Gado de Leite
Inseminação Artificial em
Tempo Fixo (IATF)
Em Bovinos Leiteiros
Erick Fonseca de Castilho
C A P Í T U L O
2
2. Fisiologia reprodutiva da fêmea bovina
2.1 Introdução
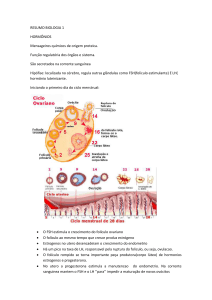
Ao longo do tempo, o controle da reprodução dos mamíferos deixou de ser considerado como regulado apenas pelo sistema nervoso central (SNC), passando
a ser visto como uma função controlada por dois sistemas separados: o sistema
nervoso central e o sistema endócrino (Figura 15). A seguir, descobriu-se que
o hipotálamo ligava esses dois sistemas por meio do sistema porta vascular
hipotálamo-hipofisário, coordenando as funções das gônadas.
Entretanto, muitos fenômenos não podiam ser explicados exclusivamente pelo
controle neuroendócrino. Portanto, nos últimos anos testemunhou-se a descoberta de determinados mensageiros químicos (fatores de crescimento), bem como
a presença de sistemas regulatórios parácrinos e autócrinos nas gônadas. Tais
avanços ajudaram a desvendar fenômenos que não conseguiram ser explicados
apenas pelo controle neuroendócrino.
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
Figura 15:. Cronologia da modificação dos conceitos sobre o controle da reprodução
em mamíferos.
»» (A) O SNC atua através das vias nervosas (controle nervoso).
»» (B) A descoberta do sistema endócrino (SE) mudou a ênfase do
SNC para controle endócrino.
»» (C) Mais tarde, o reconhecimento do sistema porta hipotalâmicohipofisário propiciou a rota para hormônios hipotalâmicos controlarem a hipófise anterior (controle neuroendócrino).
»» (D) os reguladores intragonadais ou fatores de crescimento, agindo via mecanismos autócrinos/parácrinos, modulam a secreção
dos hormônios gonadotróficos hipofisários. FONTE: HAFEZ e HAFEZ
(2004).
42
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
Ambos os sistemas, nervoso e endócrino, funcionam no sentido de iniciar, coordenar ou regular as funções do sistema reprodutivo. Diferentemente do sistema
nervoso, que controla as funções orgânicas por meio de impulsos nervosos elétricos como, por exemplo, a regulação musculoesquelética, o sistema endócrino
utiliza mensageiros químicos ou hormônios para regular processos orgânicos
mais lentos, como o crescimento e a reprodução.
Um hormônio é classicamente definido como uma substância química fisiológica,
orgânica, sintetizada e secretada por uma glândula endócrina sem ducto e transportada via corrente sanguínea. Os hormônios têm a ação básica de estimular,
inibir ou regular a atividade funcional de seus órgãos ou tecidos-alvo. Entretanto,
órgãos como o útero e hipotálamo produzem hormônios que não se enquadram
nessa definição clássica.
Além dos hormônios das glândulas endócrinas, os “fatores de crescimento” são
substâncias peptídicas relacionadas aos hormônios e que controlam o crescimento e o desenvolvimento de diversos órgãos, tecidos e células em cultivo. Diferentemente dos hormônios, os fatores de crescimento são produzidos e secretados
por células de diferentes tecidos para se difundirem por meio das células-alvo.
Os objetivos deste capítulo são:
»» Discutir os conceitos de hormônios, bem como seus tipos e modos
regulatórios;
»» Discutir a anatomia funcional das glândulas endócrinas e seus
principais hormônios envolvidos no controle reprodutivo das fêmeas bovinas;
»» Discutir a importância do eixo-hipotalâmico-hipofisário-gonadal na
O portal do agroconhecimento
43
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
regulação hormonal reprodutiva;
»» Descrever a cronologia da ocorrência da puberdade em fêmeas
bovinas;
»» Descrever os eventos celulares e endócrinos durante a foliculogênese, maturação oocitária e ovulação regulação hormonal e sua
importância na fisiologia reprodutiva dos bovinos.
»» Discutir a endocrinologia do ciclo estral das fêmeas bovinas.
2.2 Hormônios
Atualmente se conhecem mais de 50 hormônios. Na Tabela 2.1 se relacionam
os principais hormônios com efeito na função reprodutiva. Existem quatro grupos
químicos de hormônios da reprodução: peptídeos, esteroides, ácidos graxos e
aminas. Os vários tipos de hormônios têm diferentes características quanto a sua
forma de síntese, armazenagem, meia-vida, forma de transporte no sangue e
mecanismo de ação.
Tabela 2: Principais hormônios que agem na função reprodutiva
feminina.
Hormônio
Órgão secretor
Órgão-alvo
Principal ação
GnRH
Hipotálamo
Adeno-hipófise
Liberação de LH
PIF
Hipotálamo
Adeno-hipófise
Inibe a liberação
de PRL
PRF
Hipotálamo
Adeno-hipófise
Liberação de PRL
Prolactina
Adeno-hipófise
Glândula mamária
Favorece a lactação
44
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
Hormônio
Órgão secretor
Órgão-alvo
Principal ação
FSH
Adeno-hipófise
Folículo ovariano
Maturação folicular
LH
Adeno-hipófise
Ovário
Ovulação, manutenção do corpo
lúteo e maturação
oocitária
Ocitocina
Corpo lúteo
Miométrio
Favorece o parto
Ocitocina
Hipotálamo
Glândula mamária
Favorece a descida
do leite
Tiroxina
Tireoide
Todas as células
Aumento do metabolismo
Triiodotironina
Tireoide
Todas as células
Aumento do metabolismo
Estrógeno
Ovário
Órgãos sexuais
secundários
Função cíclica ovariana e caracteres
sexuais
Estrógeno
Ovário
Glândula mamária
Desenvolvimento
Progesterona
Ovário
Glândula mamária
Desenvolvimento
mamário
Progesterona
Ovário
Útero
Manutenção da
gestação
Relaxina
Ovário
Sínfise pubiana
Relaxamento do
canal do parto
hCG
Placenta
Ovário
Similar ao LH
eCG
Placenta
Ovário
Similar ao FSH
Lact. plac.
Glândula mamária
Glândula mamária
Similar ao hormônio
do crescimento
(GH) e PRL
PGF2α
Miométrio
Corpo lúteo
Luteólise
PGF2α
Miométrio
Miométrio
Contração uterina
O portal do agroconhecimento
45
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
Os hormônios peptídicos podem ter desde 3 até 200 resíduos de aminoácidos,
com peso molecular variando entre 300 e 70.000 dáltons e constituem o grupo
de hormônios mais numeroso. Os principais órgãos que produzem hormônios
peptídicos são o hipotálamo, a hipófise, as ilhotas pancreáticas, a placenta, a
glândula paratireoide e o trato gastrointestinal. Exemplos: FSH, LH e ocitocina.
Os hormônios esteroides são derivados principalmente do colesterol, com peso
molecular entre 300 e 400 dáltons. São produzidos pelo córtex adrenal, pelas
gônadas e pela placenta, e incluem os corticosteroides, os estrógenos, os andrógenos e a progesterona. Neste grupo está incluída a forma hormonal da vitamina
D (1,25-dihidroxi-colecalciferol).
Os hormônios do grupo das aminas são derivados da tirosina ou do triptofano.
Incluem as catecolaminas, que são produzidas pela medula adrenal e algumas
células nervosas, e as iodotironinas, derivadas do aminoácido tirosina, que são
produzidas exclusivamente pela tireoide. A melatonina também se enquadra no
neste grupo, sendo produzida a partir do triptofano.
Finalmente, os ácidos graxos (eicosanoides) incluem as prostaglandinas, os leucotrienos e os tromboxanos, compostos derivados do ácido araquidônico, produzidos em quase todos os tecidos e com peso molecular em torno de 400 dáltons.
Na função reprodutiva são importantes a PGF2α e a PGE2.
2.2.1 Modos de comunicação intercelular
O sistema nervoso central era considerado o principal dos sistemas orgânicos
até a descoberta das glândulas endócrinas. A partir daí, compreendeu-se que a
regulação da reprodução era compartilhada por dois sistemas distintos, com o hipotálamo servindo de interface entre ambos. Atualmente, mensageiros químicos
46
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
que não se encaixam em nenhum dos dois sistemas, os fatores de crescimento,
estão sendo descobertos como importantes fatores no controle da reprodução.
As células se comunicam entre si por meio de mensageiros químicos, como
peptídeos, esteroides, aminas e ácidos graxos. Portanto, existem quatro modos
de comunicação intercelular: endócrina, parácrina, autócrina e neural (Figura 16).
Figura 16: Modos de comunicação intercelular. Fatores de crescimento produzidos
localmente agindo de diferentes modos nas células-alvo. Fonte: HAFEZ e HAFEZ
(2004).
2.2.1.1 Hormônios do sistema endócrino hormonal
No sistema endócrino, o hormônio é sintetizado e armazenado em células especializadas de uma glândula anatomicamente definida. Estes hormônios são
liberados na corrente sanguínea e transportados (geralmente por proteínas
específicas de transporte) a um órgão-alvo, geralmente distante da origem.
O sistema endócrino inclui glândulas secretórias que liberam seus hormônios na
circulação geral (por exemplo, a insulina) ou ainda em sistemas de circulação
fechada (como o GnRH).
O portal do agroconhecimento
47
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
2.2.1.2 Hormônios parácrinos
São denominados parácrinos os hormônios que influenciam células ou órgãos em
sua vizinhança imediata. Por exemplo, os andrógenos produzidos pelas células da
teca interna, para agir nas células da granulosa adjacentes no interior do folículo
ovariano.
2.2.1.3 Hormônios autócrinos
Um processo autócrino se refere ao mecanismo em que a célula produtora é
também a célula-alvo. Exemplo: prostaglandinas.
2.2.1.4 Neurotransmissores
Os neurotransmissores estão sendo frequentemente considerados hormônios nos
dias de hoje, isto é, são mensageiros hormonais. Neurotransmissores como a
acetilcolina podem também ser considerados hormônios parácrinos.
O conhecimento a respeito das funções endócrinas é bem maior do que em
relação ao restante do sistema hormonal. Nos últimos anos, pesquisadores têm
dado mais atenção às funções parácrinas e endócrinas, mas muitos detalhes
ainda são pouco compreendidos.
Após atingir uma célula-alvo, o hormônio deve provocar uma reação, ativada pelos receptores específicos da célula-alvo. Os receptores são estruturas moleculares com alta afinidade específica para uma determinada configuração hormonal.
Assim, os receptores exercem duas importantes funções:
48
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
a) Reconhecimento do hormônio específico pela célula-alvo;
b) Tradução do sinal em uma resposta celular específica.
A estrutura bioquímica dos receptores hormonais pode variar, mas em geral, cada
um deles pode reconhecer e interagir com uma unidade hormonal altamente
específica (em contraste com o modelo chave-fechadura que rege a interação
substrato-enzima).
Todos os receptores apresentam dois componentes chave:
»» um domínio de ligação que se liga especificamente ao hormônio
correspondente;
»» um domínio efetor, que reconhece o complexo formado pela ligação entre o domínio de ligação e o hormônio, e ativa a resposta
biológica específica da célula, a qual geralmente envolve a ativação ou desativação de enzimas nas células-alvo.
Os receptores de hormônios esteroides (lipofílicos) são geralmente encontrados
no citosol e no núcleo das células-alvo, onde interagem diretamente com o DNA.
Já os receptores para peptídeos e hormônios proteicos (lipofóbicos) geralmente
se localizam na membrana externa da célula. A maioria dos receptores, especialmente os da membrana celular, requer um segundo mensageiro para transmitir
a mensagem. Um dos segundos mensageiros mais conhecidos é o AMP cíclico
(cAMP), representado na Figura 17. Após ligar-se ao receptor, o hormônio ativa
o sistema adenilciclase na membrana celular.
O ATP é então convertido em AMP cíclico. O segundo mensageiro cAMP, por
sua vez, ativa a cAMP-proteína-quinase-A que se torna uma unidade catalítica
O portal do agroconhecimento
49
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
ativa e uma unidade regulatória. A unidade catalítica ativa da proteína-quinase
estimula a fosforilação de uma proteína ou enzima, que provoca então os efeitos
celulares, como síntese proteica, crescimento ou secreção hormonal. Como as
concentrações hormonais circulantes geralmente são baixas, o receptor precisa
ter um mecanismo de captura muito eficiente.
Figura 17: AMP cíclico como segundo mensageiro. FONTE: INTERVET (2006).
O efeito da secreção de um hormônio endócrino pode variar, conforme a circunstância particular. O número e o tipo de receptores de uma célula-alvo não
são fixos, e sua formação e degradação é um processo dinâmico. A função de
um hormônio em uma célula pode ser a indução ou a degradação de receptores
para outro mensageiro. Além disso, os receptores podem ser bloqueados por
50
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
uma quantidade excessiva de hormônio. Neste caso, a superestimulação por
doses maiores que a dose normal não irá potencializar o efeito. Muitas condições
patológicas no processo reprodutivo são causadas por distúrbios em receptores
hormonais.
2.2.2 Regulação da secreção hormonal
O sistema nervoso desempenha papel essencial na regulação da atividade gonadal por meio do mecanismo de retroalimentação endócrina das vias neurais e
do controle imunoendócrino.
2.2.2.1 Retroalimentação endócrina
2.2.2.1.1 Gônadas
Um hormônio da glândula-alvo (por exemplo, o estrógeno) pode influenciar a
secreção do estímulo trófico que causou sua liberação (por exemplo, FSH). O
controle por retroalimentação (feedback) ocorre no nível do hipotálamo e da
hipófise (Figura 18). Dependendo de suas concentrações na corrente circulatória,
os hormônios esteroides podem exercem uma retroalimentação estimulatória
(positiva) ou inibitória (negativa).
O portal do agroconhecimento
51
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
Figura 18: Complexo hipotalâmico-hipófise. Células nervosas hipotalâmicas liberando neuro-hormônios nos vasos porta para transportar à hipófise anterior via vasos
hipotalâmicos-hifofisários. Partículas sólidas nas células nervosas representam
neuro-hormônios. O sangue é transportado pelo sistema venoso retrógrado de
volta para o hipotálamo. FONTE: HAFEZ e HAFEZ (2004).
2.2.2.1.2 Retroalimentação inibitória ou negativa (Feedback
negativo)
Esse sistema envolve inter-relações recíprocas entre duas ou mais glândulas e
órgãos-alvo. Por exemplo, a estimulação dos ovários pelo FSH eleva a secreção
de estrógenos, os quais, aumentando na corrente circulatória, consequentemente
diminuem a secreção de FSH. De modo semelhante, quando os hormônios hipofisário alcançam um determinado nível, alguns núcleos hipotalâmicos respondem
diminuindo a produção de seus hormônios liberadores específicos, há um declínio
na secreção do hormônio trófico hipofisário e, finalmente, uma diminuição do
nível funcional da glândula-alvo.
52
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
2.2.2.1.2 Retroalimentação estimulatória ou positiva (Feedback
positivo)
Nesse sistema, a elevação de determinado hormônio provoca o aumento de
outro hormônio. Por exemplo, o aumento dos níveis de estrógeno durante a
fase pré-ovulatória acarreta um pico do LH hipofisário. Esses dois eventos são
precisamente sincronizados, uma vez que o pico de LH é necessário para a ruptura
do folículo ovariano.
2.2.2.1.3 Retroalimentação de alça longa e curta
Tanto os hormônios hipofisário quanto os esteroides regulam a síntese, o armazenamento e a liberação dos hormônios hipotalâmicos por meio de dois
mecanismos de retroalimentação: alça longa e alça curta. O mecanismo de retroalimentação de alça longa envolve a interação entre a gônada, a hipófise e o
hipotálamo. Enquanto que o de alça curta permite às gonadotrofinas hipofisárias
influenciar a atividade secretória dos hormônios liberadores hipotalâmicos sem
a mediação das gônadas.
2.2.2.1.4 Reflexo neuroendócrino
Além dos mecanismos de retroalimentação acima citados, o sistema nervoso
também pode controlar a liberação de hormônios por meio de vias nervosas,
como, por exemplo, a descida do leite pela ocitocina.
2.2.2.1.5 Controle imunoendócrino
O portal do agroconhecimento
53
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
Os sistemas endócrino e imune interagem extensivamente regulando-se entre si.
Diversos órgãos endócrinos estão envolvidos em alguns aspectos desse processo
regulatório: hipotálamo, hipófise, gônadas, adrenais, pineal, tireoide e timo. Muitos desses órgãos são afetados pela função imune.
2.3 Glândulas endócrinas
2.3.1 Hipotálamo
O hipotálamo é uma porção especializada do sistema nervoso central (SNC) que
se encontra situada na base do cérebro (região do terceiro ventrículo), acima e
atrás do quiasma óptico, estendendo-se do quiasma óptico aos corpos mamilares
(Figura 19).
Figura 19: Desenho esquemático dos núcleos hipotalâmicos e da hipófise. AH,
adeno-hipófise; ARQ, núcleo arqueado; AHA, área hipotalâmica anterior; AHD, área
hipotalâmica dorsal; NMD, núcleo medial dorsal; EM, eminência média; NH, neurohipófise; CM, corpo mamilar; PM, núcleo pré-mamilar; QO, quiasma óptico; NPV,
núcleo paraventricular; NPO, núcleo pré-óptico; AHP, área hipotalâmica posterior; PT,
pars tuberalis; NSQ, núcleo supraquiasmático; NOS, núcleo supra-óptico; NVM, núcleo
ventro-medial. FONTE: HAFEZ e HAFEZ (2004).
54
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
Os elementos celulares hipotalâmicos que regulam a secreção da hipófise anterior não estão localizados em uma região específica. No entanto, os núcleos
nervosos mais importantes do hipotálamo foram identificados como o supraóptico e o paraventricular.
Existem conexões neurais entre o hipotálamo e a hipófise posterior através pelo
eixo hipotalâmico-hipofisário e conexões vasculares do hipotálamo com a hipófise anterior (Figura 18). O sangue arterial ganha a hipófise através da artéria
hipofisária superior e inferior. A artéria hipofisária superior forma alças capilares
no nível da eminência média e hipófise posterior. Destes capilares, o sangue
flui para o sistema porta-hipotalâmico-hipofisário, o qual começa e termina em
capilares sem a passagem pelo coração. Parte do fluxo venoso que sai da hipófise
anterior se dá através de um fluxo retrógrado, que expõe o hipotálamo a altas
concentrações dos hormônios da hipófise anterior. Tal fluxo sanguíneo possibilita
ao mecanismo hipofisário a retroalimentação negativa de regulação das funções
hipotalâmicas. Esse tipo de retroalimentação é conhecido como retroalimentação
de alça curta.
Os hormônios hipotalâmicos relacionados com a reprodução incluem o Hormônio
Liberador de Gonadotrofinas (GnRH), os Fatores Liberador (PRF) e Inibidor (PIF) da
Prolactina, o Hormônio Liberador de Tireotropina (TRH) e o Hormônio Liberador
de Corticotropina (CRH). O hipotálamo ainda é a principal fonte de ocitocina e
vasopressina, que são estocadas na hipófise posterior.
O portal do agroconhecimento
55
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
2.3.1.1 Hormônio Liberador de Gonadotrofinas (GnRH)
O GnRH é um hormônio decapeptídeo (p-Glu-His-Trp-Ser-Tyr-Gly-Leu-Arg-ProGly-NH2) com peso molecular de 1.183 dáltons, sintetizado e armazenado no
hipotálamo basal médio. O GnRH fornece uma ligação humoral entre os sistemas
nervoso e endócrino. Em resposta à estimulação nervosa, pulsos de GnRH são
liberados no sistema porta-hipotalâmico-hipofisário promovendo a liberação do
Hormônio Luteinizante (LH) e Hormônio Folículo-Estimulante (FSH) da hipófise
anterior.
A secreção dos hormônios liberadores hipotalâmicos é modulada pelos níveis
dos hormônios secretados nos órgãos-alvo primários e secundários. No caso do
GnRH, o controle da secreção na fêmea é feito pelas próprias gonadotrofinas
hipofisiárias (LH e FSH), progesterona e o estradiol.
Existem agonistas sintéticos do GnRH que são utilizados com fins terapêuticos na
prática veterinária. Um deles, a buserelina, é 17 vezes mais potente que o GnRH
natural devido a sua menor taxa de degradação e, portanto, maior meia-vida.
2.3.1.2 Fator Liberador Prolactina (PRF), Fator Inibidor de Prolactina
(PIF) e Hormônio Liberador de Tireotropina (TRH)
Os fatores PRF e PIF controlam a biossíntese e a secreção da prolactina. O efeito
inibitório parece prevalecer durante o estado basal através do PIF. A secreção
de prolactina também é estimulada por neurotensina, substância P, histamina,
serotonina e agentes α-adrenérgicos. O Hormônio Liberador de Tireotropina (TRH)
tem sido pesquisado como o PRF devido a que estimula a secreção de prolactina.
56
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
O TRH é um tripeptídeo, sendo o menor hormônio peptídico que se conhece e
tendo a seguinte sequência: p-Glu-His-Pro-NH2. O TRH estimula a liberação de
tireotropina (TSH) e prolactina (PRL) na hipófise e sua secreção é controlada pelos
hormônios tireoidianos (T3 e T4) e pelo TSH. Seu mecanismo de ação é através
do cAMP.
2.3.2 Hipófise (Glândula Pituitária)
A hipófise está localizada na sela túrcica, uma depressão óssea localizada na base
do cérebro, logo, diretamente abaixo do hipotálamo.
A hipófise ou pituitária é uma estrutura altamente complexa formada por grupos
celulares que sintetizam diferentes tipos de hormônios. É dividida em três porções: hipófise anterior; hipófise posterior; e lóbulo intermediário.
2.3.2.1 Adeno-hipófise ou hipófise anterior
Os tipos celulares da hipófise anterior têm sido tradicionalmente classificados de
acordo com suas características de coloração, em cromófilos acidófilos e basófilos.
A hipófise anterior tem cinco tipos diferentes de células, que secretam seis hormônios: os gonadótrofos, que secretam o Hormônio Folículo-Estimulante (FSH)
e o Hormônio Luteinizante (LH); os mamótrofos, responsáveis pela secreção de
prolactina; os corticótrofos, que secretam o Hormônio adenocorticotrófico (ACTH);
os tireótrofos, que secretam o Hormônio Tireotrópico (TSH); e os somatótrofos,
que secretam o Hormônio do Crescimento (GH).
O portal do agroconhecimento
57
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
2.3.2.1.1 Hormônio Folículo-Estimulante (FSH) e o Hormônio
Luteinizante (LH)
Cada subunidade proteica das gonadotropinas possui duas cadeias de oligossacarídeos unidos por ligações N-glicosídicas, sendo as unidades monossacarídicas
mais comuns manose, glicosamina, fucose e ácido siálico. Este último é o responsável pela meia-vida do hormônio devido ao fato de que antes da degradação do
hormônio, deve ocorrer a remoção dos resíduos de ácido siálico. Assim, quanto
maior é a proporção de ácido siálico na molécula, maior é a meia-vida do hormônio (Tabela 3).
Tabela 3: Conteúdo de carboidratos e meia-vida das gonadotropinas.
Hormônio
Peso
molecular
(Dáltons)
Glicódios
(%)
Ácido siálico (%)
Meia-vida
(horas)
LH
28.500
16
1-2
0,5
FSH
34.000
30
5
2
hCG
36.700
32
8,5
11
eCG
68.000
48
10,4
26
A secreção das gonadotropinas hipofisiárias está sob controle do GnRH hipotalâmico, obedecendo a uma modulação feedback negativa por parte dos esteroides
gonadais (estrógeno e progesterona na fêmea, testosterona no macho). A secreção basal das gonadotropinas é pulsátil sendo interrompida por um pico massivo
de LH durante o estro, no caso dos mamíferos que têm ovulação espontânea.
Esse pico de LH é disparado por um pico de GnRH hipotalâmico, o qual, por sua
vez, é causado por um aumento na liberação de 17β-estradiol durante o proestro
(feedback positivo).
58
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
Os agentes opioides endógenos causam diminuição, tanto da frequência quanto
da altura, dos picos de secreção de LH. Este fato pode ter importância quando
se relaciona o estresse, e a consequente secreção de opioides endógenos com
inibição da função reprodutiva.
O FSH na fêmea é responsável pelo crescimento e a maturação dos folículos
ovarianos ou folículos de De Graaf. O FSH por si só não causa a secreção de
esteroides (estrógenos) no ovário; ao contrário, o FSH necessita da presença do
LH para estimular a produção estrogênica.
O LH tem como função de maturação oocitária, induzir a ovulação e manter o
corpo lúteo, além de estimular, junto com o FSH, a secreção de esteroides no
ovário (estrógenos antes da ovulação e progesterona no corpo lúteo).
2.3.2.1.2 Prolactina
A prolactina (PRL) ou hormônio lactogênico é sintetizado nas células mamotrópicas da adeno-hipófise. É o maior hormônio peptídico que existe (199 aminoácidos, peso molecular 23,3 kDa) considerando uma cadeia só. Existe grande
variabilidade das PRLs entre as distintas espécies. A meia-vida da PRL é de 15
minutos.
A secreção de prolactina é pulsátil sendo controlada inibitoriamente por ação
da dopamina (PIF) e estimulada pelas endorfinas, pois estas inibem a secreção
de dopamina. A secreção de PRL também está favorecida por PRF, TRH, estrógenos, progesterona e por estímulos neurogênicos como a sucção das tetas pelo
lactente, a ordenha ou por sensações de calor, dor e estresse. Os estrógenos,
especialmente o 17β-estradiol, estimulam a secreção de prolactina por inibir o
fator inibidor de prolactina (dopamina) e, também, aumentam a secreção de
O portal do agroconhecimento
59
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
PRL por modular os receptores de TRH, hormônio que estimula a secreção de PRL
na hipófise. A secreção de PRL pode ser inibida por derivados do ergot como a
bromocriptina, um agonista da dopamina. A PRL pode também regular sua própria secreção atuando diretamente sobre o hipotálamo (feedback de alça curta
sobre o TRH). A dopamina, que constitui o único fator hipotalâmico não-peptídico,
parece atuar impedindo a mobilização de Ca+2 ao interior da célula lactotrópica
secretora da prolactina na adeno-hipófise. A amamentação parece inibir a secreção da dopamina e, portanto, aumenta os níveis de prolactina. A prolactina, por
sua vez, provoca a liberação de dopamina da eminência média, conformando
uma regulação feedback negativa. A PRL é secretada com flutuações durante os
diferentes estágios do ciclo reprodutivo e sua concentração se difere entre as
espécies domésticas (Tabela 4). Aumentos de PRL ocorrem durante a ovulação e
também durante a fase luteal do ciclo ovariano na vaca.
Tabela 4: Níveis sanguíneos de prolactina em várias espécies.
Espécie
Valor (ng/mL)
Cadela (anestro)
9,1 ± 1,2
Cadela (2ª semana de lactação)
86 ± 19
Cadela (pós-parto)
117 ± 24
Cadela (ovarioectomizada)
7,9 – 11,5
Gata (início de gestação)
7,0 ± 0,3
Gata (fim de gestação)
43,5 ± 4,5
Vaca (fase luteal)
23,3 ± 4,8
Vaca (fase folicular)
15,8 ± 2,7
Porca (2ª semana de lactação)
9,1 – 26,1
Porca (pós-desmame)
1,4 – 1,9
60
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
Também ocorre aumento de PRL durante a lactação e no parto. A prolactina
faz parte do complexo mamotrófico que promove o crescimento da glândula
mamária, junto com o hormônio do crescimento, estradiol, progesterona, glicocorticoides e hormônios tireoidianos.
A PRL tem efeito luteotrópico na ovelha e roedores, mas não na vaca. Em vários
animais, a PRL parece ter um efeito inibitório sobre a secreção das gonadotropinas hipofisiárias, pelo qual tem sido sugerido que a PRL seria um hormônio
anti-gonadotrópico, uma vez que estimula a biossíntese de dopamina, a qual tem
efeito negativo sobre a secreção de GnRH.
A PRL tem sido responsabilizada pela ação inibitória da amamentação sobre o
início da atividade ovárica durante o pós-parto em vacas de corte. Por outro lado,
tem sido utilizada bromocriptina, agente agonista da dopamina, com a ideia de
desbloquear o suposto efeito da PRL sobre a ciclicidade ovárica em vacas. Foi
encontrado que a bromocriptina causa diminuição dos níveis de PRL, porém sem
ocorrer redução do intervalo do parto ao primeiro cio pós-parto e nem aumento
nos níveis de LH. Os indícios levam a aceitar que é o efeito do estímulo neural
do amamentamento como tal e não a maior quantidade de PRL secretada, o
responsável pela supressão da secreção de gonadotropinas durante o pós-parto
de vacas de corte.
O portal do agroconhecimento
61
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
2.3.2.1.3 ACTH ou corticotropina
O mais importante glicocorticoide envolvido na reprodução dos animais domésticos é o cortisol. A concentração sérica média de cortisol em bovinos oscila entre
2 e 12 mg/ml.
A secreção de glicocorticoides é controlada pelo hipotálamo. O estressor atua,
via sistema nervoso central, sobre as células neuro-secretoras do hipotálamo, as
quais reagem com uma maior secreção do Hormônio Liberador de Corticotropina
(CRH). Através de um sistema portal vascular, o CRH é transportado do hipotálamo até os lóbulos anteriores da hipófise onde provoca uma elevada secreção
do hormônio adenocorticotrófico (ACTH) ou corticotrofina. Circulando na corrente
sanguínea, este estimula o córtex da adrenal a produzir glicocorticoides (cortisol).
Cinco minutos após a injeção com ACTH, soros sanguíneos de bovinos mostram
elevada concentração de glicocorticoides.
A concentração de cortisol no plasma é regulada por um rígido mecanismo de
feedback negativo. Em condições normais, a concentração sanguínea de cortisol
atua, principalmente, sobre o hipotálamo, inibindo a secreção de CRH, reduzindo produção de ACTH e, consequentemente, reduzindo a secreção dos próprios
hormônios do córtex adrenal.
Em condições de intenso e prolongado estresse (calórico, nutricional, ambiental
etc.), os estressores estimulam diretamente a liberação de CRH, com secreção
posterior de ACTH e cortisol, contornando o status “regulador” do hipotálamo e
desprezando o feedback negativo. Com a ativação do eixo hipotálamo-hipófiseadrenal, ocorrem mudanças em quase todo sistema endócrino. Por exemplo, um
organismo sob estresse, a hipófise secreta menos hormônio do crescimento, além
de menos TSH e GnRH, conduzindo, os dois últimos, a uma reduzida atividade da
tireoide e das gônadas. Dado o efeito catabólico e a gliconeogênese estimulados
62
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
pelos glicocorticoides, pode ocorrer constante degradação de tecidos musculares
e gordurosos, assim como a inibição da síntese desses tecidos, resultando com
isso a perda de peso e reduzido crescimento. Não menos importante é o efeito
dos glicocorticoides causando a atrofia do sistema timo-linfático, diminuindo a
concentração dos linfócitos e anticorpos, resultando na queda da resposta imunológica do organismo no combate a quaisquer infecções.
2.3.2.2 Neuro-hipófise ou hipófise posterior
Os hormônios da hipófise posterior diferem dos outros hormônios hipofisário
pelo fato de que não se originam da hipófise, mas são apenas armazenados ali
até quando forem necessários, pois são produzidos no hipotálamo (Figura 20).
A neuro-hipófise possui terminações axônicas de neurônios hipotalâmicos que
armazenam dois hormônios: a arginina-vasopressina (AVP) ou hormônio antidiurético (ADH) e a ocitocina (hormônio da descida do leite). Esses hormônios
são transferidos do hipotálamo para a hipófise posterior não através do sistema
porta vascular, mas ao longo dos axônios do sistema nervoso. Do ponto de vista
reprodutivo o hormônio neuro-hipofisiário de interesse é a ocitocina.
Os neurônios produtores da ocitocina no hipotálamo têm em abundância retículos
endoplasmáticos rugosos e aparelhos de Golgi, além de grânulos secretores,
os quais se localizam no corpo do neurônio e nos axônios que se estendem à
neuro-hipófise. Os neurônios secretores de ocitocina se encontram em núcleos
específicos do hipotálamo: o núcleo supraóptico e o núcleo paraventricular. Os
hormônios, dentro dos grânulos, estão unidos a uma proteína transportadora chamada neurofisina e são secretados à circulação desta forma. Existe uma grande
similaridade entre as neurofisinas de bovinos, suínos e equinos.
O portal do agroconhecimento
63
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
Figura 20: Conexão nervosa entre o hipotálamo e hipófise posterior. PVN, núcleo
paraventricular; SON, núcleo supra-óptico. Fonte: CUNNINGHAM (1999).
2.3.2.2.1 Ocitocina
A secreção da ocitocina é estimulada via neurogênica por amamentação, ordenha,
parto, dilatação cervical ou vaginal ou estímulo clitoridiano, sendo a acetilcolina
o modulador estimulante e a adrenalina e a noradrenalina os agentes inibidores.
Os níveis de ocitocina têm variações entre as espécies (Tabela 5) e durante
o ciclo estral na vaca, ovelha e cabra. A concentração sanguínea de ocitocina
aumenta depois do pico pré-ovulatório de LH e diminui depois da regressão do
corpo lúteo. Os estrógenos ovarianos estimulam a liberação de ocitocina pituitária
enquanto que a progesterona a inibe. Têm sido encontrados outros fatores nãoreprodutivos, como o estresse, que afetam a liberação de ocitocina.
64
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
Tabela 5: Concentração plasmática de ocitocina em alguns animais.
Espécie
Valor (pmol/L)
Cadela (lactação)
15 – 66
Vaca (pré-odenha)
1,6 ± 0,6
Cabra (basal)
4,5 ± 1,0
A ocitocina desempenha um importante papel na fase folicular do ciclo estral,
estimulando a contração uterina para facilitar o transporte dos espermatozoides
para o oviduto durante o estro (cio).
A dilatação da cérvix no momento do parto, causada pela passagem do feto,
estimula a liberação reflexa de ocitocina (Reflexo de Ferguson; Figura 21) que
promove a contração do miométrio, facilitando a expulsão do feto e da placenta.
Figura 21: Esquema do reflexo de Ferguson ou de expulsão do feto. Através do nervo pudendo, realizada pela presença das bolsas fetais e/ou feto no canal vaginal,
estabelece-se um estímulo na medula espinhal que resulta na ocorrência dos esforços por meio das contrações abdominais. FONTE: PRESTES e LANDIM-ALVARENGA
(2006).
O portal do agroconhecimento
65
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
A ocitocina também causa a contração das células mioepiteliais (células musculares lisas) que circundam os alvéolos da glândula mamária durante a lactação,
resultando na descida do leite. Na fêmea lactente, estímulos táteis, visuais, olfativos e auditivos associados à sucção ou ordenha induzem a liberação de ocitocina
na corrente sanguínea.
O estrógeno é necessário para a ação da ocitocina, pois estimula a síntese de
receptores para ocitocina; portanto os estrógenos aumentam a resposta do útero
à ocitocina. A progesterona, por sua parte, inibe a secreção de ocitocina, o que
explica que durante a gestação a resposta do útero à ocitocina está muito reduzida. A adrenalina, secretada no estresse, diminui a descida do leite da glândula
mamária por bloquear a ação da ocitocina mediante a inibição de sua secreção
na neuro-hipófise e também, possivelmente, por bloquear os receptores da ocitocina nas células mioepiteliais.
O corpo lúteo também secreta a ocitocina, estando envolvida no processo da luteólise (regressão do corpo lúteo) na maioria dos mamíferos. A ocitocina ovariana,
secretada sem a neurofisina, tem receptores no endométrio e sua ação estimula
a biossíntese de prostaglandina-F2α (PGF2α). A síntese dos receptores de ocitocina
no endométrio é estimulada por 17β-estradiol. Portanto, a ocitocina tem dois
sítios de origem: um hipotalâmico e outro ovariano.
2.3.3 Gônadas (ovários)
Os ovários desempenham um papel duplo: a produção de células germinativas
(gametogênese) e a secreção de hormônios gonadais (esteroidogênese). As unidades estruturais e funcionais do ovário que garantem essas funções são: folículo
ovariano (gametogênese e esteroidogênese) e o corpo lúteo (esteroidogênese).
66
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
As células intersticiais ovarianas ou células da teca interna do folículo ovariano
são as fontes primárias dos estrógenos circulantes. Após a ruptura do folículo
(ovulação), as células da granulosa e da teca são substituídas pelo Corpo Lúteo
(CL), que secreta a progesterona. O estrógeno, progesterona e testosterona são
hormônios esteroides derivados do colesterol (Figura 22). Na cascata da reação,
o colesterol é uma molécula esteroide de 27 átomos de carbono que se converte em pregnolona (20 átomos de carbono). A pregnolona é convertida em
progesterona, que por sua vez, converte-se em andrógenos e estrógenos. Entre
os sexos, a fêmea apresenta uma maior quantidade de enzimas específicas nesta
cascata, o que lhe favorece maior facilidade em converter estrógenos a partir
dos andógenos. Animais com baixa ou nenhuma reserva de gordura (colesterol)
apresentam maior dificuldade em produzir esteroides reprodutivos, resultando
em falhas reprodutivas na fêmea.
Os folículos contêm em seu interior as células germinativas ou ovócitos ou oócitos. Com cada ciclo estral, vários folículos crescem e se desenvolvem, sendo que
somente um folículo, nas espécies uníparas, ou alguns deles, nas espécies multíparas, serão ovulados. Os folículos são responsáveis pela produção de estrógenos
e o corpo lúteo pela produção de progesterona. O ovário também é responsável
pela produção de hormônios peptídicos, tais como, a inibina, o fator de inibição
de maturação do ovócito, a relaxina e a gonadocrinina.
O portal do agroconhecimento
67
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
Figura 22: Vias metabólicas envolvidas na produção dos principais hormônios esteroides. Fonte: CUNNINGHAM (1999).
Os dois ovários não funcionam de igual maneira. Em ovelhas e cabras, 55 a 60 %
das ovulações ocorrem no ovário direito, enquanto que na vaca este valor chega
a 60-65 %. Na égua e na porca, o ovário esquerdo é mais ativo, produzindo 55
a 60 % dos ovócitos.
68
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
Diferentemente dos testículos, os ovários permanecem na cavidade abdominal.
A forma do ovário varia em função da espécie e da fase do ciclo estral.
O desenvolvimento dos folículos pode ser avaliado em função do seu tamanho,
o número de camadas de células granulosas, o desenvolvimento das células da
teca, a posição do ovócito dentro do cumulus oophorus e o tamanho do antro
folicular (Figura 23). Em função desses fatores, os folículos podem ser classificados em:
»» Folículos primários ou primordiais: são ovócitos envolvidos por
uma única camada de células foliculares planas ou cuboides, rodeadas de tecido intersticial;
»» Folículos secundários: nos quais o número de células foliculares
aumenta e passam a ser chamadas de células granulosas, com
aparência irregular poliédrica e com pouco líquido folicular nos
espaços intercelulares;
»» Folículos terciários: quando as células granulosas começam a secretar uma grande quantidade de líquido folicular que se acumula
nos espaços intercelulares, formando um antro. Nesse ponto, os
capilares do estroma invadem a camada fibrosa de células que
rodeia o folículo e formam uma camada vascular chamada de teca
interna. Contudo, as células granulosas dentro do folículo estão
privadas de suprimento sanguíneo pela membrana basal. A zona
pelúcida que contorna o ovócito está rodeada por uma massa sólida de células granulosas radiais que formam a corona radiata;
»» Folículo de De Graaf ou folículo maduro: no qual as células da
granulosa aumentam de tamanho, o antro aumenta em função da
O portal do agroconhecimento
69
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
maior secreção de líquido folicular e o ovócito é pressionado pelo
líquido para um extremo do folículo. O ovócito agora é rodeado
por inúmeras células foliculares e forma o cumulus oophorus. As
células deste folículo se diferenciam em células granulosas, sob a
membrana basal, e em células das tecas interna e externa. O ovócito apresenta duas membranas distintas, a membrana vitelina ou
membrana plasmática, e a zona pelúcida, membrana homogênea
composta de proteínas suscetíveis de serem dissolvidas por enzimas proteolíticas.
Figura 23: Classificação fisiológica e morfológica dos folículos ovarianos. P, folículos
primordiais; PM, folículos primários; S, folículos secundários; T, folículos terciários.
FONTE: HAFEZ e HAFEZ (2004).
2.3.3.1 Estrógeno
O estradiol é o estrógeno primário, sendo que a estrona e o estriol representam
outros estrógenos metabolicamente ativos.
70
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
O 17β-estradiol é o estrógeno biologicamente ativo produzido pelos ovários, com
quantidades menores de estrona. Todos os estrógenos são produzidos a partir
dos andrógenos.
As células da teca interna são responsáveis pela produção de andrógenos que são
transportados para as células da granulosa, no interior dos folículos ovarianos. A
partir destes andrógenos, as células da granulosa secretam enzimas específicas
para a conversão em estrógenos e liberados no fluido folicular e na corrente
sanguínea.
Os estrógenos são carregados pela corrente sanguínea por meio de proteínas
ligadoras. As funções dos estrógenos são:
»» Atuam no SNC induzindo o comportamento de estro (cio) na vaca;
»» Atuam no útero aumentando tanto a amplitude quanto a frequência das contrações, potencializando os efeitos da ocitocina e PGF2α;
»» Desenvolvimento físico das características sexuais secundárias femininas;
»» Estimulam o crescimento dos ductos e desenvolvem as glândulas
mamárias;
»» Exercem efeitos de feedback tanto negativos quanto positivos no
controle da liberação de LH e FSH no hipotálamo. O efeito negativo ocorre no centro tônico do hipotálamo e o efeito positivo, no
centro de pré-ovulatório.
O portal do agroconhecimento
71
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
Em ruminantes, os estrógenos também possuem um efeito anabólico proteico,
aumentando o ganho de peso e o crescimento. O mecanismo pode estar relacionado à habilidade dos estrógenos em estimular a hipófise para a liberação de
maiores quantidade de hormônio do crescimento.
Dentre os estrógenos sintéticos disponíveis no mercado, destacam-se o benzoato
de estradiol e o cipionato de estradiol. A explanação destes hormônios será
abordada nos capítulos seguintes.
2.3.3.2 Progesterona
A progesterona é o progestágeno natural de maior prevalência, sendo secretada
pelas células luteínicas do corpo lúteo, pela placenta e pelas glândulas adrenais. A progesterona, assim com os estrógenos e andrógenos, é transportada
na corrente sanguínea por uma proteína de ligação. Sua secreção é estimulada
primariamente pelo LH.
A progesterona desempenha as seguintes funções:
»» Prepara o endométrio para a implantação embrionária e a manutenção da prenhez, aumentando a atividade secretora das glândulas endometriais e inibindo a motilidade do endométrio;
»» Atua sinergicamente com os estrógenos na indução do comportamento do estro;
»» Auxilia no desenvolvimento do tecido secretor (alvéolos) da glândula mamária;
72
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
»» Provoca a inibição do estro e do pico pré-ovulatório do LH quando
em níveis elevados. Portanto, a progesterona desempenha papel
fundamental na regulação hormonal do ciclo estral;
»» Inibe a motilidade uterina.
Alguns progestágenos sintéticos estão disponíveis para a sincronização do ciclo
estral de ruminantes. Tais hormônios podem ser administrados por via oral, inseridos no interior da vagina por meio de dispositivos intravaginais ou por implantes
auriculares (subcutâneos) pelo período correspondente à fase luteínica do ciclo
estral. A explanação destes hormônios será abordada nos capítulos seguintes.
2.3.3.3 Relaxina
A relaxina é um hormônio polipeptídico com peso molecular de 5.700 dáltons. A
relaxina é secretada primariamente pelo corpo lúteo durante a gestação. A principal ação biológica da relaxina é a dilatação da cérvix e da vagina durante o parto.
Ela inibe também as contrações uterinas e causa o crescimento das glândulas
mamárias quando administradas em conjunto com o estradiol.
2.3.3.4 Inibina, ativina e folistatina
A inibina e a ativina foram isoladas de fluidos gonadais por causa de seus efeitos na produção de FSH. Elas são reguladoras parácrinas, pois modulam o sinal
endócrino do LH.
As gônadas são as principais fontes de inibina e proteínas correlatas, que contribuem para a regulação endócrina do sistema reprodutivo. Na fêmea, as células
O portal do agroconhecimento
73
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
da granulosa são as produtoras de inibina. As inibinas não são esteroides, mas
proteínas compreendendo duas subunidades alfa e beta interligadas por pontes
dissulfídicas, liberadas pela corrente sanguínea.
As inibinas desempenham um papel importante na regulação hormonal da foliculogênese ovariana durante o ciclo estral. Elas atuam como sinalizadores químicos
para a hipófise sobre o número de folículos em crescimento no ovário. A inibina
reduz a secreção de FSH até certo nível, o que mantém o número de ovulações
espécie-específico tanto nas espécies mono quanto poliovulatórias. Pela inibição
da liberação de FSH sem alteração da liberação de LH, as inibinas podem ser
parcialmente responsáveis pela liberação diferenciada de LH e FSH pela hipófise.
O fluido folicular ovariano contém uma fração que estimula ao invés de inibir a
secreção de FSH. As proteínas responsáveis por essa atividade são caracterizadas
como ativinas. As ativinas são potentes liberadores de FSH, tendo ação contrária
à inibina.
A folistatina é outra proteína isolada do fluido folicular ovariano. Ela não somente
inibe a secreção de FSH como a inibina, mas também se liga à ativina, neutralizando sua atividade biológica. Portanto, constitui-se num agente modulatório
da secreção de FSH.
2.3.3.5 Prostaglandinas
As prostaglandinas, primeiramente foram isoladas de fluidos das glândulas sexuais acessórias do macho, receberam essa nomenclatura por causa de sua associação com a glândula prostática, sendo que quase todos os tecidos orgânicos
podem secretá-las. O ácido aracdônico, um ácido graxo essencial, é o precursor
da maioria das prostaglandinas mais intimamente associadas com a reprodu-
74
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
ção, principalmente a prostaglandina F2α (PGF2α) e a prostaglandina E2 (PGE2)
(Figura 24). Na reprodução, as prostaglandinas estão envolvidas na liberação de
gonadotrofinas, ovulação, regressão do corpo lúteo, motilidade uterina, parto e
transporte de espermatozoides.
A maioria das prostaglandinas atua localmente no seu sítio de produção numa
interação célula a célula e, portanto, não se encaixa perfeitamente na definição
clássica dos hormônios. Mas, também podem ser liberadas na corrente sanguínea para atuar em tecidos-alvo distantes de seu sítio de produção.
A PGF2α é o agente luteolítico natural associado ao final da fase luteínica (corpo
lúteo) do ciclo estral, permitindo o início de um novo ciclo quando da ausência
de fertilização. Uma elevação nos níveis de estrógenos, os quais promovem o
crescimento do miométrio, estimula a síntese e a liberação de PGF2α. Ela é particularmente potente na interrupção da gestação em sua fase inicial. Em fêmeas
prenhes, o embrião em desenvolvimento envia um sinal ao útero (reconhecimento materno da prenhez), prevenindo os efeitos luteolíticos da PGF2α.
Como nos casos de inflamação, na involução uterina pós-parto, as prostaglandinas exercem importante função (BENCHARIF et al., 2000). Além de acelerar o
processo de involução, a PGF2α estimula a contração da camada muscular uterina
(miométrio) após o parto. Esta substância é normalmente produzida pelo útero,
sendo responsável pela correta involução uterina pós-parto, em tempo normal.
A capacidade da PGF2α em induzir a luteólise é explorada para a manipulação do
ciclo estral (sincronização do estro) e também para a indução do parto.
O portal do agroconhecimento
75
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
2.3.3.6 Hormônios placentários
A placenta secreta diversos hormônios idênticos ou com atividades biológicas
similares aos hormônios reprodutivos dos mamíferos. Dentre eles, destacam-se:
Gonadotrofina Coriônica Equina (eCG) e a Gonadotrofina Coriônica Humana (hCG).
O hormônio eCG (originalmente denominado PMSG) foi descoberto quando se
verificou que o sangue de éguas prenhes produzia maturidade sexual quando
administrado em ratos imaturos. A eCG é uma glicoproteína com subunidades
alfa e beta similares ao LH e ao FSH, porém com conteúdo de carboidrato mais
elevado (principalmente o ácido siálico). O alto conteúdo de ácido siálico parece
contribuir para a longa meia-vida de diversos dias da eCG. Portanto, uma única
administração de eCG possui efeitos biológicos por mais de uma semana. Este
hormônio é comumente utilizado em programas de IATF, com a finalidade de
prolongar a ação folículo estimulante no folículo dominante.
O útero equino secreta gonadotrofina placentária, sendo os cálices endometriais a
fonte específica da eCG. Tais cálices, que são formados ao redor de 40 dias, permanecem até 85 dias de gestação. A eCG possui atividade biológica semelhante
ao FSH e ao LH, porém, predominantemente ao FSH. A eCG circula na corrente
sanguínea das éguas prenhes e não é excretada na urina. A secreção de eCG estimula o desenvolvimento dos folículos ovarianos. Alguns folículos ovulam, mas a
maioria transforma-se em folículos luteinizados devido à ação da eCG semelhante ao LH. Tais corpos lúteos acessórios produzem progesterona, o que mantém a
prenhez da égua. A eCG foi uma das primeiras gonadotrofinas comercialmente
disponíveis usadas para indução da superovulação em animais domésticos.
A glicoproteína hCG consiste de uma subunidade alfa e outra beta com peso molecular de 40.000 dáltons. A subunidade alfa do hCG é similar à subunidade alfa
do LH em humanos, suínos, ovinos e bovinos. A hCG é primariamente luteinizante
76
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
e luteotrófico, possuindo baixa atividade de FSH. As células do sinciciotrofoblasto
na placenta dos primatas são responsáveis pela síntese de hCG, que pode ser
encontrada tanto no sangue quanto na urina. Sua presença na urina no início da
gestação constitui a base de vários testes laboratoriais para a gravidez humana.
Ela pode ser detectada na urina por meio de imunoensaios sensíveis a partir de
8 dias após a concepção.
2.4. Regulação hormonal da reprodução na fêmea
Na maior parte da vida reprodutiva de uma fêmea fértil, ela não se apresenta em
atividade cíclica regular (ou seja, apresenta-se em anestro). Quando somados, os
períodos de inatividade durante a pré-puberdade, gestação e lactação são muito
maiores que os períodos relativamente curtos de atividade cíclica. Entretanto, os
períodos em que é possível interferir no processo reprodutivo (cruzar/não cruzar;
escolha do macho/sêmen; controle do estro; indução da ovulação etc.) são os
mais importantes e é nesta fase que a maior parte dos problemas reprodutivos
pode acontecer.
Os princípios do mecanismo hormonal da reprodução são basicamente os mesmos para todos os mamíferos domésticos, embora haja algumas diferenças entre
eles. Alguns animais são poliéstricos contínuos como os bovinos e os suínos,
ciclando durante todo o ano, enquanto outros são poliéstricos estacionais, como
os equinos, ovinos e felinos.
Além disso, há diferenças no mecanismo de ovulação. A maioria dos animais
ovula espontaneamente, mas na gata, coelha e camela a ovulação é induzida
pela estimulação de receptores sensoriais na vagina e na cérvix durante o coito.
O portal do agroconhecimento
77
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
O processo reprodutivo dos mamíferos é regulado por uma complexa cascata de
atividades combinadas do sistema nervoso central, tecidos secretórios, tecidos
alvo e vários hormônios. A Figura 24 é uma representação esquemática dos
órgãos e hormônios mais importantes envolvidos na reprodução da fêmea, com
algumas de suas funções e interações.
Figura 24: Inter-relações no controle da função reprodutiva da fêmea. FONTE: INTERVET (2006).
78
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
O sistema nervoso central recebe informações do ambiente em que o animal se
encontra (estímulo visual olfatório, auditivo e tátil) e envia a informação relevante
do ponto de vista reprodutivo para as gônadas via eixo hipotalâmico-hipofisáriogonadal. O hipotálamo e a hipófise estão firmemente ligados à parte ventral do
cérebro. Não são apenas produtores de hormônios, mas também órgãos-alvo,
formando um sofisticado sistema homeostático de feedback, por meio do qual
regulam sua própria taxa de secreção.
A partir de um estímulo do SNC, os neurônios endócrinos no hipotálamo produzem o hormônio liberador de gonadotrofinas. O GnRH é transportado pelo
sistema porta hipotálamo-hipofisário ao lobo anterior da pituitária, seu órgãoalvo, estimulando as células da pituitária a secretar o FSH e LH. O GnRH, FSH e LH
não são secretados em níveis constantes, mas em uma série de pulsos. O FSH
estimula o desenvolvimento dos folículos ovarianos. Na teca interna do folículo,
o LH estimula a síntese de androstenediona a partir do colesterol.
A androstenediona é convertida em testosterona, que é aromatizada em
17β-estradiol sob a influência do FSH, nas células da granulosa do folículo. O
estradiol exerce um feedback positivo no hipotálamo e na pituitária, aumentando
a frequência dos pulsos de GnRH. Quando o estradiol ultrapassa certo nível, o
hipotálamo responde com um pico de GnRH que, por sua vez, induz um pico de
LH que inicia a ovulação. Assim, o FSH estimula o crescimento dos folículos ovarianos, enquanto o LH estimula sua maturação, produção de estradiol e ovulação.
O LH dá suporte à formação e à função inicial do corpo lúteo.
Um dos principais efeitos do estradiol é a indução dos sintomas de estro. O
estro pode ser descrito como os sinais comportamentais e físicos que indicam
aos outros animais que a fêmea está na fase fértil de seu ciclo, e vai permitir a
cobertura pelo macho.
O portal do agroconhecimento
79
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
As células da granulosa também produzem inibina. Nem todos os efeitos deste
hormônio são compreendidos, mas seu nome é derivado do feedback negativo
que provoca na liberação de FSH da glândula pituitária, controlando assim o
desenvolvimento dos folículos. Depois da ovulação, os restos do folículo são
remodelados, formando o corpo lúteo, sob a influência do LH. A cavidade folicular
é preenchida com vasos sanguíneos, e as células da granulosa aumentam de
tamanho. O corpo lúteo é um órgão que produz basicamente progesterona e
ocitocina.
A progesterona é essencial para o ciclo normal na vaca e, após a concepção,
é o principal hormônio responsável pela manutenção da prenhez. Ela provoca
redução da liberação dos pulsos de GnRH, e assim inibe novas ovulações. Além
disso, prepara o endométrio para a implantação do embrião em desenvolvimento, e inibe as contrações da parede uterina pelo folículo durante a ovulação não
é fertilizado, não são recebidos sinais de prenhez vindos do embrião. Por volta
do dia 16 pós-ovulação, o endométrio do útero não gestante irá liberar PGF2α.
A prostaglandina-F2α dá início à regressão do corpo lúteo, denominada luteólise. O mecanismo luteolítico da prostaglandina envolve redução do suprimento
sanguíneo para o corpo lúteo via vasoconstrição, bem como um efeito direto
nas células luteínicas propriamente ditas. A ocitocina produzida no corpo lúteo
também desempenha um papel importante na luteólise.
Como resultado da regressão do corpo lúteo, as concentrações de progesterona
diminuem, removendo o bloqueio sobre a liberação de GnRH pelo hipotálamo.
Isto provoca início de uma nova fase folicular, com desenvolvimento de um folículo pré-ovulatório. A fase que envolve crescimento folicular, estro e ovulação é
denominado de fase folicular do ciclo estral. A fase dominada pela progesterona,
a partir da ovulação até a luteólise, é chamada fase luteínica ou luteal (Figura 25
e 26; observar que as duas figuras se complementam).
80
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
Figura 25: Níveis hormonais durante o ciclo estral. Fonte: INTERVET (2006).
Figura 26: Dinâmica folicular em um ciclo estral de 3 ondas foliculares, mostrando o
as secreções de FSH, LH e estradiol. FONTE: INTERVET (2006).
O portal do agroconhecimento
81
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
2.5 Endocrinologia da puberdade
A puberdade na fêmea pode representar tanto o primeiro cio, quanto a primeira
ovulação, eventos que geralmente não chegam juntos. A rigor, a puberdade é
definida como a época em que a fêmea está em capacidade fertilizante, o que
compreende tanto o aspecto comportamental (cio) como de formação de gametas e de capacidade de manter e desenvolver o embrião e o feto até o parto. A
primeira ovulação costuma ser “silenciosa”, isto é, sem sintomas de cio. Somente
a segunda ou terceira ovulações são acompanhadas de sinais evidentes de estro.
O início da puberdade vem precedido por um pico de LH, seguido de uma pequena elevação da progesterona circulante. Um 2º pico de LH marca a apresentação
da 1ª ovulação, que é silenciosa. É só no 3º pico de LH que se caracteriza a
puberdade completa, ou seja, sintomas de cio junto com ovulação.
Durante o período da pré-puberdade ocorrem pulsos de LH com amplitude similar
às observadas na puberdade, porém com frequência bem menor (2-3 h). Durante
a puberdade ocorre uma pronunciada diminuição da sensibilidade de inibição
dos esteroides às gonadotropinas, rompendo o forte feedback negativo entre
estradiol e LH. Este é o fundamento da hipótese gonadostática para explicar o
início da puberdade, a qual sugere que o eixo hipotâlamo-hipofisário-ovariano
estaria já maduro desde antes da puberdade, porém sem funcionar de forma
adulta devido ao mencionado efeito feedback. A hipótese gonadostática está
sustentada em quatro considerações.
Na primeira, o mecanismo de secreção de LH em resposta à ação estimulatória exógena de estradiol é observado desde antes da puberdade. Esta resposta
aumenta com a idade, o que supõe que a falta de uma produção permanente
de estradiol é o que impede uma secreção de LH no período da pré-puberdade.
82
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
Na segunda consideração, o ovário tem a capacidade de produzir níveis foliculares de estradiol em resposta a pulsos horários de LH desde antes da puberdade, o
que é coerente com o fato de que o ovário possui folículos terciários, portanto está
relativamente maduro desde antes da puberdade. Isto foi provado em ovelhas
ao administrar eCG/hCG na pré-puberdade obtendo indução da ovulação com
formação de corpo lúteo. Portanto, a falta de produção de estradiol não seria
devido à imaturidade do ovário, mas à falta de estímulo gonadotrópico.
Na terceira consideração, existe o potencial de produzir picos de LH com alta
frequência desde antes da puberdade. Entretanto, a frequência é pouca, não por
insuficiente desenvolvimento pituitário, mas porque o sistema é muito sensível
à ação inibitória do estradiol.
Enquanto que na quarta consideração, a inibição da secreção de LH em resposta
ao estradiol diminui no período da puberdade permitindo a secreção da gonadotropina e, consequentemente, estimulando o ovário para funcionar de forma
adulta.
Há uma quinta consideração que vem se tornando cada vez mais concreta à
medida que novos estudos são realizados, que é a influência da progesterona
na cronologia da puberdade. Quando ocorre o primeiro pico de LH, ocorre a
luteinização de pequenos folículos em crescimento, aumentando a progesterona
circulante. Quando ocorre o segundo pico de LH, a concentração de progesterona
torna-se ainda maior na corrente circulatória, o que promove uma desensibilização do hipotálamo em secretar GnRH, e consequentemente, gonadotrofinas. Com
este aumento de progesterona, ocorre um aumento da armazenagem de GnRH
no hipotálamo. Assim, a secreção de FSH, LH e estrógenos torna-se maior, o que
acarreta os sinais de estro com ovulação (terceiro pico de LH), caracterizando o
início da puberdade. Tal fato se observa em novilhas impúberes submetidas à
indução da puberdade com implantes intravaginais impregnados com progeste-
O portal do agroconhecimento
83
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
rona. Geralmente, utilizam-se implantes de terceiro ou quarto uso, dependendo
do fabricante.
A inibição da secreção de LH por parte do estradiol antes da puberdade pode
estar mediada por neurotransmissores como dopamina ou opioides. Essa inibição
diminuiria quando se aproxima a puberdade. A progesterona de origem ovárica
secretada antes da puberdade, embora em baixos níveis, pode também estar
afetando o padrão de secreção de LH.
O mecanismo controlador da secreção de LH inclui um efeito feedback inibitório
de alça curta sobre sua própria secreção determinado pela secreção pulsátil de
GnRH hipotalâmico. Portanto, cada pulso de LH está associado com um pulso
de GnRH. Assim, o hipotálamo pode ser visto como a via final que comanda os
vários estágios da atividade reprodutiva. O ritmo gerador de pulsos de GnRH
está afetado pelo estradiol de forma que durante períodos de alta sensibilidade
feedback, a atividade do gerador está diminuída.
Depois da puberdade ocorrem várias mudanças no controle da secreção de LH. O
sistema hipotâlamo-hipofisiário diminui sua capacidade para ser inibido por estradiol. O novo esteroide inibitório passa a ser a progesterona e o novo mecanismo que determina os intervalos entre os ciclos ovulatórios é a duração do corpo
lúteo. O aumento de progesterona durante a fase luteal diminui a frequência dos
pulsos de LH, inibindo o desenvolvimento dos folículos pré-ovulatórios. Com a
regressão do corpo lúteo e a queda nos níveis de progesterona, a frequência de
pulsos de LH aumenta e começa a fase folicular.
A possível explicação dos cios silenciosos no início da puberdade está no fato de
que a progesterona tem um papel importante como “iniciador” da ação estrogênica sobre o comportamento sexual. A sequência para a indução do estro seria a
seguinte: aumento de progesterona, diminuição de progesterona e aumento de
84
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
estradiol. Antes da 1ª ovulação não há formação de corpo lúteo e, portanto, não
se produz progesterona. A fase luteal subsequente é muito curta e não produz
suficiente progesterona, de forma que o estradiol circulante não consegue fazer
expressar o comportamento sexual do estro. Finalmente, após a exposição do
cérebro a altas concentrações de progesterona durante a 1ª fase luteal normal,
correspondente à 2ª ovulação, o cio pode ocorrer com os níveis de estradiol
circulantes durante a 3ª fase folicular (3ª ovulação). Contudo, os primeiros cios
podem ser curtos e pouco expressivos.
Em novilhas, a 1ª ovulação é seguida de um ciclo estral curto de 7 a 8 dias e a 2ª
ovulação por um ciclo estral normal (21 dias). Existe um período de estabilização
da função reprodutiva após o início da puberdade. A diferença entre puberdade
e capacidade fertilizante se expressa nas diferenças de idade entre o início da
puberdade em novilhas e a idade ao 1º serviço. As raças zebuínas são menos
precoces que as raças europeias.
O nível nutricional baixo aumenta a idade na qual se atinge a puberdade, talvez
por afetar a pulsatilidade do LH. Em novilhas, é desejável um peso de 350 kg para
o 1º serviço, o que pode implicar em uma alimentação consistente com ganhos
de 700 g/dia ou mais a fim de conseguir o primeiro parto com 2 anos de idade. O
mecanismo do efeito retardante da subnutrição sobre a época da puberdade não
está totalmente esclarecido. Muitas perguntas surgem, tais como, quais seriam os
sinais que inibem a atividade reprodutiva na subnutrição? Quando e como podem
ser monitorados? Trata-se de hormônios metabólicos (insulina, GH, hormônios
tireoidianos) ou de produtos de vias metabólicas? Os sensores são hipotalâmicos
ou extra-hipotalâmicos? Como se comunicam esses sensores com o gerador de
pulsos de GnRH para modificar sua atividade? De qualquer maneira, seja qual for
o mecanismo, tais sinais devem provocar aumento da sensibilidade ao feedback
negativo do estradiol causando inibição da secreção de GnRH e produzindo insuficiente LH para desenvolver os folículos.
O portal do agroconhecimento
85
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
O ambiente social é outro fator que afeta o início da puberdade. Porcas em
grupo acompanhadas de um cachaço chegam à puberdade antes das que estão
isoladas. O efeito é também observado em vacas, ovelhas e éguas. O mecanismo
pode ser através dos feromônios da urina do macho que estimulariam o sistema
feedback desencadeante da puberdade.
Finalmente, temperaturas extremas ou estresse inespecífico também são causas
de retardo ao início da puberdade, em virtude da ação do cortisol sobre o eixo
hipotalâmico-hipofisário.
2.6 Fisiologia do ciclo estral
2.6.1 Foliculogênese
O crescimento e o desenvolvimento folicular nos ruminantes se caracterizam por
duas ou três ondas foliculares consecutivas durante o ciclo estral. O advento da
ultrassonografia permitiu a coleta de muitas informações sobre os estágios do
crescimento folicular e sua seleção. Cada onda envolve o recrutamento de uma
série de folículos do total do estoque folicular do ovário, e a seleção de um folículo
dominante, que continua a crescer e a amadurecer até o estágio pré-ovulatório
enquanto os outros sofrem atresia.
Podem-se distinguir três estágios distintos no desenvolvimento folicular: crescimento, seleção e desvio.
Cada onda consiste no recrutamento simultâneo de três a seis folículos, que
crescem acima de 4 a 5 mm de diâmetro.
86
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
Dentro de alguns dias do início da onda, um folículo emerge como dominante.
O folículo dominante continua a crescer e a se diferenciar, ao passo que os
outros folículos param de crescer e regridem. Observa-se regressão do folículo
dominante da primeira onda em ciclos de duas ondas, assim como dos folículos
dominantes da primeira e segunda onda em ciclos de três ondas. Contudo, o
folículo dominante de qualquer onda folicular, inclusive da primeira, pode ovular
se forem fornecidas as condições endócrinas apropriadas pela indução de luteólise (por meio da administração de PGF2α) durante seu período de dominância.
2.6.1.1 O recrutamento de ondas de folículos
Nos bovinos e em outras espécies, as ondas foliculares são precedidas ou acompanhadas de um ligeiro aumento nas concentrações de FSH.
Todos os folículos que crescem em grupo contêm receptores específicos para
FSH e dependem dessa gonadotrofina para seu crescimento. Nesse estágio, os
folículos em crescimento não têm uma quantidade suficiente de receptores de
LH para responder a uma estimulação por LH (como na secreção pré-ovulatória),
que é a razão pela qual esse estágio de crescimento às vezes é chamado de FSH
dependente.
Nos bovinos, os aumentos sequenciais de FSH, acompanhados por ondas foliculares, ocorrem durante o ciclo estral, no período pós-parto, durante a prenhez e
antes da puberdade.
O portal do agroconhecimento
87
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
2.6.1.2 Seleção do folículo dominante
Na vaca, por razões que ainda não foram completamente esclarecidas, somente
um folículo dominante é selecionado do grupo recrutado. Uma característica que
parece definir um folículo como dominante é sua maior capacidade de produzir
estradiol e maior número de receptores para o FSH. A secreção de estradiol
pelo folículo dominante está associada ao bloqueio da liberação de FSH e sua
manutenção em níveis basais (GINTHER et al., 2000), impedindo o crescimento
dos folículos subordinados. O futuro folículo dominante adquire receptores de LH,
que lhe permitem continuar a crescer em um ambiente de baixos níveis de FSH
e com níveis crescentes de LH.
Ao provocar indiretamente a redução dos níveis de FSH, o folículo dominante
provoca redução do suporte dos folículos subordinados ao reduzir o componente
vital para seu crescimento, enquanto ao mesmo tempo se beneficia tanto do
baixo nível de FSH como do aumento dos níveis de LH.
Recentemente surgiram informações importantes sobre o papel de outros moduladores tais como fatores de crescimento, inibina e insulina na diferenciação e
seleção do folículo dominante (FORTUNE et al., 2001; MIHM et al., 2003).
2.6.1.3 Folículo dominante selecionado
O crescimento, a atividade estrogênica e o período de vida do folículo dominante
selecionado são controlados pelo padrão dos pulsos de LH. Portanto, qualquer
mudança no padrão de liberação do GnRH e, assim, do LH, terão um profundo
efeito sobre a continuidade do crescimento do folículo dominante e sua ovulação. Sabe-se bem que o aumento da frequência dos pulsos de LH observado
após os tratamentos com progestágeno, por exemplo, prolongarão o período
88
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
de dominância desse folículo de 2 a 7 dias para mais de 14 dias, o que afeta a
fertilidade do oócito (DISKIN et al., 2002). Fatores nutricionais, ambientais e até
infecciosos, que afetam direta e indiretamente o padrão de GnRH/LH em bovinos, apresentam um efeito considerável sobre o destino do folículo dominante
e, consequentemente, sobre a ovulação e a fertilidade.
Os eventos de desenvolvimento, recrutamento, seleção e desvio folicular estão
ilustrados na Figura 27, enquanto que o ciclo estral com 2 e 3 ondas foliculares
estão ilustradas nas Figuras 28 e 29, respectivamente.
Figura 27: Fases do crescimento folicular: recrutamento (dependente de FSH);
seleção e dominância (dependente de LH); ovulação ou atresia (na dependência da
presença ou não do pico pré-ovulatório de LH). FONTE: TECNOPEC.
O portal do agroconhecimento
89
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
Figura 28: Esquema de ciclo estral com 2 ondas foliculares. A primeira onda culmina
com a atresia do folículo dominante por falta do pico pré-ovulatório de LH (Fase
luteínica) e a segunda onda culmina com a ovulação (Fase folicular, baixa progesterona e pico pré-ovulatório de LH). FONTE: TECNOPEC.
Figura 29: Esquema de ciclo estral com 3 ondas foliculares. A primeira e a segunda
onda culminam com a atresia dos folículos dominantes por falta do pico pré-ovulatório de LH (Fase luteínica). A terceira onda culmina com a ovulação (Fase folicular,
baixa progesterona e pico pré-ovulatório de LH). FONTE: TECNOPEC.
90
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
2.6.2 Maturação oocitária
A maturação oocitária se divide em dois estágios:
»» Período de crescimento;
»» Período de preparação nuclear e citoplasmática final, que é prérequisito para a fertilização e o desenvolvimento final.
2.6.2.1 Crescimento oocitário
Quando um folículo primordial deixa a reserva, o oócito e seu folículo iniciam o
crescimento. O crescimento oocitário está quase completo na fase de formação
do antro. Com os processos celulares, a atividade das células internas do cumulus
cooperam ativamente com o processo de crescimento oocitário, tendo em vista
que essas células estabelecem um contato íntimo com a membrana celular do
oócito. Durante a formação da membrana externa do oócito (zona pelúcida), os
processos celulares nas células do cumulus são fortalecidos.
A maturação oocitária ocorre independentemente da natureza da estimulação
folicular, do diâmetro folicular do qual o oócito foi removido e da fonte do fluido
folicular ou seu filtrado, de diversos folículos e diferentes fêmeas.
2.6.2.2 Preparação do oócito para a fertilização
Da oogênese em diante, o núcleo do oócito, que se encontra em diplóteno,
permanece em repouso. A meiose normalmente não se reinicia antes do pico
pré-ovulatório de LH (Figura 30). A liberação de LH suprime a produção do fa-
O portal do agroconhecimento
91
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
tor inibidor da meiose pelas células da granulosa. O pico de LH é seguido por
modificações metabólicas na camada folicular. O reinício da meiose (maturação
nuclear) é apenas um aspecto da maturação oocitária. A maturação citoplasmática também deve ocorrer.
Figura 30: Oócitos em processo de maturação. FONTE: HAFEZ e HAFEZ (2004).
2.6.3 Ovulação
Na vaca, a ovulação ocorre após o estro, ou seja, no início de metaestro. O folículo
eleito recebe maior fluxo capilar, efeito influenciado pelo LH. A ovulação pode
ocorrer em qualquer ponto do ovário, exceto na égua que ocorre numa área conhecida como “fossa da ovulação”, onde fica concentrado o epitélio germinativo.
A ovulação ocorre aleatoriamente em qualquer um dos ovários, mas geralmente
92
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
ocorre de forma alternada.
Na medida em que a ovulação se aproxima, o folículo ovulatório é projetado
na superfície do ovário aumentando a sua vascularização, exceto numa área
central onde ocorrerá a ruptura do folículo. A dissociação das células do cumulus
oophorus liberta o ovócito e a meiose recomeça cerca de 3 horas após a onda de
LH, terminando 1 hora antes da ovulação, quando ocorre a extrusão do 1º corpo
polar e o ovócito secundário é formado.
As células do cumulus secretam glicoproteínas que rodeiam o ovócito e a corona
radiata. Ocorrendo a ruptura, a massa viscosa facilita a captação do ovócito pelas
fímbrias. As células granulosas dissociadas do folículo sofrem invasão das células
da teca interna e dos vasos sanguíneos.
A ação do LH e do FSH é responsável pela dissociação das células do cumulus, ao
mesmo tempo em que estimula a secreção de esteroides ovarianos. A progesterona estimula a ação da enzima colagenase na parede folicular e o estradiol
aumenta o edema da teca. As prostaglandinas, secretadas pelas células granulosas, também têm efeito sobre a dissolução da parede folicular participando da
ruptura do folículo (PGF2α) e da transformação das células granulosas em células
luteínicas (PGE2). Também ativam o plasminogênio para formar plasmina, a qual
está envolvida na migração celular (PGE2) e estimulam as contrações ovarianas,
as quais facilitam a ruptura folicular (PGF2α).
A partir daí, as células da teca e da granulosa remanescentes do folículo ovulatório se diferenciam em células luteínicas sob efeito do LH, para a formação do
corpo lúteo.
O portal do agroconhecimento
93
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
2.6.4 Ciclo estral
O ciclo estral é definido como a sequência de eventos endócrinos, morfológicos
e comportamentais que ocorrem entre dois estros ovulatórios sucessivos. O estro
ou cio é definido como período de receptividade sexual da fêmea (aceitação da
monta) e sua duração varia dependendo da espécie desde horas até dias. Na
vaca, o dia de início do estro costuma ser designado como o dia zero do ciclo e
tem duração média de 18 horas, com uma faixa de 4 a 24 horas. O ciclo estral da
vaca dura 21 dias, em média, com uma faixa de 18 a 24 dias.
O ciclo estral resulta da interação coordenada de 4 tecidos: sistema nervoso central, hipotâlamo-hipófise, ovário e útero. A comunicação entre esses órgãos ocorre principalmente mediante 6 hormônios: GnRH (do hipotálamo), LH e FSH (da
hipófise), estradiol e progesterona (do ovário) e prostaglandina F2α (do útero).
Três esteróides são produzidos pelo ovário durante o ciclo estral: progesterona,
estradiol e androstenediona. A progesterona é secretada exclusivamente pelo
corpo lúteo, o estradiol pelos folículos e a androstenediona é produzida pelo corpo
lúteo, pelos folículos e pelo estroma ovárico.
O crescimento e a maturação dos folículos ováricos nos mamíferos dependem
da ação sinérgica das gonadotropinas hipofisiárias FSH e LH. Ambas gonadotropinas participam também da síntese de estrógenos. O FSH da pituitária anterior
estimula o desenvolvimento dos folículos vesiculares e estimula as células da
granulosa para produzir estradiol, o qual se acumula no líquido folicular e sai para
a corrente circulatória.
O ciclo estral é classificado em fase folicular (proestro e estro) e fase luteal
(metaestro e diestro).
94
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
A fase folicular costuma ser definida como aquele período desde a regressão
do corpo lúteo até a ovulação e dura, em média, 4-5 dias na vaca. Entretanto,
durante a fase luteal existe crescimento de folículos apesar da natureza inibitória
da progesterona, principal hormônio secretado pelo corpo lúteo durante esta
fase, de forma que as fases luteal e folicular se sobrepõem. Ocorre uma segunda
onda de FSH, 20-30 horas após a onda pré-ovulatória de LH e FSH. A onda pósovulatória de FSH desenvolve e seleciona os folículos a crescer e ovular no ciclo
posterior.
2.6.5 Endocrinologia de vacas leiteiras de alta produção
A maioria dos conjuntos de dados disponíveis demonstra uma relação antagônica entre produção de leite e fertilidade. Todavia, a extensão desse efeito
vem sendo questionada, principalmente porque, no caso de muitos dos índices
reprodutivos, não se estabeleceram ainda relações claras com a produção de
leite. Mas, as observações no campo indicam claramente que as vacas leiteiras
de alta produção apresentam taxas de concepção muito mais baixas do que as
novilhas. O possível efeito negativo dos altos níveis de produção de leite sobre
o desempenho reprodutivo das vacas de alta produção pode ser modulado por
meio de vários aspectos da função reprodutiva.
Não existe uma confirmação uniforme na literatura de um efeito negativo dos altos níveis de produção de leite com a intensidade e duração do cio. Por outro lado,
tanto os profissionais veterinários como os criadores relatam que vacas leiteiras
de alta produção apresentam problemas em relação à detecção de estros. Em
um experimento relatado por Lopez et al. (2004), a duração do cio foi correlacionada positivamente com o pico das concentrações de estradiol e correlacionada
negativamente com a produção de leite. Wiltbank et al. (2006) sugeriram que
altos níveis de produção de leite levam a uma diminuição das concentrações de
O portal do agroconhecimento
95
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
estradiol circulante, resultando na diminuição da duração e intensidade do cio.
A diminuição das concentrações de estradiol também pode causar aumento do
tamanho do folículo devido à maior demora para a indução do cio pelo estradiol,
para o pico de GnRH-LH e ovulação das vacas de alta produção.
Parece claro que vacas de produção muito alta apresentam condições endócrinas diferentes em relação às vacas não-lactantes, em virtude de sua alta taxa
metabólica (balanço energético negativo). As vacas que produzem mais leite
desenvolvem folículos maiores, mas com concentrações menores de estradiol
circulante (LOPES et al., 2004). Além disso, vacas possuem maior volume de
tecido luteínico, mas concentrações reduzidas de progesterona circulante. A explicação mais plausível é que o metabolismo dos hormônios esteroides aumenta à
medida que aumenta a produção de leite nas vacas leiteiras lactantes, em decorrência gasto das reservas de gordura. Com isso, qualquer desequilíbrio energético
durante a fase inicial de lactação, o primeiro sistema orgânico a estagnar é o reprodutivo, pois o eixo hipotalâmico-hipofisário-gonadal é extremamente sensível
da falta de energia e exigente da mesma para seu perfeito funcionamento. Em
contrapartida, quando se retorna ao equilíbrio orgânico, o sistema reprodutivo é
o último a se reestabelecer.
Wiltbank et al. (2006) propuseram que algumas das alterações reprodutivas em
vacas leiteiras lactantes são causadas por um aumento dramático do metabolismo de esteroides devido à maior ingestão de alimentos e fluxo sanguíneo por
meio do fígado. Nas vacas leiteiras lactantes, a manutenção contínua de um alto
plano nutricional leva a uma elevação crônica do fluxo sanguíneo hepático, observando-se aproximadamente o dobro suas companheiras não-lactantes de porte e
idade semelhantes. Os resultados experimentais recentes indicam que, mesmo
com níveis semelhantes de produção hormonal, as concentrações circulantes de
hormônios esteroides são menores durante a lactação (SANGSRITAVONG et al.,
2002; WILTBANK et al., 2006).
96
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
Além das concentrações mais baixas de estradiol no início do estro, é provável
que também haja uma redução mais rápida no estradiol circulante após o pico
de LH devido ao maior metabolismo deste esteroide. Isto resultaria em menor
duração do cio em vacas de alta produção. O alto metabolismo de esteroides nas
vacas de altos níveis de produção de leite também pode apresentar um efeito
deletério mais profundo sobre a fertilidade. O folículo pré-ovulatório e o oócito
podem ficar expostos a um período prolongado de altos pulsos de LH, que, por
sua vez, podem levar à ovulação de um oócito superestimulado ou prematuramente ativado e, assim, a uma menor fertilidade. O pequeno aumento das concentrações de progesterona após a ovulação também pode reduzir a fertilidade
por causa da pior sobrevida dos embriões.
2.6.6 Fisiologia reprodutiva do Bos indicus
A fisiologia de vacas Bos indicus foi estudada em vários trabalhos de pesquisa
(BÓ et al., 2003). Hoje, sabe-se que as fêmeas Bos indicus apresentam duas,
três ou quatro ondas de crescimento folicular durante o ciclo estral e apresentam
menor diâmetro do folículo dominante e do corpo lúteo (BÓ et al., 2003), bem
como menores concentrações séricas de progesterona (SEGERSON et al., 1984),
em relação a animais Bos taurus.
Em estudos mais recentes, verificou-se que o diâmetro do folículo dominante no
momento do desvio é menor em vacas Nelore (6,0 a 6,3 mm) (SARTORELLI et
al., 2005; GIMENES et al., 2005) do que em vacas Holandesas (8,5 mm) (GINTHER
et al., 1996). Além disso, o diâmetro com o qual o folículo dominante adquire
capacidade de ovular em resposta à administração de LH em novilhas Nelore se
situa entre 7 e 8,4 mm (GIMENES et al., 2005), enquanto em vacas Holandesas a
ovulação em resposta ao LH só ocorre com diâmetro superior a 10 mm (SARTORI
et al., 2001).
O portal do agroconhecimento
97
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
O estro é o complexo de sinais fisiológicos e comportamentais que ocorrem antes
da ovulação. Os sinais do estro são: vaca em estação quando montada; vulva
edemaciada; mucosa vaginal hiperêmica; muco cervical transparente e elástico; inserção da cauda arrepiada, possivelmente com pequenas lesões cutâneas;
inquietude; formação de grupos; reflexo de Fehmen; lamber, empurrar, brigar,
montar outros animais; lordose e, possível redução da ingestão de alimento e/
ou da produção de leite.
Os sinais do estro, principalmente quando vários animais estão em proestro simultaneamente, são frequentemente interpretados erroneamente. De todos os
sinais, o reflexo de estação (em que o animal permanece em estação quando
montado) é a indicação mais confiável do cio. Diz-se, então, que a vaca está
“aceitando monta”.
Figura 31: Mudanças endócrinas, fisiológicas e comportamentais associadas ao
estro (aceitação da monta) na vaca. FONTE: HAFEZ e HAFEZ (2004).
98
IEPEC
Capítulo 2
Fisiologia reprodutiva da fêmea bovina
Em suma, durante o proestro, o folículo pré ovulatório secreta quantidades crescentes de estradiol. Nesse momento, a vaca tenta montar em outras vacas e
inicia a secreção do muco cervical. No início do estro, os níveis máximos de
estradiol desencadeiam o pico de LH, que causa a ovulação cerca de 10 a 12 horas
após o término do cio. O tônus uterino é máximo e o muco cervical é aquoso e
abundante. Enquanto que o corpo lúteo em desenvolvimento secreta quantidades crescentes de progesterona, ocorrem 2 ondas foliculares, uma na fase inicial
do ciclo estral e outra na fase luteínica intermediária (Figura 31).
2.7 Considerações finais
Para que ocorra o sucesso da manipulação hormonal na IATF e nos tratamentos de
falhas reprodutivas, faz-se necessário basicamente do conhecimento dos eventos fisiológicos e comportamentais dos animais. A partir destes conhecimentos,
podemos intervir e alterar os protocolos de IATF pré-estabelecidos no intuito
de melhorar os índices reprodutivos, baixando os custos com hormônios, bem
como melhorar a saúde reprodutiva do rebanho. Portanto, para entender todo o
contexto da IATF, é imprescindível o conhecimento da biologia reprodutiva dos
bovinos de leite.
O portal do agroconhecimento
99
Inseminação Artificial em Tempo Fixo (IATF)
em Bovinos Leiteiros
Material Complementar
.pdf
Estresse e função reprodutiva feminina.
capitulo-2-arquivo-1.pdf
Influência do Estresse Térmico na Atividade Reprodutiva de
Fêmeas Bovinas.
capitulo-2-arquivo-2.pdf
O estresse e a reprodução.
capitulo-2-arquivo-3.pdf
100
IEPEC
O Instituto de Estudos Pecuários é um portal que busca
difundir o agroconhecimento, realizando cursos e palestras
tanto presenciais quanto online. Mas este não é nosso único
foco. Com o objetivo principal de levar conhecimento à
comunidade do agronegócio, disponibilizamos conteúdos
gratuitos, como notícias, artigos, entrevistas entre outras
informações e ferramentas para o setor.
Através dos cursos on-line, o IEPEC oferece a oportunidade
de atualização constante aos participantes, fazendo com
que atualizem e adquiram novos conhecimentos sem
ter que gastar com deslocamento ou interromper suas
atividades profissionais.
w w w. i e p e c . c o m