio cilíndrico simples. A maior parte das células deste epitélio estão adaptadas à função de absorção dos
nutrientes, água e iões. Estas células, designadas células absortivas ou enterócitos, apresentam na sua face apical,
inúmeras microvilosidades, que são prolongamentos citoplasmáticos que aumentam a área de absorção da célula. Na
superfície das microvilosidades encontram-se enzimas digestivas, nomeadamente dissacaridases e peptidases, fazendo crer
que uma parte da digestão intestinal se faz no local de absorção dos nutrientes. Entre estas células especializadas na
3
absorção, há células caliciformes produtoras de muco .
A mucosa apresenta inúmeras evaginações designadas vilosidades intestinais, elevações digitiformes do epitélio e
tecido conjuntivo subjacente, que se projectam para o lúmen intestinal. Estas projecções da mucosa tem 0,5 a 1mm de altura e
aumentam a área de absorção intestinal. O tecido conjuntivo subjacente ao epitélio, também designado lâmina própria, penetra
nas vilosidades intestinais arrastando consigo vasos sanguíneos, linfáticos e nervos. Cada vilosidade intestinal tem uma porção
central de tecido conjuntivo revestido por epitélio. Neste tecido conjuntivo há um vaso linfático central (vaso quilífero) que se
inicia no ápice da vilosidade e se dirige em direcção à sua base, terminando num plexo de vasos linfáticos da lâmina própria.
2, 3
Paralelamente há vasos sanguíneos que formam um plexo capilar à volta do vaso linfático
.
Entre as vilosidades há inúmeros orifícios onde se abrem glândulas intestinais tubulosas, colocadas profundamente na
mucosa intestinal, designadas glândulas intestinais ou de Lieberkühn que são tubos formados pelo aprofundamento do
epitélio em direcção ao tecido conjuntivo subjacente, constituindo invaginações tubulares que se abrem em orifícios situados na
superfície da mucosa, entre as vilosidades intestinais. O epitélio que forma estas estruturas tubulares é a continuação do
epitélio que reveste a vilosidade, sendo constituído por uma única camada de células colunares ou cilíndricas. Estas células
2, 3
colunares são morfologicamente e funcionalmente distintas entre si
. As células caliciformes produzem muco com acção
3
lubrificante e protectora. As células de Paneth segregam lisozima, uma enzima antibacteriana . As células M são
responsáveis por captar antigénios do lúmen intestinal e colocá-los em contacto com macrófagos e linfócitos, situados em
aglomerados de tecido linfóide no tecido conjuntivo subjacente ao epitélio. Sendo assim, estas células são extremamente
importantes nos processos imunitários. Citologicamente caracterizam-se por invaginações da membrana plasmática do polo
basal que envolvem células de defesa e por prolongamentos citoplasmáticos no polo apical que captam antigénios do conteúdo
intestinal
2, 3
. As células entero-endócrinas produzem inúmeras hormonas entre as quais a colecistoquinina e a secretina, a
primeira promove a secrecção enzimática do pâncreas e a contracção da vesícula biliar, a segunda estimula células secretoras
pancreáticas a produzirem um fluído alcalino, rico em iões bicarbonato, importante para alcalinizar o pH extremamente ácido
3, 4, 5
dos alimentos que saiem do estômago
.
2.2 – CAMADA SUBMUCOSA
É uma camada de tecido conjuntivo frouxo que separa a membrana mucosa da camada muscular. Na submucosa
encontram-se nódulos linfáticos que no íleo formam as designadas placas de Peyer. Aliás, o tecido conjuntivo da mucosa e a
camada submucosa de todo o tubo digestivo apresentam inúmeros nódulos linfáticos, ricos em linfócitos e macrófagos. Estes
aglomerados de tecido linfóide são importantes no desencadeamento de respostas imunitárias e constituem o designado gut
associated linphoide tissue (GALT). Em parceria com as células M do epitélio intestinal, o tecido linfóide existente na parede do
intestino delgado constitui uma barreira defensiva contra micorganismos existentes no lúmen intestinal, sendo responsáveis
3
directa ou indirectamente pela libertação de imunoglobulina A para o lúmen intestinal .
A membrana mucosa e camada submucosa elevam-se para o interior do lúmen intestinal, formando as pregas circulares
(ou válvulas coniventes), visíveis a olho nu. As pregas circulares aumentam a superfície interna do intestino e são mais
2, 3, 5
abundantes na porção proximal do intestino delgado
.
Na camada submucosa do duodeno há um tipo de glândulas, designadas glândulas duodenais ou de Brünner, situadas
na submucosa mas que se abrem as glândulas intestinais ou de Lieberkühn. As glândulas de Brünner produzem uma secrecção
alcalina que protege a mucosa intestinal da acidez do suco gástrico e, conjuntamente com secreções alcalinas pancreáticas,
3
permite o pH ideal para acção das enzimas do suco pancreático .
2.3 – CAMADA MUSCULAR
A camada muscular é constituída por uma subcamada mais interna com fibras com disposição circular e uma subcamada
mais externa com disposição longitudinal.
3 – Aspectos Fisiológicos: Absorção, Secreção Digestão e Motilidade
Bibliografia
1 - Netter FH. Atlas de Anatomia Humana. Segunda Edición. East Hanover. Novartis; 1999.
2 - Williams PL, Warwick R. Gray’s Anatomy. 38th Edition. Edinburgh: Churchill Livingstone; 1995. pp 1683-1815
3 - Junqueira LC, Carneiro J. Histologia Básica. 10ª Edição. Rio de Janeiro: Guanabara Koogan; 2004. pp 284-414.
4 - Berne RM, Levy MN. Physiology. 4th Edition. St Louis: Mosby; 1998. pp 589-674.
5 - Guyton AC, Hall JE. Textbook of Medical Physiology. 10th Edition. Philadelphia: WB Saunders Company; 2000. pp 718-833.
1, 2, 3
VIII - INTESTINO GROSSO
Estende-se da porção final do íleo até ao ânus e circunda o intestino delgado. Tem maior diâmetro do que o intestino
delgado e as fibras musculares longitudinais formam fitas ou faixas cólicas. Como estas fitas são mais curtas do que o estrato
muscular circular, o intestino grosso tem aparência exterior pregueada com saliências designadas haustras.
No intestino grosso não há vilosidades nem pregas circulares. Com excepção do canal anal, a camada mucosa
apresenta um epitélio de revestimento idêntico ao do intestino delgado, apresentando uma única camada de células cilíndricas
com microvilosidades no seu polo apical. Estas células são responsáveis pela principal função do intestino grosso, a absorção
de água e iões do lúmen intestinal e, consequente, formação da matéria fecal. Este epitélio aprofunda-se em direcção ao tecido
conjuntivo subjacente, constituindo as glândulas intestinais ou de Lieberkühn. Nestas glândulas são encontradas células
caliciformes que produzem grande quantidade de muco, no entanto há também algumas células entero-endócrinas. Sob o
epitélio encontra-se a lâmina própria ou tecido conjuntivo subjacente rico em nódulos linfáticos que invadem a submucosa.
Externamente à camada submucosa encontra-se a dupla camada muscular constituída por fibras circulares internamente e
fibras longitudinais externamente.
O intestino grosso começa no lado direito, na porção mais inferior do abdómen, numa porção dilatada designada ceco. O
apêndice vermiforme é um tubo estreito alongado que parte da face póstero-medial do ceco. O ceco tem percurso ascendente
continuando como cólon ascendente até à face inferior do fígado e neste ponto volta-se para a esquerda ao nível da flexura
cólica direita. O íleo desemboca entre o ceco e o cólon ascendente através da válvula ileocecal. Da flexura cólica direita dirigese horizontalmente para a esquerda como cólon transverso. Na região superior e esquerda do abdómen, o cólon transverso
curva-se para baixo, ao nível da flexura cólica esquerda e dirige-se para baixo em direcção à pelve menor como cólon
descendente. Na pelve menor, o intestino grosso forma uma alça sinuosa, o cólon sigmóide. A partir deste ponto continua a
dirigir-se para baixo, junto à parede posterior da pelve menor, designando-se recto, cuja porção inferior se dilata para formar a
ampola rectal. O canal anal começa na extremidade inferior da ampola rectal e termina no ânus. Nesta região o epitélio deixa
de ser cilíndrico simples e passa a ser pavimentoso e estratificado. A porção superior do canal anal apresenta uma série de
pregas internas verticais, as colunas anais, cada uma com uma artéria terminal e uma veia. As extremidades inferiores das
colunas estão unidas por pregas da mucosa, as válvulas anais que no seu conjunto formam a linha pectinada. Abaixo desta
linha há uma zona de transição que termina numa porção revestida internamente por pele. As paredes do canal anal estão
envolvidas por uma espessa camada muscular esfinctérica dividida em esfíncter interno e esfíncter externo do ânus. O esfíncter
anal interno é constituído por fibras musculares lisas e o esfíncter anal externo é formado por fibras musculares estriadas. O
ânus é o orifício distal do tubo digestivo.
Bibliografia
1 - Netter FH. Atlas de Anatomia Humana. Segunda Edición. East Hanover. Novartis; 1999.
2 - Williams PL, Warwick R. Gray’s Anatomy. 38th Edition. Edinburgh: Churchill Livingstone; 1995. pp 1683-1815
3 - Junqueira LC, Carneiro J. Histologia Básica. 10ª Edição. Rio de Janeiro: Guanabara Koogan; 2004. pp 284-414.
IX - PERITONEU
É uma membrana serosa que reveste as paredes internas da cavidade abdominal e reflecte-se nas vísceras
intra-abdominais, envolvendo-as. A parte que reveste as paredes abdominais constitui a lâmina parietal e a parte que envolve
as vísceras constitui a lâmina visceral. As lâminas visceral e parietal estão em contacto e o espaço virtual entre elas constitui a
cavidade peritonial. Pregas de peritoneu permitem ligar as vísceras à parede abdominal. Uma destas pregas é o mesentério
que envolve o intestino delgado, constituído por duas lâminas de peritoneu entre as quais se encontram vasos sanguíneos,
linfáticos e nervos.
X - FÍGADO
O fígado ocupa a porção superior direita da cavidade abdominal. É um órgão extremamente vascularizado,
apresenta lobos direito e esquerdo, sendo o primeiro muito maior que o segundo. Tem uma face superior convexa voltada para
a face inferior do diafragma, a face posterior é sulcada pela veia cava inferior e a face inferior apresenta uma concavidade que
aloja a vesícula biliar
1, 2
.
1 – ESTRUTURA DO PARÊNQUIMA HEPÁTICO
Estruturalmente o fígado é uma estrutura constituída principalmente por células epiteliais, os hepatócitos,
organizados em inúmeros lóbulos hepáticos, envolvidos por bainhas de tecido conjuntivo de suporte e atravessado por
11
inúmeros vasos sanguíneos .
1
Considerando a descrição clássica, cada lóbulo hepático clássico é um pequeno polígono com cerca de 0,7
3
1
milímetros de diâmetro e cerca de 2 milímetros de comprimento , em corte transversal tem contorno hexagonal , delimitado por
septos incompletos de tecido conjuntivo que constituem os espaços porta. Os espaços porta ocupam os ângulos do hexágono
3
e em cada espaço porta aparecem grupos de três estruturas tubulares . Cada um destes grupos designa-se tríade portal, que é
formada por: um ramo da veia porta, um ramo da artéria hepática e um canalículo biliar
1, 3
.
No centro de cada lóbulo há uma pequena veia ou vénula centrolobular da qual irradiam fileiras de
hepatócitos colocados lado a lado do centro para a periferia do lóbulo (lembrando os raios da roda da bicicleta). Entre fileiras
adjacentes de hepatócitos estão colocados capilares sinusóides, os sinusóides hepáticos, resultantes da ramificação dos
ramos da veia porta. Os capilares sinusóides são constituídos por uma camada descontínua de células endoteliais e na face
luminal aparecem macrófagos com morfologia particular, designados células de Kupfer. As células endoteliais estão separadas
1, 3
dos hepatócitos por um espaço designado espaço de Disse
.
As membranas plasmáticas de dois hepatócitos vizinhos, colocados lado a lado, delimitam uma minúscula
estrutura tubular designada canalículo biliar. Em cada lóbulo hepático clássico, os canalículos biliares constituem uma rede
densa convergindo para a periferia do lóbulo para desembocarem em cada canalículo biliar da tríade portal. Este sistema tubular
recolhe a bílis, segregada pelos hepatócitos, que percorre o lóbulo fazendo o trajecto do centro para a periferia
1, 3
.
.
2 – SISTEMA PORTA HEPÁTICO
É um sistema de drenagem de sangue venoso que inclui as veias que recolhem o sangue do pâncreas, do baço,
da vesícula biliar e das porções abdominal e pélvica do tubo digestivo, incluindo a porção abdominal do esófago,. Estas veias
convergem para a veia esplénica, a veia mesentérica superior, as veias gástricas e as veias para-umbilicais e císticas. A
convergência destas veias dá origem à veia porta. Esta veia tem um trajecto de cerca de 8 centímetros, desde a 2ª vértebra
lombar onde tem origem pela união de duas grandes veias, a esplénica e a mesentérica superior, localizando-se a este nível
anteriormente à veia cava inferior e posteriormente ao pâncreas. O percurso da veia porta é ascendente e para a direita,
posteriormente ao duodeno, em direcção ao fígado onde se ramifica várias vezes como uma artéria para dar origem aos
1
sinusóides hepáticos .
Os sinusóides recebem sangue proveniente da veia porta, mas também proveniente da artéria hepática, o
sangue percorre o lóbulo da periferia para o centro onde é drenado para as veias centrolobulares, que por sua vez,
convergem para formar veias de maior calibre que drenam para as veias hepáticas. Estas emergem da face posterior do fígado
1
e após um curtíssimo trajecto, abrem-se na veia cava inferior que sulca a face posterior do fígado .
3 – ARTÉRIA HEPÁTICA
A artéria hepática acompanha a veia porta em direcção ao fígado e penetram juntas neste órgão. Ao lado
destes dois vasos encontra-se o canal hepático comum que deixa o fígado, após ter sido formado pela convergência de vários
pequenos canalículos biliares provenientes dos inúmeros lóbulos hepáticos. A artéria hepática ramifica-se no parênquima
hepático. Um ramo da veia porta, um ramo da artéria hepática e um canalículo biliar, percorrem juntos o mesmo trajecto na
periferia dos lóbulos constituindo a tríade portal. Ramos da artéria hepática dão origem a plexos capilares que drenam para as
veias hepáticas. No entanto, algum sangue arterial entra nos sinusóides hepáticos misturando-se com o sangue proveniente da
veia porta
1, 2
.
4 – A VESÍCULA BILIAR
A vesícula biliar é um pequeno saco piriforme colocado na face inferior do fígado que armazena a bílis
segregada pelos hepatócitos e recolhida por um sistema canalicular que converge para o canal hepático comum que emerge
do fígado. A vesícula biliar tem 7 a 10 centímetros de comprimento e cerca de 3 centímetros de diâmetro na zona mais larga,
1
com uma capacidade que ronda 30 a 50 mililitros .
Da vesícula biliar parte um canal, o canal cístico, que se une ao canal hepático comum, formando o canal
biliar ou canal colédoco. Este une-se ao canal pancreático para formarem a ampola hepatopancreática que perfura a parede
do duodeno desembocando no seu interior. O local onde desemboca a ampola hepatopancreática é marcado por uma saliência
1
2
no interior do duodeno designada papila duodenal (de Vater) e o orifício de passagem para o lúmen duodenal é marcado por
3
um espessamento da camada muscular constituindo o esfínter de Oddi .
5 – FUNÇÔES METABÓLICAS DO FÍGADO:
1
O fígado é essencial para a sobreviivência do indivíduo sendo responsável por inúmeras actividades metabólicas . Os
hepatócitos têm elevada taxa metabólica, participando activamente no metabolismo glicídico, lipídico e proteico, bem como
4,5,6
numa miscelânia de outras funções metabólicas
.
Metabolismo Glicídico:
Os hepatócitos são responsáveis pela manutenção da normoglicemia. Em jejum, a concentração de glicose no plasma
considerada normal (normoglicemia) varia, entre 60 a 110 mg/dl
7, 8, 9
. Valores de glicemia acima de 110 mg/dl definem
8, 9
hiperglicemia se eses valores forem superiores a 126 mg/dl permitem diagnosticar diabetes mellitus
.
O fígado é um órgão efector de extrema importância na manutenção da normoglicemia, armazenando e libertando
glicose sempre que a glicemia aumenta ou diminui. Nos hepatócitos há uma série de complexos enzimáticos que permitem ao
4
hepatócitos armazenar o excedente de glicose ou libertar glicose, consoante as necessidades do organismo . Desta forma os
hepatócitos conseguem manter a glicemia dentro de estreitos limites,
Quando aumenta a glicemia, os hepatócitos removem a glicose do plasma que percorre os sinusóides hepáticos,
aumentando a concentração de glicose no citoplasma dos hepatócitos. Nestas circunstâncias, uma forma de armazenamento do
4,5
excedente de glicose é sob a forma de glicogénio, um polissacarídeo sintetizado a partir das moléculas de glicose , através de
6
um processo bioquímico designado glicogénese .
5, 6
Se diminuir a glicemia os hepatócitos degradam o glicogénio, um processo bioquímico designado glicogenólise
,a
6
capacidade de libertar glicose, ao contrário do que sucede noutras células, permite aos hepatócitos elevar a glicemia . O fígado
5
é o principal órgão onde ocorre gliconeogénese , um processo bioquímico inerente ao metabolismo glicídico que consiste na
6
conversão de moléculas não glicídicas em glicose ou glicogénio . Os hepatócitos utilizam o lactato, o glicerol e alguns
aminoácidos como substratos para o processo da gliconeogénese, convertendo-os em moléculas de glicose, este processo
6,7
permite aumentar a glicemia em situação de hipoglicemia .
Metabolismo Lipídico:
Embora muitas células do organismo tenham capacidade de metabolizar os lipídeos, certos aspectos do metabolismo
4
lipídico ocorrem principalmente nos hepatócitos . Um destes aspectos é a síntese de ácidos gordos a partir de acetil-coenzima
A. Em situação de hiperglicemia, o excedente de glicose captada pelos hepatócitos é convertida em acetilcoenzima A pelo
processo de glicólise. As moléculas de acetilcoenzima A são utilizadas para sintetizar ácidos gordos através de um mecanismo
bioquímico complexo que tem início na carboxilação da acetilcoenzima A convertendo-a em malonilcoenzima A. A adição de
acetilcoenzima A à malonilcoenzima A dá origem a um composto que sofre descarboxilação. A partir daqui são construídos os
6
ácidos gordos pela adição de novas moléculas de malonilcoenzima A seguida de descarboxilação . A combinação de ácidos
4
gordos com moléculas de glicerol permite a síntese de triglicerídeos .
As moléculas de acetilcoenzima A são também utilizadas pelos hepatócitos para sintetizar colesterol, 80% do qual é
4
utilizado na síntese de ácidos biliares . Os triglicerídeos e o colesterol são exportados para tecidos extra-hepáticos, sendo
6,7
transportados no plasma em complexos lipoproteicos de densidade variável (VLDL; LDL; HDL) sintetizados no fígado .
Quando é necessário disponibilizar substrato alternativo para as células produzirem ATP, os ácidos gordos são
clivados em segmentos de dois átomos de carbono, dando origem a moléculas de acetilcoenzima A, um processo bioquímico
4,6
mitocondrial designado β-oxidação . Embora este processo possa ocorrer noutras células do organismo, os hepatócitos são
4
particularmernte eficientes a executar a β-oxidação , dando origem a corpos cetónicos pela condensação de dois radicais acetil.
4,6
Os corpos cetónicos são exportados para tecidos extra-hepáticos para serem utilizados como substrato na produção de ATP .
Metabolismo das proteínas:
Praticamente todas as proteínas plasmáticas com excepção das imunoglobulinas são sintetizadas pelos hepatócitos. O
processo de desaminação dos aminoácidos é também uma das funções dos hepatócitos, inerente ao metabolismo proteico, um
processo obrigatório para que os aminoácidos possam ser utilizados como substrato para a produção de ATP ou para serem
4
convertidos em glicose . A amónia obtida no processo de desaminação é eliminada através da ureia, uma molécula inócua
4,5
formada nos hepatócitos .
Outras funções metabólicas do fígado:
Os hepatócitos inactivam substâncias endógenas e exógenas potencialmente tóxicas, nomeadamente alguns
fármacos, armazenam ião ferro sob a forma de ferritina e vitaminas e sintetizam factores que participam na cascata de
4
coagulação .
A bílis é segregada pelos hepatócitos nos canalículos biliares. É constituída por ácidos biliares, colesterol, fosfolipídeos
e pigmentos biliares. Os sais biliares são sintetizados a partir das moléculas de colesterol e normalmente são conjugados com
+
os aminoácidos taurina e glicina e em pH neutro ligam-se a iões, nomeadamente Na dando origem a sais biliares. Os ácidos
biliares são moléculas anfipáticas com domínios hidrofóbico e hidrofílico. Devido ao carácter anfipático, quando em solução, os
ácidos biliares formam agregados moleculares designados micelas. Dadas estas características, os ácidos biliares funcionam
como agentes de emulsão dos lipídeos, fragmentam as gotas de lipídeos em minúsculas gotículas, aumentando a área de
exposição às enzimas lipolíticas. Este aspecto é fulcral na digestão dos lipídeos. As micelas transportam os produtos de
digestão lipídica até às microvilosidades das células epiteliais do intestino delgado. Grande parte dos sais biliares libertados no
lúmen intestinal, são absorvidos pelo epitélio intestinal e regressam aos hepatócitos pelo sistema porta-hepático para voltarem a
ser segregados na bílis, apenas 10 a 20% dos sais biliares são excretados nas fezes. Esta circulação dos sais biliares é
5
conhecida como circulação entero-hepática .
Aplicação de conhecimentos: A utilização de fármacos que inibem a absorção de ácidos biliares permite aumentar a
5
excreção de colesterol nas fezes e, consequentemente reduzir os níveis de colesterol plasmático . Tem sido
demonstrado por estudos epidemiológicos que a ingestão de alimentos ricos em fibra, nomeadamente cereais como a
aveia ricos em β-glucan, fazem baixar os níveis de colesterol plasmático, por inibirem não só a absorção de colesterol da
10
dieta, mas também a absorção de sais biliares, pelas células epiteliais do intestino delgado .
Os hepatócitos são responsáveis por colocar na bílis substâncias que precisam de ser eliminadas, desta forma, essas
substâncias são excretadas nas fezes. Uma destas substâncias é um pigmento amarelo esverdeado designado bilirrubina
resultante da degradação da hemoglobina. Quando os eritrócitos envelhecidos são destruídos, a hemoglobina é degradada. O
anel porfirínico do grupo heme é clivado originando a bilirrubina que é libertada no plasma. Os hepatócitos absorvem a
bilirrubina em circulação e conjugam a bilirrubina com ácido glucorónico ou com sulfato. A bilirrubina conjugada é então
4
segregada na bílis pelos hepatócitos, para ser excretada através das fezes .
Aplicação de conhecimentos: A medição da concentração de bilirrubina no plasma é utilizada como técnica de
diagnóstico. No caso de intensa destruição dos eritrócitos, a concentração plasmática de bilirrubina não conjugada
aumenta. Em caso de lesão dos hepatócitos (hepatite), há dificuldade de colocar a bílis no intestino delgado, logo a
4
bilirrubina conjugada acumula-se no organismo, aumentando a sua concentração no plasma .
Bibliografia:
1 - Williams PL, Warwick R. Gray’s Anatomy. 38th Edition. Edinburgh: Churchill Livingstone; 1995. pp 1683-1815
2 - Netter FH. Atlas de Anatomia Humana. Segunda Edición. East Hanover. Novartis; 1999.
3 - Junqueira LC, Carneiro J. Histologia Básica. 10ª Edição. Rio de Janeiro: Guanabara Koogan; 2004. pp 284-338.
4 - Guyton AC, Hall JE. Textbook of Medical Physiology. 10th Edition. Philadelphia: WB Saunders Company; 2000. pp 718-833.
5 - Berne RM, Levy MN. Physiology. 4th Edition. St Louis: Mosby; 1998. pp 589-674.
6 - Murray RK, Granner DK, Mayes PA, Rodwell VW. Harper’s Illustrated Biochemistry. 26th Edition. New York: Lange McGraw Hill; 2000. pp 145-218.
7
- Berne RM, Levy MN. Physiology. 4th Edition. St Louis: Mosby; 1998. pp 800-821.
8
- The Expert Committee on the Diagnosis and Classification of Diabetes Mellitus. Report of the Expert Committee on the Diagnosis and Classification
of Diabetes Mellitus. Diabetes Care ( 2001 ). 24 Suppl. 1, S5 – S20.
9 - American Diabetes Association. Screening for Diabetes. Diabetes Care ( 2001 ). 24 Suppl. 1, S21 – S24.
10- Queenan KM, Stewart ML, Smith KN, Thomas W Fulcher RG, Slavin JL. Concentrated oat β-glucan, a fermentable fiber, lowers serum cholesterol in
hypercholesterolemic adults in a randomized controlled trial. Nutrition Journal 2007; 6:6.
XI - PÂNCREAS
1 – Aspectos Anatómicos e Histológicos
O pâncreas é uma glândula ímpar, colocada horizontalmente na parede posterior da cavidade abdominal,
posteriormente ao estômago e com cerca de 12 a 15 centímetros de comprimento
18, 13
. É mais largo na sua extremidade
direita, esta porção é designada a cabeça do pâncreas e está encaixada na curvatura do duodeno (o primeiro segmento do
intestino delgado). A porção restante da glândula é mais estreita mas mais longa do que a cabeça, constitui o corpo do
pâncreas, terminando na cauda do pâncreas, uma porção ainda mais estreita na extremidade esquerda da glândula,
anatomicamente relacionada com o baço
1, 2
.
Histologicamente o pâncreas é constituído por dois tipos de tecido glandular, a maior quantidade de tecido
são células glandulares exócrinas entre as quais aparecem pequenos grupos de células endócrinas constituindo os ilhéus
pancreáticos ou de Langerhans
2, 3
.
A porção exócrina do pâncreas está organizada em túbulos microscópicos cujo fundo cego está rodeado por
células poligonais, os ácinos. As secreções produzidas pelas células acinares são recolhidas nos canais intercalados que
drenam para canais mais largos designados canais intralobulares, drenando para canais extralobulares que convergem
4
para um canal excretor único . O canal excretor pancreático une-se ao canal colédoco para constituírem a ampola
hepatopancreática que desemboca no duodeno
1, 2
.
2 – Aspectos Fisiológicos
O suco pancreático é composto por duas componentes, uma componente aquosa e uma componente
enzimática. A componente aquosa é produzida pelas células epiteliais que constituem a parede dos canais excretores
-
pancreáticos, é um fluído isotónico em relação ao plasma com elevada concentração de HCO3 , tanto mais elevada quanto
maior a taxa de secreção, fortemente estimulada pela secretina. A componente enzimática é sintetizada e libertada pelas
células acinares e contém enzimas digestivas: proteases (tripsina, quimotripsina e carboxipeptidase), amilases e lipases
(triacilglicerol hidrolase, colesterol hidrolase e fosfolipase
4, 5
.
Bibliografia:
1 - Williams PL, Warwick R. Gray’s Anatomy. 38th Edition. Edinburgh: Churchill Livingstone; 1995. pp 1683-1815
2 - Netter FH. Atlas de Anatomia Humana. Segunda Edición. East Hanover. Novartis; 1999.
3 - Junqueira LC, Carneiro J. Histologia Básica. 10ª Edição. Rio de Janeiro: Guanabara Koogan; 2004. pp 284-338.
4 - Berne RM, Levy MN. Physiology. 4th Edition. St Louis: Mosby; 1998. pp 589-674.
5 - Guyton AC, Hall JE. Textbook of Medical Physiology. 10th Edition. Philadelphia: WB Saunders Company; 2000. pp 718-833.
Bibliografia (ordem alfabética):
1.
Atherton JC. The Pathogenesis of Helicobacter Pylori- Induced Gastroduodenal Diseases. Annual Review of Pathology: Mechanisms of Disease.
1:63-93; 2006.
2.
Barret KE, Keely SJ. Chloride Secretion by the Intestinal Epithelium: Molecular Basis and Regulatory Aspects. Annual Review of Physiology.
62:535-572; 2000.
3.
Berne RM, Levy MN. Physiology. 4th Edition. St Louis: Mosby; 1998. pp 589-674
4.
Daniel H. Molecular and Integrative Physiology of Intestinal Peptide Transport. Annual Review of Physiology. 66:361-384; 2004.
5.
Dockray GJ. Varro A, Dimaline R, Wang T. The Gastrins. Annual Review of Physiology. 63:119-139; 2001.
6.
Geibel JP. Secretion and Absortion by Colonic Crypts. Annual Review of Physiology. 67:471-490; 2005.
7.
Guyton AC, Hall JE. Textbook of Medical Physiology. 10th Edition. Philadelphia: WB Saunders Company; 2000. pp 718-833
8.
Junqueira LC, Carneiro J. Histologia Básica. 10ª Edição. Rio de Janeiro: Guanabara Koogan; 2004. pp 284-414.
9.
Kierszenbaum AL. Histologia e Biologia Celular. Uma Introdução à Patologia. Rio de Janeiro: Elsevier 2004.
10. Kunze WAA, Furness JB. The Enteric Nervous System and Regulation of Intestinal Motility. Annual Review of Physiology. 61:117-142; 1999.
11. Liddle RA. Cholecystokinine Cells. Annual Review of Physiology. 59:221-242; 1997.
12. Melvin JE. Yule D, Shuttleworth T, Begenisich T. Regulation of Fluid and Electrolyte Secretion in Salivary Gland Acinar Cells. Annual Review of
Physiology. 67:445-469; 2005.
13. Netter FH. Atlas de Anatomia Humana. Segunda Edición. East Hanover. Novartis; 1999.
14. Prinz C, Zanner R, Gratzl M. Physiology of Gastric Enterochromaffin-Like Cells. Annual Review of Physiology. 65:371-382; 2003.
15. Sachs G, Zeng N, Prinz C. Physiology of Isolated Gastric Endocrine Cells. Annual Review of Physiology. 59:243-256; 1997.
16. Samuelson LC, Hinkle KL. Insights Into the Regulation of Gastric Acid Scretion Through Analysis of Genetically Engineerd Mice. Annual Review of
Physiology. 65:383-400; 2003.
17. Sawada M, Dickinson CJ. The G Cell. Annual Review of Physiology. 59:273-298; 1997.
18. Williams PL, Warwick R. Gray’s Anatomy. 38th Edition. Edinburgh: Churchill Livingstone; 1995. pp 1683-1815
19. Williams PL, Warwick R. Gray’s Anatomy. 38th Edition. Edinburgh: Churchill Livingstone; 1995. pp 1504 -1605.
20. Yao X, Forte JG. Cell Biology of Acid Secretion by the Parietal Cell. Annual Review of Physiology. 65:103-131; 2002.