Bio-Electricidade
2º Semestre - 2008/2009
1º ano - Mestrado Integrado em Engenharia Biomédica
IST / FMUL
Baseado nas aulas e no livro Textbook of Medical Physiology,
Guyton & Hall
Resumo por Inês Amorim
2
Índice
Transporte através de membranas ............................................................................................ 4
Membrana celular................................................................................................................. 4
Tipos de transporte ............................................................................................................... 5
Transporte em quantidade .................................................................................................. 10
Potenciais electrotónicos ........................................................................................................ 11
Potencial de Repouso .......................................................................................................... 13
Potencial de Acção .............................................................................................................. 14
Condução Nervosa .................................................................................................................. 17
Sinapses.................................................................................................................................. 19
Músculo .................................................................................................................................. 22
Músculo Esquelético ........................................................................................................... 24
EMG e ENMG – Electromiografia e Electroneuromiografia .................................................. 30
Músculo Liso ....................................................................................................................... 33
Músculo Cardíaco................................................................................................................ 39
ECG – Electrocardiograma ................................................................................................... 42
Sono e EEG - Electroencefalograma......................................................................................... 50
Efeitos da electricidade ........................................................................................................... 53
Efeitos fisiológicos da corrente eléctrica .............................................................................. 53
Aplicações da corrente eléctrica .......................................................................................... 55
Interacção laser-tecido............................................................................................................ 57
Ultrasons ................................................................................................................................ 60
Tópicos essenciais ................................................................................................................... 63
3
Transporte através de membranas
Membrana celular
A membrana celular reveste todas as células do organismo contribuindo para a sua protecção,
para o controlo da entrada e saída de substâncias da célula e, consequentemente, para a
manutenção da concentração correcta de diversas substâncias no fluido intra e extra-celular. É
formada essencialmente por fosfolípidos, proteínas, colesterol e glicoproteínas.
Na membrana celular, os fosfolípidos organizam-se numa camada dupla, a bicamada
fosfolipídica. As suas cabeças polares hidrofílicas orientam-se para o exterior da camada
enquanto as caudas apolares hidrofóbicas estão orientadas para o interior. Esta camada é
impermeável a substâncias solúveis em água, tais como iões e glicose, mas é permeável a
substâncias lipossolúveis, como o oxigénio, o dióxido de carbono ou o álcool.
Ao longo da camada fosfolipídica existem proteínas membranares, maioritariamente
glicoproteínas, com função de transporte. Essas proteínas podem ser intrínsecas/integradas
se atravessarem completamente a membrana plasmática, ou extrínsecas/periféricas, se se
mantiverem apenas à superfície.
Algumas proteínas intrínsecas inserem-se na membrana e formam poros que permitem a
passagem constante de substâncias hidrofílicas; outras dão origem a canais que se podem
encontrar abertos ou fechados e que permitem a difusão de determinadas substâncias entre
os meios intra e extra-celulares; algumas funcionam como bombas e são responsáveis pelo
transporte activo; encontramos ainda proteínas transportadoras que, através de um sistema
de comporta, alteram a sua configuração (sem gasto de energia metabólica) e se ligam a
moléculas e iões para permitir a sua difusão.
As proteínas extrínsecas encontram-se frequentemente ligadas a proteínas intrínsecas e
assumem um papel enzimático ou de receptores hormonais
4
Tipos de transporte
O transporte através das membranas celulares pode dar-se com ou sem a intervenção de
proteínas transportadoras, respectivamente transporte mediado e transporte não mediado, e
com ou sem dispêndio de energia metabólica, respectivamente transporte activo e transporte
passivo.
O tipo de transporte de cada substância depende da sua natureza e da sua concentração nos
meios intra e extra-celular. Enquanto substâncias lipossoluveis podem atravessar directamente
a membrana plasmática a favor do gradiente de concentração sem necessidade de mediação,
substâncias não lipossoluveis só são transportadas através de canais proteicos. Quando o
transporte se dá contra o gradiente de concentração dos meios, o movimento das substâncias
dá-se, necessariamente, por transporte activo.
Transporte não mediado
Difusão simples
Osmose
Transporte mediado
Difusão facilitada
Transporte activo
Osmose
Osmose é o nome que se dá ao movimento de moléculas de água, através de uma membrana
semipermeável, de um meio onde a concentração de soluto é menor para um meio onde essa
concentração é maior, ou seja, contra o gradiente de concentração. À pressão necessária para
travar este processo chama-se pressão osmótica (quanto maior for o gradiente de
concentração entre os dois meios maior será a pressão osmótica) e é dado pela mesma
fórmula que permite determinar a pressão de um gás. Na prática, podemos dizer que a
pressão osmótica é dada pela expressão:
P RTic
em que R representa a constante de gás ideal, T a temperatura absoluta, a coeficiente
osmótico, i o número de iões e c a concentração molar do soluto.
Osmole é a unidade (não SI) que mede o número de moles de partículas activas (1 mole de
glicose – 1 osm; 1 mole de NaCl (Na+ Cl-) – 2 osm) de um componente químico que contribuem
para a pressão osmótica. A osmolaridade define o nº de osmoles por litro de solução
enquanto a osmolalidade define o nº de osmoles por kilograma de solução.
5
Difusão
A difusão de partículas em líquidos ou gases corresponde ao movimento espacial aleatório
dessas mesmas partículas, a que também se dá o nome de movimentos Brownianos,
determinado pela sua energia térmica. Quando dois meios estão separados por uma
membrana, a difusão de partículas tende a equilibrar a concentração entre os dois lados da
membrana, sem dispêndio de energia, ou seja, as substâncias movem-se a favor do seu
gradiente de concentração – da solução hipertónica (mais concentrada) para a solução
hipotónica (menos concentrada).
A difusão que ocorre entre os meios intra e extra-celulares pode ser facilitada ou simples,
conforme intervenham ou não, respectivamente, proteínas membranares.
A difusão simples dá-se directamente através dos interstícios da membrana lipídica, para
moléculas lipossolúveis, ou através de poros, ou canais iónicos, e canais proteicos, para
substâncias hidrossolúveis. A sua taxa de difusão é directamente proporcional à solubilidade
das partículas.
Os canais proteicos são, na maioria das vezes, selectivos, permitindo apenas a passagem de
determinadas substâncias, e muitos deles apresentam ainda mecanismos de abertura e fecho.
A permeabilidade destes canais depende de características como o seu diâmetro, forma, carga
eléctrica e natureza química. A abertura e fecho dos canais controla a permeabilidade da
membrana a determinadas partículas e é facultado por extensões das próprias proteínas
transmembranares que, pela alteração da sua forma, abrem ou fecham os canais. A regulação
deste processo é controlada por dois mecanismos principais: voltagem (a proteína responde a
uma mudança do potencial eléctrico entre o os dois lados da membrana) ou ligação química (a
ligação de uma molécula à proteína promove a mudança da sua configuração).
Num canal proteico podemos distinguir:
- Porta, ou comporta: determina quando o canal está aberto ou fechado, controlando
a sua permeabilidade;
- Sensores: respondem a estímulos/sinais que regulam a abertura e fecho dos canais;
- Filtro de selectividade: características que determinam o tipo de moléculas que
atravessam o canal;
- Poro real do canal aberto: canal que permite a passagem de partículas por difusão
quando o canal se encontra aberto.
Dois dos canais proteicos mais importantes são os canais de sódio e potássio. Os canais de
sódio, também chamados canais lentos, têm cargas negativas nas suas paredes que atraem os
iões de sódio, que têm carga positiva. Já os canais de potássio, ou canais lentos, são mais
pequenos e as suas paredes têm carga neutra.
6
A difusão facilitada caracteriza-se por ser um transporte mediado passivo em que proteínas
transportadoras têm afinidade química com algumas moléculas, o que permite a sua ligação.
Quando a molécula se encontra ligada à transportadora, esta altera a sua configuração e abrese no lado oposto da membrana. Como a força de ligação entre a proteína e a molécula é
muito baixa, a agitação térmica da partícula transportada é suficiente para quebrar essa
ligação e permitir a sua libertação. Terminado o transporte, a proteína volta ao seu estado
inicial e está pronta para dar inicio a um novo processo de difusão facilitada.
A velocidade da difusão facilitada, ao contrário da da difusão simples, não é directamente
proporcional ao gradiente de concentração e atinge um valor máximo quando todas as
moléculas transportadoras estão activas, i.e., quando atinge o ponto de saturação. Por outro
lado, a velocidade do transporte também é limitada na medida que nunca ultrapassa a
velocidade com que a proteína transportadora é capaz de efectuar o transporte e voltar à sua
configuração inicial.
7
Transporte mediado
Maior velocidade
Para moléculas de peso molecular e solubilidade nos lípidos semelhantes, a velocidade do
transporte por difusão facilitada é superior à da difusão simples.
Saturação
A intensidade da difusão apresenta saturação devido ao número limitado de proteínas
transportadoras.
Especialização química
As proteínas transportadoras têm especificação química pelo que apenas permitem a ligação,
e consequente transporte, de moléculas específicas.
Inibição competitiva
Moléculas estruturalmente semelhantes podem competir no transporte.
Inibição por compostos
O transporte pode ser inibido por compostos que não estão estruturalmente relacionados
com o substrato a transportar.
Transporte Activo
Por vezes é necessário manter uma maior ou menor concentração intracelular de uma
substância do que a sua concentração no meio extracelular. Tal acontece, por exemplo, com os
iões de sódio, potássio, cálcio, hidrogénio, ferro e cloro, ureia, açucares, aminoácidos, etc. Para
manter essa diferença é necessário realizar transporte contra o gradiente de concentração, o
que requer o uso de energia por parte das proteínas transportadoras. A este tipo de transporte
dá-se o nome de transporte activo e, de acordo com a fonte de energia utilizada, pode
classificar-se em transporte activo primário ou secundário.
Transporte activo primário
A energia utilizada provém de
reacções exergónicas, como a
hidrólise de ATP ou de outros
metabolismos energéticos.
Transporte activo secundário
A energia utilizada é fornecida,
indirectamente, por gradientes
iónicos.
Um dos exemplos mais importantes de transporte activo primário é a bomba de sódiopotássio (Na+/K+), que transfere activamente iões sódio para o exterior das células e,
simultaneamente, iões potássio para o seu interior. Esta bomba electrogénica mantém a
diferente concentração destes iões nas regiões intra e extracelular e é assim responsável pela
criação de uma diferença de potencial entre os dois lados da membrana (o interior da células
está a um potencial inferior) que permite a transmissão nervosa.
8
A bomba Na+/K+ é formada por uma proteína transportadora que tem receptores para o sódio
(3 na zona intracelular) e para o potássio (2 na zona extracelular). Quando se ligam iões a
todos estes receptores verifica-se actividade de ATPase: uma molécula de ATP é transformada
em ADP e a energia libertada é utilizada para gerar uma alteração química e estrutural da
proteína que permite a expulsão de 3 Na+ do interior da célula e a entrada de 2 K+.
Em situações em que a energia gerada pelo gradiente de concentrações destes iões é superior
à energia libertada na hidrólise do ATP, a bomba Na+/K+ pode funcionar ao contrário, ou seja, o
sódio passa a entrar na célula, o potássio a sair e forma-se ATP a partir de ADP+Pi.
A bomba de sódio-potássio existe na maioria das células do nosso organismo e é responsável
pelo consumo de cerca de 60%-70% de toda a energia mobilizada para a célula. Para além da
sua importância crucial nos mecanismos de transmissão nervosa, esta bomba têm ainda um
papel muito importante no controlo do volume celular.
No interior da célula encontram-se muitas proteínas e moléculas orgânicas que não podem
escapar para o fluido extra-celular e que, por isso, favorecem a entrada de água na célula por
osmose. Sem controlo, a célula corre o risco de inchar indefinidamente e sofrer lise. A bomba
Na+/K+ ajuda a manter um gradiente de K+ para fora da célula, contrabalançando a tendência
de entrada de água e participando assim activamente no controlo do volume celular.
A elevada concentração de sódio no exterior da célula gera um gradiente que pode ser
interpretado como energia, participando no transporte activo secundário. Devido a esse
gradiente, a difusão de sódio para o meio intracelular, através de proteínas
transmembranares, pode promover a entrada ou saída de substâncias da célula contra o seu
gradiente de concentração, respectivamente, co-transporte e contra-transporte.
Co-Transporte
Contra-Transporte
São transportadas duas moléculas no As substâncias são transportadas em
mesmo sentido.
sentidos diferentes.
Ex: Na-Glucose, Na-AA (aminoácidos)
Ex: Na-Ca, Na-H
Para além do transporte através de membranas, por vezes é necessário realizar transporte de
substâncias através de camadas celulares, como, por exemplo, através do epitélio intestinal,
dos túbulos renais, das glândulas exócrinas e da vesícula biliar. Este transporte dá-se por um
mecanismo que engloba duas etapas básicas:
9
Transporte activo através da membrana celular de um lado da camada;
Difusão simples ou facilitada através da membrana do outro lado da camada.
Transporte em quantidade
A maioria das substâncias entra e sai das células por difusão ou transporte activo. No entanto,
estes meios não permitem o transporte de partículas de grandes dimensões, sendo esse
assegurado pela endocitose e pela exocitose.
Endocitose
Exocitose
Absorção de partículas por parte da célula.
Expulsão de substâncias que se encontram
Expansões da membrana envolvem as
em vesículas no interior da célula, por fusão
substâncias e dão origem a vesículas no meio
com a membrana plasmática.
intracelular.
Pinocitose: ingestão de solutos ou partículas
singulares.
Permite a libertação de secreções, tais como
Permite a absorção de substâncias cujas toxinas, produtos metabólicos, hormonas ou
dimensões ou características não permitem o neurotransmissores.
seu transporte por difusão ou transporte
activo, como as proteínas.
Fagocitoce: ingestão de partículas de grandes
dimensões, tais como bactérias, vírus ou
células,
através
de
expansões
citoplasmáticas.
Apenas é possível por alguns tipos de células,
como os macrófagos e outros tipos de
glóbulos brancos. Possibilita a ingestão de
microrganismos tendo um papel crucial na
defesa do organismo.
10
Potenciais electrotónicos
Existem potenciais eléctricos entre as membranas de todas as células do corpo mas apenas
algumas são excitáveis, ou seja, apenas algumas, como as células nervosas e musculares, são
capazes de gerar transformações rápidas desses potenciais através de toda a membrana. Para
além de constituírem um método de transmissão nervosa, as alterações locais do potencial
comandam variadas funções celulares, como acontece em diversas células glandulares,
macrófagos e células ciliadas.
A diferença de potencial através de uma membrana – potencial de membrana, deve-se à
concentração de substâncias e ao potencial eléctrico entre os dois lados da membrana. As
contribuições relativas desses dois factores podem ser definidas pelo potencial electroquímico
- numa célula, o potencial electroquímico da sua membrana é dado pela diferença de potencial
electroquímico entre os meios intra e extracelular.
Uma célula em repouso deve estar em equilíbrio electroquímico, i.e., a concentração dos
diversos iões e o potencial entre os dois lados da sua membrana deve manter-se constante ao
longo do tempo e não deve apresentar tendência para se modificar. A Equação de Nernst
permite calcular a diferença de potencial para a qual determinados iões, com determinada
concentração, entram em equilíbrio e, no caso de não se verificar esse equilíbrio, permite
predizer qual a direcção do fluxo iónico.
Equação de Nernst:
(soma do produto de RT pelo logaritmo neperiano da concentração de X no exterior sobre a
concentração de X no interior, com o produto de zF pela diferença de potencial electroquímico
entre os dois lados da membrana)
X
( X ) ext ( X ) int ( X ) RT ln
X
ext
int
zF Eext Eint
R – constante de gás ideal
T – temperatura absoluta (Kelvin)
[X+]int – concentração iónica no interior da membrana
[X+]ext – concentração iónica no exterior da membrana, geralmente considerada nula
F – constante de Faraday
z – carga eléctrica do ião
Eext-Eint – diferença de potencial electroquímico entre o exterior e o interior da membrana
Em equilíbrio, a diferença de potencial electroquímico é igual a zero. Portanto da equação
anterior, temos:
X
RT ln
X
ext
int
zF Eext Eint 0
11
Resolvendo para (Eext-Eint)
Eext Eint RT ln X int
zF
X
ext
Simplificando, se se pensar que o logaritmo de base 10 é igual a 2,303 ln, que as unidades de
RT/F podem ser em mV (a unidade que normalmente se usa em potenciais de membrana) e
que a 30 ºC, (2,303 x R x T) /F = 60 mV então, sabendo que este valor praticamente não muda
com a temperatura, pode usar-se a fórmula simplificada:
60 X
EMF
log
z
X
ext
mV
int
EMF: Força electromotriz
(O valor do potencial no exterior da membrana considera-se, tipicamente, nulo. Deste modo, o
potencial calculado pela equação anterior diz respeito ao potencial no interior da membrana.)
Quando se mede o potencial de uma membrana o valor encontrado, quando comparado com
o calculado na Eq. Nernst, pode indicar várias situações:
Se a diferença de potencial medida através da membrana for igual à calculada pela
equação de Nernst, está-se num estado de equilíbrio em que não há movimento iónico
apesar de se verificar um gradiente de concentração;
Se a diferença de potencial medida for do mesmo sinal mas superior à calculada, a
força eléctrica é superior à das concentrações e portanto determina a direcção dos
iões;
Se a diferença de potencial medida for do mesmo sinal mas inferior à calculada, a força
de diferença de concentrações é superior à eléctrica determinando a direcção dos
iões, que se movem de acordo com o seu gradiente de concentração;
Se a diferença de potencial medida for de sinal contrário à calculada, ambas as forças
têm a mesma direcção, não podendo haver equilíbrio. As duas forças vão puxar o ião
exactamente para o mesmo lado da membrana e portanto vai haver um desequilíbrio
iónico franco.
Apesar de muito útil, a equação de Nernst não tem em conta a concentração de diferentes
iões em simultâneo nem a permeabilidade (P) das membranas a esses mesmos iões,
assumindo que esta é total. Para entrar com estas variáveis em estudo utiliza-se a Equação de
Goldman:
P( Na ) Na
EMF 60 log
P( Na ) Na
P( K ) K
ext P( K ) K
int
P(Cl ) Cl
ext P(Cl ) Cl
int
ext
int
mV
Nesta equação, quanto maior for a permeabilidade a determinado ião maior será a sua
contribuição para o potencial da membrana, pelo que se torna importante definir o conceito
12
de condutância. Do ponto de vista da electrónica, a condutância é o inverso da resistência,
correspondendo a uma maior condutância uma menor resistência à passagem da corrente
eléctrica. No caso da permeabilidade iónica das membranas, quanto maior for a condutância
de determinado ião menor será a resistência oferecida pela membrana à sua passagem e,
portanto, maior será a permeabilidade desse ião.
Potencial de Repouso
Designa-se potencial de repouso o potencial da membrana de células/fibras nervosas quando
estas não estão a transmitir qualquer tipo de sinais nervosos e se encontram em equilíbrio
electroquímico. Na maioria das células, este potencial é negativo e tem o valor aproximado de
-90mV.
Para o estabelecimento do potencial de repouso contribuem diversos factores, sendo o mais
importante a elevada permeabilidade das membranas aos iões potássio e a sua baixa
permeabilidade aos iões sódio:
Potencial de difusão de K+: a concentração de potássio no interior das células é cerca
de 35x superior à sua concentração no exterior. A eq. de Nernst para estes iões diz-nos
que a sua contribuição para o potencial de repouso é de -94mV (Fig. A);
Potencial de difusão de Na+: a concentração de sódio no meio extracelular é 10x
superior à sua concentração intracelular. Pela eq. Nernst, a contribuição deste ião para
o potencial de repouso é de 61mV. Porém, há que ter em conta que a condutância do
sódio é cerca de 100x inferior à do potássio, pelo que as suas contribuições têm pesos
diferentes. Ao aplicar a eq. de Goldman para o sódio e potássio chega-se a um valor de
-86mV para o potencial de repouso (Fig. B);
Potencial de difusão do Cl-: o resultado da eq. de Nernst para o cloro indica um
potencial de -90mV, que coincide com o potencial da célula em repouso. Deste modo,
pode desprezar-se a contribuição deste ião;
Bomba Na+/K+: a actividade da bomba de sódio-potássio (bomba electrogénica) faz
com que saia mais sódio de uma célula do que entre potássio, levando a uma perda
contínua de cargas positivas no meio interno. Este desequilíbrio de cargas dá origem a
um potencial de -4mV no interior das membranas. Conjugando a contribuição desta
bomba com a dos canais de sódio e potássio, obtemos um potencial de repouso de 90mV (Fig. C);
Aniões intracelulares: a existência de aniões intracelulares incapazes de atravessar a
membrana pode contribuir com cargas negativas para um potencial de até -10mV em
alguns casos.
13
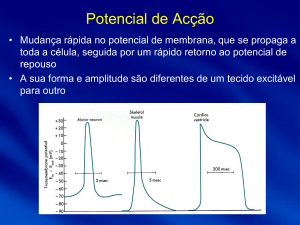
Potencial de Acção
Potencial de acção é uma mudança rápida do potencial da membrana, que se torna positivo
para de seguida retornar ao potencial de repouso, que se propaga a toda a célula. Esta
mudança dá-se em resposta a um estímulo eficaz (de origem variada: eléctrico, químico,
mecânico, …) e é característica dos tecidos excitáveis.
A propagação do potencial de acção compreende 3 etapas:
Repouso: antecede o potencial de acção. A membrana encontra-se polarizada, ou
seja, está ao potencial de repouso (mais negativo no interior);
Despolarização: em resposta a um estímulo eficaz a permeabilidade da membrana ao
sódio aumenta e a difusão destes iões para o interior da célula provoca um aumento
rápido do potencial da membrana. Em determinadas células, como nas fibras
nervosas, verifica-se overshooting - o potencial atinge valores positivos (o meio
intracelular fica a um potencial superior ao meio extracelular);
Repolarização: os canais de sódio começam a fechar e a permeabilidade ao potássio
aumenta gradualmente (mais lentamente do que a abertura dos canais de sódio na
despolarização) permitindo a rápida difusão destes iões para o exterior e o
restabelecimento do potencial de repouso. A determinada altura, o potencial da
membrana é inferior ao potencial de repouso - hiperpolarização, voltando depois a
subir ligeiramente até ao valor de equilíbrio.
14
Os canais de sódio e potássio que intervêm no potencial de acção são canais controlados por
voltagem, ou seja, a sua abertura/fecho está dependente da variação do potencial do meio.
Com o aumento do potencial da membrana (por exemplo, por um estímulo) os canais de sódio
abrem e permitem a difusão desses iões para o interior da célula, provocando um aumento
cada vez maior do potencial que, num ciclo vicioso, leva à abertura de mais canais de sódio.
Quando o potencial da membrana atinge determinado valor, os canais de potássio abrem
lentamente e os canais de sódio começam a ficar desactivados, dando-se início à
repolarização.
A abertura dos canais de sódio que dá inicio ao potencial de acção deve-se ao aumento do
potencial da membrana para um valor suficientemente elevado. Designa-se por limiar de
excitabilidade (limiar de acção ou threshold) o valor mínimo necessário para gerar um
potencial de acção, que é cerca de -65mV para a maioria das células nervosas. Caso esse limiar
não seja atingido, não há propagação de um potencial e os estímulos podem ser classificados
de acordo com a sua capacidade de atingir ou não esse limiar:
Estímulo não eficaz: gera uma resposta local mas não um potencial de acção;
o Estímulo sublimiar: não é suficiente para atingir o limiar de excitabilidade;
Estímulo eficaz: gera um potencial de acção;
o Estímulo limiar: é suficiente para atingir o limiar de excitabilidade;
o Estímulo supra-limiar: ultrapassa o limiar de excitabilidade.
Após atingido o limiar de excitabilidade, a resposta produzida é independente do estímulo - Lei
do tudo ou nada. Para um mesmo tipo de células, o potencial de acção gerado tem sempre a
15
mesma forma e amplitude. Por outro lado, apesar de ser necessário um estímulo limiar ou
supra-limiar para dar origem a um potencial de acção, há períodos em que, por mais elevado
que seja o estímulo, não é iniciada uma nova fase de despolarização. A esses períodos dá-se o
nome de períodos refractários:
Período refractário absoluto: período em que, qualquer que seja o estímulo, não é
possível gerar um novo potencial de acção. Limita a frequência de repetição dos
potenciais de acção.
Pouco após o estímulo inicial, os canais de sódio ficam inactivados e não podem ser
reactivados durante determinado período de tempo. Durante o período refractário
absoluto a maioria dos canais de Na+ encontram-se inactivados por voltagem e a célula
não é capaz de iniciar um novo potencial;
Período refractário relativo: período em que é necessário um estímulo maior que o
inicial para gerar um potencial de acção.
Alguns canais de Na+ ainda estão inactivados e a condutância ao K+ ainda está
aumentada, como no período de hiperpolarização.
No caso da acomodação, o limiar de excitabilidade também pode ser ultrapassado sem
desencadear um potencial de acção. Isto acontece quando uma célula é despolarizada
lentamente pois, nesse caso, alguns canais de sódio abertos têm tempo para ficar inactivados
antes de o seu número crítico desencadear um potencial de acção. Adicionalmente, alguns
canais de potássio abrem em resposta à ligeira despolarização, aumentando assim a
condutância ao K+ e hiperpolarizando a célula.
Após a repolarização, as concentrações de sódio e potássio ficam ligeiramente alteradas. A
actividade da bomba Na+/k+ restabelece as concentrações de equilíbrio desses dois iões e,
embora tal não seja necessário após cada potencial de acção, com tempo torna-se
imprescindível.
Como os potenciais de acção de uma célula têm todos a mesma forma e amplitude, a
informação que é transmitida tem de estar relacionada com a quantidade e frequência destes
impulsos.
16
Condução Nervosa
Na condução electrotónica de um potencial a condução decresce ao longo ao tempo, não se
propagando, geralmente, por mais de 10mm ao longo de uma célula nervosa. Este tipo de
condução pode ser utilizada a curtas distâncias e a sua eficácia depende essencialmente da
resistência da fibra nervosa e da distância de propagação: quanto maior for a distância e
quanto maior for a resistência oferecida pela fibra mais fraco será o sinal obtido. De um modo
geral, a velocidade da condução aumenta quando as fibras apresentam um revestimento de
mielina.
A condução regenerativa é a utilizada para fibras não mielinizadas e para condução por
grandes distâncias, pois o potencial de acção vai-se regenerando ao longo da fibra. Quando as
áreas adjacentes ao estímulo inicial atingem o limiar, regeneram o potencial e propagam-no
progressivamente a toda a célula, sempre com a mesma forma e amplitude.
A condução por este meio pode iniciar-se para dois sentidos da fibra, no entanto, devido aos
períodos refractários, uma vez iniciada a propagação num sentido esta mantém-se
unidireccional, ou seja, não regride.
Nas fibras mielinizadas a condução designa-se por condução saltatória, uma vez que o
potencial é propagado de nodo de Ranvier em nodo Ranvier em vez de continuamente ao
longo de toda a fibra.
As trocas iónicas nos locais cobertos pela bainha de mielina são praticamente nulas, pelo que
só é possível gerar um potencial de acção nos nodos de Ranvier. Quando se inicia um potencial
de acção num nodo criam-se correntes nas regiões próximas do fluido extracelular e também
17
no axoplasma (citoplasma dos axónios) que dão inicio a um novo potencial de acção no nodo
mais próximo.
Este modo de condução tem duas vantagens principais:
Velocidade de condução: permite uma condução mais rápida do potencial (5x-50x);
Poupança de energia: como só os nódulos é que sofrem despolarização há menos
perda de iões e, consequentemente, poupa-se energia a restabelecer as
concentrações iónicas de sódio-potássio.
A velocidade de propagação dos potenciais nas fibras nervosas depende essencialmente de
dois factores:
Diâmetro: quanto maior for o diâmetro da fibra menor será a resistência por ela
oferecida e mais rápida será a condução;
Revestimento de mielina: a mielina aumenta a velocidade de condução porque
diminui a capacitância da membrana (a membrana funciona como um condensador) e
restringe os potenciais aos nódulos de Ranvier. Uma fibra mielinizada chega a conduzir
mais rapidamente do que uma fibra não-mielinizada com um diâmetro 100x maior.
18
Sinapses
Os neurónios são constituídos pelo corpo celular, pelo axónio e pelas dendrites. As dendrites e
o corpo celular recebem informação e transmitem-na através do axónio para outro neurónio
ou para um órgão receptor. A zona específica de contacto/interacção intercelular,
morfologicamente específica, que faz a comunicação entre nervos (ou entre nervos e músculo
ou glândulas) e que permite a transmissão de potenciais designa-se sinapse. Nas dendrites e
soma (corpo celular) das células pós-sinápticas encontram-se terminais pré-sinápticos,
terminações de fibrilhas nervosas originárias nas células pré-sinápticas.
No caso dos neurónios, a comunicação entre si pode dar-se por:
Conexão directa (corrente, 2º mensageiro);
Moléculas de adesão;
Libertação de substâncias (receptores específicos, neurotransmissores).
A maioria das sinapses do sistema nervoso central são sinapses químicas, embora nas fibras do
músculo liso e cardíaco sejam comuns as sinapses eléctricas. Estes dois tipos de sinapses
apresentam diferentes características, que podem ser resumidas no quadro seguinte:
Distância
Continuidade citoplasmática
Ultra-estrutura
Sinapse Eléctrica
3,5 nm
Sim
Gap-Junction
Agente Transmissor
Direcção
Atraso sináptico
Corrente iónica
Bidireccional
Ausente
Sinapse Química
20 - 40 nm
Não
Vesículas pré-sinápticas
receptores pós-sinápticos
Neurotransmissores
Unidireccional
0,3 – 5 ms
e
Nas sinapses eléctricas as células pré-sináptica e pós-sináptica unem-se por canais proteicos
tubulares de baixa resistência – gap junctions, que permitem passagem directa de correntes
iónicas entre as duas células e a transmissão do potencial de acção. A transmissão do impulso
neste tipo de sinapses é bidireccional e muito rápida.
Nas sinapses químicas a resposta gerada é mais complexa e, consequentemente, mais lenta
do que nas sinapses eléctricas. A célula pré-sináptica secreta substâncias químicas, os
neurotransmissores, que são libertadas para a fenda sináptica e captadas por receptores
proteicos da membrana da célula pós-sináptica. De acordo com a sua natureza, os
neurotransmissores vão actuar nessa célula desencadeando uma resposta inibitória ou
excitatória.
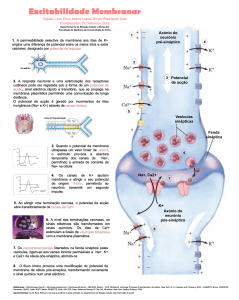
Quando um potencial chega à zona pré-sináptica, a despolarização da membrana provoca a
abertura de inúmeros canais de cálcio controlados por voltagem. Ao entrarem no terminal présináptico, os iões Ca2+ promovem a exocitose das vesículas sinápticas e a libertação de
neurotransmissores para a fenda sináptica.
19
Os transmissores que agora se encontram na fenda sináptica vão-se ligar a receptores
proteicos da membrana da célula pós-sináptica e desencadear aí uma resposta. Como já foi
dito, essa resposta pode ser excitatória ou inibitória. É de referir que a resposta final de um
neurónio se deve à acção conjunta de várias sinapses.
Resposta excitatória: neurotransmissores como o glutamano, a acetilcolina (SN
parassimpático) ou a noradrenalina (SN simpático), quando captadas pelos receptores
pós-sinápticos, actuam sobre a permeabilidade da membrana a diversos iões,
contribuindo para o aumento do seu potencial. Na maioria dos casos, há abertura dos
canais de sódio, o que leva à entrada de cargas positivas para o interior da célula, mas
também se pode verificar uma diminuição na condutância do cloro e do potássio, o
que dificulta, respectivamente, a entrada e saída destes iões do meio intracelular. É
ainda possível que a célula altere o seu metabolismo interno de forma a excitar a sua
actividade, a aumentar o número de receptores excitatórios da sua membrana ou a
diminuir a quantidade de receptores inibitórios.
Todas estas estratégias contribuem para elevar o potencial intramembranar (EPSP excitatory postsynaptic potential) e, se esse aumento for suficiente para atingir o
limiar de excitabilidade, gera-se um potencial de acção;
Resposta inibitória: a interacção dos neurotransmissores com os receptores póssinápticos provoca uma hiperpolarização da membrana sem transmissão de impulso
nervoso. A diminuição do potencial no interior na célula (IPSP – inhibitory postsynaptic
potential) pode dever-se à abertura dos canais de cloro (Cl- passa a entrar na célula),
20
ao aumento da condutância do potássio (sai mais K+ do meio intracelular) ou à
activação de receptores enzimáticos que aumentam o número de receptores
inibitórios e diminuem o número de receptores excitatórios.
Após a transmissão da informação nas sinapses químicas, sistemas enzimáticos destroem o
excesso de neurotransmissores que se encontram nas fendas sinápticas.
Nas sinapses químicas podem distinguir-se receptores que actuam de dois modos distintos:
Receptores ionotrópicos: actuam ao nível dos canais iónicos, promovendo a sua
abertura ou fecho e influenciando o movimento de iões;
Receptores metabotrópicos: entram no sistema de mensageiros secundários,
provocam alterações metabólicas.
De acordo com a zona onde ocorre a ligação sináptica, as sinapses químicas podem classificarse em:
Axo-axonal: ligação axónio-axónio;
Axo-somática: ligação axónio-corpo celular;
Axo-dendrítica: ligação axónio-dendrite.
21
Músculo
Músculo
Estriado
Esquelético
Liso
Cardíaco
22
23
Músculo Esquelético
O músculo esquelético é o tecido mais abundante no organismo humano (40%-45%). É ele que
liga os ossos e articulações, permite o seu movimento e a produção de força, e oferece ainda
protecção e resistência.
O músculo esquelético é um músculo estriado que tem uma componente contráctil
(miofibrilhas) e uma componente elástica (tendões, epimísio, perimísio). A sua unidade
estrutural é a fibra muscular, uma célula polinucleada, cilíndrica e alongada, revestida por uma
membrana, o sarcolema. Cada fibra estende-se ao longo de todo o músculo e é inervada
apenas por uma terminação nervosa, na região intermédia. As várias fibras que constituem o
músculo dispõem-se paralelamente entre si e são formadas por inúmeras miofibrilhas que
contêm filamentos de actina (finos) e de miosina (espessos), duas proteínas polimerizadas
responsáveis pela contracção muscular. No sarcómero (unidade funcional contráctil do m.e.),
esses filamentos interligam-se com outras proteínas contrácteis – tropomiosina e troponina,
que facilitam e promovem a contracção.
No sarcómero distinguem-se várias bandas causadas pelo intercalar dos filamentos de actina e
miosina, o que confere ao músculo um aspecto estriado. Essas bandas são:
Lina Z: linha em zig-zag, de grande densidade proteica, que delimita as laterais direita
e esquerda do sarcómero;
Banda I: bana clara, na periferia, constituída apenas por filamentos de actina;
Banda A: banda escura, entre as bandas I, composta por filamentos de miosina e, nas
extremidades, pela sobreposição de filamentos de actina;
Banda H: zona central do sarcómero onde apenas existem filamentos de miosina.
24
O mecanismo de contracção do músculo esquelético pode ser descrito pelos seguintes passos:
1. Um potencial de acção propaga-se ao longo de um nervo motor que se ramifica na sua
porção terminal e comunica com várias fibras nervosas (um ramo para cada fibra)
através de junções neuromusculares. Ao conjunto de todas as fibras inervadas por um
mesmo nervo dá-se o nome de unidade motora. Esta unidade é ainda a menor porção
de músculo que se contrai autonomamente.
Uma junção neuromuscular é um tipo particular de sinapse que faz a comunicação
entre um nervo e o músculo esquelético. É constituída por uma parte nervosa, a
terminação do nervo, e por um espessamento da parede do músculo, a chamada placa
motora;
2. Quando o potencial de acção atinge as terminações do nervo provoca a abertura de
canais de cálcio controlados por voltagem. A entrada de Ca2+ para o elemento présináptico promove a exocitose de vesículas com o neurotransmissor acetilcolina;
3. Na membrana da fibra muscular encontram-se receptores de acetilcolina que abrem
canais iónicos controlados por acetilcolina;
4. A abertura desses canais permite a difusão de grandes quantidades de Na + para o
interior da membrana o que dá origem a um potencial positivo, o potencial de placa
terminal. Esse potencial origina um potencial de acção.
Embora a maioria dos canais abertos não seja específica para o sódio, permitindo
também a difusão de outros iões positivos como K+ e Ca2+, verifica-se principalmente a
entrada de Na+ uma vez que o potencial negativo no interior da membrana evita a
saída de K+. Por outro lado, a concentração de Ca2+ é muito baixa quando comparada
com a sódio e potássio, o que lhe confere menor importância nesta fase;
5. O potencial de acção propaga-se pela fibra muscular do mesmo modo que nas fibras
nervosas. As miofibrilhas estão rodeadas por um sistema de túbulos-T (invaginações
do sarcolema) que permitem a rápida propagação do impulso ao longo de toda a fibra;
6. A despolarização da membrana leva à abertura dos retículos sarcoplasmáticos
(armazenam iões cálcio) e de canais de cálcio dos túbulos-T adjacentes. A saída de Ca2+
para o sarcoplasma que rodeia as miofibrilhas acciona o mecanismo de contracção do
músculo.
Essa contracção dá-se segundo a teoria do filamento deslizante, segundo a qual os
filamentos de actina deslizam sobre os filamentos de miosina, fazendo com que as
25
bandas H e I estreitem e acabem por desaparecer. Apesar de o sarcómero diminuir de
tamanho, os filamentos mantêm as mesmas dimensões.
Cada molécula de miosina é formada por cadeias enroladas em hélice que terminam
em duas cabeças. Essas moléculas agrupam-se para dar origem aos filamentos de
miosina e as suas cabeças organizam-se de modo a formar pontes cruzadas na lateral
do corpo do filamento.
Os filamentos de actina são constituídos por três proteínas:
A actina é a molécula central, que polimerizada forma uma dupla hélice que
contém sítios activos para ligação com a miosina;
A tropomiosina enrola-se em torno da actina e impede a ligação
actina/miosina bloqueando os sítios activos;
A troponina (existem 3 tipos) faz a ligação da tropomiosina à actina.
Quando o músculo está relaxado, os filamentos de actina e miosina não se encontram
ligados devido à acção do complexo tropomiosina-troponina. Na presença de cálcio
esse complexo é desactivado e as pontes cruzadas ligam-se aos sítios activos dos
filamentos de actina. A actividade da ATPase, que faz a hidrólise de uma molécula de
ATP, solta de novo os dois filamentos e permite que a ponte se ligue a um novo sítio
activo. O repetir deste ciclo leva ao encurtamento da fibra e à contracção do músculo;
26
7. Após uma fracção de segundo, uma bomba de Ca2+ armazena de novo esses iões no
retículo sarcoplasmático e a contracção termina. Até que chegue um novo potencial
de acção o músculo mantém-se relaxado.
Distinguem-se diferentes tipos de contracção:
Contracção isométrica ou estática: não há encurtamento do músculo mas há
desenvolvimento de tensão;
Contracção isotónica ou dinâmica: o músculo está preso apenas numa extremidade e
encurta após ser estimulado.
Define-se abalo muscular como a contracção rápida de um músculo em resposta a um
estímulo único. Um abalo compreende várias fases:
Período de latência: período entre a recepção do estímulo e a contracção efectiva;
Contracção;
Velocidade máxima de contracção;
Fase de relaxamento.
A força exercida por um músculo depende de um fenómeno chamado somação que consiste
na adição de contracções singulares de modo a aumentar a contracção final. Pode ocorrer
somação tanto pelo aumento do número de unidades motoras a contraírem em simultâneo
como pelo aumento da frequência de estimulação.
27
Há medida que se aumenta a frequência de estimulação as contracções ficam cada vez menos
espaçadas, acabando por se sobrepor e permitindo um aumento da força de contracção.
Nesses casos estamos na presença de um tétano fisiológico:
Tétano imperfeito: há sobreposição parcial de estímulos com períodos de relativo
relaxamento. A contracção resultante é mais forte do que a de um abalo simples;
Tétano perfeito: a frequência de excitação é tal que o músculo se mantém sempre
contraído e a força de contracção é máxima, não aumentando mais em função da
frequência. O tétano perfeito ocorre porque se mantêm no sarcoplasma iões de cálcio
suficientes, mesmo entre potenciais, para que não ocorra relaxamento do músculo;
Tétano de fadiga: ocorre normalmente depois de uma contracção forte e prolongada.
O músculo é incapaz de contrair devido a processos contrácteis ou metabólicos.
A força de contracção de uma fibra muscular está directamente relacionada com o
comprimento do sarcómero. Para uma fibra individual, verifica-se que para um comprimento
de cerca de 2m a tensão produzida é máxima, diminuindo depois tanto com o aumento como
com a diminuição do comprimento das fibras.
28
Relativamente à contracção do músculo como um todo, quando este se encontra no
comprimento normal de relaxamento, a tensão produzida após uma estimulação é máxima. A
variação da tensão exercida em função do comprimento do músculo está representada no
gráfico abaixo.
A velocidade de contracção está, por sua vez, associada à carga a que o músculo está sujeito.
À medida que a carga aumenta, a velocidade de contracção diminui.
Existem ainda diferentes tipos de fibras musculares que se diferenciam entre si essencialmente
pela velocidade de encurtamento e pela via de síntese do ATP. As características principais
dessas fibras encontram-se no quadro seguinte:
29
%
Resistência à fadiga
Velocidade de Fadiga
Velocidade de
contracção
Actividade miosinaATPásica
Fonte ATP
Activ. Enzimas
Glicolíticas
Nº mitocôndrias
Capilares
[Mioglobina]
Cor do músculo
[Glicogénio]
Diâmetro
Fibras Tipo I
(Lentas oxidativas)
50-55%
Resistentes à fadiga
Lenta
Fibras Tipo IIa
(Rápidas oxidativas)
30-35%
Resistentes à fadiga
Média
Fibras Tipo IIb
(Rápidas glicolíticas)
10-20%
Fatigáveis (10/20%)
Rápida
Lenta
Rápida
Rápida
Baixa
Alta
Alta
Fosforilação
oxidativa
Fosforilação
oxidativa
Glicólise anaeróbica
Baixa
Média
Alta
Muitas
Muitos
Alta
Vermelho
Baixa
Pequeno
Muitas
Muitos
Alta
Vermelho
Média
Médio
Poucas
Poucos
Baixa
Branco
Alta
Grande
A falta de utilização dos músculos leva à diminuição do seu volume - atrofia. Essa atrofia pode
ser derivada ao desuso (falta de exercício ou impossibilidade de utilizar determinados
músculos), à desnervação ou à diminuição do potencial oxidativo.
Enquanto num indivíduo sedentário a atrofia se inicia pelas fibras tipo I, num atleta inicia-se
pelas fibras mais desenvolvidas (por exemplo, tipo I nos maratonistas, tipo II nos
halterofilistas). Por outro lado, a prática regular de exercício pode levar à hipertrofia, ou seja,
ao desenvolvimento dos músculos. De acordo com o tipo de exercício, são desenvolvidos
diferentes tipos de fibras.
EMG e ENMG – Electromiografia e Electroneuromiografia
Entende-se por dermátomo a área cutânea inervada por fibras sensitivas de uma mesma raiz
nervosa e por miótomo o grupo de músculos inervados primariamente pelas fibras motoras de
uma única raiz nervosa.
O potencial de unidade motora (PUM) é a soma de todos os potenciais de fibras individuais de
unidade/s motora/s. Como nem todas as fibras se contraem em simultâneo, o PUM tem uma
duração mais longa do que um potencial singular e pode apresentar picos.
30
A Electromiografia (EMG) é uma técnica utilizada para monitorizar a actividade eléctrica dos
músculos. Os potenciais eléctricos são captados por eléctrodos de superfície ou por eléctrodos
em forma de agulha introduzidos no músculo e a interpretação da amplitude, número de fases
e duração de um potencial de unidade motora pode fornecer dados valiosos para a detecção
de patologias como neuropatias (inervação colateral) e miopatias (perda de fibras
musculares), e para a diferenciação das suas causas.
A Electroneuromiografia (ENMG) permite medir a actividade neuromuscular (tempo de
condução, tipo de potencial) através da estimulação eléctrica de nervos e monitorização dos
potenciais propagados. A interpretação da amplitude do sinal recebido, da sua duração, do
período de latência e da área, bem como o cálculo da velocidade de condução, da diferença de
amplitudes e da dispersão temporal, permite tirar conclusões relevantes quanto às condições
das fibras nervosas e quanto à possível existência de anomalias.
31
Duas patologias possíveis de detectar através de uma neurografia são:
Desmielinização: alteração do revestimento de mielina das fibras nervosas. Um
estímulo demora mais tempo a percorrer a mesma distância mas não apresenta
alteração do potencial;
Degenerescência axonal: diminuição do número de axónios fisiologicamente activos.
O tempo de transmissão é o normal mas o potencial é mais fraco do que o esperado.
O ENMG tem aplicações em diversas patologias, tais como:
Síndrome do túnel cárpico;
Neuropatia cubital do cotovelo;
Polineuropatia;
Radiculopatia;
Miopatia;
Etc
Em suma, o ENMG permite:
Localizar topograficamente lesões;
Diferenciar entre compromisso neuropático desmielinizante e axonal;
Caracterizar a gravidade das lesões;
Determinar um prognóstico.
32
Músculo Liso
O músculo liso encontra-se, por exemplo, nas artérias e no aparelho digestivo, e tem
contracção involuntária. Apesar dos seus princípios gerais de contracção serem idênticos ao do
músculo esquelético, o arranjo interno dos filamentos de actina e miosina é diferente (não
apresenta aspecto estriado) e as suas fibras são menores, mais finas e uninucleadas.
As características do tecido muscular variam de órgão para órgão, mas podem distinguir-se
dois tipos principais:
Músculo liso multi-unitário: é formado por fibras individuais que se contraem de
forma independente, maioritariamente por acção de sinais nervosos, e que são
inervadas por uma terminação nervosa simples. Cada fibra está envolta por tecido
conjuntivo rico em colagénio e glicoproteínas, o que ajuda ao seu isolamento.
Ex: íris e músculo erector do pelo;
Músculo liso unitário (visceral ou sincicial): as fibras contraem-se em conjunto, como
um todo (sincício), e as suas membranas estão ligadas em vários pontos por gapjunctions, o que permite a transmissão de impulsos (com origem em vários tipos de
estímulos) directamente entre fibras.
Ex: vasos sanguíneos, estômago, ureteres.
O músculo liso contém filamentos de actina e miosina com características químicas similares
aos filamentos do músculo esquelético: embora não apresentem o complexo de troponina, a
contracção destas fibras é activada por iões cálcio (processo diferente do do ME) e a energia
necessária a esse processo é fornecida pela degradação de ATP.
Relativamente às características físicas, o arranjo de actina e miosina não confere ao músculo
um aspecto estriado. As fibras de actina encontram-se ligadas a corpos densos que podem
estar dispersos na célula ou associados à membrana celular, e que se podem ligar entre si por
proteínas estruturais ou a corpos densos de outras células por pontes proteicas intercelulares.
Os filamentos de miosina apresentam um diâmetro maior e encontram-se, numa proporção
baixa, entre filamentos de actina. Os corpos densos podem ser comparados à linha Z de um
33
sarcómero, apesar de no músculo liso não existir a mesma regularidade que se encontra no
músculo esquelético, e a contracção das fibras também se dá por deslizamento dos filamentos.
As fibras do sistema nervoso autónomo que inervam o músculo liso ramificam-se na sua zona
terminal e o revestimento de mielina é interrompido ao longo do fim dos axónios, formando
varicosidades, que permitem a comunicação com o músculo.
Podem distinguir-se dois tipos de junções:
Junções difusas: a terminação nervosa está afastada da fibra nervosa (de alguns
nanómetros a alguns micrómetros) e a comunicação dá-se pela libertação de
substâncias transmissoras (acetilcolina e noradrenalina) para o meio extracelular;
Junções de contacto: em tudo semelhantes a uma junção neuromuscular, permitem
uma velocidade de contracção mais elevada do que nas junções difusas.
Apesar das semelhanças entre o processo de contracção dos músculos liso e esquelético,
existem algumas diferenças fundamentais, de entre as quais podemos destacar:
Ciclo de ligação entre pontes cruzadas lento: este ciclo é muito mais lento (10x-300x)
do que o do músculo esquelético o que indica uma menor actividade da ATPase;
34
Menor requerimento de energia: devido à velocidade reduzida do ciclo de ligação
entre pontes cruzadas, a energia necessária para manter o músculo em contracção
(gasta-se um ATP por ciclo) é também cerca de 10x a 300x menor;
Inicio de contracção e relaxamento lento: após recebido um estímulo, o início da
contracção e a sua duração é de cerca de 30x (este valor varia com o tipo de músculo
liso) mais lento do que o do músculo esquelético. Este facto deve-se à duração mais
prolongada do ciclo de ligação da miosina e também à resposta mais demorada à
libertação de cálcio;
Maior força de contracção: devido ao maior período de ligação actina-miosina;
Maior percentagem de encurtamento: as ligeiras diferenças no mecanismo de
contracção do músculo liso permitem-lhe uma percentagem de encurtamento cerca
de 2/3 maior;
“Latch Mechanism”: apenas sob a acção de pequenos e contínuos estímulos nervosos
ou hormonais, o ML é capaz de manter contracções prolongadas (por vezes durante
horas) com um gasto de energia muito reduzido;
Stress-relaxamento e reverso stress-relaxamento: permite que um órgão oco
mantenha a pressão no seu lúmen constante face a alterações de volume, excepto por
curtos períodos de tempo;
Regulação pelos iões cálcio: os iões cálcio também têm um papel fundamental na
contracção do músculo liso. Contudo, devido à ausência de troponina, o mecanismo de
contracção é activado por um processo totalmente diferente do do músculo
esquelético.
O inicio da contracção é modulado pela calmodulina, em vez da troponina, e ocorre
em três etapas:
1. 4 iões cálcio ligam-se à calmodulina, formando o complexo cálciocalmodulina, um complexo fosforilativo (só ligada a 4 iões cálcio a calmodulina
se torna eficaz);
2. O complexo cálcio-calmodulina activa a miosina-quinase e liga-se ao
caldesmon (tem um papel semelhante ao da tropomiosina), retirando-o do
contacto com a actina;
3. A miosina-quinase fosforila uma das cadeias leves de cada cabeça da miosina,
permitindo a ligação desta ao filamento de actina. Ocorre depois o
deslizamento entre os filamentos de actina e miosina, levando à contracção
do músculo.
Quando a concentração de iões cálcio baixa até um nível crítico o processo é invertido,
passando a miosina-fosfatase a desfosforalizar a cadeia leve da cabeça de miosina,
que se separa do filamento de actina e facilita o relaxamento do músculo;
35
Fonte dos iões cálcio: o retículo sarcoplasmático das células musculares lisas é pouco
desenvolvido, pelo que a maioria dos iões cálcio utilizados para activar o mecanismo
de contracção entra nas fibras por difusão a partir do fluido extracelular. Como este é
um processo mais demorado, o período latente (período entre a recepção do estímulo
e a contracção) é cerca de 50x mais longo do que o do músculo esquelético.
Nalgumas células existem clavéolas (pequenas invaginações da membrana análogas
aos túbulos-T do ME) adjacentes aos restículos sarcoplasmáticos que, na presença de
um potencial de acção, estimulam a libertação de Ca2+ por parte dos RS.
Ao contrário do que acontece no músculo liso, pequenas variações da concentração de
cálcio (diminuição de 1/3 a 1/10) levam ao fim da contracção. Esse facto faz com que a
36
força de contracção do músculo esteja dependente da concentração extracelular de
iões cálcio: quanto maior for essa concentração maior será a força de contracção.
Muito importante é também o papel da bomba de cálcio no relaxamento do músculo.
Esta bomba transfere Ca2+ do fluido intracelular de novo para o meio extracelular ou
para o RS. A sua actividade é lenta quando comparada com a bomba do músculo
esquelético, proporcionando uma contracção mais longa;
Estímulos de contracção: a contracção do músculo liso pode ser estimulada por
diferentes factores, desde potenciais de acção a hormonas e estiramento. Verifica-se
que cerca de metade de todas as contracções são por estímulos não nervosos e sem
propagação de potenciais de acção. Independentemente do tipo de estímulo, pode
ocorrer despolarização ou hiperpolarização da membrana.
Os estímulos podem ser devidos a:
o Estiramento: o estiramento do músculo liso unitário pode provocar potenciais
de acção espontâneos;
o Hormonas: em alguns órgãos existem receptores hormonais excitatórios ou
inibitórios que são activados na presença de determinadas hormonas. Ex:
noraepinefrina, epinefrina, acetilcolina, oxitocina, serotonina, histamina, etc;
o Efeitos tecidulares locais: falta de oxigénio, excesso de dióxido de carbono,
temperatura, presença de determinadas substâncias, entre outros factores,
podem influenciar a contracção/relaxamento do músculo;
o Estímulo nervoso: transmitido através de fibras nervosas.
O potencial de membrana do músculo liso ronda os -50mV a -60mV e pode ser modificado
pelos factores já mencionados e dar origem a potenciais locais ou a potenciais de acção. Um
potencial de acção ocorre quase exclusivamente no músculo liso unitário e pode apresentar-se
sob várias formas:
Potencial em ponta (tipo “spkie”): potencial de acção típico, como o que ocorre no
músculo esquelético. Tem uma duração de 10ms a 50ms;
Potencial em plateu (planalto): a fase de despolarização é semelhante à do potencial
em ponta mas a repolarização é longa (pode durar até 1s), mantendo o músculo
contraído durante mais tempo;
Potencial de ondas lentas (ondas de pace-maker): localmente, as fibras musculares
podem excitar-se espontaneamente com determinado ritmo (por razões ainda
desconhecidas). Se a despolarização resultante for suficiente para atingir o limiar
então gera-se um potencial de acção.
No músculo liso os potenciais de acção são gerados principalmente por canais de cálcio
controlados por voltagem, em vez de canais de sódio. Estes canais abrem mais lentamente e
mantêm-se abertos durante mais tempo, o que permite explicar a origem dos potenciais em
plateu.
37
No músculo liso multi-unitário é comum ocorrer despolarização sem potencial de acção, pois
as fibras são muito pequenas para desencadear um potencial. Dá-se então uma despolarização
local que é transmitida electrotonicamente.
38
Músculo Cardíaco
O músculo cardíaco é um tipo de músculo estriado. As fibras musculares comunicam entre si
através de discos intercalares que permitem o movimento livre de iões e a transmissão do
potencial de acção. As células actuam assim sob a forma de sincícios – sincício auricular e
sincício ventricular, que se contraem a comandos diferentes de forma síncrona. A
coordenação da contracção do coração é feita pelo tecido cardionector, ou tecido nodal, um
tecido condutor especializado na produção e condução dos impulsos eléctricos e que é
formado por:
Nódulo Sinusal, ou nódulo sinoauricular (SA): localizado na parede postero-superior da
aurícula direita é o local onde se originam os impulsos. O esqueleto fibroso do coração
não permite a propagação directa dos potenciais para os ventrículos, dando-se essa ao
nível do feixe de His. Dispara 60 a 100 vezes por minuto;
Nódulo aurículo-ventricular (AV): localiza-se na zona mediana da válvula aurículoventricular direita e atrasa a propagação dos impulsos eléctricos permitindo a
separação temporal entre a contracção auricular e ventricular;
Feixe de His: tem origem no nódulo aurículo-ventricular e atinge o septo
interventricular, dividindo-se no ramo direito e esquerdo. O ramo esquerdo estendese por baixo do endocárdio e atinge a região apical dos ventrículos onde termina na
rede de Purkinje;
Rede de Purkinje: rede de fibras musculares cardíacas modificadas.
As principais características do tecido cardíaco são:
Automatismo: capacidade de gerar estímulos que podem originar contracção;
Dromotropismo (condutibilidade): poder de condução do estímulo pelas fibras de
modo a que o estímulo chegue a todo o sincício;
Batmotropismo (excitabilidade): capacidade de reagir quando estimulado;
Ionotropismo (contractilidade): pode ser negativa (diminui força de contracção) ou
positiva (aumenta);
Tonacidade.
39
O potencial de repouso do músculo cardíaco normal é de -85mV a -95mV e o potencial das
fibras de Purkinje atinge os -90mV a -100mV. Os seus potenciais de acção têm características
singulares:
Longa duração: são 3 a 15 vezes mais longos que os do músculo esquelético;
Plateu (planalto): após o pico inicial, o potencial da membrana mantém-se elevado
durante um período de tempo relativamente longo ao qual se segue uma rápida
repolarização. A abertura dos canais rápidos de sódio leva ao pico inicial e o plateu é
mantido pela acção dos canais lentos de cálcio e pela baixa permeabilidade da
membrana ao potássio;
Período refractário: devido à existência de plateu, o período refractário absoluto é
mais longo. Músculo ventricular: absoluto – 0,25s a 0,3s; relativo – 0,05s. Músculo
auricular: absoluto – 0,15s; relativo – 0,03s;
Velocidade de condução: nas fibras musculares a velocidade de condução dos
impulsos é de cerca de 0,3 a 0,5m/s, 1/10 da das fibras musculares esqueléticas. No
tecido cardionector essa velocidade é de 0,02 a 4 m/s.
As características referidas ilustram mais fielmente o potencial de acção que se propaga ao
nível do ventrículo. Ao nível nas aurículas e do nódulo sinusal, o potencial apresenta algumas
diferenças que estão relacionadas com o tipo de canais iónicos que intervêm na
despolarização e repolarização.
O mecanismo de contracção do músculo cardíaco é semelhante ao do músculo esquelético:
Um potencial de acção atravessa a membrana do músculo cardíaco, espalhando-se
para o seu interior através dos túbulos T;
40
Abrem-se dois tipos de canais: canais rápidos de sódio e canais lentos de cálcio. A
permeabilidade da membrana ao potássio baixa para 1/5 (K+ é impedido de sair da
célula, contribuindo para que o potencial se mantenha positivo);
O potencial actua na membrana do retículo sarcoplasmático levando à libertação de
iões de cálcio aí armazenados;
A troponina C, fosforilada previamente pelas catecolaminas, liga-se aos iões cálcio;
O complexo formado liberta a actina/miosina do efeito de inibição a que estes
filamentos estavam sujeitos e permite a sua interacção;
Dá-se o encurtamento do sarcómero e a contracção do músculo cardíaco;
O potencial de acção termina e, pouco depois, o músculo relaxa.
A rápida repolarização após o plateu deve-se à acção conjunta do fecho dos canais de
cálcio, do aumento da condutância do sódio e da actividade do fosfolambam.
O fosfolambam é uma proteína que capta Ca2+ (com gasto de ATP) para o RS para que
este não fique nas imediações da troponina e estabeleça novas ligações, dando origem
a um novo ciclo de contracção.
Para o acoplamento excitação-contracção (mecanismo pelo qual o potencial de acção causa a
contracção das miofibrilhas) contribui grandemente a concentração extracelular de cálcio e as
características do túbulos T: os túbulos T do músculo cardíaco têm um diâmetro 5x maior do
que os do músculo esquelético e encerram um volume 25x maior. Durante o potencial de
acção, para além do Ca2+ proveniente do RS, grandes quantidades de cálcio difundem dos
túbulos T para o sarcoplasma.
41
O ciclo cardíaco engloba a sístole (contracção) e diástole (relaxamento) auriculares e a sístole e
diástole ventriculares. O tempo de sístole é sempre o mesmo e é mais curto que o de diástole,
pelo que quando a frequência cardíaca aumenta é a duração da diástole que diminui.
A diástole, nomeadamente a ventricular, é extremamente importante porque permite que os
ventrículos se encham de sangue que vai ser bombeado. Se a frequência cardíaca for muito
elevada (taquicardia), o ventrículo não enche totalmente e o débito cardíaco diminui. Se o
ritmo normal não for reposto rapidamente, o fluxo sanguíneo pode parar.
ECG – Electrocardiograma
O Electrocardiograma (ECG) regista o gráfico dos potenciais eléctricos gerados pelo coração
(despolarização e repolarização do miocárdio), detectados por eléctrodos metálicos colocados
nas extremidades e na parede torácica.
Esta técnica é muito utilizada como exame complementar porque apresenta várias
características muito vantajosas:
Não invasiso;
Simples;
Versátil;
Colheita e interpretação simples;
Reprodutível, económico;
Detecta múltiplas patologias.
Pode ser muito útil para conhecer:
A orientação anatómica do coração;
O tamanho relativo das diversas câmaras cardíacas;
Uma variedade de alterações do ritmo e condução;
A extensão, localização e progressão de lesões isquémicas do miocárdio;
Os efeitos de alterações de concentrações de electrólitos;
A influência de determinados fármacos.
Em qualquer momento, o potencial gerado no coração pode ser representado por um dipolo,
constituído por uma carga positiva e outra negativa, separadas por uma curta distância.
Se analisarmos uma célula estimulada, quer a partir do exterior quer a partir do interior,
vemos que na zona de transição (entre a zona despolarizada e em repouso) existe um dipolo
42
que progride ao longo do tempo. Esse dipolo gera uma força que tem direcção, sentido e
amplitude definidas, pelo que pode ser representada por um vector.
Quando um dipolo é emerso num volume condutor, é gerado um campo eléctrico que se
distribui de forma simétrica, caso o volume seja homogéneo. No organismo humano, tal
volume não é homogéneo e, até atingir a superfície onde é captado por eléctrodos, o campo
eléctrico passa por tecidos com propriedades distintas. De entre os factores que influenciam a
transmissão podemos destacar:
Factores celulares: resistência intra e extracelular;
Factores cardíacos: anisotropia e tecido conjuntivo;
Factores extra-cardíacos: sangue inrta e extra-cardíaco, pericárdio, pulmões, músculo
esquelético, gordura, pele;
Factores físicos: quadrado da distância, ângulos.
Dependendo do local onde é efectuada a medição e do sentido do dipolo resultante da
actividade cardíaca num dado momento, um ECG pode apresentar dois tipos de deflexão:
Deflexão positiva: o dipolo é representando por um vector que se aproxima da
posição de registo;
Deflexão negativa: o dipolo afasta-se da posição de registo.
Num ECG normal é possível identificar um sinal típico, com uma forma e intervalos
característicos, embora com variações de indivíduo para indivíduo. Podem identificar-se
diversas ondas e intervalos que correspondem a diferentes etapas do ciclo cardíaco:
43
Onda P: activação do nódulo sinusal e despolarização das aurículas (a direita
despolariza primeiro que a esquerda);
Intervalo PQ ou PR: activação completa das aurículas e atraso no nodo AV. Linha
isoeléctrica;
Onda Q: activação do nódulo AV e repolarização das aurículas;
Onda R: despolarização do miocárdio interno;
Onda S: despolarização do miocárdio externo;
Complexo QRS: despolarização ventricular;
Segmento ST: activação completa do ventrículo. Linha isoeléctrica;
Onda T: repolarização dos ventrículos. Esta onda é positiva porque a repolarização
começa no exterior, seguindo de fora para dentro (ao contrário da despolarização);
Onda U: potenciais tardios do inicio da diástole. Só está presente nalguns ECG,
sobretudo quando as frequências são mais baixas em determinadas derivações;
Intervalo QT: despolarização e repolarização dos ventrículos.
Os aspectos mais importantes a notar num ECG são:
Onda P;
Intervalo PR;
Complexo QRS;
Segmento ST;
Onda T.
44
45
O registo é feito em papel milimétrico colocado no electrocardiógrafo. O papel corre a
velocidade standardizadas, geralmente 25 mm/s (cada uma das quadrículas mínimas
corresponde a 0,04 s) e a amplitude é também standardizada, tendo-se uma calibração no
registo de tal maneira que cada mV corresponde a 10 mm (10mm/mV).
Apesar de a forma geral das ondas e dos intervalos de um ECG se manter qualquer que seja a
derivação utilizada (localização dos eléctrodos), o registo efectuado vai sempre depender das
derivações e da orientação do eixo eléctrico do coração.
Utilizam-se diversas derivações que, no seu conjunto, permitem obter uma representação tridimensional da actividade eléctrica cardíaca. Essas derivações são:
Derivações bipolares dos membros: DI, DII, DIII;
Derivações unipolares aumentadas dos membros: aVR, aVL, aVF;
Derivações precordiais: V1, V2, V3, V4, V5, V6.
As derivações biploares dos membros (clássicas) registam os potenciais eléctricos no plano
frontal e representam uma diferença de potencial entre dois locais seleccionados:
DI: diferença de potencial entre o braço esquerdo e o braço direito (VLA - VRA);
DII: diferença de potencial entre a perna esquerda e o braço direito (VLL - VRA);
DIII: diferença de potencial entre a perna esquerda e o braço esquerdo (VLL - VLA).
Em 1913, Einthoven desenvolveu um método de estudo da actividade eléctrica do coração
representando-a graficamente numa figura geométrica bi-dimensional: um triângulo
equilátero. No triângulo de Einthoven, o coração está localizado no centro e os vértices do
triângulo são indicados pelas 3 derivações.
Embora não seja matematicamente verdade, este método fornece ao clínico um conceito
prático com que pode trabalhar. Einthoven foi ainda mais longe e formulou uma lei baseada
nas leis de Kirchoff, a lei de Einthoven, que diz que se o potencial eléctrico de duas quaisquer
derivações bipolares for conhecido num dado instante, a terceira pode ser calculada pela
fórmula: DII = DI + DIII.
46
As derivações unipolares (dos membros e pré-cordiais) foram introduzidas por Wilson em
1932 e medem a diferença de potencial entre um eléctrodo indiferente e um eléctrodo
explorador.
Eléctrodo indiferente: formado por três fios eléctricos que estão ligados em série com
resistências de 5M e ligados entre si a um terminal central. As extremidades livres
destes fios ligam-se aos eléctrodos do braço esquerdo (LA), braço direito (RA) e perna
esquerda (LL). Considera-se que a soma dos três potenciais LA+RA+LL é igual a zero, ou
seja, o potencial do eléctrodo indiferente é zero;
Terminal central: liga-se ao pólo negativo do electrocardiógrafo;
Eléctrodo explorador: liga-se ao pólo positivo. Por princípio, as derivações unipolares
tentam medir potenciais locais e não diferenças de potencial.
Nas derivações unipolares aumentadas dos membros mede-se o potencial em três membros
(RA, LA, LF) ligando o terminal positivo do electrocardiógrafo directamente ao membro em
questão e mantendo os outros eléctrodos ligados ao terminal central e acoplados às
resistências. O potencial registado vem aumentado.
As derivações utilizadas são:
aVR: potencial no braço direito;
aVL: potencial no braço esquerdo;
aVF: potencial na perna esquerda.
47
As derivações unipolares pré-cordiais permitem fazer o registo de potenciais no plano
horizontal. O eléctrodo indiferente permanece ligado às três extremidades, enquanto o
eléctrodo explorador varia de posição ao longo da parede torácica. Uma derivação unipolar
feita por este método é denominada pelo prefixo V (de voltagem) seguido de um número, que
indica a sua posição correspondente:
V1: quarto espaço intercostal direito junto ao esterno;
V2: quarto espaço intercostal esquerdo junto ao esterno;
V3: equidistante de V2 e V4;
V4: quinto espaço intercostal esquerdo na linha médio-clavicular;
(todos os eléctrodos seguintes são registados no mesmo plano horizontal de V4)
V5: linha axilar anterior;
V6: linha axilar média.
O eixo eléctrico do coração pode ser definido por um vector com origem no centro do
triângulo de Einthoven e indica qual a direcção preponderante do fluxo de corrente durante a
despolarização dos ventrículos. Embora o termo eixo eléctrico possa ser usado relativamente a
qualquer um dos principais componentes do ECG (ondas P, T e complexo QRS), é geralmente
aplicado apenas ao complexo QRS. Existem vários métodos para determinar o eixo eléctrico
médio.
48
O vector médio do QRS no plano frontal, ou eixo eléctrico médio do complexo QRS,pode ser
calculado a partir das derivações bipolares dos membros DI, DII e DIII usando o sistema de
referência triaxial derivado do triângulo de Einthoven. Uma forma acessível de calcular o eixo é
usar a amplitude total (subtraindo os potenciais negativos aos positivos) dos complexos QRS
nas derivações bipolares dos membros.
Os valores do eixo eléctrico médio, em indivíduos adultos, variam entre -30º e 110º. Valores
diferentes são reconhecidos como desvios do eixo:
Desvio esquerdo do eixo: Valores entre -30º e -90º (DI positivo e DII negativo);
Desvio direito do eixo: Valores entre +110º e +180º;
Desvio extremo do eixo: Valores entre -90º e +180º (DI e DII negativos).
49
Sono e EEG - Electroencefalograma
O ciclo sono-vigília é um ciclo, com periodicidade de 24 horas, durante o qual o organismo
altera entre um estado de sono e de vigília.
O sono é um estado de alteração de consciência reversível, que ocorre de forma cíclica e que é
imprescindível à vida. Do ponto de vista comportamental, pode ser definido por 4 critérios:
Diminuição da actividade motora;
Diminuição da resposta à estimulação externa;
Posturas estereotipadas, como estar deitado de olhos fechados;
Relativa facilidade de reversibilidade (ao contrário do coma).
Pensa-se que a principal função do sono seja a de restaurar o balanço natural dos centros
nervosos e, embora ainda existam factos contraditórios por explicar, reconhecem-se-lhe
outras funções:
Conservação de energia: durante o sono o metabolismo baixa cerca de 15% e a
privação de sono leva a uma maior ingestão de alimentos;
Cognição: privação de sono afecta o desempenho intelectual;
Termorregulação: a temperatura do corpo e do cérebro baixa durante o sono;
Maturação neuronal e saúde mental: privação de sono REM pode dar alterações
comportamentais e está associado à aprendizagem.
Os estudos poligráficos do sono podem ser feitos recorrendo a diferentes técnicas e
parâmetros:
EOG (Electroculograma): mede o potencial da retina;
EMG(Electromiograma) submentoniano e tibial: monotoriza movimentos musculares
durante o sono;
EEG (Electroencefalograma): regista a actividade eléctrica cerebral;
ECG (Electrocardiograma): regista a actividade cardíaca durante o sono;
Fluxo nasal, movimentos torácicos e abdominais: monitorizam os movimentos
respiratórios;
Saturação de O2: mede os níveis de saturação de oxigénio do sangue.
O Electroencefalograma (EEG) é um exame complementar de diagnóstico que, por intermédio
da colocação de vários eléctrodos segundo um sistema internacional e das derivações entre
esses eléctrodos, permite registar a actividade eléctrica cerebral.
A actividade eléctrica registada num EEG resulta das correntes extracelulares criadas pelos
neurónios excitados e depende de vários factores:
Distância entre o gerador e o eléctrodo: quanto maior for a distância mais fraco é o
sinal registado;
50
Orientação dos geradores: num EEG só é detectada a actividade dos geradores radiais
(perpendiculares ao córtex);
Número e sincronização dos geradores: quanto maior for o número de geradores
maior será a amplitude do sinal captado. No entanto, caso a excitação seja
dessincronizada, é possível que os sinais emitidos se anulem entre si e o registo seja de
baixa amplitude.
Num EEG, para além de ondas dessincronizadas, podem registar-se 4 tipos de actividade
ritmada:
Actividade Alfa : ondas de baixa amplitude e frequência entre 8Hz e 13Hz.
Predomina na região occipital. É registada quando um indivíduo se encontra em
descanso, deitado e de olhos fechados, mas acordado;
Actividade Beta : ondas de baixa amplitude e frequência maior que 13Hz
(geralmente entre 14Hz e 80Hz). Predomina no lobo frontal e parietal. Verifica-se
quando um indivíduo está acordado, de olhos abertos e num estado normal de
actividade;
Actividade Teta : ondas de baixa frequência, entre 4Hz e 7Hz. Presente nas regiões
temporais e central. Regista-se numa fase de sonolência;
Actividade Delta : ondas de alta amplitude e muito baixa frequência, entre 0,5Hz e
4Hz. É registada em fases de sono profundo ou coma, ou em zonas localizadas que
apresentem lesões cerebrais.
Um EEG ajuda a caracterizar os diferentes tipos e estágios do sono:
Sono Não-REM: apresenta 4 estágios durante os quais as ondas registadas num EEG se
tornam cada vez mais lentas e com maior amplitude. O sono vai-se tornando
sincronizado e progressivamente mais profundo.
o Fase 1 (4%-5%): fase de sonolência em que se dá a transição entre a vigília e o
sono. Regista-se alguma actividade teta. Ainda há alguma actividade muscular
e movimentos oculares, as frequências respiratória e cardíaca vão diminuindo,
a temperatura corporal baixa. Não se registam fusos de sono nem complexos
K;
51
o
Fase 2 (45%-55%): fase de sono leve. Maior relaxamento muscular e menor
frequência respiratória. Predominam os fusos de sono (actividades rítmicas
entre os 12 e 14 Hz, com um aumento gradual de amplitude, seguido de uma
diminuição também gradual) e os complexos K (elementos gráficos
transitórios, geralmente bifásicos);
o Fase 3 (4%-6%): apresenta 20%-50% de actividade delta e pode, ou não, ter
fusos de sono. A respiração e a pressão arterial baixam ainda mais. É muito
semelhante ao estágio 4;
o Fase 4 (12%-15%): fase de sono profundo. Mais de 50% da actividade registada
é do tipo delta e pode apresentar fusos de sono. Fase na qual o corpo se
restaura através da libertação de hormonas reguladoras e reparadoras,
favorecendo o crescimento do corpo e o melhoramento do sistema imunitário.
Sono REM (20%-25%): é caracterizado por um EEG dessincronizado e de baixa
amplitude, pela ocorrência episódica de movimentos rápidos dos olhos (REM - Rapid
Eye Movement), por atonia muscular, pelo aumento da frequência cardíaca e
respiratória, e por erecção peniana. É nesta fase que se dão os sonhos mais vividos e
aqueles que são normalmente recordados.
Num noite de sono, passamos por ciclos de sono que se repetem 4 a 6 vezes e que têm a
duração aproximada de 90 minutos. À medida que o sono chega ao fim, a profundidade
diminui e a duração do sono REM aumenta.
52
Efeitos da electricidade
Efeitos fisiológicos da corrente eléctrica
A passagem de corrente eléctrica no organismo pode danificar diversos tecidos,
nomeadamente os tecidos nervoso, cerebral e cardíaco.
A gravidade de uma lesão por passagem de corrente eléctrica não é determinada pela
extensão/gravidade da lesão à superfície. Devido à diferente resistência oferecida pelos
diversos tipos de tecidos do nosso organismo, é possível que à superfície a lesão seja
superficial mas que tenha causado graves danos internos. Por outro lado, a uma queimadura
severa da pele pode não corresponder uma lesão interna com a mesma gravidade, pois a
resistência da pele dissipou a maior parte da energia. É também importante salientar que a
electricidade não mata por lesão cardíaca. Pode, no entanto, levar a fibrilhação ventricular e
provocar a morte do indivíduo (causa de morte mais comum).
Outros mitos que persistem em relação aos efeitos da electricidade são que a alta voltagem e
mais perigosa e que os acidentes eléctricos são raros.
A gravidade e o tipo das lesões varia com diversos factores, sendo que os mais determinantes
são a voltagem da corrente e a resistência do corpo.
Factores determinantes da gravidade das lesões:
Voltagem e tipo de corrente (AC: alterna / DC: contínua):
o Baixa voltagem (< 1000 V)
Queimaduras ausentes ou ligeiras;
AC de baixa frequência (40 a 150 HZ) é 3x mais perigosa que a DC. Lesões
frequentes (iluminação e aparelhos eléctricos);
Pode provocar morte imediata por fibrilhação ventricular, paragem do
centro respiratório, asfixia devido a contracções tetânicas dos músculos
respiratórios, etc.
o Alta voltagem (> 1000 V)
AC e DC igualmente letais;
Queimaduras graves muito frequentes.
Intensidade da corrente:
I (mA)
0,1-2
1-9
9 - 20
Perturbações prováveis
Limiar da sensação.
Sensação cada vez mais
desagradável à medida que a
intensidade aumenta.
Contracções musculares.
Sensação dolorosa,
contracções violentas,
perturbações circulatórias.
Estado após o
choque
Normal
Salvamento
Resultado Final
-----
Normal
Normal
Desnecessário
Normal
Morte
aparente
Respiração
artificial
Restabelecimento
53
20 - 100
>100
Vários
Amperes
Sensação insuportável,
contracções violentas,
asfixia, perturbações
circulatórias graves,
fibrilhação ventricular.
Asfixia imediata, fibrilhação
ventricular.
Asfixia imediata,
queimaduras graves.
Morte
aparente
Morte
aparente
Morte
aparente ou
imediata
Respiração
artificial
Restabelecimento
ou morte
Muito difícil
Morte
Praticamente
impossível
Morte
Resistência dos tecidos: a resistência das várias células e tecidos depende de diversos
factores, como a concentração iónica (quanto maior esta for menor será a resistência)
e, no caso da pele, a sua espessura (maior espessura = maior resistência), humidade
(maior humidade = menor resistência), limpeza, existência de calosidades (mais
calosidades = maior resistência), etc.
o Resistência baixa – Predominam efeitos sistémicos (coração e cérebro);
o Resistência alta – Predomina a destruição local dos tecidos e queimaduras.
Resistência: nervos < vasos < músculo < pele < gordura < osso
Trajecto da corrente e tipo de contacto: o tipo de contacto influencia o trajecto da
corrente, que por sua vez, determina a gravidade das lesões. Por exemplo, se a
corrente passa pelo coração pode causar fibrilhação ventricular e se atravessa a
cabeça pode causar convulsões, hemorragias cerebrais, paragem respiratória,
alterações psicológicas e até arritmias cardíacas.
Duração do contacto: quanto mais for o tempo de contacto mais graves serão as
lesões causadas pela passagem da corrente eléctrica.
As principais lesões causadas pela passagem de corrente eléctrica no organismo são:
Queimaduras provocadas por efeito de Joule (transformação da energia eléctrica em
calor) ou por chama (directa ou através das roupas), no local de entrada e saída da
corrente. São muito frequentes nas mãos, punhos, cotovelos e axilas;
Insuficiência renal aguda;
Lesões neurológicas imediatas frequentemente transitórias. Podem também verificarse lesões corticais e medulares como encefalopatia, hemiplegia (paralesia de uma
metade do corpo), tetraparésia, paraperésia ou afasia, e lesões dos nervos periférios
quer directamente por queimadura quer indirectamente por compressão;
Cataratas: associadas a choques de alta tensão e queimaduras na cabeça, são lesões
tardias que se manifestam, geralmente, 6 meses após a ocorrência do choque;
Lesões directas dos vasos, tromboses venosas/arteriais. Podem levar a amputações;
Fracturas por contracções tetânicas (espasmos musculares).
Morte imediata por fibrilhação ventricular, paragem do centro respiratório, asfixia
devido a contracções tetânicas dos músculos respiratórios, etc.
54
No caso particular de um indivíduo ser atingido por um relâmpago, distinguem-se 4 tipos de
choques:
“Direct Strike”: o relâmpago atinge o indivíduo directamente;
“Flashover Phenomenon”: quando o indivíduo tem o corpo e as roupas
húmidas/molhadas a passagem de corrente provoca um efeito luminoso;
“Side Flash”: o relâmpago não atinge a vítima directamente mas salta para ela depois
de atingir um outro objecto como, por exemplo, uma árvore;
“Stride potencial”: o relâmpago atinge o solo perto da vítima e atravessa-a de uma
perna para outra.
O tratamento de um indivíduo sujeito a um choque eléctrico faz-se por três fases:
No local:
o Desligar a corrente;
o Separar o doente da fonte;
o Extinguir as chamas (caso estas existam);
No transporte:
o Suporte cardio-respiratório;
o Reanimações sucessivas e agressivas (o tempo de reanimação é mais longo
que o usual uma vez que estes doentes têm tendência a recuperar bem);
o Reposição de líquidos;
No hospital:
o Acidentes com baixa voltagem: queimaduras geralmente limitadas sujeitos a
estudo de 7-10 dias da viabilidade dos tecidos;
o Acidentes com alta voltagem: estabilizar o doente, desbridar tecidos
desvitalizados e evitar infecções através de desbridamento e, se necessário,
amputações.
Aplicações da corrente eléctrica
Para além dos efeitos nocivos que a passagem de corrente eléctrica no nosso organismo
provoca, quando utilizada e/ou interpretada correctamente, ela proporciona importantes
métodos de diagnóstico e de tratamento.
O estudo dos campos eléctricos e magnéticos gerados no organismo tem aplicações em
diversas áreas da medicina. Podemos salientar os seguintes exames:
EMG: Electromiograma;
ECG: Electrocardiograma;
EEG: Electroencefalograma;
ERG: Electroretinograma;
EOG: Electroculograma;
MCG: Magnetocardiograma;
MEG: Magnetoencefalograma.
55
A aplicação directa e controlada de corrente eléctrica pode ser utilizada para fins terapêuticos.
De entre os métodos da Electroterapia podem destacar-se:
Iontoforese: utilização de corrente eléctrica com o objectivo de introduzir substâncias
com interesse terapêutico através da pele. Permite uma acção local, com recurso a
elevada concentração subcutânea e evita a via sistémica. No entanto, pode provocar
queimaduras (eléctricas ou químicas), alergia e toxicidade;
TENS (Transcutaneos Electrical Nerve Stimulation): utilização da corrente eléctrica
para provocar analgesia (alivio da dor) através da estimulação de receptores neuronais
específicos.
56
Interacção laser-tecido
Um Laser (Light Amplification by Stimulated Emission of Radiation) tem diversas propriedades
que tornam possíveis a sua aplicação nas mais variadas áreas, incluindo em medicina:
Monocromaticidade;
Direccionalidade – colimação;
Coerência espacial e temporal;
Intensidade ou brilho;
Polarização.
Quando um laser interage com tecidos pode provocar vários efeitos que dependem não só das
propriedades ópticas dos tecidos (reflexão, refracção, absorção e dispersão) mas também das
características da radiação utilizada (comprimento de onda, tempo de exposição, densidade
de potência, densidade de energia). Para obter os efeitos desejados é preciso conjugar várias
características: por exemplo, sabe-se que a água não absorve luz visível, que as proteínas têm
preferência pela radiação UV e que a hemoglobina tem dois picos de absorção
(correspondentes à oxihemoglobina e a desoxihemoglobina), logo o comprimento de onda
utilizado para trabalhar com cada uma dessas substância é diferente; É também importante
definir o conceito de densidade de potência (quantidade de energia por unidade de área e de
tempo) uma vez que é daí que advêm os diferentes efeitos da radiação laser.
Podemos destacar os seguintes efeitos da interacção laser-tecido:
Efeitos térmicos: geralmente foto-coagulação (oncologia, angiomas);
Efeitos fotoquímicos: indução de reacções químicas. Foto-terapia dinâmica (lesões
vasculares);
Efeitos fotodisruptivos: quebram algumas estruturas. Foto-disrupção (calcos renais);
Efeitos fotoablativos: tornam possível a ablação de tecidos. Foto-ablação (miopia).
Os efeitos fototérmicos representam cerca de 80% das utilizações médico-cirúrgicas dos lasers
e compreendem 3 etapas:
Etapa óptica: gera-se uma fonte de calor primária que resulta da absorção de um
fotão por parte de uma molécula. Esta aumenta a sua energia vibracional e,
consequentemente, a sua energia cinética, levando assim a um aumento da
temperatura. Esta etapa depende tanto das características ópticas do tecido como das
propriedades da radiação;
Etapa térmica: ocorre transferência de calor da área irradiada directamente (alvo
primário) para áreas adjacentes (alvos secundários). Depende das propriedades
térmicas dos tecidos, nomeadamente da condução e difusão térmicas;
57
Etapa de desnaturação térmica: o aquecimento induzido pela absorção da radiação
provoca desnaturação ou destruição tecidular. A diferentes temperaturas obtêm-se
diferentes efeitos. Os mais utilizados são:
o Hipertermia: utilizada em oncologia (laser Nd: YAG, 1064nm);
o Coagulação: utilizada no tratamento de angiomas planos;
o Volatilização: provoca a ablação instantânea de tecidos.
T (ºC)
45
50
60
70
80
100
>150
>300
Alterações histológicas
Vasodilatação, lesão endotelial, hipertermia
Redução da actividade enzimática, imobilidade celular
Desnaturação proteica e do colagénio, coagulação
Permeabilização de membranas
Contracção das fibras de colagénio, necrose de coagulação
Vaporização
Carbonização
Fusão
O efeito fotoquímico consiste na administração de uma substância fotossensibilizante (FS) que
é captada selectivamente pelo órgão/tecido alvo e activada secundariamente por radiação de
modo a produzir substâncias tóxicas e provocar a morte do tecido alvo.
No caso da Terapia FotoDinâmica (PDT) são necessários três componentes:
Fotossensibilizantes (porfirias, clorinas, ftalocianinas, etc) administrados por via
envenosa, tópica ou endocavitária;
Luz. Os efeitos fotoquímicos ocorrem a baixa densidade de potência com tempos de
acção prolongados. O comprimento de onda da radiação deve ser escolhido tendo em
conta o espectro de absorção do fotossensibilizante e os coeficientes de absorção
(probabilidade de o fotão ser absorvido) e dispersão (probabilidade de o fotão ser
desviado) do tecido;
Presença de oxigénio leva a substância fotossensibilizante a reagir com substratos
(reacção tipo I) ou directamente com o O2 (reacção tipo II).
Na fotodisrupção utilizam-se lasers Nd: YAG para produzir efeitos mecânicos, como por
exemplo, partir calcos renais ou calcos da vesícula. A densidade de potência utilizada é muito
elevada e é distribuída numa fracção de tempo extremamente curta.
Através da fotoablação podem fazer-se cortes muito precisos sem efeitos secundários no
tecido circundante (efeito térmico, mecânico). A energia da radiação utilizada quebra as
ligações das moléculas que constituem os tecidos (por ex, proteínas) e faz com que estas se
“desintegrem”.
Este efeito da radiação laser é utilizado, por exemplo, em oftalmologia para o tratamento da
miopia.
Tanto a fotodisrupção como a fotoablação induzida por plasma têm a origem na disrupção
óptica e na formação de plasma. Os acontecimentos que levam a estes dois efeitos podem ser
sumarizados:
1. O impulso laser de alta energia vai provocar a ionização de átomos do tecido alvo;
2. O excesso de energia acelera electrões livres;
58
3. Estes, por sua vez, vão provocar uma cascata electrónica, levando à ionização de
outros átomos;
4. Ao fim de algum tempo, o local irradiado é elevado para um estado da matéria
denominado plasma (estado físico da matéria que se consegue a altas temperaturas e
altas pressões, onde se perde a individualidade dos elementos químicos que o
constituem. Só restam electrões livres, uma amálgama de electrões, núcleos, protões,
etc.);
5. Como o plasma não pode existir à temperatura e pressão ambientes, desaparece do
local, provocando cortes extremamente precisos - fotoablação. É possível prever qual
a profundidade do corte, controlando a intensidade de energia;
Apenas para a fotodisrupção:
6. O plasma provoca uma onda de choque, ou seja, depois do plasma desaparecer,
origina-se um ponto de descontinuidade para onde cai a matéria circundante. Neste
momento, gera-se uma onda de choque que se propaga, não conseguindo, no
entanto, passar pelo local onde existe uma grande quantidade de energia, mas apenas
daí para trás. Ao progredir em sentido inverso, se encontrar uma superfície dura, a
onda de choque quebra-a - fotodisrupção;
Esta propriedade é aproveitada apontando-se o laser para o lado de lá de uma
determinada estrutura e gerando uma onda de choque numa direcção que a parta.
59
Ultrasons
As ondas sonoras são ondas mecânicas longitudinais que se propagam num meio elástico.
Ondas sonoras de frequência entre 20 Hz e 20 kHz (janela de frequências) têm a capacidade
de impressionar a membrana do tímpano, sendo vulgarmente chamadas sons. Ondas de
frequência inferior são infra-sons e as de frequência superior são ultra-sons (que vamos focar
neste capítulo).
O estudo do som e das suas propriedades tem importância em medicina:
Linguagem;
Audição;
Ruídos fisiológicos e patológicos;
Ultrassonoterapia: utilização de ultra-sons com objectivo terapêutico;
Ultrassonografia: utilização de ultra-sons com objectivo diagnóstico.
A produção de ultra-sons é feita com recurso a geradores piezoeléctricos que utilizam o efeito
piezoeléctrico, obtido sobretudo em determinado tipo de cerâmicas (PZT), aproveitam o valor
da frequência própria dessas estruturas e o efeito de ressonância.
O efeito piezoeléctrico consiste na deformação elástica de um material pela aplicação de um
campo eléctrico. Já o efeito piezoeléctrico inverso é caracterizado pelo aparecimento de
cargas eléctricas na superfície de um material quando este é sujeito a forças de pressão
(compressões e expansões). De entre os materiais capazes de produzir estes efeitos podemos
destacar o quartzo e algumas cerâmicas.
Relativamente à emissão dos ultra-sons, é possível dividir a zona em que se vai sentir a sua
acção em duas partes:
Zona de Fresnel: zona próxima da emissão onde a intensidade do som é superior à
esperada teoricamente;
Zona de Fraunhofer: zona mais afastada de intensidade igual à esperada e onde o som
sofre um certa dispersão. Depende sobretudo da frequência e da forma da cabeça do
emissor do ultra-som.
60
Existem outros fenómenos físicos que são importantes quando se utilizam ultra-sons:
Interferência: a ocorrência de interferência pode levar a que, em determinados locais,
a intensidade do ultra-som seja superior àquela que se está a introduzir. Pode
provocar queimaduras internas;
Cativação: quando um ultra-som se propaga num líquido que contém gases
dissolvidos, esses gases podem formar bolhas que podem implodir. É possível utilizar
este efeito, por exemplo, para esterilização – as bactérias são destruídas;
Pressão de radiação: a emissão do ultra-som dá origem a uma determinada pressão,
que depende da sua intensidade. Pode utilizar-se este fenómeno para medir a
intensidade da radiação ou para introduzir substâncias na pele sob a forma de creme
ou gel.
A ultrasonoterapia (terapia com ultra-sons) faz uso dos efeitos biofísicos dos ultra-sons:
Efeitos térmicos: a emissão de um ultra-som de forma contínua produz calor por
dissipação. A profundidade a que a temperatura sofre maiores alterações depende da
frequência da radiação: por exemplo, uma sonda de 3 MHz é menos penetrante e a
produção de calor dá-se mais à superfície. Já uma sonda de 1Mhz provoca o
aquecimento de estruturas mais profundas;
Efeitos mecânicos: emitem-se ultra-sons de forma pulsátil;
Efeitos biológicos: resultam da combinação dos efeitos térmicos e mecânicos.
A ecografia é a técnica de ultrassonografia (diagnóstico com ultra-sons) mais utilizada e
baseia-se na interpretação do som reflectido (eco) nas descontinuidades dos tecidos quando
estes são atingidos por uma onda sonora, para produzir imagens.
Esta técnica tem inúmeras vantagens:
Não invasiva;
Inócua;
Baixo custo.
A ecografia tem diversas aplicações:
Visualização de órgãos e massas em várias zonas:
o Pescoço (tiroideia, paratiroideias);
o Abdómen (fígado, vesícula, pâncreas, baço);
o Aparelho urinário (rins, bexiga e próstata);
o Aparelho cardiovascular (coração, artérias, veias);
o Partes moles (músculos, tendões);
É muito utilizada em ginecologia e obstetrícia;
Só não pode ser aplicada nos pulmões, devido à quantidade de gás presente nesses
órgãos, e nos ossos, estes reflectem totalmente as ondas e não permitem a
visualização em profundidade.
61
Possibilita diferentes métodos de representação:
Modo A: modo de amplitude. Representa a medição da distância a que o ultra-som é
emitido e reflectido e permite representar, num sistema de coordenadas (x,y), a
profundidade do sinal a que o ultra-som foi reflectido. Possibilita medições com
precisão e representação bidimensional. É o modo mais antigo;
Modo B: modo de brilho. Representa também um sistema de coordenadas espaciais e
permite medir a distância resultante do varrimento que se faz a uma estrutura. Já
permite obter imagens (estáticas) de tipo topográfico e ter uma imagem
bidimensional. Por exemplo, relativamente a órgãos, permite ter uma imagem do seu
contorno. É mais avançado que o modo A;
Modo M: modo de movimento. Tecnologicamente mais complexo, é obtido por
processos mecânicos ou electrónicos (uma sonda de ultra-sons com vários cristais
emite ondas com várias frequências). Permite fazer um varrimento, representando a
profundidade em função do tempo, e registar os movimentos de estruturas. Utiliza-se,
por exemplo, para visualizar ecograficamente as válvulas do coração a funcionar, temse uma ideia do movimento em tempo real (ecografias mais sofisticadas);
Dopller: utiliza feixes contínuos e baseia-se no efeito de doppler (a altura do som
depende não só da frequência mas da velocidade relativa entre o emissor e receptor).
Permite medir débitos, como a velocidade de circulação do sangue numa artéria.
62
Tópicos essenciais
Transporte através de membranas
Osmose: movimento de moléculas de água, através de uma membrana semipermeável, de um
meio onde a concentração de soluto é menor para um meio onde essa concentração é maior,
ou seja, contra o gradiente de concentração.
Difusão simples: movimento de partículas a favor do seu gradiente de concentração sem
intervenção de proteínas transportadoras - dá-se directamente através dos interstícios da
membrana lipídica, para moléculas lipossolúveis; ou através de poros (ou canais iónicos) e
canais proteicos, para substâncias hidrossolúveis. A taxa de difusão é directamente
proporcional à solubilidade das partículas.
Difusão facilitada: transporte mediado passivo em que proteínas transportadoras têm
afinidade química com algumas moléculas, o que permite a sua ligação. Só possibilita o
movimento de substâncias a favor do seu gradiente de concentração.
Transporte activo: movimento de substâncias contra o seu gradiente de concentração. Requer
o uso de energia por parte das proteínas transportadoras:
Transporte activo primário: A energia utilizada provém de reacções exergónicas, como
a hidrólise de ATP ou de outros metabolismos energéticos. Ex: bomba Na +/K+;
Transporte activo secundário: A energia utilizada é fornecida, indirectamente, por
gradientes iónicos. A passagem de uma substância a favor do seu gradiente de
concentração permite o movimento de outra substância contra o gradiente:
o Co-Transporte: São transportadas duas moléculas no mesmo sentido. Ex: NaGlucose, Na-AA (aminoácidos);
o Contra-Transporte: As substâncias são transportadas em sentidos diferentes.
Ex: Na-Ca, Na-H.
Endocitose: Absorção de partículas por parte da célula envolvendo-as na sua membrana e
dando origem a vesículas no meio intracelular.
Pinocitose: ingestão de solutos ou partículas singulares;
Fagocitoce: ingestão de partículas de grandes dimensões, tais como bactérias, vírus ou
células.
Exocitose: Expulsão de substâncias que se encontram em vesículas no interior da células, por
fusão com a membrana plasmática.
63
Potenciais bioeléctricos
Equação de Nernst: EMF
60 X
log
z
X
ext
int
mV
Se a diferença de potencial medida através da membrana for igual à calculada pela
equação de Nernst, está-se num estado de equilíbrio, em que não há movimento
iónico apesar de se verificar um gradiente de concentração;
Se a diferença de potencial medida for do mesmo sinal mas superior à calculada, a
força eléctrica é superior à das concentrações e portanto determina a direcção dos
iões;
Se a diferença de potencial medida for do mesmo sinal mas inferior à calculada, a força
de diferença de concentrações é superior à eléctrica determinando a direcção dos
iões, que se movem de acordo com o seu gradiente de concentração;
Se a diferença de potencial medida for de sinal contrário à calculada, ambas as forças
têm a mesma direcção, não podendo haver equilíbrio. As duas forças vão puxar o ião
exactamente para o mesmo lado da membrana e portanto vai haver um desequilíbrio
iónico franco.
Equação de Goldman: tem em conta a permeabilidade da membrana a diferentes iões.
P( Na ) Na
EMF 60 log
P( Na ) Na
P( K ) K
ext P( K ) K
int
P(Cl ) Cl
ext P(Cl ) Cl
int
ext
int
mV
Potencial de repouso: potencial da membrana de células/fibras nervosas quando estas não
estão a transmitir qualquer tipo de sinais nervosos e se encontram em equilíbrio
electroquímico. Na maioria das células, este potencial é negativo e tem o valor aproximado de
-90mV.
Depende dos canais de sódio e potássio, sendo o potencial de difusão de K + o que mais
contribui (-94mV). Com a acção dos canais de Na+ (+61mV) atinge-se -86mV (a condutância do
sódio é 100x menor que a do potássio). A bomba de sódio potássio contribui com -4mV. Os
aniões intracelulares podem contribuir com até -10mV.
Potencial de repouso
Neurónio
Músculo esquelético
Músculo liso
Tecido condutor cardíaco
-70mV
-90mV
-60mV
-90mV
Potencial de acção: mudança rápida do potencial da membrana. Tem 3 fases:
Repouso: antecede o potencial de acção. Potencial de repouso;
Despolarização: aumento do potencial no interior da célula. Overshooting - o
potencial atinge valores positivos. Deve-se à abertura dos canais de sódio;
64
Repolarização: reposição do potencial de repouso. Hiperpolarização - potencial da
membrana é inferior ao potencial de repouso. Deve-se ao fecho dos canais de sódio e
abertura dos canais de potássio.
Os canais de sódio e potássio que intervêm no potencial de acção são controlados por
voltagem.
Limiar de excitabilidade (limiar de acção ou threshold): o valor mínimo necessário para gerar
um potencial de acção, que é cerca de -65mV para a maioria das células nervosas. Caso esse
limiar não seja atingido não há propagação de um potencial. Depois de atingido o limiar a
resposta é independente do estímulo – Lei do tudo ou nada.
Estímulo não eficaz: gera uma resposta local mas não um potencial de acção;
o Estímulo sublimiar: não é suficiente para atingir o limiar de excitabilidade;
Estímulo eficaz: gera um potencial de acção;
o Estímulo limiar: é suficiente para atingir o limiar de excitabilidade;
o Estímulo supra-limiar: ultrapassa o limiar de excitabilidade.
Período refractário absoluto: período em que, qualquer que seja o estímulo, não é possível
gerar um novo potencial de acção. Maioria dos canais de Na + encontra-se inactivada por
voltagem.
Período refractário relativo: período em que é necessário um estímulo maior que o inicial para
gerar um potencial de acção. Alguns canais de Na+ ainda estão inactivados e a condutância ao
K+ ainda está aumentada.
Acomodação: a despolarização é lenta e permite que seja ultrapassado o limiar de
excitabilidade sem que seja gerado um novo potencial de acção.
Condução no nervo
Condução electrotónica: condução a curtas distâncias. O potencial decresce ao longo do
tempo e propaga-se mais eficazmente em fibras revestidas por mielina.
Condução regenerativa: utilizada para grandes distâncias e para fibras não mielinizadas. Um
potencial gera correntes locais que estimulam as áreas adjacentes até ao limiar e regeneram o
potencial de acção, propagando-o sucessivamente ao longo de toda a fibra.
Condução saltatória: dá-se em fibra mielinizadas. O potencial é propagado de nodo de Ranvier
em nodo Ranvier. Quando se inicia um potencial de acção num nodo criam-se correntes nas
regiões próximas do fluido extracelular e também no axoplasma (citoplasma dos axónios) que
dão inicio a um novo potencial de acção no nodo mais próximo. Este modo de condução é
mais rápido e permite poupar energia.
65
A velocidade de propagação dos potenciais nas fibras nervosas depende de:
Diâmetro: maior diâmetro corresponde a maior velocidade;
Revestimento de mielina: uma fibra mielinizada chega a conduzir mais rapidamente
do que uma fibra não-mielinizada com um diâmetro 100x maior.
Sinapse
Sinapse é uma zona específica de contacto/interacção intercelular, morfologicamente
específico, que faz a comunicação entre um nervo e outro nervo, músculo ou glândula.
Distância
Continuidade citoplasmática
Ultra-estrutura
Sinapse Eléctrica
3,5 nm
Sim
Gap-Junction
Agente Transmissor
Direcção
Atraso sináptico
Corrente iónica
Bidireccional
Ausente
Sinapse Química
20 - 40 nm
Não
Vesículas pré-sinápticas
receptores pós-sinápticos
Neurotransmissores
Unidireccional
0,3 – 5 ms
e
Sinapse eléctrica: o potencial propaga-se directamente por correntes iónicas através de gapjunctions.
Sinapse química: a comunicação dá-se através de neurotransmissores. Um potencial na célula
pré-sináptica provoca exocitose de vesículas com transmissores que são libertados para a
fenda sináptica e captados por receptores pós-sinápticos.
Nas sinapses químicas podem ocorrer dois tipos de resposta:
Resposta inibitória: há uma hiperpolarização da célula devido à abertura dos canais de
cloro, ao aumento da condutância do potássio ou à activação de receptores
enzimáticos que aumentam o número de receptores inibitórios e diminuem o número
de receptores excitatórios;
Resposta excitatória: é gerado um potencial por abertura dos canais de sódio, há
diminuição da condutância do cloro e potássio, aumento do número de receptores
excitatórios da sua membrana ou diminuição da quantidade de receptores inibitórios.
Nas sinapses químicas podem distinguir-se receptores que actuam de dois modos distintos:
Receptores ionotrópicos: actuam ao nível dos canais iónicos, promovendo a sua
abertura ou fecho e influenciando o movimento de iões;
Receptores metabotrópicos: entram no sistema de mensageiros secundários,
provocam alterações metabólicas.
66
De acordo com a zona onde ocorre a ligação sináptica, as sinapses químicas podem classificarse em:
Axo-axonal: ligação axónio-axónio;
Axo-somática: ligação axónio-corpo celular;
Axo-dendrítica: ligação axónio-dendrite.
Músculo Esquelético
Músculo esquelético: liga os ossos e articulações, permite o seu movimento e a produção de
força, oferece protecção e resistência. É o tecido mais abundante no organismo humano e tem
actividade voluntária. A sua unidade estrutural é a fibra muscular e a unidade funcional é o
sarcómero.
músculo fibras musculares miofibrilhas (actina e miosina) sarcómero
A contracção do músculo é provocada pela acção de uma série de proteínas específicas, as
proteínas contrácteis:
Miosina: possui cabeças globulares e forma filamentos espessos;
Actina: enroladas em dupla hélice, forma filamentos finos;
Tropomiosina: encontra-se entre as hélices de actina e bloqueia os sítios activos a que
se liga a miosina durante a contracção;
Troponinas: aderentes à tropomiosina e espaçadas entre si, fazem a ligação entre a
actina e a tropomiosina.
Mecanismo de contracção:
1. Potencial de acção do nervo chega à junção neuromuscular;
2. Provoca um aumento da permeabilidade ao cálcio;
3. Junção do cálcio a uma vesícula e abertura na fenda sináptica, com libertação de
acetilcolina;
4. Acetilcolina une-se ao receptor pós-sináptico, que é um receptor próprio para a
acetilcolina (tipo nicotínico), abrindo canais de sódio;
5. Há despolarização da membrana e propagação do potencial de acção ao longo de toda
a membrana da fibra (túbulos-T);
6. A despolarização da membrana leva à abertura dos retículos sarcoplasmáticos e à
libertação de cálcio;
7. O cálcio interage com Troponina-C e elimina o efeito inibidor da tropomiosina sobre a
interacção actina/miosina;
8. Filamentos de actina deslizam ao longo da miosina e dá-se o encurtamento do
sarcómero;
9. O cálcio reentra nos retículos e a contracção cessa.
Contracção isométrica ou estática: não há encurtamento do músculo mas há desenvolvimento
de tensão;
67
Contracção isotónica ou dinâmica: o músculo está preso apenas numa extremidade e encurta
após ser estimulado.
Abalo muscular: contracção rápida de um músculo em resposta a um estímulo único. Um
abalo compreende várias fases:
Período de latência;
Contracção;
Velocidade máxima de contracção;
Fase de relaxamento.
O aumento da frequência de estimulação dos músculo pode levar a:
Tétano imperfeito: há fusão parcial das respostas. A força aumenta com a frequência;
Tétano perfeito: fusão completa das respostas. Atinge-se a força máxima, que não
aumenta mais;
Tétano fadiga: o músculo é incapaz de contrair.
A força de contracção depende do comprimento do sarcómero e do músculo como um todo.
A velocidade de contracção diminui com o aumento da carga.
%
Resistência à fadiga
Velocidade de Fadiga
Velocidade de
contracção
Actividade miosinaATPásica
Fonte ATP
Activ. Enzimas
Glicolíticas
Nº mitocôndrias
Capilares
[Mioglobina]
Cor do músculo
[Glicogénio]
Diâmetro
Fibras Tipo I
(Lentas oxidativas)
50-55%
Resistentes à fadiga
Lenta
Lenta
Fibras Tipo IIa
(Rápidas oxidativas)
30-35%
Resistentes à fadiga
Média
Rápida
Fibras Tipo IIb
(Rápidas glicolíticas)
10-20%
Fatigáveis (10/20%)
Rápida
Rápida
Baixa
Alta
Alta
Fosforilação
oxidativa
Baixa
Fosforilação
oxidativa
Média
Glicólise anaeróbica
Muitas
Muitos
Alta
Vermelho
Baixa
Pequeno
Muitas
Muitos
Alta
Vermelho
Média
Médio
Poucas
Poucos
Baixa
Branco
Alta
Grande
Alta
EMG e ENMG – Electromiografia e Electroneuromiografia
Dermátomo: área cutânea inervada por fibras sensitivas de uma mesma raiz nervosa.
Miótomo: grupo de músculos inervados primariamente pelas fibras motoras de uma única raiz
nervosa.
68
Potencial de unidade motora (PUM): soma de todos os potenciais de fibras individuais de
unidade/s motora/s.
Electromiografia (EMG): monitoriza a actividade eléctrica dos músculos.
Electroneuromiografia (ENMG) permite medir a actividade neuromuscular (tempo de
condução, tipo de potencial) através da estimulação eléctrica de nervos e captação
monitorização dos potenciais propagados.
Neuropatia: invervação colateral.
Miopatia: perda de fibras musculares.
Desmielinização: um estímulo demora mais tempo a percorrer a mesma distância mas não
apresenta alteração do potencial.
Degenerescência axonal: o tempo de transmissão é o normal mas o potencial é mais fraco do
que o esperado.
O ENMG tem aplicações na detecção de diversas patologias, tais como:
Síndrome do túnel cárpico;
Neuropatia cubital do cotovelo;
Polineuropatia;
Radiculopatia;
Miopatia.
O ENMG permite:
Localizar topograficamente lesões;
Diferenciar entre compromisso neuropático desmielinizante (um estímulo demora
mais tempo a percorrer a mesma distância mas não apresenta alteração do potencial)
e axonal (o tempo de transmissão é o normal mas o potencial é mais fraco do que o
esperado);
Caracterizar a gravidade das lesões;
Determinar um prognóstico.
Músculo Liso
Músculo liso multi-unitário: formado por fibras individuais que se contraem de forma
independente, maioritariamente por acção de sinais nervosos, e que são inervadas por uma
terminação nervosa simples. Cada fibra está envolta por tecido conjuntivo rico em colagénio e
glicoproteínas que ajudam ao seu isolamento.
Músculo liso unitário (visceral ou sincicial): as fibras contraem-se em conjunto, como um todo,
e as suas membranas estão ligadas em vários pontos por gap-junctions, o que permite a
transmissão de impulsos (com origem em vários tipos de estímulos) entre fibras.
69
Bases químicas do processo contráctil:
Características (químicas) das fibras de actina e miosina idênticas às do músculo
esquelético;
Não têm complexo de troponina (têm calmodulina);
A contracção também é activa por iões cálcio (mas por um processo diferente);
A energia necessária é fornecida pelo ATP.
Bases físicas do processo contráctil:
Fibras de actina ligadas a corpos densos;
Alguns corpos densos associados à membrana celular ou dispersos na célula;
Corpos ligados entre si por proteínas estruturais e ligados a corpos de outras células
por pontes proteicas intercelulares;
Filamentos de miosina escassos e intercalados em filamentos de miosina.
Diferenças entre o processo de contracção do músculo liso em relação ao músculo esquelético:
Ciclo de ligação entre pontes cruzadas lento: menor actividade da ATPase;
Menor requerimento de energia;
Inicio de contracção e relaxamento lento: varia de órgão para órgão;
Maior força de contracção: devido ao maior período de ligação actina-miosina;
Maior percentagem de encurtamento;
“Latch Mechanism”: apenas sob a acção de pequenos e contínuos estímulos nervosos
ou hormonais, o ML é capaz de manter contracções prolongadas (por vezes durante
horas) com um gasto de energia muito reduzido;
Stress-relaxamento e reverso stress-relaxamento: permite que um órgão oco
mantenha a pressão no seu lúmen constante face a alterações de volume, excepto por
curtos períodos de tempo;
Regulação pelos iões cálcio: o início da contracção é modulado pela calmodulina, em
vez da troponina, e ocorre em três etapas:
4. 4 iões cálcio ligam-se à calmodulina, formando o complexo cálciocalmodulina, um complexo fosforilativo (só ligada a 4 iões cálcio a calmodulina
se torna eficaz);
5. O complexo cálcio-calmodulina activa a miosina-quinase e liga-se ao
caldesmon (tem um papel semelhante ao da tropomiosina), retirando-o do
contacto com a actina;
6. A miosina-quinase fosforila uma das cadeias leves de cada cabeça de miosina,
permitindo a ligação desta ao filamento de actina. Ocorre depois o
deslizamento entre os filamentos de actina e miosina, levando à contracção
do músculo.
Quando a concentração de iões cálcio baixa até um nível crítico o processo é invertido,
passando a miosina-fosfatase a desfosforalizar a cadeia leve da cabeça de miosina,
que se separa do filamento de actina e facilita o relaxamento do músculo;
Fontes dos iões cálcio: retículo sarcoplasmático rudimentar, algumas células têm
clavéolas adjacentes ao RS; a entrada de cálcio extracelular é a maior responsável pela
contracção muscular; a força de contracção depende da concentração de cálcio
70
extracelular; bombas de cálcio têm um papel importante no relaxamento do músculo
(relaxamento lento);
Estímulos de contracção: a contracção do músculo liso pode ser estimulada por
diferentes factores: estímulo nervoso (eléctrico ou químico), estiramento, hormonas,
efeitos tecidulares locais.
Potencial em ponta (tipo “spkie”): potencial de acção típico, como o que ocorre no músculo
esquelético. Tem uma duração de 10ms a 50ms;
Potencial em plateu: a fase de despolarização é semelhante à do potencial em ponta mas a
repolarização é longa (pode durar até 1s), mantendo o músculo contraído durante mais
tempo. Deve-se principalmente à acção dos canais de cálcio (mais abundantes que os canais
de sódio);
Potencial de ondas lentas (ondas de pace-maker): localmente, as fibras musculares podem
excitar-se espontaneamente com determinado ritmo (por razões ainda desconhecidas). Se a
despolarização resultante for suficiente para atingir o limiar então gera-se um potencial de
acção.
Despolarização sem potencial de acção: ocorre principalmente no músculo liso multi-unitário.
Dá-se uma despolarização local que é transmitida electrotonicamente.
Músculo Cardíaco
Músculo cardíaco: músculo estriado que faz a contracção do coração. As fibras musculares
comunicam entre si através de discos intercalares que permitem o movimento livre de iões e a
transmissão do potencial de acção. Sincício auricular, sincício ventricular, tecido cardionector.
Principais características:
Automatismo: capacidade de gerar estímulos que podem originar contracção;
Dromotropismo (condutibilidade): poder de condução do estímulo pelas fibras de
modo a que o estímulo chegue a todo o sincício;
Batmotropismo (excitabilidade): capacidade de reagir quando estimulado;
Ionotropismo (contractilidade): pode ser negativa (diminui força de contracção) ou
positiva (aumenta);
Tonacidade.
Potencial de repouso:
Fibras musculares: -85mV a -95mV;
Fibras de Purkinje: -90mV a -100mV.
Potencial de acção: varia com o local onde tem origem (nodo SA, AV, miocárdio, etc).
Longa duração: são 3 a 15 vezes mais longos que os do músculo esquelético;
Plateu (planalto): após o pico inicial, o potencial da membrana mantém-se elevado
durante um período de tempo relativamente longo ao qual se segue uma rápida
71
repolarização. A abertura dos canais rápidos de sódio leva ao pico inicial e o plateu é
mantido pela acção dos canais lentos de cálcio e pela baixa permeabilidade da
membrana ao potássio;
Período refractário: devido à existência de plateu, o período refractário absoluto é
mais longo. Músculo ventricular: absoluto – 0,25s a 0,3s; relativo – 0,05s. Músculo
auricular: absoluto – 0,15s; relativo – 0,03s;
Velocidade de condução: nas fibras musculares a velocidade de condução dos
impulsos é de cerca de 0,3 a 0,5m/s, 1/10 da das fibras musculares esqueléticas. No
tecido cardionector essa velocidade é de 0,02 a 4 m/s.
Mecanismo de contracção:
Propagação do potencial;
Abertura de canais rápidos de sódio e de canais lentos de cálcio. Baixa condutância ao
potássio;
Libertação de Ca2+ do RS e dos túbulos T (volume 25x maior que os do ME);
Ligação do cálcio à troponina C e contracção muscular;
Após o plateu, rápida repolarização. Fecho dos canais de cálcio, aumento da
condutância do sódio e actividade de fosfolambam (transporta activamente Ca2+ para
o interior dos RS).
ECG – Electrocardiograma
Electrocardiograma (ECG): registo gráfico dos potenciais eléctricos gerados pelo coração
(despolarização e repolarização do miocárdio), detectados por eléctrodos metálicos colocados
nas extremidades e na parede torácica.
Vantagens:
Não invasiso;
Simples;
Versátil,
Reprodutível, económico;
Detecta múltiplas patologias;
Colheita e interpretação simples.
Pode ser muito útil para conhecer:
A orientação anatómica do coração;
O tamanho relativo das diversas câmaras cardíacas;
Uma variedade de alterações do ritmo e condução;
A extensão, localização e progressão de lesões isquémicas do miocárdio;
Os efeitos de alterações de concentrações de electrólitos;
A influência de determinados fármacos.
72
Dipolo: vector com amplitude, drecção e sentido definidos, gerado por uma carga positiva e
outra negativa separadas por uma curta distância.
Deflexão positiva: o dipolo é representando por um vector que se aproxima da posição de
registo.
Deflexão negativa: o dipolo afasta-se da posição de registo.
Uma curva de ECG apresenta diversas ondas e intervalos a que correspondem a etapas
diferentes do ciclo cardíaco:
Onda P: activação do nódulo sinusal e despolarização das aurículas (a direita
despolariza primeiro que a esquerda);
Intervalo PQ ou PR: activação completa das aurículas e atraso no nodo AV. Linha
isoeléctrica;
Onda Q: activação do nódulo AV e repolarização das aurículas;
Onda R: despolarização do miocárdio interno;
Onda S: despolarização do miocárdio externo;
Complexo QRS: despolarização ventricular;
Segmento ST: activação completa do ventrículo. Linha isoeléctrica;
Onda T: repolarização dos ventrículos. Esta onda é positiva porque a repolarização
começa no exterior, seguindo de fora para dentro (ao contrário da despolarização);
Onda U: potenciais tardios do inicio da diástole. Só está presente nalguns ECG,
sobretudo quando as frequências são mais baixas em determinadas derivações;
Intervalo QT: despolarização e repolarização dos ventrículos.
Derivações biploares dos membros (clássicas): registam os potenciais eléctricos no plano
frontal e representam uma diferença de potencial entre dois locais seleccionados.
DI: diferença de potencial entre o braço esquerdo e o braço direito (VLA - VRA);
DII: diferença de potencial entre a perna esquerda e o braço direito (VLL - VRA);
DIII: diferença de potencial entre a perna esquerda e o braço esquerdo (VLL - VLA).
Triângulo de Einthoven: o coração está localizado no centro e os vértices do triângulo são
indicados pelas 3 derivações.
lei de Einthoven: se o potencial eléctrico de duas quaisquer derivações bipolares for conhecido
num dado instante, a terceira pode ser calculada pela fórmula: DII = DI + DIII.
Derivações unipolares (dos membros e pré-cordiais): medem a diferença de potencial entre
um eléctrodo indiferente e um eléctrodo explorador.
Eléctrodo indiferente: formado por três fios eléctricos que estão ligados em série com
resistências de 5M e ligados entre si a um terminal central. As extremidades livres
destes fios ligam-se aos eléctrodos do braço esquerdo (LA), braço direito (RA) e perna
esquerda (LL). Considera-se que a soma dos três potenciais LA+RA+LL é igual a zero, ou
seja, o potencial do eléctrodo indiferente é zero;
Terminal central: liga-se ao pólo negativo do electrocardiógrafo;
73
Eléctrodo explorador: liga-se ao pólo positivo. Por princípio, as derivações unipolares
tentam medir potenciais locais e não diferenças de potencial.
Derivações unipolares aumentadas dos membros: retira-se do terminal central o eléctrodo do
membro a ser explorado.
aVR: potencial no braço direito;
aVL: potencial no braço esquerdo;
aVF: potencial na perna esquerda.
Derivações unipolares pré-cordiais: registam potenciais no plano horizontal. O eléctrodo
indiferente permanece ligado às três extremidades, enquanto o eléctrodo explorador varia de
posição ao longo da parede torácica.
V1: quarto espaço intercostal direito junto ao esterno;
V2: quarto espaço intercostal esquerdo junto ao esterno;
V3: equidistante de V2 e V4;
V4: quinto espaço intercostal esquerdo na linha médio-clavicular;
(todos os eléctrodos seguintes são registados no mesmo plano horizontal de V4)
V5: linha axilar anterior;
V6: linha axilar média.
Eixo eléctrico médio do complexo QRS: vector com origem no centro do triângulo de
Einthoven e que indica a direcção preponderante do fluxo de corrente durante a
despolarização dos ventrículos. Calculado a partir da amplitude total (subtraindo os potenciais
negativos aos positivos) dos complexos QRS nas derivações bipolares dos membros.
Os valores do eixo eléctrico médio, em indivíduos adultos, variam entre -30º e 110º. Valores
diferentes são reconhecidos como desvios do eixo:
Desvio esquerdo do eixo: Valores entre -30º e -90º (DI positivo e DII negativo);
Desvio direito do eixo: Valores entre +110º e +180º;
Desvio extremo do eixo: Valores entre -90º e +180º (DI e DII negativos).
Sono e EEG – Electroencefalograma
Ciclo sono-vigília: ciclo, com periodicidade de 24 horas, durante o qual o organismo altera
entre um estado de sono e de vigília.
Sono: estado de alteração de consciência reversível, que ocorre de forma cíclica e que é
imprescindível à vida.
Critérios comportamentais:
Diminuição da actividade motora;
Diminuição da resposta à estimulação externa;
Posturas estereotipadas, como estar deitado de olhos fechados;
Relativa facilidade de reversibilidade (ao contrário do coma).
74
Funções:
Restauração do balanço natural dos centros nervosos;
Conservação de energia;
Cognição;
Termorregulação;
Maturação neuronal e saúde mental.
Os estudos poligráficos do sono podem ser feitos recorrendo a diferentes técnicas e
parâmetros:
EOG;
EMG submentoniano e tibial;
EEG;
ECG;
Fluxo nasal, movimentos torácicos e abdominais;
Saturação de O2.
Electroencefalograma (EEG): exame complementar de diagnóstico que, por intermédio da
colocação de vários eléctrodos segundo um sistema internacional e das derivações entre esses
eléctrodos, permite registar a actividade eléctrica cerebral.
Depende de:
Distância entre o gerador e o eléctrodo;
Orientação dos geradores;
Número e sincronização dos geradores.
Tipos de actividade registada:
Actividade Alfa : ondas de baixa amplitude e frequência entre 8Hz e 13Hz. São
registadas quando um indivíduo se encontra em descanso, deitado e de olhos fechado,
mas acordado;
Actividade Beta : ondas de baixa amplitude e frequência maior que 13Hz
(geralmente entre 14Hz e 80Hz). Verificam-se quando um indivíduo está acordado, de
olhos abertos e num estado normal de actividade;
Actividade Teta : ondas de baixa frequência, entre 4Hz e 7Hz. Registam-se numa fase
de sonolência;
Actividade Delta : ondas alta amplitude e muito baixa frequência, entre 05Hz e 4Hz.
São registadas em fases de sono profundo ou coma, ou em zonas localizadas que
apresentem lesões cerebrais.
Fases do sono:
Sono Não-REM: ondas cada vez mais lentas e com maior amplitude. O sono vai-se
tornando sincronizado e progressivamente mais profundo;
o Fase 1 (4%-5%): fase de sonolência em que se dá a transição entre a vigília e o
sono. Regista-se alguma actividade teta. Ainda há alguma actividade muscular
e movimentos oculares, as frequências respiratória e cardíaca vão diminuindo,
75
a temperatura corporal baixa. Não se registam fusos de sono nem complexos
K;
o Fase 2 (45%-55%): fase de sono leve. Maior relaxamento muscular e menor
frequência respiratória. Predominam os fusos de sono e os complexos K;
o Fase 3 (4%-6%): apresenta 20%-50% de actividade delta e pode, ou não, ter
fusos de sono. A respiração e a pressão arterial baixam ainda mais. É muito
semelhante ao estágio 4;
o Fase 4 (12%-15%): fase de sono profundo. Mais de 50% da actividade registada
é do tipo delta e pode apresentar fusos de sono. Fase na qual o corpo se
restaura através da libertação de hormonas reguladoras e reparadoras,
favorecendo o crescimento do corpo e o melhoramento do sistema imunitário;
Sono REM (20%-25%): é caracterizado por um EEG dessincronizado e de baixa
amplitude, pela ocorrência episódica de movimentos rápidos dos olhos (REM - Rapid
Eye Movement), por atonia muscular, pelo aumento da frequência cardíaca e
respiratória, e por erecção peniana. É nesta fase que se dão os sonhos mais vividos e
aqueles que são normalmente recordados.
Ciclos de sono: repetem-se 4 a 6 vezes e têm a duração aproximada de 90 minutos. À medida
que o sono chega ao fim, a profundidade diminui e a duração do sono REM aumenta.
Efeitos da electricidade
Mitos:
A electricidade mata por lesão cardíaca (mata por fibrilhação ventricular);
A alta voltagem é mais perigosa (depende de diversos factores);
A extensão da lesão superficial determina a gravidade da lesão (pouca queimadura
superficial não significa que os tecidos internos não tenham sido lesados);
Os acidentes eléctricos são raros (são até bastante frequentes).
A resistência das várias células e tecidos depende de diversos factores, como a concentração
iónica (quanto maior esta for menor será a resistência) e, no caso da pele, a sua espessura
(maior espessura = maior resistência), humidade (maior humidade = menor resistência),
limpeza, existência de calosidades (mais calosidades = maior resistência), etc.
o Resistência baixa – Predominam efeitos sistémicos (coração e cérebro);
o Resistência alta – Predomina a destruição local dos tecidos e queimaduras.
Resistência: nervos < vasos < músculo < pele < gordura < osso
As principais lesões causadas pela passagem de corrente eléctrica no organismo são:
Queimaduras provocadas por efeito de Joule (transformação da energia eléctrica em
calor) ou por chama (directa ou através das roupas), no local de entrada e saída da
corrente. São muito frequentes nas mãos, punhos, cotovelos e axilas;
Insuficiência renal aguda;
76
Lesões neurológicas imediatas frequentemente transitórias. Podem também verificarse lesões corticais e medulares como encefalopatia, hemiplegia (paralesia de uma
metade do corpo), tetraparésia, paraperésia ou afasia, e lesões dos nervos periférios
quer directamente por queimadura quer indirectamente por compressão;
Cataratas: associadas a choques de alta tensão e queimaduras na cabeça, são lesões
tardias que se manifestam, geralmente, 6 meses após a ocorrência do choque;
Lesões directas dos vasos, tromboses venosas/arteriais. Podem levar a amputações;
Fracturas por contracções tetânicas (espasmos musculares).
Morte imediata por fibrilhação ventricular, paragem do centro respiratório, asfixia
devido a contracções tetânicas dos músculos respiratórios, etc
O estudo dos campos eléctricos e magnéticos gerados no organismo tem aplicações em
diversas áreas da medicina. Podemos salientar os seguintes exames:
EMG: Electromiograma;
ECG: Electrocardiograma;
EEG: Electroencefalograma;
ERG: Electroretinograma;
EOG: Electroculograma;
MCG: Magnetocardiograma;
MEG: Magnetoencefalograma.
A aplicação directa e controlada de corrente eléctrica pode ser utilizada para fins terapêuticos.
De entre os métodos da Electroterapia pode destacar-se:
Iontoforese: utilização de corrente eléctrica com o objectivo de introduzir substâncias
com interesse terapêutico através da pele;
TENS (Transcutaneos Electrical Nerve Stimulation): utilização da corrente eléctrica
para provocar analgesia (alivio da dor) através da estimulação de receptores neuronais
específicos.
Interacção Laser-Tecido
Laser: Light Amplification by Stimulated Emission of Radiation.
Características:
Monocromaticidade;
Direccionalidade – colimação;
Coerência espacial e temporal;
Intensidade ou brilho;
Polarização.
Propriedades ópticas dos tecidos:
Reflexão;
Refracção;
77
Absorção;
Dispersão.
Propriedades de radiação:
Comprimento de onda;
Tempo de exposição;
Densidade de potência;
Densidade de energia.
Efeitos da interacção lase-tecido:
Efeitos fototérmicos: foto-coagulação (oncologia, angiomas).
o Etapa óptica: gera-se uma fonte de calor primária que resulta da absorção de
um fotão por parte de uma molécula. Esta aumenta a sua energia vibracional
e, consequentemente, a sua energia cinética, levando assim a um aumento da
temperatura;
o Etapa térmica: ocorre transferência de calor da área irradiada directamente
(alvo primário) para áreas adjacentes (alvos secundários);
o Etapa de desnaturação térmica: o aquecimento induzido pela absorção da
radiação provoca desnaturação ou destruição tecidular.
Efeitos fotoquímicos: foto-terapia dinâmica (lesões vasculares). Administração de uma
substância fotossensibilizante (FS) que é captada selectivamente pelo órgão/tecido
alvo e activada secundariamente por radiação de modo a produzir substâncias tóxicas
e provocar a morte do tecido alvo;
Efeitos fotodisruptivos: foto-disrupção (calcos renais). Onde de choque produz efeito
mecânico;
Efeitos fotoablativos: foto-ablação (miopia). Cortes muito precisos sem efeitos
térmicos ou mecânicos no tecido circundante.
Ultra-sons
Importância em medicina:
Linguagem;
Audição;
Ruídos fisiológicos e patológicos;
Ultrassonoterapia: utilização de ultra-sons com objectivo terapêutico;
Ultrassonografia: utilização de ultra-sons com objectivo diagnóstico.
Geradores piezoeléctricos: produzem ultra-sons.
Efeito piezoeléctrico: deformação elástica de um material (pode produzir um som) pela
aplicação de um campo eléctrico.
Efeito piezoeléctrico inverso: aparecimento de cargas eléctricas na superfície de um material
quando este é sujeito a forças de pressão.
78
Relativamente à emissão dos ultra-sons, é possível dividir a zona em que se vai sentir a sua
acção em duas partes:
Zona de Fresnel: zona próxima da emissão onde a intensidade do som é superior à
esperada teoricamente;
Zona de Fraunhofer: zona mais afastada de intensidade igual é esperada e onde o som
sofre um certa dispersão. Depende sobretudo da frequência e da cabeça do emissor
do ultra-som.
Fenómenos físicos importantes quando se utilizam ultra-sons:
Interferência: em determinados locais, a intensidade do ultra-som pode ser superior
àquela que se está a introduzir e, consequentemente, pode provocar queimaduras
internas;
Cativação: quando um ultra-som se propaga num líquido que contém gases
dissolvidos, esses gases podem formar bolhas que podem implodir. Pode ser utilizado
em esterilização;
Pressão de radiação: a emissão do ultra-som dá origem a uma determinada pressão,
que depende da sua intensidade. Pode utilizar-se este fenómeno para medir a
intensidade da radiação ou para introduzir substâncias na pele sob a forma de creme
ou gel.
A ultrasonoterapia faz uso dos efeitos biofísicos dos ultra-sons: efeitos térmicos, mecânicos e
biológicos.
Ecografia: técnica de ultrassonografia. Baseia-se na interpretação do som reflectido (eco) nas
descontinuidades dos tecidos, quando estes são atingidos por uma onda sonora, para produzir
imagens.
Vantagens:
Técnica não invasiva;
Inócua;
Baixo custo.
Aplicações:
Visualização de órgãos e massas em várias zonas:
o Pescoço (tiroideia, paratiroideias);
o Abdómen (fígado, vesícula, pâncreas, baço);
o Aparelho urinário (rins, bexiga e próstata);
o Aparelho cardiovascular (coração, artérias, veias);
o Partes moles (músculos, tendões);
É muito utilizada em ginecologia e obstetrícia;
Só não pode ser aplicada nos pulmões e não traz vantagens para a observação dos
ossos.
Possibilita diferentes métodos de representação:
Modo A: modo de amplitude. Possibilita medições com precisão e representação
bidimensional;
79
Modo B: modo de brilho. Imagens topográficas em modo estático, representação
bidimensional;
Modo M: modo de movimento. Tecnologicamente mais complexo, permite registar os
movimentos de estruturas;
Dopller: permite medir débitos, como a velocidade de circulação do sangue numa
artéria.
80