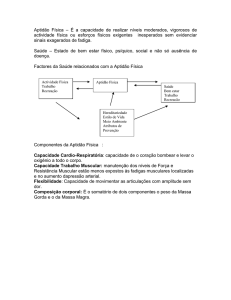
Revisão:
INSETOS EUSOCIAIS E O DESAFIO PARA A IDÉIA
DE SELEÇÃO NATURAL
Pedro Leite Ribeiro
Departamento de Fisiologia, Instituto de Biociências, USP
Recebido 26mar09 / Aceito 24nov09 / Publicação inicial 24nov09
Pedro Leite Ribeiro
Resumo. Este texto versa sobre a clássica discussão a respeito da dificuldade que a teoria da
Seleção Natural tem em explicar o surgimento de indivíduos não férteis. Para tanto é apresentado o
conceito de eusocialidade, as idéias de seleção de parentesco, altruísmo, altruísmo forçado e seleção de
grupo.
Palavras-chave. Seleção natural, Seleção de parentesco, Hamilton
EUSOCIAL INSECTS AND THE CHALLENGE TO THE IDEA OF NATURAL SELECTION
Abstract. This article is about the classic debate about the difficulty that the theory of natural selection
has in explaining the emergence of non-fertile individuals. Therefore, it is presented the concept of
eusociality, the ideas of kin selection, altruism, enforced altruism and group selection.
Keywords. Natural Selection, Kin selection, Hamilton
Darwin, ao propor a teoria da Seleção
Natural em seu livro Origem das Espécies,
questiona a abrangência de sua teoria quando,
por exemplo, menciona os insetos sociais: “há
que se admitir a existência de casos que
apresentam especial dificuldade com relação à
teoria da seleção natural. Um dos mais curiosos é
o da existência de duas ou três castas definidas
de formigas-operárias ou fêmeas estéreis na
mesma comunidade de insetos” (Darwin, 1859).
Wilson, em 1971, estipula três critérios, que
têm a pretensão de classificar todas as espécies
animais com relação aos níveis de socialidade, a
saber: 1) que indivíduos da mesma espécie
ajudam de forma cooperativa na criação dos
jovens; 2) que há uma divisão de tarefas
reprodutivas em uma sociedade em que um
grupo de indivíduos é infértil e colabora com a
criação dos filhos dos indivíduos férteis da
colônia, além de desempenharem outras funções
de manutenção da colônia; e 3) a existência de
uma sobreposição de pelo menos duas gerações,
de forma que os filhos possam ajudar seus pais.
Sob a perspectiva de Wilson, os animais ditos
eusociais são apenas aqueles que exibem os três
critérios por ele estipulados (Wilson, 1971).
Então, é importante notar que existem também
vários estágios considerados intermediários,
como, por exemplo, os seres humanos, que são
capazes de comportamentos solidários com a
prole alheia, sem, no entanto, abdicarem de sua
própria capacidade reprodutiva.
Portanto, a idéia de que a seleção natural
agiria exclusivamente sobre o indivíduo traz uma
dificuldade para a explicação de como teriam sido
selecionadas as relações altruísticas das castas
inférteis de uma colônia de formigas. Afinal,
nesse caso, como a esterilidade de um indivíduo
pode ter sido selecionada? Se um indivíduo é
estéril, jamais poderá deixar descendentes e não
terá qualquer aptidão, (capacidade de deixar
descendentes diretos) algo que, pensando sob o
olhar clássico da evolução, torna-se paradoxal. A
dificuldade
torna-se
ainda
maior
se
considerarmos que estas castas inférteis
apresentam marcadas diferenças morfológicas:
como
essas
diferenças
poderiam
ser
selecionadas num sistema no qual não há
reprodução (Darwin, 1859)?
Hamilton em 1964 publicou dois trabalhos
que procuravam explicar, sob a luz da seleção
natural, o comportamento eusocial nas diferentes
espécies animais (Hamilton, 1964a; Hamilton,
1964b) Sob essa perspectiva, ambas as
pesquisas introduziram uma nova e importante
idéia no contexto da evolução, a idéia da seleção
de parentesco. Para entendermos a concepção
desta idéia, temos, antes de tudo, que explanar
outro conceito fundamental, a saber, o conceito
de aptidão abrangente (fitness inclusivo, ou
fitness total), que é a base da teoria de Hamilton
ao definir aptidão (fitness) como a capacidade de
um indivíduo transmitir seus genes para as
gerações futuras.
De acordo com as idéias de Hamilton
aptidão abrangente é o resultado da soma da
aptidão direta com a aptidão indireta. A aptidão
direta é oriunda da capacidade reprodutiva do
indivíduo em questão, enquanto que a aptidão
indireta é dada pela capacidade reprodutiva dos
outros membros da comunidade em que o animal
vive, e que guardam algum tipo de parentesco
com o indivíduo de referência. Como o cálculo da
aptidão indireta leva em conta, necessariamente,
a consideração do grau de parentesco dos outros
membros da comunidade com o indivíduo em
questão, quanto maior o grau de parentesco com
os parentes reprodutivamente ativos, maior seria,
então, a aptidão indireta desse indivíduo. Assim,
a
capacidade
reprodutiva
de
membros
geneticamente relacionados (parentes) de um
determinado indivíduo tem uma importante
participação na aptidão abrangente (total) do
indivíduo. Dessa forma, temos a aptidão
abrangente de um indivíduo parcialmente
desvinculada da capacidade do indivíduo de gerar
6
REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009
filhos, pois os seus genes podem ser transmitidos
para as gerações futuras, por exemplo, pelos
seus irmãos, através dos sobrinhos, consistindo,
nisso, o principal argumento sobre o qual
Hamilton teceu a sua teoria. Portanto, do ponto
de vista de Hamilton, comportamentos altruísticos
poderiam ser justificados pelo aumento da
aptidão indireta, desde que este aumento seja
maior do que o prejuízo na aptidão direta que o
comportamento altruístico possa provocar no
indivíduo que faz a generosidade. Ou seja, tratase, nesse caso, de um aumento na aptidão total.
Assim, em situações particulares em que exista
um alto grau de parentesco entre os membros de
uma comunidade, pode ser mais vantajoso, do
ponto de vista da transmissão de genes para
gerações futuras, abdicar da aptidão direta em
prol da aptidão indireta. É em sociedades
partenogênicas que este raciocínio ganha força,
afinal, as irmãs compartilham, em média, 75%
dos genes umas com as outras, enquanto que
mães e filhas compartilham apenas 50% dos
genes, o que torna a aptidão indireta
potencializada. Quanto à transmissão gênica,
pode tornar-se mais interessante que se tenha
maior cuidado com as irmãs do que com os
próprios filhos. Estas idéias parecem, portanto,
acalmar as críticas com relação à espetacular
contradição de que a seleção natural teria levado
ao surgimento de indivíduos que não têm
qualquer capacidade de transmitir os seus genes
diretamente, algo que, à primeira vista, pode
parecer improvável de ser selecionado. Afinal, a
teoria da seleção natural versa: os indivíduos que
são selecionados são aqueles que têm maior
capacidade de deixar descendentes.
Desse modo, uma vez que temos uma
explicação lógica e coerente sobre o status quo
dos sistemas biológicos que nos propusemos a
estudar, por que não tentarmos explicar o
surgimento dos sistemas biológicos eusociais?
Podemos, com isso, nos perguntar: quais eram –
ou deveriam ser – as condições ecológicas dos
ancestrais desses animais eusociais que hoje
conhecemos? De fato há artigos de autores
consagrados que lidam com essas questões, por
exemplo, Arillo (2007) e Wilson e Holldobler
(2005).
Dessa
forma,
a
descoberta
de
características comuns aos ancestrais destes
grupos pode ser de grande valor para o
entendimento de quais são as condições
necessárias ou pelo menos favoráveis ao
surgimento de animais eusociais. Infelizmente, o
estudo de fósseis é pouco revelador com relação
ao comportamento e às condições ecológicas de
um determinado momento da história evolutiva.
Como há poucas informações extraídas das
estruturas morfológicas que podem evidenciar
algum tipo de comportamento, torna-se
necessário recorrer aos estudos sobre a
fossilização de grupos de insetos, que, ocorrida
em único evento, pôde eternizar um momento da
vida social de uma determinada espécie. As
fossilizações de formigas aladas e de outras
ápteras podem revelar a existência de indivíduos
tanto férteis como estéreis, o que seria um
indicativo de eusocialidade. Infelizmente, as
condições ecológicas em que estes animais
vivem são de difícil fossilização, e os estudiosos
do assunto devem construir inferências teóricas
para demonstrar como deveriam ser as condições
ecológicas dos ancestrais dos animais eusociais
que conhecemos hoje. Tais inferências, por sua
vez, podem estar apoiadas nas poucas
evidências
fósseis,
como
também
nas
características comportamentais e ecológicas dos
animais que hoje se encontram em estágios
intermediários de socialidade, ou, ainda, no
próprio estudo dos animais eusociais da
atualidade.
De maneira geral, acredita-se que
algumas condições devam ser satisfeitas para
que exista a possibilidade do surgimento da
eusocialidade. Em primeiro lugar, é preciso que
exista uma situação ecológica tal que grupos de
indivíduos da mesma espécie sejam obrigados a
viver juntos. Tanto por viverem em ninhos, ou
pela necessidade de manutenção de um território
que, dificilmente, possa ser mantido por um único
indivíduo. Em segundo lugar, devido às
necessidades alimentares restritas, a procura por
alimento torne-se demasiado intensa, que acabe
por inviabilizar o cumprimento das outras tarefas
que este indivíduo precisa executar, como, por
exemplo, o seu cuidado com a prole (Wilson,
2008). Portanto, dados todos esses fatores,
parece existir uma situação em que as condições
ecológicas adversas obriguem o trabalho coletivo
por meio da imposição de dificuldades à vida
solitária.
A idéia de seleção de parentesco pode
parecer, às vezes, insuficiente, por exemplo,
quando se leva em consideração o surgimento da
eusocialidade em cupins (diplobiontes), como
também em formigas, cujas fêmeas foram
fecundadas por vários machos. Afinal, nesses
casos a relação de parentesco entre os irmãos
deixa de ser de 75% e pode passar para menos
de 30%. Nesse contexto a aptidão indireta nunca
será maior que a aptidão direta – pois cuidar da
própria prole seria sempre mais vantajoso do que
abdicar dos próprios filhos em favor dos irmãos.
Surge, então, uma idéia alternativa ou, pelo
menos, complementar às idéias de Hamilton
(seleção de parentesco). Esta idéia – inicialmente
apresentada por Richard Alexander (1974) e que
ganha apoio mais de 20 anos depois (Foster e
col., 2002; Foster e Ratnieks, 2000; Foster e
Ratnieks, 2001; Ratnieks e Reeve, 1992;
Ratnieks e Visscher, 1989; Ratnieks e
Wenseleers, 2005; Ratnieks e Wenseleers, 2008)
– introduz um novo conceito dentro do contexto
7
REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009
da evolução da eusocialidade: o altruísmo
forçado.
Idéias e concepções como essas ganham
suporte em recentes estudos que descrevem uma
série de comportamentos agressivos dentro da
sociedade de insetos (Boomsma e Ratnieks,
1996; Chaline e col., 2004; Foster e Ratnieks,
2000; Foster e col., 2001; Foster e col., 2006;
Ratnieks, 1988; Ratnieks, 1991; Wenseleers e
col., 2005; Wenseleers e Ratnieks, 2006;
Wenseleers e col., 2003) que podem ser
resumidos em três tipos: 1) Coerção –
comportamento social agressivo, que pune e
policia o comportamento individual egoísta. 2)
Manipulação parental – comportamento exibido
pelos pais, que visa à persuasão dos filhos para
que cuidem dos irmãos. 3) Policiamento –
comportamento de inibição da atividade de
reprodução de determinada operários (formas
não reprodutivas presentes em colônias de
insetos socias) que pode acontecer pela
destruição dos ovos postos ou agressão física a
ela (Ratnieks e Wenseleers, 2008). Foi a
descrição de tais comportamentos que levou os
estudiosos do assunto a acreditarem que os
comportamentos tidos como altruístas poderiam
não ser voluntários como Hamilton supunha. Há,
a partir desse posicionamento, uma sutil e
importante diferença na compreensão de como se
deu o surgimento do comportamento eusocial, já
que a questão permissiva e causal do surgimento
de castas inférteis não mais estaria pautada
somente nas relações genéticas entre os
indivíduos que vivem conjuntamente; mas,
preponderantemente, esse surgimento consistiria
no comportamento agressivo de membros
dominantes dentro do grupo, o que levaria à
esterilidade de alguns indivíduos que a ele
pertencem. É claro que essas descobertas não
necessariamente são contrárias às idéias de
seleção de parentesco, e podem inclusive servir
de substrato para uma nova classe de
argumentação que suporte as idéias de Hamilton.
É evidente que as idéias de Hamilton não
podem ser desconsideradas ou totalmente
substituídas, afinal, a razão pela qual se deu a
origem do comportamento eusocial é um assunto
que ainda não está totalmente elucidado.
Poderíamos argumentar do ponto de vista
evolutivo que, por exemplo, a manutenção das
relações altruísticas pacíficas em animais
eusociais pode ter como explicação parcial – ou
até mesmo total – as idéias de seleção de
parentesco. De qualquer modo, uma idéia que
permanece
válida,
com
relação
ao
comportamento social das diferentes espécies
animais, é a de que, dentro do grupo, o
comportamento altruísta parece ser prejudicial
para o indivíduo que pratica a generosidade,
enquanto que, quando há uma comparação entre
grupos, parece-nos que o grupo que exibe
comportamento altruísta entre os seus membros
angaria alguma vantagem (Wilson e Wilson,
2007).
Bibliografia:
Alexander, R. (1974). The evolution of social
behaviour. Annual Review of Ecology and Systematics.
Arillo,
A.
(2007).
Paleoethology:
fossilized
behaviours in amber. Geologica Acta 5, 159-166.
Boomsma, J. J. e Ratnieks, F. L. W. (1996).
Paternity in eusocial Hymenoptera. Philosophical Transactions
Of The Royal Society Of London Series B-Biological Sciences
351, 947-975.
Chaline, N., Martin, S. J. e Ratnieks, F. L. W. (2004).
Worker policing persists in a hopelessly queenless honey bee
colony (Apis mellifera). Insectes Sociaux 51, 113-116.
Darwin, C. (1859). The origin of species (By Means
of Natural Selection). London: John Murray.
Foster, K. R., Gulliver, J. e Ratnieks, F. L. W. (2002).
Worker policing in the European hornet Vespa crabro. Insectes
Sociaux 49, 41-44.
Foster, K. R. e Ratnieks, F. L. W. (2000). Social
insects - Facultative worker policing in a wasp. Nature 407,
692-693.
Foster, K. R. e Ratnieks, F. L. W. (2001).
Convergent evolution of worker policing by egg eating in the
honeybee and common wasp. Proceedings Of The Royal
Society Of London Series B-Biological Sciences 268, 169-174.
Foster, K. R., Wenseleers, T. e Ratnieks, F. L. W.
(2001). Spite: Hamilton's unproven theory. Annales Zoologici
Fennici 38, 229-238.
Foster, K. R., Wenseleers, T. e Ratnieks, F. L. W.
(2006). Kin selection is the key to altruism. Trends In Ecology
& Evolution 21, 57-60.
Hamilton, W. D. (1964a). Genetical Evolution Of
Social Behaviour 2. Journal of Theoretical Biology 7, 17-&.
Hamilton, W. D. (1964b). Genetical Evolution Of
Social Behaviour I. Journal of Theoretical Biology 7, 1-&.
Ratnieks, F. L. W. (1988). Reproductive Harmony
Via Mutual Policing By Workers In Eusocial Hymenoptera.
American Naturalist 132, 217-236.
Ratnieks, F. L. W. (1991). Evolution Of
Discriminatory Aggression In Marine-Invertebrates. Journal of
Theoretical Biology 152, 557-565.
Ratnieks, F. L. W. e Reeve, H. K. (1992). Conflict In
Single-Queen Hymenopteran Societies - The Structure Of
Conflict And Processes That Reduce Conflict In Advanced
Eusocial Species. Journal of Theoretical Biology 158, 33-65.
Ratnieks, F. L. W. e Visscher, P. K. (1989). Worker
Policing In The Honeybee. Nature 342, 796-797.
Ratnieks, F. L. W. e Wenseleers, T. (2005). Policing
insect societies. Science 307, 54-56.
Ratnieks, F. L. W. e Wenseleers, T. (2008). Altruism
in insect societies and beyond: voluntary or enforced? Trends
In Ecology & Evolution 23, 45-52.
Wenseleers, T., Badcock, N. S., Erven, K., e col.
(2005). A test of worker policing theory in an advanced
eusocial wasp, Vespula rufa. Evolution 59, 1306-1314.
Wenseleers, T. e Ratnieks, F. L. W. (2006).
Enforced altruism in insect societies. Nature 444, 50-50.
Wenseleers, T., Ratnieks, F. L. W. e Billen, J.
(2003). Caste fate conflict in swarm-founding social
Hymenoptera: an inclusive fitness analysis. Journal Of
Evolutionary Biology 16, 647-658.
Wilson, D. S. e Wilson, E. O. (2007). Rethinking the
theoretical foundation of sociobiology. Quarterly Review of
Biology 82, 327-348.
Wilson, E. O. (1971). The insect societies.
Cambridge, MA: Belknap Press of Harvard University Press.
Wilson, E. O. (2008). One giant leap: How insects
achieved altruism and colonial life. Bioscience 58, 17-25.
Wilson, E. O. e Holldobler, B. (2005). Eusociality:
Origin and consequences (vol 102, pg 13367, 2005).
Proceedings of the National Academy of Sciences of the
United States of America 102, 16119-16119.
8
REVISTA DA BIOLOGIA – www.ib.usp.br/revista – publicado no volume 3 – dezembro de 2009