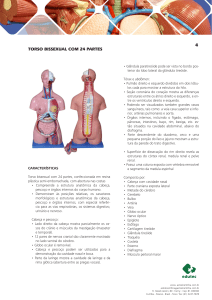
PAPEL DOS HORMÔNIOS NA LACTAÇÃO*
Introdução.
A glândula mamária é um tecido especializado, anatomo-morfologicamente adaptado
a processos filtrativos e bioquimicamente complexo. Seu desenvolvimento caracteriza
uma vitória ontogênica dos mamíferos mediante a qual muitos processos reprodutivos
puderam ser mais eficientes (redução do tempo de gestação nos marsupiais; nascimento
relativamente imaturo -desenvolvimento mandibular sem dentes que favorece a
expulsão da cabeça do feto no momento do parto- e manutenção de crias em fases
posteriores em toda a classe mammalia). Por isto, alguns consideram que a glândula
mamária é um órgão reprodutivo anexo, dado que através de sua secreção a
descendência dos mamíferos, que nasce em estado de total dependência nutricional
obtém o alimento básico para continuar com seu desenvolvimento extra-uterino.
Todos os mamíferos são absolutamente dependentes da mãe, quem mediante a
secreção conhecida como leite entrega à cria um produto nutricionalmente balanceado
que garante a vida do neonato. Na ausência desta, o recém-nascido não sobreviveria
depois do parto.
Reprodução e lactação são partes do mesmo processo. A fisiologia da lactação está
relacionada com a fisiologia dos processos reprodutivos. A maior parte do
desenvolvimento estrutural da glândula mamária ocorre durante a gestação. Neste
período, a fisiologia da gestação desenvolve duas tarefas independentes mas sinérgicas
que garantem a sobrevivência da espécie: de uma parte, o sistema materno fornece
ambiente estéril, proteção, umidade, calor, nutrientes, intercâmbio gasoso, realiza
funções metabólico-sensoriais; de outro lado, provê, mediante o desenvolvimento da
glândula, a possibilidade de fornecer alimento (água, minerais, vitaminas, proteína e
energia) ao recém-nascido que garantem sua sobrevivência e desenvolvimento, uma vez
que no momento do parto ocorrem dramáticas transformações metabólicas e
fisiológicas.
Alguns hormônios envolvidos na gestação são também responsáveis pelo
crescimento e desenvolvimento da glândula mamária, em uma relação estreita entre
crescimento fetal e desenvolvimento mamário (mamogênese). Próximo do parto e
intimamente ligado aos eventos fisiológicos que causam a expulsão do feto, é iniciada a
função secretora por parte da glândula (lactogênese).
Zootecnicamente, o homem aproveitou o alto poder nutritivo da secreção láctea de
alguns mamíferos (bovinos, caprinos, búfalos), como fonte alimentar de alto poder
calórico, fácil obtenção e baixo custo, desenvolvendo na produção de leite uma indústria
de amplos níveis técnicos em todas as áreas envolvidas no processo.
Desenvolvimento da glândula mamária.
A glândula mamária, é uma glândula apocrina modificada, pertencendo portanto, às
glândulas cutâneas, mas com desenvolvimento funcional especial. Inicialmente, aparece
na forma de um engrossamento epidérmico conhecido como linha mamária que se
estende desde a região torácica até a região lombar. A linha mamária, por proliferação
celular, origina a cresta mamária, a qual se destaca sobre a superfície e oferece uma
dilatação regional. Na cresta, aparecem logo protuberâncias a intervalos definidos,
dando o aspecto de um cordão chamado eminência mamária. A cresta desaparece, o
*
Seminário apresentado na disciplina Bioquímica do Tecido Animal (VET00036) no Programa de PósGraduação em Ciências Veterinárias da UFRGS, pelo aluno RÓMULO CAMPOS GAONA no primeiro
semestre de 2001. Professor da disciplina: Félix H.D. González.
esboço da glândula mamária se desloca em direção ventral e as eminências mamárias
originam os processos mamários, característicos de cada espécie. O desenvolvimento do
mamilo vai acompanhado de transformações do mesênquima que rodeia a gema
mamária, a qual se eleva desde a circunvolução cutânea que a delimita. O duto
originado no broto primário está fechado ao exterior por um tampão córneo em nível do
extremo do mamilo. Posteriormente, este se desfaz e origina o duto papilar e a cisterna
láctea, em comunicação com o exterior. Até este momento, todos os eventos do
desenvolvimento embriológico ocorrem como resultado da expressão genética primária.
A glândula mamária fica em estado latente até a puberdade, momento em que
começam a formar-se os alvéolos verdadeiros a partir do sistema glandular juvenil,
quando é substituído o tecido adiposo de recheio por tecido glandular, apenas na fêmea,
ficando no macho a glândula em estado juvenil.
A partir da glândula mamária juvenil, os hormônios gonadotróficos hipofisiários
(FSH, LH) estimulam seu crescimento, ao tempo que a fêmea desenvolve sua maturação
sexual e regulariza sua ciclicidade estral. Entretanto, apenas ao momento da primeira
gestação, e por influência dos estrógenos e da progesterona, quando se desenvolve
plenamente o sistema de dutos lactíferos (efeito de estrógenos, hormônio do
crescimento e esteróides adrenais) e a diferenciação completa dos alvéolos (efeito da
progesterona e a prolactina).
O tecido glandular sofre desenvolvimento cada vez mais acentuado, deslocando por
completo o tecido adiposo e constituindo o parênquima. A camada epitelial de
revestimento e as células que a rodeiam se formam por diferenciação gradual dos
alvéolos. Todo o processo chama-se mamogênese, evento controlado de forma genética
e endócrina. Existem ao menos dois eventos que confirmam este sistema de controle:
primeiro, se o mesênquima mamário for transplantado para outro lugar, observa-se a
formação do botão mamário no lugar do transplante; segundo, é possível conseguir que
a glândula mamária em machos ou fêmeas tenha secreção ativa ao nascimento com a
administração exógena de hormônios na mãe.
O controle hormonal, no desenvolvimento da glândula mamária tem sido estudado
mediante extirpação de glândulas endócrinas, especialmente através de hipofisectomias,
adrenalectomias, isolamento de ilhotas pancreáticas, ovarectomias e tireoidectomias.
Aos animais tratados cirurgicamente lhes são fornecidas diversas combinações de
hormônios para medir o efeito das doses com relação ao crescimento e à diferenciação
dos dutos e do tecido alveolar. Tanto os estudos in vivo como in vitro têm demonstrado
o grande número de hormônios que estimula e controla o desenvolvimento da glândula
mamária. Por exemplo, estrógenos e progesterona exógenos podem causar crescimento
e desenvolvimento da glândula em animais ovarectomizados e hipofisectomizados, mas
o efeito é menor quando somente são animais ovarectomizados, o que demonstra a
influência na mamogênese de FSH e LH.
Terapias de substituição, como a suplementação de hormônios tireoidianos em
animais hipofisectomizados, mostram algum grau de resposta, especialmente na
secreção de esteróides ováricos, mas somente se a terapia incluir ao mesmo tempo
corticosteróides.
Os hormônios envolvidos no crescimento e diferenciação do sistema da glândula
mamária estimulam a síntese de DNA (estrógenos) ou a síntese de RNA e proteínas,
como a caseína, ação em que a prolactina participa ativamente.
Nesta fase de desenvolvimento fica claro que os estrógenos produzem crescimento
extensivo dos dutos, pois na ovarectomia se interrompe o crescimento alométrico da
glândula mamária, tornando-o isométrico. Também, nesta fase a placenta contribui com
estrógenos na regulação do crescimento, embora os níveis desses hormônios não devem
2
ser elevados, uma vez que se conhece que a administração exógena de estrógenos
resulta em glândulas mamárias pequenas.
Quanto à progesterona, o maior desenvolvimento da glândula ocorre quando suas
concentrações são altas, sendo a sua ação sinérgica com os estrógenos, juntos
promovendo a multiplicação de células epiteliais mamárias, ação produzida mediante a
redução do tempo de síntese de DNA mamário. À medida que a gestação avança, os
receptores celulares para estrógenos e progesterona passam progressivamente da forma
4s a 8s, mudança que é induzida pela prolactina. Existe um efeito regulatório sobre o
receptor de progesterona no citosol, pelo qual ele não está ativo durante a lactação, o
que explica porquê a administração exógena de progesterona nesse período não afeta o
desenvolvimento da glândula e, portanto, não induz síntese de leite.
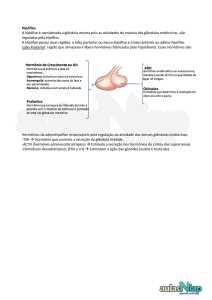
A hipófise participa na mamogênese através de vários hormônios como prolactina
(PRL), hormônio do crescimento (GH), hormônio adrenocorticotrópico (ACTH),
FSH/LH e hormônio estimulante da tiréoide (TSH). Essas participações têm sido
estudadas a partir da radioimunoanálise e outras técnicas altamente sensíveis
desenvolvidas nas últimas décadas. Também foi conhecido que a ação dos hormônios
não é individual mas sinérgica, especialmente regulada pela atividade das
gonadotrofinas (FSH/LH) que, por sua vez, induzem na gônada o desenvolvimento
folicular ou luteal, com a respectiva secreção de estrógenos e progestágenos
relacionados diretamente com o desenvolvimento da glândula.
Os hormônios em maior grau envolvidos com o processo de crescimento são a PRL e
o GH, atualmente classificados na família das somatotropinas e, aparentemente, com
efeito aditivo.
Em algumas espécies, um hormônio de origem placentária, denominado
genericamente como lactogênio placentário similar à PRL, também participa na
estimulação do crescimento da glândula. Em estudos específicos na fase de gestação, os
receptores das células mamárias para o lactogênio placentário deslocam os receptores da
PRL, o que explica parcialmente porquê as concentrações de PRL são baixas nos
primeiros quatro meses da gestação em bovinos e depois se elevam consideravelmente.
O GH não tem níveis circulantes elevados durante a gestação, tendo níveis
significativos apenas nas duas últimas semanas.
O hormônio adrenocorticotrópico (ACTH) não tem ação direta comprovada sobre a
glândula mamária, mas seus efeitos sobre o metabolismo geram efeito sobre a glândula.
Por isto, alguns corticóides aceleram o desenvolvimento mamário mediados pelo
aumento das poliaminas espermidina e espermina (derivados de metionina ou ornitina,
envolvidos no empacotamento do DNA). Similarmente ao GH, os níveis séricos de
cortisol são baixos durante a gestação e só aumentam dois a três dias antes do parto.
Os hormônios tireoidianos T3 e T4 tampouco possuem ação direta sobre a glândula
mamária para estimular seu crescimento, tendo efeitos secundários, embora o
hipotireoidismo detém o desenvolvimento da glândula em ratos, podendo reiniciar-se
com suplementação exógena desses hormônios.
O papel da insulina não é claro, pois ela estimula a mitose das células mamárias invitro, mas não é essencial para o crescimento in-vivo. Ainda sem conhecer a sua ação
específica, têm sido isolados receptores para insulina nas células da glândula.
Finalmente, os processos de crescimento da glândula mamária, ocorrem de forma
crítica no final da gestação, quando a profunda integração endócrina direciona os
processos metabólicos mediante ações sinérgicas diretas ou indiretas para obter
hipertrofia (síntese de RNA) e diferenciação celular (síntese de DNA) do sistema
glandular e do tecido de suporte (tecido conjuntivo), com o qual, a glândula estará
3
preparada para iniciar os processos de síntese bioquímica dos diferentes componentes
de sua secreção após o início da lactogênese.
GESTAÇÃO
ESTRÓGENO
PROLIFERAÇÃO
PROGESTERONA
CÉLULA MAMÁRIA
PLACENTA
ESTÍMULO
EXTEROCEPTIVO
SNC
ESTRÓGENO
PROGESTERONA
MANUTENÇÃO
CÉLULAS MAMÁRIAS
OVÁRIOS
EFEITO
SECUNDÁRIO
CORTICÓIDES T3/T4
GH
PRL
FSH/LH
HIPOTÁLAMO
RF
IF
HIPÓFISE
Figura 1. Resumo da participação endócrina no processo da mamogênese.
Considerações morfológicas da glândula mamária.
Existe na literatura abundante material com descrições detalhadas dos aspectos
anatômico-estruturais, de irrigação, inervação e de drenagem linfática da glândula
mamária que não é do caso analisar aqui. Entretanto, vale a pena salientar a estrutura
dos alvéolos com seu especial sistema de sustentação que na forma de rede invertida
permite que a secreção láctea flua à cisterna da glândula, a qual é capaz de armazenar
100 a 400 ml de leite e serve como ponto de acumulação dos dutos principais (10-20),
os quais se ramificam repetidamente para finalizar nos alvéolos, verdadeiro tecido
secretor do leite.
O alvéolo, pode considerar-se como unidade funcional da glândula mamária. Está
composto por uma linha de células epiteliais rodeada de tecido conjuntivo e por células
mioepiteliais e vasos sangüíneos. As células epiteliais são capazes de extrair nutrientes
desde o sangue, sintetizar componentes do leite (gordura, proteína, lactose) e secretar
leite para a luz do alvéolo. As células mioepiteliais são células musculares
especializadas que envolvem os alvéolos e os dutos menores e se contraem na ejeção do
leite.
Dentro dos componentes celulares, a membrana celular possui ampla especialização
e, aparentemente, tem elementos tisulares únicos que lhe permitem drásticos trabalhos
de filtração. Para produzir um litro de leite, a glândula deve filtrar cerca de 500 litros de
sangue a uma taxa de filtração de 800 litros/hora. Também permite o trabalho de ligação
de receptores de membrana ou ativação de receptores de segundos mensageiros. O
retículo endoplasmático sintetiza proteína e esterífica ácidos graxos, e o aparelho de
Golgi sintetiza lactose e processa proteína.
A inervação autônoma da glândula mamária só conta com o sistema simpático, não
encontrando-se fibras nervosas parassimpáticas. O sistema eferente simpático inerva os
músculos lisos que circundam os vasos sangüíneos, os túbulos coletores e os esfíncteres
4
do mamilo. A estimulação do mamilo envia informação aferente ao sistema nervoso
central, que é recebida na hipófise posterior, onde se libera ocitocina e se inicia a ejeção
do leite, mediante contração das células mioepiteliais. A hipófise anterior libera outros
hormônios necessários para manter a galactopoiese. Quando ocorre excitação do animal,
a adrenalina diminui o fluxo de sangue para a glândula mamária e reduz a saída de leite.
Foi postulada a expressão de elementos na glândula de forma independente em cada
quarto da glândula, quanto ao volume produzido sob um mesmo nível sangüíneo de
hormônios ou de metabolitos circulantes.
Lactogênese: generalidades fisiológicas.
A lactogênese é o início da secreção de leite. A prolactina (PRL) desempenha um
importante papel neste processo, liberando-se quando o mamilo é manipulado, seja pelo
neonato (sucção) ou mediante estimulação mecânica ou manual (ordenha).
Os estímulos sensoriais são levados ao hipotálamo, onde se sintetiza PRF (Fator de
Liberação de Prolactina) e se secreta a dopamina, principal fator de inibição da
prolactina, conhecido também como PIF (Fator Inibidor da Prolactina). Há evidência
que o VIP (Peptídeo Intestinal Vasoativo) induz a secreção de PRF.
Dois fatores possuem estreita relação com a liberação de PRL: em primeiro lugar,
requer um tempo após o início da remoção do leite para atingir o pico máximo de
secreção do hormônio, normalmente de 30 minutos. Em segundo lugar, é necessária a
remoção permanente do leite em intervalo próximo a 12 horas para estimular a liberação
de PRL. Os valores de PRL em circulação diminuem na medida que a lactação avança.
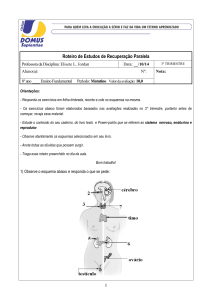
Regulação endócrina da lactogênese.
Dentro do complexo processo de síntese láctea, a regulação hormonal induz, modula
ou bloqueia os diferentes eventos relacionados com a lactogênese. Os requerimentos
endócrinos específicos variam entre as espécies, mas em geral as necessidades mínimas
incluem PRL, glicocorticóides, ausência relativa de progesterona, hormônio do
crescimento, parathormônio, calcitonina e ocitocina.
Desde a década de 1960, se conhece que PRL, ACTH e estrógenos iniciam a lactação
e que a progesterona inibe a lactogênese, o que explica porquê a secreção láctea
aumenta gradualmente depois do parto, uma vez que os níveis séricos de progesterona,
de origem luteal ou placentária caem.
Basicamente se requerem três fatores para que a ação endócrina possa controlar a
lactação: o número de células mamárias, a atividade sintética das células epiteliais
mamárias e a manutenção de um eficiente reflexo para a ejeção do leite. Na Figura 2, se
mostra um esquema resumido da interação endócrina no processo da lactação.
Prolactina (PRL).
Possui um peso molecular de 32 kDa, com estrutura similar ao hormônio do
crescimento (GH). É um hormônio protéico, com receptor na membrana. É produzido
nos lactotropos (células acidófilas) da hipófise anterior, cujo número aumenta
dramaticamente durante a fase final da gestação.
A PRL está envolvida na iniciação e manutenção da lactação. Ela tem um papel
importante na diferenciação das células da glândula mamária e controla os passos
bioquímicos envolvidos na síntese do leite. Induz a acumulação de mRNA da caseína,
estimulando a expressão de genes desta proteína e provavelmente de outros genes. Os
receptores para prolactina na glândula mamária aumentam paralelamente com o
aumento da secreção de PRL no período do peri-parto. A PRL não atua sozinha, mas de
forma sinérgica com outros hormônios. Estudos in-vitro têm demonstrado que a PRL
5
produz aumento na membrana da mitocôndria e induz a síntese de α-lactalbumina,
lactose e gordura.
HIPOTÁLAMO
RF/IF
HIPÓFISE
ANTERIOR
GH/PRL
TSH
FSH/LH
ACTH
ADRENAL
glicocorticóides
POSTERIOR
TIRÉOIDE
T3/T4
PTH/CT
OT
OVÁRIO
E2/P4
INSULINA/GLUCAGON
PROLIFERAÇÃO E REGULAÇÃO DAS CÉLULAS MAMÁRIAS
Figura 2. Resumo da interação endócrina na lactação.
As concentrações séricas de PRL têm correlação positiva mas de baixo nível com a
secreção de leite em bovinos, não sendo possível aumentar a produção de leite mediante
administração exógena de PRL. Igualmente, foi encontrado que a supressão da secreção
de PRL durante uma lactação já estabelecida não afeta a produção de leite. Portanto, a
PRL pode ser importante, porém não limitante para a manutenção da lactação. A
hipofisectomia no início da lactação resulta em completa interrupção da secreção de
leite.
Várias drogas e compostos biológicos naturais reduzem a secreção de PRL, entre os
quais estão a dopamina, a bromocriptina e os alcalóides derivados do “ergot”. O
bloqueio da PRL ocorre em nível de sua secreção e não no receptor, o que foi
evidenciado porque o único hormônio que bloqueia o receptor de PRL de forma
específica é a progesterona. Igualmente, o bloqueio da PRL, não ocasiona inibição total
da lactação, mas apenas a reduz em cerca de 45%.
Para a sua ação biológica, a PRL ativa seu receptor, se internaliza na célula e causa
indução da expressão gênica. Posteriormente, o complexo receptor-hormônio é
degradado nos lisossomos.
Com relação à PRL, há estudos sobre seus níveis de secreção, sua participação na
lactação, o grau de inibição por bloqueadores farmacológicos ou pela ação de outros
hormônios (progesterona, lactogênio placentário), que mostram marcadas diferenças
conforme a espécie estudada. A secreção de PLR é estimulada por estímulos de sucção.
Em geral, em ruminantes a ação da PRL é muito mais intensa que nos roedores.
6
Lactogênio placentário (LP).
Tem sido encontrado nas diferentes espécies. Sua secreção começa desde poucos
dias após a fecundação até metade da gestação. Possui atividade luteotrópica e
lactogênica, embora esta última não suficientemente elucidada. Os efeitos metabólicos
são similares aos ocasionados pelo hormônio do crescimento (GH).
Os níveis circulantes de lactogênio placentário caem à medida que o parto se
aproxima. Contudo, no primeiro estágio da lactação ainda persistem alguns níveis tão
baixos (de forma prática indetectáveis), que há dúvida que por si só possam ter
atividade lactogênica.
Diferentes lactogênios placentários (ovino, humano) apresentam atividade
lactogênica em cultivos de células mamárias in-vitro. A injeção de lactogênio
placentário humano em coelhos ocasiona secreção de caseína, tendo, portanto, atividade
lactogênica, porém muito inferior àquela desenvolvida pela PRL.
A ação do lactogênio placentário é mediada através do receptor da PRL. Por si só o
lactogênio bloqueia o receptor ocupando seu sítio de ativação, mas esta ação é de maior
nível se existe progesterona em circulação.
Acredita-se que os LP têm maior ação no processo de mamogênese que no de
lactogênese, o que explica sua ação similar ao hormônio do crescimento. Conhece-se
que o lactogênio placentário bovino é secretado primariamente na circulação fetal, mas
o significado fisiológico deste fato não se sabe.
A informação existente sobre os lactogênios placentários indica que eles teriam uma
menor importância na lactogênese.
Hormônio do crescimento (GH).
O GH é um polipeptídio simples (uma cadeia) com peso próximo a 22 kDa em todos
os mamíferos. Sintetizado nos somatotrofos (cédulas acidófilas da hipófise),é o mais
abundante dos hormônios hipofisiários e liberado por estímulo hipotalámico. É
considerado como hormônio pertencente ao grupo das somatotropinas junto com a PRL
e os LP.
Os hormônios somatotrópicos são um grupo de hormônios protéicos com homologia
na sua seqüência, contendo entre 190-199 aminoácidos, com resíduos de triptofano e
duas pontes dissulfeto, provenientes de um mesmo gene ancestral, embora a sua
expressão precisa de múltiplos genes.
O receptor para GH é membro da citocina-hematopoietina, cuja união ativa
diferentes vias que atuam como sinais ativos em processos específicos como
fosforilação e transcrição de genes, ativação da MAP-quinase, fosforilação e ativação da
PI3-quinase e ativação da proteína C-quinase.
O GH é essencial no crescimento pós-natal e no metabolismo de lipídios, proteínas e
minerais. Todos os efeitos são mediados por IGFI e IGFII. O GH aumenta o transporte
de aminoácidos nas células musculares, favorece a síntese de proteína, induz a síntese
de RNA/DNA nos tecidos, provocando, em geral, um balanço energético positivo.
No metabolismo dos carboidratos, o GH antagoniza os efeitos da insulina, sendo
hiperglicemiante, aumenta a glicogenólise e, com isto, dando maior aporte de glicose
aos tecidos. No metabolismo dos lipídios, promove a liberação de ácidos graxos e
glicerol desde o tecido adiposo, aumenta a circulação de ácidos graxos livres e aumenta
a oxidação desses ácidos no fígado. Ocasiona aumento dos valores séricos de cálcio,
magnésio e fosfato, com ação sobre o osso. Também causa retenção de sódio.
Por ter ampla participação em todas as vias metabólicas relacionadas com a
lactogênese é talvez o hormônio com maior incidência direta sobre a lactação, atuando
de forma sinérgica com PRL, ACTH e hormônios tireoidianos.
7
Conhece-se que o GH, junto com a PRL e os corticóides estão em pequenas
quantidades na circulação durante grande parte da gestação. A progesterona freia sua
ativação, mas permanecendo níveis basais. Pouco antes do parto começam mudanças
nos níveis circulantes com pico significativo ao redor do parto.
Desde 1937 se conhece que o GH é, claramente, galactopoiético em ruminantes, o
que motivou sua síntese por DNA recombinante e sua produção industrial. Foi
comprovado que o GH pode aumentar entre 6 a 35% a produção de leite, dependendo da
forma a freqüência da suplementação exógena e das condições ambientais externas.
A administração de GH, em vacas com balanço energético negativo (BEN), aumenta
a quantidade de gordura no leite e causa diminuição dos níveis de proteína, sem afetar
os níveis de lactose. No leite de animais tratados com GH há maior quantidade de
ácidos graxos de cadeia média e longa que de cadeia curta e a α-lactalbúmina aumenta
em até 32%. Em nível hepático, o GH estimula a capacidade do fígado para metabolizar
propionato.
Em contraste, quando a suplementação de GH é feita em animais em balanço
energético positivo, não ocorrem mudanças na secreção láctea, o que indica que o GH
modula efetivamente a homeorrese, direcionando a distribuição de nutrientes,
favorecendo o uso de metabolitos pela glândula mamária.
O uso prolongado de GH, natural ou recombinante, não produz mudanças a longo
prazo nas características químicas da secreção (gordura, proteína, lactose). O GH
produz mudanças no balanço energético nos animais suplementados, embora, após um
período de adaptação, aumenta o consumo voluntário de alimentos retornando a um
balanço energético positivo após 70 dias.
As concentrações basais de GH e de TRH aumentam no início da lactação quando a
glândula sintetiza volumes crescentes de leite. Foi comprovado estímulo positivo do
GRH (Fator de Liberação do GH, de origem hipotalámica) em qualquer fase da
lactação.
Os mecanismos mediante os quais o GH atua em ruminantes envolvem mudanças
coordenadas no metabolismo de gordura, proteína e carboidratos na glândula mamária.
Entretanto, o GH, não atua modificando as taxas de digestibilidade de nutrientes, nem
os requerimentos para manutenção corporal. As mudanças energéticas se associam ao
aumento metabólico necessário para compensar o aumento na síntese láctea. O anterior
confirma a hipótese de que o GH atua mediante modificação pós-absortiva de
nutrientes.
Diversos trabalhos têm estudado os mecanismos de ação do GH. Ele aumenta a taxa
de uso de glicose para sua conversão em lactose, sem que os níveis basais de glicose
abaixem, o que consegue mediante redução da oxidação da glicose em outros tecidos.
Também, o GH participa na mobilização de lipídios de reserva, o que tem como
conseqüência o aumento de ácidos graxos de cadeia longa no leite. Com relação aos
minerais, o GH não modifica os níveis de cálcio e fósforo na secreção.
As modificações que ocasiona o GH no organismo são, no geral, para produzir
mudanças homerrésicas que garantam o aumento da secreção. Entretanto, o hormônio
não se une especificamente a receptores na glândula mamária somente. De fato, se une
com maior especificidade a receptores para GH no tecido hepático, mas ainda assim,
exerce efeitos galatopoiéticos significativos.
Tem sido postulado que a somatomedina C (IGFI) pode mediar a resposta
galatopoiética do GH, através de receptores específicos localizados tanto na glândula
mamária como nos tecidos responsáveis pelo fornecimento de metabolitos precursores
dos diversos componentes da secreção (fígado, tecido adiposo).
8
Glicocorticóides.
Diversos trabalhos têm demonstrado que os glicocorticóides têm um papel
importante no processo de lactogênese. Na célula alveolar, o cortisol induz a
diferenciação do retículo endoplasmático rugoso e do aparelho de Golgi, participando
no metabolismo de carboidratos e no balanço eletrolítico.
A adrenalectomia bloqueia a síntese de proteínas (caseína) e de RNA que
normalmente ocorre durante a lactogênese. Por isto, se deduz que o papel dos
corticóides adrenais é importante durante toda a lactação. Os glicocorticóides começam
a liberar-se, de forma crescente, no final da gestação. Mas os receptores para eles na
glândula mamária estão bloqueados pela progesterona. Assim que os níveis dela caem,
os corticóides tornam-se ativos. Adicionalmente, a globulina, que é a proteína
transportadora dos corticóides no sangue, aumenta também no final da gestação.
Têm sido encontrados maiores níveis circulantes de cortisol em vacas lactantes do
que em não lactantes. Também, o número de receptores para cortisol na glândula
mamária ativa supera em até quatro vezes o número de receptores na glândula mamária
inativa, evidencia que dá suporte adicional sobre o papel dos corticóides na lactação.
Por outra parte, quando se administram glicocorticóides exógenos em grandes
quantidades, a síntese de leite é bloqueada.
Hormônios tireoidianos.
Os estudos sobre efeitos diretos dos hormônios tireoidianos na lactação, têm
mostrado resultados contraditórios. Não se encontrou efeito direto dos hormônios sobre
as células da glândula mamária, embora alguns trabalhos têm demonstrado que a
triiodotironina (T3) afeta a síntese de DNA nas células epiteliais mamárias.
A administração de caseína iodada, que possui atividade como tiroxina (T4) aumenta
entre 10-30% a produção de leite, dependendo do momento de sua aplicação, sendo
mais efetiva no final da lactação. Todavia, o aumento da produção de leite ocasiona
maiores necessidades de nutrientes, ocasiona perda de peso, aumento do gasto cardíaco,
maior consumo de oxigênio e aumento da temperatura corporal.
Em diversos estudos se encontrou correlação negativa entre os níveis circulantes de
hormônios tireoidianos e a produção de leite. Também, se conhece que a atividade de T3
e de T4 varia conforme a temperatura ambiental e o conteúdo energético da dieta.
Assim, vacas em balanço energético negativo não respondem a tiroxina exógena, o que
sugere que as mudanças na produção de leite estão mais relacionadas com efeitos de
homeorrese do que com uma ação única da tiroxina.
A aplicação de TRH, produz liberação de GH, PRL e TSH, aumentando a produção
de leite, o que sugere um trabalho sinérgico entre os diferentes hormônios na lactação.
Durante a lactação são excretadas grandes quantidades de iodo no leite, o que pode
ocasionar redução nos níveis de T3 e T4, ocasionando um estado de hipotireoidismo
temporário durante a lactação. Em recentes estudos se encontrou na glândula mamária
um inibidor de hormônios tireoidianos em animais gestantes e em não lactantes, o que
explica parcialmente o papel dos hormônios tireoidianos como hormônios
galactopoiéticos. É possível, que a ação de T3 sobre a lactação não seja especificamente
sobre as células epiteliais da glândula mamária, mas de forma indireta através do
metabolismo basal e da homeorrese.
Calcitonina.
A calcitonina é um peptídio de 32 aminoácidos, sintetizado nas células C da glândula
tiréoide como pró-calcitonina. A calcitonina regula os níveis de cálcio, sendo secretada
quando existe aumento dos níveis circulantes, prevenindo o aumento sérico de cálcio e
9
de fósforo durante a lactação, por inibição da ressorção óssea. Os níveis de calcitonina
se elevam ao redor do parto, mas não existe evidência se sua depleção bloqueia a
secreção normal de leite.
Hormônio da paratiréoide (PTH).
A ação do paratohormônio (PTH) é contrária àquela da calcitonina, aumentando
portanto, os níveis de cálcio sangüíneo. A hipocalcemia estimula a liberação do
hormônio.
No peri-parto aumentam os níveis séricos do hormônio paratireoidiano que mobiliza
de forma drástica cálcio desde o osso e estimula a atividade da 1,25 di-hidroxi-vitamina
D3, al qual aumenta a absorção de cálcio desde o intestino. Durante a lactação, e devido
às altas taxas de secreção de cálcio no leite, a paratireoidectomia reduz a produção de
leite.
Insulina e glucagon.
Os efeitos da insulina, de forma específica sobre a glândula mamária, não são claros.
De fato, a insulina induz a utilização de glicose pelas células, mas, a glândula mamária
pode utilizar glicose, acetato, β-OH-butirato e triglicerídeos de forma independente da
insulina.
Acredita-se que a insulina atua na lactação modulando a homeorrese, estimulando a
síntese de proteína e lipídios e aumentando a utilização de acetato no tecido adiposo. A
glândula mamária requer níveis basais de insulina.
O glucagon atua de forma oposta à insulina, estimulando a glicogenólise e a
gliconeogênese. Os níveis de glucagon aumentam entre os dias 5 a 30 da lactação, o que
sugere um papel de controle do metabolismo energético durante o início da lactação.
Sabendo que os níveis de insulina, glucagon, GH e T4 se elevam no período do parto
(duas semanas antes até um mês depois) isto se associa a sua participação no controle da
lactação mediante o controle da absorção/utilização de nutrientes, da homeorrese e do
metabolismo basal.
Esteróides ováricos.
Estabelecida a lactação, a ovarectomia não afeta a produção de leite de forma direta.
Todavia, os estrógenos a diminuem de forma temporária, o que se evidencia com a
queda de produção ao redor do cio. Existem receptores para estrógenos no tecido
mamário. Estes hormônios, devido a sua estrutura esteróide (lipídica) se unem à gordura
do leite, sendo secretados nele.
O bloqueio dos receptores para estrógenos ou sua inibição, ocasiona redução no
reflexo da ejeção, talvez mediada pela redução da ação das células mioepiteliais e das
células contráteis dos alvéolos.
A progesterona tem sido associada ao bloqueio da lactação. Os receptores de
progesterona no tecido mamário são 75% menos em vacas lactantes do que em não
lactantes. Pela alta afinidade da molécula de progesterona pela gordura, ela pode ser
seqüestrada e excretada no leite anulando a sua função biológica na glândula.
Os estrógenos ováricos têm sido usados para a indução da lactação mediante a sua
administração exógena em doses controladas. Se utiliza em animais cuja lactação está
no fim e estão gestantes. Procura-se obter produção de leite com rentabilidade
econômica, em animais de alta produção, porém com falhas reprodutivas. O volume
obtido nas lactações induzidas atinge 70% do volume de uma lactação normal.
10
Ocitocina (OXT).
A ocitocina se sintetiza no hipotálamo e se armazena na hipófise posterior, sendo
similar quanto a sua composição química ao hormônio antidiurético (ADH). Sua
meia-vida é curta (2-4 minutos), possui ação sobre o músculo liso e sobre as células
mioepiteliais na glândula mamária.
A ocitocina se considera o hormônio da ejeção do leite, requisito básico para a
lactogênese. Além disso, é considerado como hormônio galactopoiético. Encontram-se
maiores valores basais de OXT no início da lactação do que no final. A secreção de
ocitocina é inibida pela adrenalina.
BIBLIOGRAFIA
Bauman, D. E. 1992. Bovine somatotropin: review of an emerging animal technology. J. Dairy Sci.
66:371-380.
Collier, D, et al. 1984. A review of endocrine regulation of metabolism during lactation. J. Animal Sci.
59:498-510.
Forsyth, I.A. 1989. Growth factors in mammary gland function. J. Reprod. Fert. 85:759.770.
Hurley, W. L. 2002. Lactation Biology: General References. Departament of Animal Sciences, University
of Illinois, Urbana.
Jensen, R.G., Ferris, A.M. & Lammi-Keefe, C.J. 1991. The composition of milk fat. J. Dairy Sci.
74:3228-3243.
Kronfeld, D.S. 1982. Major metabolic determinants of milk volume, mammary efficiency and spontaneus
ketosis in dairy cows. J. Dairy Sci. 65:2205-2212.
McDonald, L. 1989. Endocrinología Veterinaria y Reproducción. Cuarta Edición. InteramericanaMcGraw-Hill, México.
Plaut, K. 1993. Role of Epidermal Growth Factor and transforming growth factors in mammary
development and lactation. J. Dairy Sci. 76:1526-1538.
Tucker, A., 1988. Lactation and Its Hormonal Control. In: The Physiology of Reproduction. Knobill &
Neill Editors. Reven Press, New York.
11