Glicólise e desidrogénase do piruvato; Rui Fontes
Glicólise e desidrogénase do piruvato
Índice
1O catabolismo dos nutrientes fornece energia para a síntese de ATP ................................................................ 1
2A glicólise, sua localização e papel metabólico ................................................................................................. 1
3O transporte transmembranar de glicose ............................................................................................................ 1
4As enzimas da glicólise catalisam a conversão da glicose em piruvato ............................................................. 2
5A ação da aldólase permite compreender que a partir de uma molécula de glicose se formem duas de piruvato3
6A energia libertada na cisão e na oxidação de uma molécula de glicose por duas moléculas de NAD+ permite a
formação líquida de duas moléculas de ATP ...................................................................................................................... 3
7Nos eritrócitos, todo o NADH formado na ação da desidrogénase do gliceraldeído-3-fosfato é reoxidado por
ação da desidrogénase do lactato ........................................................................................................................................ 3
8Nos eritrócitos, a glicose é cindida a lactato e o processo (que não é uma oxidação) designa-se por glicólise
anaeróbica ........................................................................................................................................................................... 4
9O destino do lactato é a sua reoxidação a piruvato e ocorre em células distintas das que o produzem ............. 4
10Quando o NADH é reoxidado na cadeia respiratória (pelo oxigénio) a glicólise diz-se aeróbica: uma molécula
de glicose é oxidada gerando duas de piruvato ................................................................................................................... 4
11A desidrogénase do piruvato catalisa a descarboxilação oxidativa do piruvato gerando acetil-CoA ................ 4
12A inativação da desidrogénase do piruvato pela cínase da desidrogénase do piruvato e a sua ativação pela
fosfátase da desidrogénase do piruvato. .............................................................................................................................. 5
13Em algumas células do organismo a glicólise é, normalmente, predominantemente aeróbica e noutras
predominantemente anaeróbica .......................................................................................................................................... 5
14A glicólise anaeróbica permite manter a concentração de ATP estacionária nas fibras musculares aquando do
exercício físico anaeróbico ................................................................................................................................................. 6
15Durante o exercício há mobilização de GLUT4 para a membrana sarcoplasmática e ativação da desidrogénase
do piruvato .......................................................................................................................................................................... 6
16As concentrações citoplasmáticas de AMP e ADP podem subir marcadamente durante o exercício físico e
ativar a glicólise .................................................................................................................................................................. 7
17A estimulação da captação celular de glicose e da oxidação da glicose no estado pós-prandial ....................... 7
18Mecanismos envolvidos na estimulação da captação e da oxidação muscular de glicose no estado pós-prandial
8
19Mecanismos envolvidos na estimulação da captação e da oxidação hepática de glicose no estado pós-prandial8
20A oxidação de glicose pelo cérebro ................................................................................................................... 8
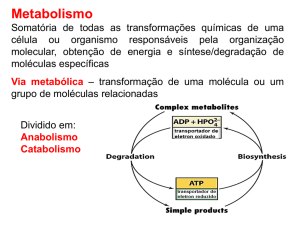
1- O catabolismo dos nutrientes fornece energia para a síntese de ATP
O metabolismo energético dos seres vivos pode ser interpretado como um processo no qual os
nutrientes são oxidados pelo O2 (catabolismo) gerando a energia necessária para a síntese de ATP (a
partir de ADP + Pi), ao mesmo tempo que o ATP formado é hidrolisado (a ADP + Pi) fornecendo a
energia utilizada na síntese de múltiplas substâncias (anabolismo), na atividade muscular e na
manutenção dos gradientes iónicos.
Juntamente com outros mecanismos, a fome é um importante mecanismo homeostático: a
ingestão de alimentos fornece ao organismo as substâncias cuja oxidação permite manter a concentração
celular de ATP praticamente constante (“estacionária”). Numa determinada célula do organismo a
concentração de ATP praticamente não varia, mesmo quando a sua velocidade de hidrólise aumenta
porque, quando isto acontece (por exemplo, nas fibras musculares, durante o esforço físico), aumenta
igualmente a sua velocidade de síntese (formação de ATP a partir de ADP e Pi). Este aumento da
velocidade de síntese de ATP implica um aumento da velocidade do catabolismo: um aumento na
velocidade de oxidação dos nutrientes.
2- A glicólise, sua localização e papel metabólico
A glicólise é uma via metabólica do citoplasma das células (de todas as células) em que a glicose
é, num processo exergónico, convertida em piruvato (ou lactato) e se forma, concomitantemente e num
processo endergónico, ATP a partir de ADP + Pi. As enzimas da glicólise são, no seu conjunto, “a
máquina” que permite a acoplagem dos dois processos.
3- O transporte transmembranar de glicose
Na maioria das células do organismo a entrada da glicose do sangue para o citoplasma dá-se a
favor do gradiente e o transporte é catalisado por proteínas da membrana que são uniporters. Estes
Página 1 de 9
Glicólise e desidrogénase do piruvato; Rui Fontes
uniporters designam-se por GLUTs e podem ser produtos de genes distintos em órgãos distintos. Nos
casos do polo basal dos enterócitos e dos hepatócitos a concentração de glicose pode, em certas
condições, ser mais alta no citoplasma que no sangue: nestas condições o GLUT (nestes casos, o GLUT2)
catalisa a saída de glicose da célula. Noutras condições ocorre o inverso e a glicose entra para estas
células.
No caso dos músculos, porque toda a glicose que entra é imediatamente fosforilada a glicose-6fosfato (catálise pela hexocínase II1; ver Equação 1) e não existem, aqui, enzimas capazes de converter a
glicose-6-fosfato em glicose, o gradiente de concentrações favorece sempre a entrada de glicose. Nos
músculos, o transporte transmembranar de glicose é catalisado pelo GLUT4 mas o número de
transportadores na membrana sarcoplasmática de uma fibra muscular varia consoante o estado hormonal e
a atividade contráctil da fibra muscular. O número de moléculas de GLUT4 na membrana
sarcoplasmática e, consequentemente, a velocidade com que uma fibra muscular capta a glicose do
sangue aumenta quando aumenta a concentração de insulina no sangue ou quando aumenta a
atividade contráctil da fibra muscular em questão [1]. Ambos os estímulos promovem a migração para
a membrana sarcoplasmática de vesículas que estavam no citoplasma e que contêm GLUT4: as vesículas
fundem-se com a membrana que incorpora as moléculas de GLUT4 na sua estrutura.
Equação 1
glicose + ATP → glicose-6-fosfato + ADP
No caso dos eritrócitos e dos neurónios os transportadores de glicose designam-se,
respetivamente, por GLUT1 e GLUT3.
4- As enzimas da glicólise catalisam a conversão da glicose em piruvato
As enzimas da glicólise são: a cínase da glicose ou a cínase das hexoses (ver nota 1 e Equação
1), a isomérase das fosfohexoses (ver Equação 2), a cínase-1 da frutose-6-fosfato (ver Equação 3), a
aldólase (ver Equação 4), a isomérase das trioses-fosfato (ver Equação 5), a desidrogénase do
gliceraldeído-3-fosfato (ver Equação 6), a cínase do 3-fosfoglicerato (ver Equação 7), a mútase do
fosfoglicerato (ver Equação 8), a enólase (ver Equação 9) e a cínase do piruvato (ver Equação 10).
Equação 2
Equação 3
Equação 4
Equação 5
Equação 6
Equação 7
Equação 8
Equação 9
Equação 10
glicose-6-fosfato ↔ frutose-6-fosfato
frutose-6-fosfato + ATP → frutose-1,6-bisfosfato + ADP
frutose-1,6-bisfosfato ↔ gliceraldeído-3-fosfato + dihidroxiacetona-fosfato
dihidroxiacetona-fosfato ↔ gliceraldeído-3-fosfato
gliceraldeído-3-fosfato + Pi + NAD+ ↔ 1,3-bisfosfoglicerato + NADH
1,3-bisfosfoglicerato + ADP ↔ 3-fosfoglicerato + ATP
3-fosfoglicerato ↔ 2-fosfoglicerato
2-fosfoglicerato ↔ fosfoenolpiruvato + H2O
fosfoenolpiruvato + ADP → piruvato + ATP
Nas membranas citoplasmáticas não existem transportadores para os intermediários fosforilados
da glicólise: por isso a ação catalítica das hexocínases (ver Equação 1) converte uma substância que pode
atravessar a membrana (a glicose, via GLUTs) numa outra (a glicose-6-fosfato) que já não tem essa
característica.
Tal como em todas as vias metabólicas, um dos produtos formados pela ação catalítica de uma
dada enzima é substrato de uma outra enzima permitindo compreender que se possa desenhar uma cadeia
sequenciada de reações e que, no seu conjunto, as enzimas da glicólise convertam glicose em piruvato. Se
ignorarmos momentaneamente a formação de NADH e o consumo e formação de ATP poderemos
escrever a seguinte sequência de transformações: glicose → glicose-6-fosfato → frutose-6-fosfato →
frutose-1,6-bisfosfato → (dihidroxiacetona-fosfato + gliceraldeído-3-fosfato); dihidroxiacetona-fosfato →
1
O reconhecimento recente da estrutura primária das diferentes isoenzimas que, nos mamíferos, são capazes de catalisar
a fosforilação da glicose motivou uma mudança na nomenclatura das isoenzimas com esta atividade catalítica. Os estudos
funcionais já haviam demonstrado, há mais de 50 anos, que havia, pelo menos, duas isoenzimas com esta atividade que
foram, então, designadas por cínase da glicose e por cínase das hexoses. Atualmente a cínase da glicose também é
designada por hexocínase IV. A atividade da cínase das hexoses é, sabe-se hoje, partilhada por três hexocínases
diferentes designadas por hexocínases I, II e III.
Página 2 de 9
Glicólise e desidrogénase do piruvato; Rui Fontes
gliceraldeído-3-fosfato → 1,3-bisfosfoglicerato
fosfoenolpiruvato → piruvato.
→
3-fosfoglicerato
→
2-fosfoglicerato
→
5- A ação da aldólase permite compreender que a partir de uma molécula de glicose se
formem duas de piruvato
A aldólase (ver Equação 4) é a enzima da glicólise que catalisa a cisão (lise) da molécula de
frutose-1,6-bisfosfato (6C) em gliceraldeído-3-fosfato (3C) e dihidroxiacetona-fosfato (3C). A
transformação da dihidroxiacetona-fosfato em gliceraldeído-3-fosfato (ver Equação 5) e a posterior
transformação deste composto em piruvato (3C; ver Equações 6-10) permite compreender que uma
molécula de glicose (6C) dê origem a duas de piruvato (2 × 3C).
6- A energia libertada na cisão e na oxidação de uma molécula de glicose por duas moléculas
de NAD+ permite a formação líquida de duas moléculas de ATP
Antes da lise da frutose-1,6-bisfosfato, duas moléculas de ATP servem como substratos dadores
de fosfato em reações de fosfotransferência catalisadas pelas cínases da glicose e da frutose-6-fosfato
(ver Equação 1 e Equação 3). Depois da lise da frutose-1,6-bisfosfato ocorre a formação de ATP em duas
reações de fosfotransferência em que o substrato aceitador é o ADP e os substratos dadores são o 1,3bisfosfoglicerato (cínase do 3-fosfoglicerato; ver Equação 7) e o fosfoenolpiruvato (cínase do piruvato;
ver Equação 10). As reações de formação de ATP a partir de ADP através de reações de
fosfotransferência designam-se, comummente, de “fosforilações ao nível do substrato” (por contraponto
com à “fosforilação oxidativa” mitocondrial).
Também já depois da lise da frutose 1,6-bisfosfato ocorre a redução do NAD+ (e consequente
formação do NADH) numa reação em que, concomitantemente com a reação de oxirredução ocorre a
adição de fosfato inorgânico (Pi): esta reação é a catalisada pela desidrogénase do gliceraldeído-3-fosfato
(ver Equação 6). O NAD+ e o NADH são, respetivamente, as formas oxidada e reduzida do dinucleotídeo
de adenina e nicotinamida.
O somatório das reações envolvidas na formação do piruvato a partir de glicose pode ser expressa
pela Equação 11:
Equação 11
glicose (C6H12O6) + 2NAD+ + 2ADP + 2Pi →
2 ác. pirúvico (C3H4O3) + 2NADH + 2ATP + 2H2O
A energia libertada no processo de cisão e oxidação de um mole de glicose por dois moles de
NAD+ permite a formação líquida de 2 moles de ATP (a partir de ADP e Pi). O processo oxidativo
envolve dois pares de eletrões que são cedidos pela glicose (que se oxida a piruvato) e aceites por dois
NAD+ (que se reduzem a NADH).
7- Nos eritrócitos, todo o NADH formado na ação da desidrogénase do gliceraldeído-3-fosfato
é reoxidado por ação da desidrogénase do lactato
Os eritrócitos são as células mais simples do organismo. Porque não têm mitocôndrias, todo o
ATP que hidrolisam (por exemplo, na ação da ATPase do Na+/K+) formando ADP e Pi é reposto na
transformação expressa pela Equação 11, ou seja, na glicólise. Existe aqui, tal como em todas as células
do organismo, um ciclo de formação e hidrólise de ATP onde as concentrações de ATP, ADP e Pi se
mantêm estacionárias.
A concentração de NAD+ (e NADH) dentro dos eritrócitos é (como em todas as células) muito
baixa (cerca de 78 µM [2]) e, na ausência de um mecanismo que permitisse reoxidar o NADH a NAD+,
todo o NAD+ do eritrócito se esgotaria em menos de 1 minuto. De facto, a concentração de NAD+ (e
NADH) é estacionária porque cada molécula de NADH que se forma na glicólise é imediatamente
oxidada a NAD+ por ação catalítica da desidrogénase do lactato (ver Equação 12).
Equação 12
2 NADH + 2 ácido pirúvico ↔ 2 NAD+ + 2 ácido láctico
Página 3 de 9
Glicólise e desidrogénase do piruvato; Rui Fontes
8- Nos eritrócitos, a glicose é cindida a lactato e o processo (que não é uma oxidação) designase por glicólise anaeróbica
O somatório da Equação 11 e da Equação 12 e de uma outra (Equação 13) que representa todos
os processos que, no eritrócito, hidrolisam ATP à mesma velocidade com que se forma permitem escrever
a Equação 14.
Equação 13
Equação 14
2 ATP + 2 H2O → 2 ADP + 2 Pi
glicose (C6H12O6) → 2 ácido láctico (C3H6O3)
A Equação 14 resume uma ideia importante: os eritrócitos captam glicose do plasma (via ação do
GLUT1) e convertem-na em ácido láctico que é libertado para o plasma. (Este ácido láctico vai depois ser
metabolizado por outras células do organismo.) À glicólise que tem como produto o ácido láctico (=
lactato + protão) chama-se glicólise anaeróbica porque não consome oxigénio.
A Equação 14 mostra que o processo não é globalmente oxidativo: o número de oxidação médio
dos carbonos da glicose e do ácido láctico são iguais (em ambos os casos, 0).
9- O destino do lactato é a sua reoxidação a piruvato e ocorre em células distintas das que o
produzem
Nos mamíferos, a única reação em que o ácido láctico é substrato é a que é catalisada pela
desidrogénase do lactato (Equação 12), uma enzima que catalisa uma reação fisiologicamente reversível.
Nos órgãos onde o ácido láctico é captado (fígado e coração, por exemplo) a única reação em que o ácido
láctico intervém é a que é expressa pela Equação 12 mas, agora, desenvolve-se no sentido de formar ácido
pirúvico que será depois oxidado nas mitocôndrias desses órgãos ou, eventualmente, sofrer outras
transformações (como, no fígado, a conversão em glicose).
O transporte transmembranar de lactato é mediado por um simporter (designado por
transportador de monocarboxilatos) que faz o cotransporte do anião lactato com um protão (ou seja,
transporta ácido láctico) a favor do gradiente. O mesmo transportador também pode transportar ácido
pirúvico [3].
10- Quando o NADH é reoxidado na cadeia respiratória (pelo oxigénio) a glicólise diz-se
aeróbica: uma molécula de glicose é oxidada gerando duas de piruvato
Ao contrário dos eritrócitos todas as outras células têm mitocôndrias e, em condições “normais”,
o ácido láctico que vertem para o sangue é relativamente escasso. Nestes casos, cada molécula de NADH
que se forma durante a glicólise é imediatamente oxidada (ver Equação 15), não por ação da
desidrogénase do lactato, mas sim pelo O2 na cadeia respiratória das mitocôndrias:
Equação 15
2 NADH + O2 → 2 NAD+ + 2 H2O
Desta forma as concentrações de NADH são mantidas baixas na célula ao mesmo tempo que o
NAD+ é regenerado. O somatório da Equação 11 e da Equação 15 é a Equação 16 que expressa a ação
conjugada das enzimas da glicólise e da cadeia respiratória, ou seja, a glicólise aeróbica:
Equação 16
glicose (C6H12O6) + O2 + 2 ADP + 2 Pi → 2 ácido pirúvico (C3H4O3) + 2 ATP + 2 H2O
11- A desidrogénase do piruvato catalisa a descarboxilação oxidativa do piruvato gerando
acetil-CoA
Em condições geralmente designadas por aeróbicas, o ácido pirúvico entra para as mitocôndrias
onde, por ação catalítica da desidrogénase do piruvato se converte em acetil-CoA. A reação catalisada
pela desidrogénase do piruvato é fisiologicamente irreversível e é descrita pela Equação 17:
Equação 17
piruvato + CoA + NAD+ → acetil-CoA + CO2 + NADH
O processo catalítico é frequentemente descrito como uma “oxidação descarboxilativa” porque, a
par de uma reação de oxi-redução, ocorre uma descarboxilação (saída de CO2) de tal forma que o número
de carbonos do resíduo acetilo da acetil-CoA (dois) é menor que o do piruvato (três). Permitindo a
oxidação das novas moléculas de ácido pirúvico formadas durante a oxidação da glicose, cada molécula
Página 4 de 9
Glicólise e desidrogénase do piruvato; Rui Fontes
+
de NADH formada é imediatamente oxidada a NAD pelo O2 na cadeia respiratória (ver Equação 15). O
resíduo acetilo do acetil-CoA é oxidado a CO2 no ciclo de Krebs e neste processo também se formam
ATPs.
12- A inativação da desidrogénase do piruvato pela cínase da desidrogénase do piruvato e a sua
ativação pela fosfátase da desidrogénase do piruvato.
A desidrogénase do piruvato (ver Equação 17) é, de facto, um complexo multienzímico
composto por 5 tipos diferentes de proteínas que se designam por (E1) desidrogénase do piruvato, (E2)
transacetílase do dihidrolipoato, (E3) desidrogénase do dihidrolipoato, (E4) cínase da desidrogénase do
piruvato e (E5) fosfátase da desidrogénase do piruvato. Estas proteínas estão organizadas em agregados
na matriz da mitocôndria.
As proteínas E1, E2 e E3 são coletivamente responsáveis pela catálise expressa pela Equação 17 e
contêm, como grupos prostéticos tiamina-pirofosfato, ácido lipóico e FAD (dinucleotídeo de flavina e
adenina), respetivamente2.
A cínase da desidrogénase do piruvato (E4; ver Equação 18) e a fosfátase da desidrogénase do
piruvato (E5; ver Equação 19) têm papéis reguladores da atividade catalítica do complexo. Quando a
proteína E1 de um dado complexo está na forma desfosforilada este complexo está ativo enquanto a
forma fosforilada é inativa. A cínase da desidrogénase do piruvato catalisa a fosforilação da proteína E1
catalisando a transferência do fosfato γ do ATP (ver Equação 18) e, consequentemente, inativando o
complexo. A fosfátase da desidrogénase do piruvato catalisa a hidrólise de E1 fosforilada (ver Equação
19) e, consequentemente, promove a ativação do complexo.
Equação 18
Equação 19
ATP + desidrogénase do piruvato desfosforilada (ativa) →
ADP + desidrogénase do piruvato fosforilada (inativa)
desidrogénase do piruvato fosforilada (inativa) + H2O →
desidrogénase do piruvato desfosforilada (ativa) + Pi
A atividade da desidrogénase do piruvato depende da proporção de moléculas de enzima que está
na forma desfosforilada, a forma ativa.
13- Em algumas células do organismo a glicólise é, normalmente, predominantemente aeróbica
e noutras predominantemente anaeróbica
Na maioria das células as glicólises aeróbica e anaeróbica coexistem: algumas das moléculas de
ácido pirúvico formadas a partir da glicose são reduzidas a ácido láctico e outras são oxidadas a acetilCoA. A proporção depende da percentagem de moléculas de NADH citoplasmático que é oxidada pelo
piruvato e pelo oxigénio e das atividades relativas das desidrogénases do lactato e do piruvato (ver
Equação 12 e Equação 17).
Contudo, no caso do cérebro e do coração, praticamente todas as moléculas de glicose que entram
para as células sofrem glicólise aeróbica. Nestes órgãos, as moléculas de ácido pirúvico não são reduzidas
a lactato, mas sim oxidadas pela ação da desidrogénase do piruvato (Equação 17) e, concomitantemente,
2
A ação catalítica do complexo desidrogénase do piruvato resulta da ação sequenciada das três proteínas (unidades
enzímicas) que o constituem.
E1 (desidrogénase do piruvato) tem duas atividades que decorrem sequencialmente: descarboxilação do piruvato (ver
reação E1a) seguida de oxidação do grupo hidroxietilo do intermediário formado na reação de descarboxilação, o
hidroxietil-tiamina-pirofosfato (ver reação E1b). O oxidante é o lipoato que é o grupo prostético de E2 (transacetílase do
dihidrolipoato): na oxidação do grupo hidroxietilo (2C) forma-se o grupo acetilo (2C) havendo concomitantemente
redução do lipoato a dihidrolipoato. Simultaneamente com a reação redox ocorre transferência da unidade de dois
carbonos da tiamina-pirofosfato para o dihidrolipoato. Assim, as reações E1a e E1b são catalisadas por E1.
(E1a) piruvato + TPP → hidroxietil-TPP + CO2
(E1b) hidroxietil-TPP + lipoato → acetil-dihidrolipoato + TPP
A ação catalítica de E2 (transacetílase do dihidrolipoato) resume-se à transferência do resíduo acetilo do acetildihidrolipoato para a CoA.
(E2) acetil-dihidrolipoato + CoA → acetil-CoA + dihidrolipoato
A possibilidade E1 poder continuar a oxidar o hidroxietil-TPP (ver E1b) depende da reoxidação do dihidrolipoato a
lipoato e este processo vai ser catalisado por E3 (desidrogénase do dihidrolipoato). Na oxidação do dihidrolipoato, o
agente oxidante direto é o FAD que, depois de reduzido a FADH2, é reoxidado pelo NAD+.
(E3a) dihidrolipoato + FAD → lipoato + FADH2
(E3b) FADH2 + NAD+ → FAD + NADH
Página 5 de 9
Glicólise e desidrogénase do piruvato; Rui Fontes
os NADH formados na oxidação do piruvato e na glicólise são oxidados pelo O2 (Equação 15). Na
realidade, estes órgãos podem captar lactato do sangue e oxidá-lo a piruvato.
No entanto, pelo menos no caso do coração, em situações em que o fluxo sanguíneo está
perturbado (situações de isquemia como o infarto ou angina de peito) o fornecimento de O2 não é
suficiente para oxidar todas as moléculas de NADH formadas. Assim, ocorre aumento da concentração
intracelular de NADH (e diminuição na de NAD+) que faz com que a reação catalisada pela
desidrogénase do lactato (Equação 12) passe a evoluir no sentido da formação de ácido láctico. Ao
contrário do que acontece no coração saudável (que oxida ácido láctico), o coração isquémico produz
ácido láctico: parte da glicólise do coração isquémico é anaeróbica [4] sendo a Equação 14 a que
melhor descreve o processo.
Uma situação semelhante acontece em tumores sólidos mal irrigados.
A glicólise dos eritrócitos é 100% anaeróbica, mas existem outras células no organismo onde a
glicólise é predominantemente anaeróbica: nestas células a maior parte das moléculas de glicose originam
lactato (que sai para o plasma) e não acetil-CoA. É o caso das células da medula dos rins: a medula renal
ao contrário do córtex renal tem uma irrigação sanguínea escassa e grande parte da glicose que é cindida
na medula renal acaba por formar lactato. Outros casos semelhantes são o cristalino e a córnea (tecidos do
olho).
14- A glicólise anaeróbica permite manter a concentração de ATP estacionária nas fibras
musculares aquando do exercício físico anaeróbico
Apesar do seu baixo rendimento “energético” (apenas 2 moles de ATP formadas por mole de
glicose cindida), a glicólise anaeróbica tem um papel essencial na sobrevivência dos indivíduos e, em
última análise, na sobrevivência das espécies. Nas fibras musculares esqueléticas, em particular nas que
são mais pobres em mitocôndrias (fibras musculares brancas), a glicólise pode aumentar explosivamente
de velocidade em situações em que a velocidade de hidrólise do ATP aumenta de forma dramática (ao
subir a uma árvore, por exemplo). Numa situação deste tipo, embora a velocidade de oxidação do ácido
pirúvico pela cadeia respiratória aumente, este aumento de velocidade não é suficiente para acompanhar o
aumento explosivo da velocidade da glicólise. A atividade da desidrogénase do lactato das fibras
musculares permite este aumento explosivo da velocidade da glicólise porque converte o NADH e o ácido
pirúvico em NAD+ e ácido láctico (Equação 12): em situações deste tipo a maior parte do ATP formado
na fibra muscular (essencial para manter a sua síntese à mesma velocidade com que se hidrolisa) resulta
da glicólise anaeróbica [4-5].
De facto, mesmo em repouso, os músculos produzem, normalmente, algum lactato, mas o
exercício físico intensifica marcadamente o processo [5]. Durante o tipo de exercício físico que é costume
designar-se por “anaeróbico” (halterofilismo, corrida de 100 m, chutar com força, etc.) a concentração de
lactato nas fibras musculares pode aumentar cerca de 30 vezes e é comum afirmar-se que é esta
acumulação do ião lactato que provoca a fadiga. Contudo as evidências experimentais demonstram que,
embora a concentração de lactato formado endogenamente esteja diretamente relacionada com o grau de
fadiga, o ião lactato não interfere na atividade contráctil. Como mostra a Equação 14, na glicólise
anaeróbica uma substância aprótida (a glicose) transforma-se numa outra que emite protões (ácido
láctico). O pKa do ácido láctico (≈4) é mais baixo que o pH do citoplasma das fibras musculares (≈7) e,
por isso, a esmagadora maioria das moléculas de ácido láctico dissocia-se formando o ião lactato e
provocando uma descida do pH (de cerca 7 para valores tão baixos quanto 6,5). No esforço muscular
contráctil em regime anaeróbico a descida do pH inibe a atividade muscular contráctil e é um dos
fatores envolvidos no processo de fadiga que impede um sprinter de continuar a correr depois de terminar
a sua prova. Durante os processos isquémicos a descida do pH intracelular que ocorre devido à glicólise
anaeróbica também contribui para a diminuição da capacidade contráctil e este efeito tem uma enorme
importância clínica: no coração isquémico a capacidade de bombeamento está diminuída.
15- Durante o exercício há mobilização de GLUT4 para a membrana sarcoplasmática e
ativação da desidrogénase do piruvato
No músculo esquelético a velocidade de hidrólise do ATP pode sofrer variações muito marcadas:
nas fibras que estão a contrair pode ser 300 vezes superior à do estado de repouso mas, em condições
fisiológicas, a velocidade de síntese sobe de forma paralela.
Isto só é possível porque a velocidade de consumo de nutrientes, incluindo o de glicose na
glicólise (e subsequente oxidação do piruvato pelo O2) também aumenta marcadamente em
situações de esforço. Os mecanismos moleculares que, nos mamíferos, permitem explicar estas subidas
Página 6 de 9
Glicólise e desidrogénase do piruvato; Rui Fontes
de velocidade da glicólise são ainda mal compreendidos e, por isso, são atualmente objeto de intensa
investigação e especulação [5].
(a) Um dos mecanismos que se crê terem importância nesta adaptação do consumo de glicose às
necessidades de ATP já foi referido acima: o esforço muscular contráctil induz o aumento do número de
moléculas de GLUT4 na membrana das fibras musculares e o consequente aumento na velocidade da
entrada de glicose para essas fibras. Esse aumento dá-se de forma muito rápida porque não implica a
síntese de novas moléculas de transportador nos ribossomas, mas sim a fusão de vesículas intracelulares
(que já contêm GLUT4) com a membrana celular. (b)
(b) Um outro mecanismo que poderá ser importante na adaptação do consumo de glicose às
situações de esforço é o efeito do ião Ca2+ nos processos oxidativos. Quando uma fibra muscular é
excitada pelo seu nervo motor aumenta a concentração citoplasmática e mitocondrial de Ca2+: no caso do
citoplasma este aumento pode ser de 100 vezes (de 0,1 µM para 10 µM) [6]. A fosfátase da
desidrogénase do piruvato (ver Equação 19) é o componente do complexo desidrogénase do piruvato
que catalisa a sua desfosforilação e a consequente ativação do processo de conversão do piruvato em
acetil-CoA (ver Equação 17). A fosfátase da desidrogénase do piruvato é ativada pelo Ca2+ e,
consequentemente, quando a concentração de Ca2+ aumenta em resposta ao estímulo nervoso aumenta
também a velocidade de oxidação do piruvato a acetil-CoA.
16- As concentrações citoplasmáticas de AMP e ADP podem subir marcadamente durante o
exercício físico e ativar a glicólise
É comum escrever-se que nas situações fisiológicas em que há aumento do consumo de ATP nas
células a sua concentração desce. De facto é possível observar descida da concentração de ATP em
condições experimentais que não parecem refletir situações fisiológicas [4, 7, 8]. Atualmente, crê-se que
essas descidas são artefactos experimentais ou refletem condições extremas dificilmente observáveis num
ser vivo saudável; no atual estádio do conhecimento pensa-se que, em condições fisiológicas, a
concentração de ATP não sofre qualquer variação apreciável dentro das células [7, 9-12].
No entanto, no caso dos músculos esqueléticos, parece poder ocorrer em situações de esforço,
aumentos na concentração de ADP e AMP [4, 10-11]. Pode parecer estranho que, resultando o ADP da
hidrólise do ATP (ATP + H2O → ADP + Pi) e o AMP da conversão do ADP (catálise pela cínase do
adenilato; ver Equação 20) possam ocorrer aumentos de concentração do ADP e AMP sem que,
praticamente, ocorram variações na concentração de ATP. Em repouso a concentração de ATP pode ser
cerca de 250 vezes superior à de ADP e cerca de 100000 vezes superior à de AMP [9]. Assim, tendo em
conta a reação de hidrólise do ATP, à descida de 0,4% na concentração de ATP corresponde a subida para
o dobro na concentração de ADP. Por sua vez, tendo em conta o equilíbrio químico da reação catalisada
pela cínase de adenilato (ver Equação 20), a uma subida percentual modesta no ADP corresponde uma
variação percentual muitíssimo marcada na de AMP [10].
Equação 20
2 ADP ↔ AMP + ATP
Quer o AMP quer o ADP são potentes ativadores alostéricos de uma das enzimas da glicólise que
se pensa ter maior importância na regulação da velocidade de fluxo nesta via metabólica: a cínase-1 da
frutose 6-fosfato (ver Equação 3). Curiosamente, um dos substratos desta enzima é o ATP mas, em
concentrações fisiológicas, o ATP, para além de se ligar no centro ativo, pode também ligar-se ao centro
alostérico e inibir a atividade da enzima. A ação ativadora do ADP e do AMP resulta da sua capacidade
para competir com o ATP pelo centro alostérico da enzima: quando as concentrações de AMP e/ou ADP
aumentam substituem o ATP no centro alostérico e impedem a sua ação inibidora.
17- A estimulação da captação celular de glicose e da oxidação da glicose no estado pósprandial
Após a ingestão de glicose ou de compostos que, como o amido, possam originar glicose, a
concentração de glicose sobe no plasma sanguíneo. No entanto esta subida é muito menos marcada que a
que seria de esperar quando se compara a pequena quantidade de glicose existente no organismo (cerca de
12 g) com a que é absorvida após uma refeição normal (cerca de 100 g). A moderação na subida da
concentração plasmática da glicose deve-se em grande parte à estimulação da secreção de insulina e ao
seu papel estimulador na captação e na oxidação da glicose pelas células. A insulina é segregada nas
células β dos ilhéus de Langerhans do pâncreas e vai estimular a oxidação da glicose em muitas células
do organismo nomeadamente nas dos músculos, do tecido adiposo, dos rins e do fígado.
Página 7 de 9
Glicólise e desidrogénase do piruvato; Rui Fontes
18- Mecanismos envolvidos na estimulação da captação e da oxidação muscular de glicose no
estado pós-prandial
Os músculos, que podem representar cerca de 40-50% do peso total de um ser humano, têm um
papel muitíssimo importante nos processos de limitação da subida e na subsequente normalização da
glicemia. As ações da insulina nos músculos têm algumas semelhanças com as que forma descritos para o
exercício físico. Da mesma forma que o exercício também a insulina provoca a migração de GLUT4
para a membrana sarcoplasmática e também provoca a ativação da desidrogénase do piruvato.
Parte da glicose que é captada para dentro das fibras musculares após as refeições vai ser
armazenada (como glicogénio) mas, porque a desidrogénase do piruvato também é ativada pela insulina,
a oxidação do piruvato e, em última análise a oxidação da glicose, também são ativadas. A ativação da
desidrogénase do piruvato pela insulina é indireta e ocorre via ativação (não dependente do Ca2+) da
fosfátase da desidrogénase do piruvato (ver Equação 19) e inibição da síntese da cínase da desidrogénase
do piruvato (ver Equação 18). O aumento da atividade da fosfátase da desidrogénase do piruvato e a
diminuição da da cínase têm a mesma consequência: aumento da proporção das moléculas de
desidrogénase de piruvato que ficam na forma ativa [11].
19- Mecanismos envolvidos na estimulação da captação e da oxidação hepática de glicose no
estado pós-prandial
O fígado também participa na moderação da subida da glicemia e na sua normalização
durante e após a absorção intestinal de glicose. No entanto, os mecanismos envolvidos têm algumas
diferenças relativamente aos músculos.
Ao contrário do que acontece nos músculos, a concentração de glicose dentro dos hepatócitos é
semelhante à do sangue: o transportador GLUT2 (o transportador de glicose na membrana dos
hepatócitos) tem grande atividade e permite manter uma situação de “quase equilíbrio” entre as duas faces
da membrana. Assim, quando aumenta a concentração de glicose no sangue também aumenta, de forma
paralela, a concentração de glicose dentro dos hepatócitos. Quando a glicemia é baixa, grande parte da
cínase da glicose (ou hexocínase IV; ver Equação 1) está sequestrada dentro do núcleo ligada a uma outra
proteína que a inibe. A ligação da glicose ao complexo hexocínase IV- proteína inibidora leva à sua
dissociação e à subsequente translocação da hexocínase IV do núcleo para o citoplasma onde inicia a
glicólise [14]. O aumento da atividade da hexocínase IV (ver Equação 1) faz com que a glicose baixe de
concentração dentro dos hepatócitos criando o gradiente que permite a sua captação a partir do sangue.
O aumento da concentração plasmática da insulina (que acompanha a da glicose) também
estimula a glicólise hepática: no fígado, os efeitos ativadores da insulina na velocidade da glicólise
resultam, em parte, da indução da síntese de enzimas chave da glicólise, como a cínase da glicose e a
cínase do piruvato (ver Equação 10).
Uma outra enzima chave da glicólise cuja atividade aumenta no fígado após as refeições é a
cínase-1 da frutose-6-fosfato (ver Equação 3). Neste caso o aumento é também uma consequência da
ação insulínica, mas não se deve a aumento da síntese de moléculas da enzima. A ativação deve-se ao
aumento da concentração intracelular de um composto (frutose-2,6-bisfosfato) que é um ativador
alostérico da cínase-1 da frutose-6-fosfato.
Tal como nos músculos, a insulina também estimula indiretamente (via ações antagónicas na
cínase e na fosfátase) a desidrogénase do piruvato hepática [11].
20- A oxidação de glicose pelo cérebro
Em muitas situações, o órgão que está num dado momento a oxidar mais glicose é o cérebro.
Isto só não acontece no período pós-prandial ou quando se se faz exercício físico; nestes casos os
músculos esqueléticos considerados no seu todo podem ser os maiores consumidores de glicose.
A entrada e a oxidação de glicose no cérebro só são influenciadas pela insulina porque, ao
contrário do que acontece noutros órgãos, estes processos podem ser indiretamente prejudicados. Se um
indivíduo se injetar com uma dose alta de insulina a glicose entra para os músculos e para o tecido
adiposo e a glicemia pode baixar para níveis tão baixos que a velocidade de entrada de glicose para o
cérebro diminua prejudicando a síntese de ATP nos neurónios. Excetuando situações de jejum muito
prolongado (vários dias) o único combustível do cérebro é a glicose sanguínea e se a glicemia baixar
demasiado o indivíduo sofre um desmaio; se a situação se mantiver pode haver lesões no cérebro.
A glicólise que ocorre no cérebro é aeróbica: ou seja, o NADH formado na glicólise cerebral é
oxidado pelo O2 (ver Equação 15) e o piruvato formado não se converte em lactato mas sim, via ação da
Página 8 de 9
Glicólise e desidrogénase do piruvato; Rui Fontes
desidrogénase do piruvato (ver Equação 17), em acetil-CoA. Em condições normais a velocidade de
hidrólise de ATP no cérebro considerado como um todo não sofre variações nem com o estado nutricional
nem com a atividade cerebral (o cérebro hidrolisa tanto ATP a dormir como a resolver problemas
complexos) e o mesmo acontece com a velocidade de oxidação da glicose (cerca de 80 mg/min; 120
g/dia) [12].
1. Russell, R. R., 3rd, Bergeron, R., Shulman, G. I. & Young, L. H. (1999) Translocation of myocardial GLUT-4 and
increased glucose uptake through activation of AMPK by AICAR, Am J Physiol. 277, H643-9.
2. Millman, M. S. & Omachi, A. (1972) The role of oxidized nicotinamide adenine dinucleotide in fluoride inhibition of
active sodium transport in human erythrocytes, J Gen Physiol. 60, 337-50.
3. Halestrap, A. P. & Price, N. T. (1999) The proton-linked monocarboxylate transporter (MCT) family: structure,
function and regulation, Biochem J. 343 Pt 2, 281-99.
4. Stanley, W. C. (2004) Myocardial energy metabolism during ischemia and the mechanisms of metabolic therapies, J
Cardiovasc Pharmacol Ther. 9 Suppl 1, S31-45.
5. Korzeniewski, B. & Liguzinski, P. (2004) Theoretical studies on the regulation of anaerobic glycolysis and its
influence on oxidative phosphorylation in skeletal muscle, Biophys Chem. 110, 147-69.
6. Balaban, R. S. (2002) Cardiac energy metabolism homeostasis: role of cytosolic calcium, J Mol Cell Cardiol. 34,
1259-71.
7. Korzeniewski, B. (2006) Oxygen consumption and metabolite concentrations during transitions between different
work intensities in heart, Am J Physiol Heart Circ Physiol. 291, H1466-74.
8. Ihlemann, J., Ploug, T., Hellsten, Y. & Galbo, H. (2000) Effect of stimulation frequency on contraction-induced
glucose transport in rat skeletal muscle, Am J Physiol Endocrinol Metab. 279, E862-7.
9. Spriet, L. L., Howlett, R. A. & Heigenhauser, G. J. (2000) An enzymatic approach to lactate production in human
skeletal muscle during exercise, Med Sci Sports Exerc. 32, 756-63.
10. Sacktor, B. & Wormser-Shavit, E. (1966) Regulation of metabolism in working muscle in vivo. I. Concentrations of
some glycolytic, tricarboxylic acid cycle, and amino acid intermediates in insect flight muscle during flight, J Biol Chem.
241, 624-31.
11. Strumilo, S. (2005) Short-term regulation of the mammalian pyruvate dehydrogenase complex, Acta Biochim Pol.
52, 759-64.
12. Frayn, K. N. (2012) Regulação Metabólica. Uma perspetiva focada no organismo humano., U.P. Editorial, Porto.
Página 9 de 9