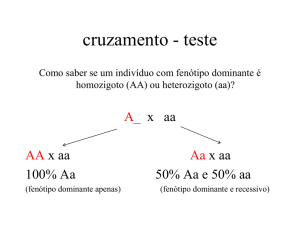
GENÉTICA DE TRANSMISSÃO:
EXTENSÕES DO MENDELISMO
52
EXTENSÕES DO MENDELISMO
INTRODUÇÃO
Os experimentos de Mendel estabeleceram que os genes existem em formas
alternativas (alelos). Na simplificação Mendeliana, para cada característica só
existiam dois alelos: o dominante que contribuía de forma definitiva para produzir
o fenótipo, e o recessivo que só se expressava na ausência do dominante. Hoje
sabemos que cada gene pode apresentar várias formas alélicas na natureza (que
surgem por mutação) e que essas várias formas podem apresentar efeitos
diferentes sobre o fenótipo. Evidentemente cada indivíduo diploide só pode
apresentar dois desses alelos, já que esses alelos se encontram na mesma posição
(locus) nos pares de cromossomos homólogos.
1. ALTERAÇÕES NAS PROPORÇÕES FENOTÍPICAS MENDELIANA
1. 1. DOMINÂNCIA INCOMPLETA
Na dominância completa, o alelo é dominante se tiver o mesmo efeito
fenotípico em dose dupla (AA) ou simples (Aa), e o organismo para essa
característica só apresenta dois fenótipos distintos, o dominante e o recessivo, (
representado pelo genótipo aa). No estudo da característica cor da flor em bocade-leão, Antirrhinum majus, foram observados três fenótipos diferentes: o
vermelho, o branco e o rosa. Ao ser realizado o cruzamento entre plantas de cor
vermelha(R1R1) e branca(R2R2), todos os descendentes F1 nasceram com uma cor
intermediária, rosa(R1R2). Ao cruzar as plantas de flores rosa (F 1) nasceram em F2:
1/4 de plantas de flores vermelha; 2/4 plantas de flores rosa e 1/4 de plantas de
flores brancas, semelhante com a proporção genotípica de 1/4 R 1R1; 2/4 R1R2 e
1/4 R2R2 obtida. A explicação para essa alteração na proporção fenotípica em
relação à proporção obtida por Mendel é que o alelo FV daria como produto gênico
final uma certa quantidade de pigmento, se ele aparece em dose dupla (F VFV =
fenótipo vermelho), ele produzirá duas vezes mais pigmento do que quando ele
aparece em dose simples (FVFB = fenótipo rosa), já o FBFB não produz pigmento,
53
resultando na cor branca. Esse tipo de herança não invalida a 1ª lei de Mendel,
mas apresenta uma proporção fenotípica diferente da obtida por ele em F 2, e como
o fenótipo do heterozigoto é intermediário entre os dos homozigotos, foi
denominado de dominância incompleta. (fig. 2.1).
Figura 2.1: Dominância incompleta mostrada na cor da flor boca-de-leão. Fonte: Klug et
al.; 2010.
54
1.2. CODOMINÂNCIA
Outra exceção ao princípio de dominância completa surge quando um
heterozigoto apresenta característica encontrada em cada um dos homozigotos
associados, produzindo dois produtos gênicos detectáveis; nesse caso, a expressão
conjunta dos dois alelos no heterozigoto é denominada codominância. Um
exemplo desse tipo de herança é a do grupo sanguíneo do sistema MN, descoberto
por Karl Landsteiner e Philip Levin, controlado por alelos presentes no
cromossomo 4, indivíduos homozigotos para o alelo L M, produzem uma molécula
glicoprotéica na superfície dos eritrócitos que é um antígeno natural,
apresentando, dessa forma, o fenótipo grupo sanguíneo M; os que apresentam
somente alelos LN já produzem um outro tipo de glicoproteína na superfície das
hemácias que também funcionam como antígeno natural e o fenótipo é grupo
sanguíneo N; já o heterozigoto, que apresenta tanta o alelo L M, quanto o alelo LN
produzem os dois tipos de glicoproteínas e o fenótipo é grupo sanguíneo MN.
Como é previsto, um cruzamento entre dois genitores heterozigotos MN
pode produzir filhos com os três tipos de fenótipos, M, MN e N, na proporção de
1:2:1, semelhante à proporção genotípica.
Como não existe dominância completa entre os alelos, a notação genética
mais utilizada é a de representar os alelos com a mesma letra maiúscula e
sobrescrito a letra dos alelos alternativos. (fig.2.2).
Figura 2.2: Exemplo de codominância
Tabela representando os genótipos e fenótipos para a característica sistema sanguíneo MN
Genótipos
Fenótipos
LM LM
Grupo sanguíneo M
LM LN
Grupo sanguíneo MN
LNLN
Grupo sanguíneo N
55
1.3. ALELOS MÚLTIPLOS OU POLIALELIA
O conceito mendeliano de que os genes existem em não mais que dois
estados alélicos foi modificado quando se descobriu que a sequência de DNA, que
determina um gene, pode sofrer inúmeras mutações, em pontos diferentes,
originando diversos tipos de alelos. Esses diversos alelos só podem ser
identificados em um estudo genético populacional, já que nos organismos
diploides cada indivíduo só herda 2 alelos, presentes em um mesmo locus nos
cromossomos homólogos. Quando na população existem mais de dois estados
alélicos de um mesmo gene, estamos falando de alelos múltiplos ou polialelia.
Um dos exemplos clássicos de polialelia é a cor da pelagem em coelhos, que
apresenta 4 formas alélicas, cuja notação utilizada é c determina pelagem albina
(todo branco), ch, pelagem himalaia (corpo branco e as extremidades pretas, patas,
focinho e orelhas), cch, pelagem chinchila (pelagem branca com a ponta dos pelos
preta, o que dá uma ideia de conjunto acinzentado) e c+, pelagem selvagem (pelo
colorido por todo o corpo, normalmente castanho). O estudo de diversos
cruzamentos na população de coelhos permitiu determinar a relação de
dominância entre os diversos alelos.
c+ > cch > ch > c, o sinal > indicando
dominância. (fig.2.3)
Figura 2.3: Exemplo de alelos múltiplos em coelhos
Tabela representando os diversos genótipos e fenótipos para a característica cor da
pelagem em coelhos.
Genótipos
Fenótipos
c+c+, c+cch , c+ch, , c+c
Pelagem tipo selvagem
cchcch, cchch, , cchc
Pelagem tipo chinchila
chch, chc
Pelagem tipo himalaia
cc
Pelagem branca ou albina
56
Outro exemplo de alelos múltiplos é o sistema sanguíneo ABO em humanos,
descoberto por Landsteiner, no início da década de 1900, e caracterizado pela
presença de antígenos na superfície dos eritrócitos. Com três alelos alternativos de
um gene, IA, IB e IO, a designação I representa isoaglutinogênio, outro termo para
antígeno, localizados em um locus do cromossomo 9. Mais uma vez lembrando que
apesar de na população encontrarmos três tipos de alelos para a determinação do
sistema sanguíneo ABO, cada indivíduo só é capaz de herdar dois desses alelos, um
que vem no cromossomo 9 de origem paterna e outro que vem no cromossomo 9
de origem materna.
O fenótipo ABO de qualquer indivíduo é averiguado mediante mistura de
uma amostra de sangue com um antissoro que contém anticorpos anti-A ou anti-B.
Se o antígeno estiver presente na superfície dos eritrócitos da pessoa, reagirá com
o anticorpo correspondente e causará agregação, ou aglutinação, dessas células
sanguíneas. Quando o indivíduo é testado desse modo, será revelado um entre
quatro fenótipos, se o indivíduo tiver o antígeno A ele será do grupo sanguíneo A,
caso tenha o antígeno B, ele será do grupo sanguíneo B, se tiver ambos os
antígenos, A e B, ele será do grupo sanguíneo AB, e caso não seja detectado
nenhum dos dois antígenos, ele será do grupo sanguíneo O.
Com relação aos genótipos, após estudos em muitas famílias diferentes,
chegou-se à conclusão de que, entre os três alelos encontrados na população, os
alelos IA e IB apresentam uma relação de codominância, e os alelos IA e IB com o
alelo IO uma relação de dominância, podendo ser encontrados os seguintes
genótipos e fenótipos na população, ver tabela abaixo na figura 2.4.
O conhecimento sobre os grupos sanguíneos humanos tem várias
aplicações. Uma das mais importantes é testar a compatibilidade das transfusões
de sangue. Outra aplicação envolve os casos de investigação de paternidade, em
que os recém-nascidos são inadvertidamente trocados no hospital, ou quando é
incerto se um homem específico é o pai de uma criança. Um exame dos grupos
sanguíneos ABO, assim como de outros antígenos hereditários, dos genitores e da
criança, pode ajudar a excluir a paternidade ou a maternidade, mas jamais prova a
paternidade ou maternidade.
57
Figura 2.4: Exemplo de alelos múltiplos em humanos
Tabela representando os genótipos e fenótipos para a característica sistema sanguíneo ABO
Genótipos
Fenótipos
IAIA, IAIO
Grupo sanguíneo A
IBIB, IBIO
Grupo sanguíneo B
IAIB
Grupo sanguíneo AB
IOIO
Grupo sanguíneo O
1.3.1. MECANISMOS BIOQUÍMICOS PARA FORMAÇÃO DOS ANTÍGENOS A E B
Os antígenos A e B são carboidratos que se ligam a moléculas de
lipídeos(ácidos graxos) na superfície externa da membrana celular dos eritrócitos.
Tanto o antígeno A como o antígeno B têm como substância precursora uma
substância denominada substância H ou antígeno H, constituído por três
moléculas de carboidrato; galactose(Gal), N-acetilglicosamina (AcGluNH) e
fucose ligadas quimicamente. A especificidade dos antígenos A e B é dada pela
ligação química na porção terminal da substância H de mais um grupamento
carboidrato.
O produto gênico do alelo IA é uma enzima que adiciona à substância H o
carboidrato N-acetilglicosamina (AcGluNH). O alelo IB tem como produto uma
enzima modificada que só consegue adicionar a porção terminal da substância H
uma galactose(Gal). Indivíduos IAIB adicionam ou um ou outro na porção
terminal, e podemos encontrar, nesse caso, tanto substância H acrescida de
acetilglicosamina (AcGluNH), formando o antígeno A, ou acrescida de
galactose(Gal), formando o antígeno B nas superfícies dos eritrócitos. O alelo IO
apresenta uma mutação que não permite que seu produto gênico acrescente
58
nenhum carboidrato na porção terminal da substância H, sendo encontrada, em
indivíduos de fenótipo O, somente a substância H.(fig. 2.5)
Figura
2.5:
Mecanismo
bioquímico para formação dos
antígenos A e B, a partir da
substância
H,
com
a
participação dos genes IA e IB e
FUT1 na formação das enzimas
envolvidas. Fonte: Klug et al.;
2010.
1.3.2. O FENÓTIPO BOMBAIM
Em 1952, uma situação muito rara propiciou informações sobre a base
genética da substância H. Uma mulher, em Bombaim, Índia, ao necessitar de uma
transfusão, fez uma tipagem sanguínea e diagnosticou-se que ela não possuía
nenhum dos antígenos, A ou B sendo, portanto, do grupo sanguíneo O. Porém ao se
fazer a árvore genealógica dela, observou-se que um dos pais era do grupo AB e ela
59
tinha doado a dois filhos o alelo IB, o que é era inconsistente com a tipagem
sanguínea.
Posteriormente, demonstrou-se que a mulher era homozigota para uma
mutação recessiva rara em um gene denominado FUT1(codificador da enzima
fucosil-transferase), responsável pela ligação química na porção terminal da
substância H, da fucose. A substância H incompleta (sem fucose) não é reconhecida
pelas enzimas produzidas pelos genes IA e IB , não podendo formar os antígenos A
ou B e apresentando-se funcionalmente como do grupo O. Os filhos que herdaram
o alelo IB são heterozigotos para o gene FUT1, logo formam a substância H e
consequentemente os antígenos B.
1.4. GENES LETAIS
Um alelo que é capaz de causar a morte de um organismo é chamado de
alelo letal. Muitos produtos gênicos são essenciais ao desenvolvimento normal e à
sobrevivência de um organismo. Quando os genes que os produzem mutam, pode
resultar na morte prematura do organismo, dependendo da fase do
desenvolvimento (embrionário, primeira infância ou adulto) em que seu produto
gênico vai ser necessário. Quando, para ocorrer a morte do indivíduo, são
necessários dois alelos mutantes, chamamos a letalidade de recessiva, mas se um
único alelo mutante já determinar a morte do indivíduo, chamamos a letalidade de
dominante.
Existem alelos que podem determinar mais de uma característica, já que
seus produtos podem interferir em mais de uma via metabólica, quando isso
ocorre, chamamos o processo de pleiotropia
Um exemplo de genes letais é um gene pleiotrópico que participa da
determinação da cor da pelagem em camundongo e da sobrevida. O alelo AY
determina pelagem amarela, quando em
heterozigose, enquanto o alelo A
determina pelagem aguti(cinzenta), quando em homozigose, porém o genótipo
AYAY mata ainda no período embrionário, não sendo encontrados camundongos
amarelos homozigotos. Logo o comportamento do alelo AY em relação à
sobrevivência é de letal recessivo, já que são necessários 2 alelos iguais para
60
causar a morte do camundongo, enquanto que, em relação à cor da pelagem, ele
comporta-se como um alelo dominante.
A letalidade também altera as proporções fenotípicas e genotípicas
mendelianas, já que alguns embriões morrem antes do nascimento, mudando
assim a proporção de nascidos vivos. Na figura 2.6, estão representados alguns
cruzamentos e as proporções fenotípicas e genotípicas resultantes desses
cruzamentos.
Figura 2.6: Exemplos de cruzamentos com alelo letal onde se percebe
alterações nas proporções fenotípicas e genotípicas. Fonte: Klug et al.;
2010.
61
O fenótipo sem cauda Manx em gatos também é produzido por um alelo que
é letal no estado homozigoto. Uma única dose do alelo Manx, ML, interfere
gravemente no desenvolvimento da coluna dorsal, resultando na falta de cauda no
heterozigoto MLM. Mas, no homozigoto MLML, a dupla dose do gene produz uma
anomalia tão extrema no desenvolvimento da coluna, que o embrião não
sobrevive.
Os alelos para cor da pelagem em camundongos e para o fenótipo sem
cauda Manx sendo genes pleiotrópicos apresentam fenótipos visíveis em
heterozigose, mas a maioria dos letais recessivos são silenciosos no heterozigoto.
Em tal situação, a letalidade recessiva é diagnosticada observando a morte de 25%
da prole em algum estágio do desenvolvimento.
Um exemplo de gene letal dominante é o da doença de Huntington, que se
caracteriza pela degeneração motora e nervosa, em humanos. Causada pelo alelo
autossômico dominante H, essa doença só se manifesta nos heterozigotos (Hh) na
idade adulta, permitindo assim que esses indivíduos cheguem à idade reprodutiva
e transmitam seus genes para os descendentes.
Genes letais dominantes são raros na população, quando causam morte
antes da idade reprodutiva, pois não permitem a perpetuação do alelo.
1.5. PENETRÂNCIA E EXPRESSIVIDADE
A herança monogênica estudada até agora produz mutantes e selvagens que
produzem claras proporções mendelianas. Em tais casos, podemos usar o fenótipo
para distinguir os genótipos mutantes e selvagens com quase 100% de certeza.
Mas existem muitos casos em que, mesmo o genótipo estando presente, o fenótipo
não é expresso. Sabemos, hoje, que a expressão do fenótipo não só depende do
genótipo e sim da interação desse genótipo com o meio interno celular, inclusive
com a possibilidade de interação com outros genes não caracterizados, com efeitos
epistáticos ou supressores, como também com o meio externo.
62
Definimos Penetrância como a porcentagem de indivíduos com um
determinado alelo que exibem o fenótipo associado a esse alelo. Nos casos em que
um determinado alelo está presente e expressa com 100% de certeza o fenótipo,
dizemos que a penetrância é completa. Mas quando um determinado alelo, como o
da polidactilia postaxial (herança autossômica dominante caracterizada por um
dedo extranumerário próximo ao quinto dedo da mão ou do pé) está presente, mas
o fenótipo só é expresso em 64,9% dos indivíduos, segundo estudo populacional na
África, a penetrância é dita incompleta, e a penetrância desse alelo é de 64,9%, ou
seja, 35,1% apesar de terem o genótipo para polidactilia, não apresentam o
fenótipo.
Outra medida para descrever a gama de expressão fenotípica é a chamada
de expressividade. A expressividade mede o grau em que determinado alelo é
expresso em nível fenotípico; isto é, a expressividade mede a intensidade do
fenótipo. Por exemplo, em cães da raça beagles, o alelo dominante, S, determina
pelagem homogênea, sem manchas, decorrente da distribuição homogênea dos
melanócitos. O alelo recessivo, s, determina uma distribuição heterogênea dos
melanócitos durante o desenvolvimento embrionário. Animais com genótipo SS ou
Ss, apresentam pelagem sem manchas, entretanto os com genótipos ss,
apresentam pelo menos 10 tipos diferentes de padrão de manchas( fenótipo
variegado), indo desde quase sem manchas até uniformemente pigmentados, como
o do genótipo dominante. Alelos que produzem fenótipos tão variados em seus
portadores, fala-se em expressividade gênica variável. (fig. 2.7)
63
Figura
2.7:
Representação
esquemática. Em cães da raça beagle
podem-se distinguir 10 padrões de
pelagem (ver no esquema, 1 a 10)
devidos à expressividade variável do
alelo que condiciona a variegação da
pelagem (Baseado em Griffiths, A. J.
F e cols., 1998). Fonte: Amabis, José
M.,Martho, Gilberto R., 2006.
1.6. INTERAÇÕES GÊNICAS NÃO ALÉLICAS
Caracteriza-se pela interação entre dois ou mais alelos, presentes no mesmo
ou em cromossomos homólogos diferentes, determinando uma mesma
característica.
A análise da proporção fenotípica entre os descendentes de um
cruzamento, além de informar quantos genes estão envolvidos na formação da
característica pode também revelar o tipo de interação entre eles.
1.6.1. INTERAÇÃO GÊNICA SIMPLES
Algumas das primeiras evidências de que uma característica pode ser
influenciada por mais de um gene foram obtidas por Bateson e Punnett, em 1905,
de experimentos de cruzamentos em galinhas. Tipos diferentes de galinhas
domésticas têm formas de cristas diferentes. As da raça Wyandottes têm cristas
rosa, as Brahmas têm cristas ervilhas, do cruzamento de Wyandottes e Brahmas
(cristas rosa e ervilha); apareceu outro tipo de crista denominada noz, e do
cruzamento de duas aves noz, obteveram-se quatro tipos de fenótipos: os três já
64
conhecidos, crista rosa, ervilha, noz e outro fenótipo chamado de crista simples em
galinhas da raça leghorns, pela proporção fenotípica da descendência, crista
simples é representado pelo genótipo duplo-recessivo.( fig. 2.8) Bateson e Punnett
descobriram que o tipo de crista é determinado pela interação de dois pares de
alelos que se segregam independentemente. Usando a notação E e e para
representar os alelos do par que determina a forma crista ervilha; a notação R e r
a forma da crista rosa; e E_ expressando que o genótipo pode ser EE ou Ee o
mesmo aplicável para o R_, representamos os genótipos da seguinte forma: O
genótipo E_ rr, determinaria o fenótipo crista ervilha;
o genótipo
ee R_ o fenótipo crista rosa; E_ R_, resultante do cruzamento de aves de crista
ervilha com crista rosa, o fenótipo crista noz; e o genótipo ee rr o fenótipo crista
simples.(fig.2.8)
Figura 2.8: Formas das cristas de galinha de
raças diferentes. (a) Rosa, Wyandottes; (b)
Ervilha, Brahmas; (c) noz, híbrida do
cruzamento entre galinhas com cristas rosa e
ervilha; (d) simples, Leghorns.Fonte: Snustad,
D.Peter; Simmons, Michael J.,2008.
65
P
Gametas
F1
Wyandotte
(rosa)
ee RR
eR
X
Brahma
(ervilha)
EE rr
Er
Híbrido
X
Híbrido
Ee Rr
Ee Rr
Gametas masculinos
ER
Er
eR
er
F2
ER
EE RR
EE Rr
Ee RR
Ee Rr
Gametas
femininos
Er
EE Rr
EE rr
Ee Rr
Ee rr
eR
Ee RR
Ee Rr
ee RR
ee Rr
er
Ee Rr
Ee rr
ee Rr
ee rr
Figura 2.9: O experimeto de Bateson e Punnett sobre a forma das cristas em galinhas. O
entrecruzamento na F1 produz quatro tipos de fenótipos, cada um destacado por uma cor
diferente no quadrado de Punnett, em uma proporção 9:3:3:1
Outro exemplo de interação gênica simples é o que ocorre com a cor da
plumagem em periquitos australianos, esses periquitos apresentam um grande
gama de cores, determinadas por dezenas de genes. No entanto, na determinação
das cores básicas da plumagem dessas aves – verde, azul, amarela e branca - estão
envolvidos somente dois pares de alelos, o par A e a e o par B e b, que se segregam
independentemente. Periquitos homozigóticos recessivos apresentam genótipo aa
bb e um fenótipo branco para a plumagem; Periquitos aa B_ são amarelos; já os A_
bb são azuis e os A_ B_ são verdes. O cruzamento de periquitos verdes
heterozigotos produz os 4 tipos de fenótipos na proporção de 9/16 verde: 3/16
amarelo: 3/16 azul e 1/16 branco.
66
Tanto o alelo A como o alelo B produzem pigmentos. O alelo A produz o
pigmento melanina (um pigmento escuro) que, devido à dispersão da luz na
superfície da pena, contra o fundo escuro de melanina no centro da pena, aparece
como azul. O alelo B produz um pigmento amarelo chamado psitacina, que se
deposita na pena. Os alelos a e b são formas alteradas e não produzem,
respectivamente, melanina e psitacina, dando um fenótipo branco. Quando os
alelos A e B estão constituindo um mesmo genótipo, a cor da plumagem é verde,
essa cor resulta da mistura do efeito visual azul, causado pela presença de
melanina, e do amarelo, causado pela presença do pigmento psitacina.(fig 2.10)
Á Figura 2.10: Esquemas de cortes
transversais das penas de periquitos para
mostrar como a presença e a distribuição dos
pigmentos melanina e psitacina determinam a
cor da plumagem. (Baseado em Campbell, N. A.
e cols., 1994).Fonte: Amabis, José M.,Martho,
Gilberto R., 2006.
1.6.2. EPISTASIA
É um exemplo de interação onde o efeito de um gene ou de um par de genes
dissimula ou modifica o efeito de outro gene ou de outro par gênico. Às vezes, os
genes envolvidos influem na mesma característica fenotípica de modo antagonista,
o que leva à dissimulação. Em outros casos, entretanto, os genes envolvidos
exercem sua influência reciprocamente, de maneira complementar ou cooperativa.
67
A epistasia pode ser recessiva, quando o par de alelos localizados em um
locus impede ou suprime a expressão, do par de alelos, em outro locus. O par que
tem o efeito supressor é dito epistático e o par suprimido é dito hipostático.
Um exemplo de epistasia recessiva, a que já nos referimos, é a do
fenótipo Bombaim, o gene H quando em homozigose recessiva suprime a
expressão dos genes IA ou IB, apresentando o portador desse genótipo, fenótipo do
grupo sanguíneo O.(fig.2.5 e 2.11)
F1
X
G. sanguíneo AB
Hh IAIB
G. sanguíneo AB
Hh IAIB
Gametas masculinos
A
HI
HI
h IA
h IB
H IA
HH IAIA
HH IAIB
HH IAIA
Hh IAIB
Gametas
H IB
HH IAIB
HHIBIB
Hh IAIB
HhIBIB
femininos
h IA
Hh IAIA
Hh IAIB
hh IAIA
hh IAIB
h IB
Hh IAIB
Hh IBIB
hhIAIB
hhIBIB
F2
B
Proporção Fenotípica: 6/16 G.S.AB; 3/16 G.S. B;
3/16 G.S.A; 4/16 G.S. O
Figura 2. 11: Representação do cruzamento de indivíduos do grupo sanguíneo AB, heterozigotos para
os genes H, com conseqüente surgimento do fenótipo Grupo sanguíneo O (G.S. O).
Outro exemplo de epistasia recessiva é o da cor da pelagem em
camundongos. O gene A produz um pigmento que funciona como precursor dos
alelos P que determina cor aguti (base do pelo preto com ponta amarela), e do
alelo p que determina cor preta, o gene a é alterado e não origina esse precursor.
68
Quando o genótipo do camundongo é A_P_ ou A_ pp os camundongos são aguti ou
pretos, respectivamente. Mas se o gene for aaP_ ou aapp o fenótipo é
albino.(fig.2.12a e b)
Figura 2.12a: Representação esquemática da seqüência de reações
bioquímicas que levam à síntese do pigmento melanina no pêlo de
camundongos aguti, preto e albino. Cada transformação química é controlada
por uma enzima, fabricada por um gene específico. Fonte: Amabis, José
M.,Martho, Gilberto R., 2006.
69
Figura 2.12b: Representação esquemática do cruzamento de camundongos
em que a cor da pelagem resulta da epistasia recessiva. No cruzamento entre
animais duplo-heterozigóticos surge a proporção 9: 3 : 4, característica
desse tipo de epistasia. Fonte: Amabis, José M.,Martho, Gilberto R., 2006.
Quando um único alelo de um par já impede ou suprime o par de alelos de
outro locus, falamos em epistasia dominante.
O exemplo de epistasia dominante é o da cor do fruto em abobrinhas, o alelo
A impede ou suprime a expressão, enquanto o alelo a permite a expressão dos
alelos B e b, que se encontram em outro locus gênico, e determinam a cor amarela
70
e cor verde respectivamente. Como esses alelos segregam independentemente a
proporção em F2 do cruzamento de duas plantas de abobrinhas brancas
heterozigotas é de 12 brancas para 3 amarelas para uma verde.(fig. 2.13)
P
X
Gametas
Abobrinha branca
AA BB
AB
Abobrinha verde
aa bb
ab
F1
Aa Bb
X
Aa Bb
Gametas
F2
Gametas
AB
Ab
aB
ab
AB
AA BB
AA Bb
Aa BB
Aa Bb
Ab
AA Bb
AA bb
Aa Bb
Aa bb
aB
Aa BB
Aa Bb
aa BB
aa Bb
ab
Aa Bb
Aa bb
aa Bb
aa bb
Proporção fenotípica: 12/16 abobrinha branca; 3/16 abobrinha amarela;
1/16 abobrinha verde
Figura 2.13: Quadrado de Punnett representativo de um cruzamento de plantas de abobrinhas em
que a determinação da cor da abobrinha é resultante de epistasia dominante, originando uma
proporção fenotípica modificada de 12: 3: 1.
A cor da plumagem em galinhas também exemplifica a interação epistática
dominante, os pares de alelos que participam são denominados I e i, e o outro par
C e c. A presença de I já suprime ou impede a expressão do C ou c. (fig 2.14)
71
Figura 2.14: Representação
esquemática do cruzamento de
galináceos para coloração das penas,
resultante de epistasia dominante,
originando uma proporção fenotípica
modificada de 13: 3, Essa proporção
difere do esperado para a epistasia
dominante pois o genótipo ii cc também
é branco pois os alelos hipostáticos cc
não produzem pigmentos. Fonte:
Amabis, José M.,Martho, Gilberto R.,
2006.
1.6.3. INTERAÇÃO GÊNICA COMPLEMENTAR (GENES DUPLOS RECESSIVOS)
Bateson e Punnett, descobriram em ervilha-doce (Lathyrus odoratus) ao
cruzar duas plantas de flores brancas homozigotas que a F 1 obtida eram todas de
flores púrpuras, e o resultado da F 2 foi de 9 púrpuras para sete brancas, indicando
que ocorre segregação independente de dois pares de alelos, assim
denominados: em um locus B e b e no outro locus A e a. A explicação para esse
resultado é que a cor da flor da ervilha é dada pela interação complementar de dois
alelos, se o os dois alelos A e B estiverem presentes o pigmento será produzido e a
flor será púrpura, caso falte um dos dois – aa B_, A_ bb ou aa bb – a planta
72
apresentará flor branca, os alelos recessivos aa ou bb dissimulam o expressão do
alelo dominante do outro locus . (fig. 2.15 e 2.16)
Gene A
Gene B
MOLÉCULA
PRECURSORA
(INCOLOR)
PRODUTO
INTERMEDIÁRIO
(INCOLOR)
PRODUTO
FINAL
(PÚRPURA)
A_
B_
Figura 2.15: Representação esquemática da seqüência de reações bioquímicas que levam à síntese do
pigmento púrpura. Cada transformação química é controlada por uma enzima, fabricada por um gene
específico. São necessários ao dois alelos A e B para produção do pigmento. Interação gênica
complementar( genes duplos recessivos)
1.6.4. GENES DUPLOS COM EFEITO CUMULATIVO
Em abobrinha (Cucurbita pepo) a forma do fruto também é um exemplo de
interação
determinada
por
dois
pares
de
alelos
que
se
segregam
independentemente. O fruto apresenta os fenótipos discoide, alongado e esférico. E
representaremos os alelos de um locus com as letras A e a e o outro par de alelos
com as letras B e b. Quando cruzamos uma planta de fruto discóide com uma de
fruto alongado, todos os descendentes em F 1 apresentam fruto discoide, mas na
geração F2, resultantes do cruzamento de F1, aparecem plantas com fenótipo do
tipo esférico, diferente dos outros dois já apresentados. A explicação para esse
resultado é que a presença dos dois alelos A e B determinam o fenótipo discoide,
enquanto a ausência dos dois determina o fenótipo alongado, porém se só um dos
alelos A ou B estão presentes o fenótipo é esférico. Os genes A e B influenciam
igualmente na determinação dos fenótipos. (fig.2.17)
73
Figura 2.16: Representação esquemática de cruzamento entre duas linhagens de ervilhadoce. Nessas linhagens, a coloração das flores depende da interação de dois pares de
alelos que se segregam independentemente. (Interação genes duplos recessivos ou
interação gênica complementar). Fonte: Amabis, José M.,Martho, Gilberto R., 2006.
74
P
Gametas
Abobrinha discoide
AA BB
AB
X
Abobrinha alongados
aa bb
ab
Discoide
Aa Bb
X
Discoide
Aa Bb
F1
Gametas
F2
Gametas
AB
Ab
aB
ab
AB
AA BB
AA Bb
Aa BB
Aa Bb
Ab
AA Bb
AA bb
Aa Bb
Aa bb
aB
Aa BB
Aa Bb
aa BB
aa Bb
ab
Aa Bb
Aa bb
aa Bb
aa bb
Fonte:Klug et. al.,2010
Proporção fenotípica: 9/16 abobrinha fruto discoide; 6/16 abobrinha
fruto esférico; 1/16 abobrinha fruto alongados
Figura 2.17: Quadrado de Punnett representativo de um cruzamento de plantas de abobrinhas em que a
determinação da forma do fruto é resultante de genes duplos de efeitos cumulativos, originando uma
proporção fenotípica modificada de 9:6:1.
Além dessas interações gênicas já mencionadas existem outras que
modificam as proporções fenotípicas de um cruzamento di-híbrido, representadas
na tabela abaixo.(fig.2.18)
75
Tipos de Interação
Interação gênica simples
Genótipos
A_ B_
A_ bb
aa B_
aabb
9
3
3
1
3
1
Epistasia dominante
12
Epistasia recessiva
9
Genes duplos com efeito
9
3
4
6
1
cumulativo
Genes duplos dominantes
Genes duplos recessivos
15
9
Interação dominante e recessiva
1
7
13
3
Figura 2.18: Tabela representativa dos principais tipos de interações gênicas, onde se encontra
relacionado os possíveis genótipos com as proporções fenotípicas do cruzamento de dois duploheterozigotos
1.7. HERANÇA QUANTITATIVA OU POLIGÊNICA
Até agora a maior parte dos nossos exemplos sobre variações fenotípicas
eram tipos que podiam ser classificados em categorias diversas e separadas: as
características das ervilhas de Mendel eram bem contrastantes, cor do cotilédone
verde ou amarelo, textura da semente da ervilha, lisa ou rugosa, mesmo nos casos
de dominância incompleta, os fenótipos eram bem pontuais, no exemplo da flor
boca-de-leão, ela apresenta fenótipos, branco, rosa e vermelho; o grupo sanguíneo
ABO, pode ser A, B, AB, O. Cada uma das características citadas apresenta
genótipos distintos que determinam fenótipos distintos. Quando isso ocorre,
falamos que essas características apresentam variação descontínua. Embora
76
fenômenos como penetrância, expressividade variável, pleiotropia e epistasia
possam confundir um pouco a relação genótipo-fenótipo.
Na herança quantitativa, nós vamos estudar características que apresentam
uma variedade contínua de fenótipos, essa variedade contínua pode ser medida e
descrita em termos quantitativos, onde cada genótipo contribui com uma pequena
parcela para determinar o fenótipo, logo esses fenótipos são resultantes da
contribuição aditiva de dois, três ou um grande número de genes, por isso o nome
dado a esse estudo é herança quantitativa ou poligênica.
São exemplos de herança poligênica ou quantitativa a cor da pele, a altura,
ou o peso na espécie humana, a produção de leite ou de carne no gado, a
produtividade das colheitas e o conteúdo proteico das sementes.
Na herança quantitativa, o genótipo estabelece os limites quantitativos na
fertilização, mas os fenótipos sofrem grande influência do ambiente, por exemplo,
a altura humana é parte geneticamente determinada, mas se a pessoa tem uma boa
alimentação, pratica esportes etc. pode atingir, dentro de uma determinada faixa
de variação da altura genética, a maior altura. Os fenótipos que resultam da ação
gênica e de influências ambientais, às vezes, são denominados características
complexas ou multifatoriais.
Além das características quantitativas contínuas, em que a variação
fenotípica pode situar-se em qualquer ponto de uma variedade de medidas, há
duas outras classes de características poligênicas:
Características merísticas são aquelas em que os fenótipos são descritos
por números inteiros. Exemplo: o número de sementes por vagem, ou o número de
ovos postos por uma galinha, em um ano. São características quantitativas, mas
não têm uma infinidade de fenótipos: Por exemplo, a vagem pode conter 2 ou 4 ou
6 sementes, mas não 5,75.
Características com limiar são características poligênicas, mas que
apresentam só poucos tipos de fenótipos distintos na população. Sofrem grande
influência dos fatores ambientais, sendo, portanto, características multifatoriais.
São de grande interesse dos geneticistas humanos, pois um grande número de
doenças apresenta esse padrão de herança. Exemplo: a diabetes tipo II, a
esquizofrenia, o transtorno afetivo bipolar etc. Em humanos, a evidência de que
77
tais características são influenciadas por fatores genéticos vem de comparações
entre parentes, especialmente gêmeos.
1.7.1. AS CARACTERÍSTICAS QUANTITATIVAS PODEM SER EXPLICADAS EM
TERMOS MENDELIANOS
No início da década de 1900, a explicação da variação fenotípica contínua,
em termos mendelianos, causou muita controvérsia, mas Bateson e Gudny Yule
propuseram a hipótese dos fatores múltiplos ou genes múltiplos, na qual
muitos genes, cada um comportando-se mendelianamente, contribuíam para o
fenótipo de forma cumulativa ou quantitativa. Essa hipótese foi sustentada pelos
resultados experimentais, publicados pelo trabalho com a característica cor do
grão de trigo desenvolvido por Hermann Nilsson-Ehle.
Nilsson-Ehle iniciou seu trabalho cruzando plantas de trigo de grão
vermelho escuro com plantas de trigo de grãos branco, obtendo em F 1 todas as
plantas com uma cor intermediária (vermelha), o que inicialmente o fez suspeitar
de dominância incompleta entre dois alelos de um mesmo locus, mas ao cruzar as
plantas F1, obteve em F2 15/16 plantas com grão que variavam em tons de
vermelho, podendo ser distinguidos até 4 tons de vermelho, e o 1/16 branco,
sugerindo que era uma herança com dois pares de alelos que se segregavam
independentemente.
Cada par tinha um alelo que contribuía de forma aditiva para compor o
fenótipo grão vermelho, e outro que não adicionava nada na formação do fenótipo.
Representando os alelos como A e a e B e b, os que apresentavam fenótipo grão
vermelho escuro possuíam um genótipo com o máximo de genes aditivos AA BB, já
os que eram brancos, apresentavam seu genótipo sem nenhum gene aditivo, aa bb.
F2 então ficou com cinco classes fenotípicas, a primeira representada por 4 alelos
aditivo, AA BB, e fenótipo vermelho escuro; a segunda por 3 alelos aditivos, Aa BB
ou AA Bb e fenótipo vermelho médio; a terceira com 2 alelos aditivos, AA bb, aa BB,
Aa Bb e fenótipo vermelho; a quarta com 1 alelo aditivo, Aa bb ou aa Bb e fenótipo
vermelho claro e a quinta e última classe com 0 alelos aditivo e fenótipo grão
branco. (fig.2.19)
78
Figura 2.19: Representação esquemática
do cruzamento entre plantas de trigo
produtoras de grão vermelhos-escuro e
brancos. A proporção obtida na geração F2
mostra tratar-se de um caso de herança
quantitativa ou poligênica. Fonte: Amabis,
José M.,Martho, Gilberto R., 2006.
Se fizermos um gráfico da distribuição da cor do grão em trigo, ou da
estatura, ou da cor da pele em humanos etc. observaremos que todas essas
características quantitativas apresentam uma mesma curva de distribuição, que
chamamos de curva de distribuição normal, ou curva em forma de sino. (fig. 2.20)
79
6/16
4/16
1/16
Vermelho Vermelho Vermelho Vermelho
escuro
médio
claro
Branco
Figura 2.20: Representação esquemática de um gráfico de distribuição normal da
herança poligênica da cor do grão de trigo , determinada por 2 pares de alelos que se
segregam independentemente , com efeito aditivo
1.7.2. CALCULANDO O NÚMERO DE POLIGENES
Para estimar o número de genes (quantos pares de alelos) envolvidos na
determinação de uma característica quantitativa, pode-se usar a fórmula 1/4n =
relação entre os indivíduos F2 que expressam um dos dois fenótipos extremos. Ou
1/2n relação entre os indivíduos F2 que expressam um dos dois fenótipos
extremos, nesse caso obteremos o número de alelos envolvidos na herança
quantitativa (fig. 2.21).
Quando o número de poligenes é pequeno, às vezes, é mais fácil usar a
equação (2n+1) = número observado de categorias fenotípicas distintas.
Do cruzamento onde os dois genitores são heterozigotos para todos os
genes, pode-se determinar a proporção fenotípica de cada uma das classes
fenotípicas formadas, usando-se o triângulo de Pascal. Vamos supor que desejamos
saber a proporção fenotípica obtida no cruzamento entre dois híbridos para três
pares de genes de efeito cumulativo. Sabemos, pela fórmula, que o número de
fenótipos será sete. Construímos um triângulo com sete linhas. Na primeira,
colocamos o número 1. Os números das linhas começam sempre por 1, e os
números seguintes são obtidos somando o número imediatamente acima com o
80
que está à esquerda dele (quando não houver número acima ou à esquerda,
considera-se zero). Todas as linhas terminam novamente com o número 1:
1
1
1
1
2
1
1
3
3
1
1
4
6
4
1
1
5
10
10
5
1
1
6
15
20
15
6
1
Na sétima linha podemos ver que a proporção fenotípica para três pares de
genes na herança quantitativa (no cruzamento de dois indivíduos heterozigotos) é
1 : 6: 15 : 20 : 15 : 6 : 1.
Determinação do número de poligenes(n) envolvidos em
uma característica quantitativa
n
Indivíduos que expressam um dos
fenótipos extremos
Classes fenotípicas
distintas
1
1/41 = 1/4
3
2
1/42 = 1/16
5
3
1/43 = 1/64
7
4
1/44 =1/256
9
5
1/45 = 1/1024
11
Figura 2.21: Representação na tabela do número de classes fenotípicas , e do
número de genes envolvidos em uma característica quantitativa.Fonte: Klug
et. al., 2010
81